
Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway

 ,
,  and
and 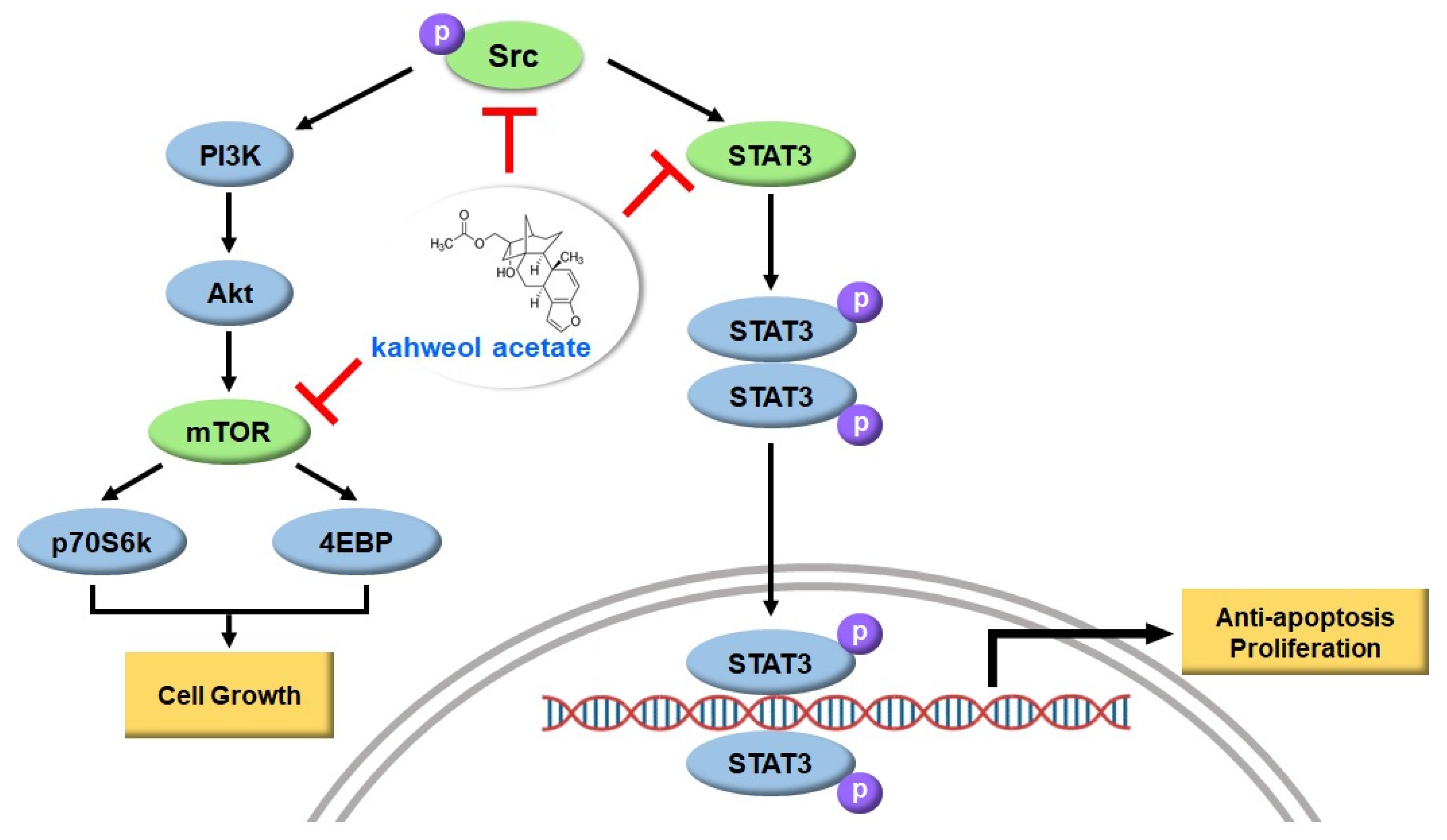{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
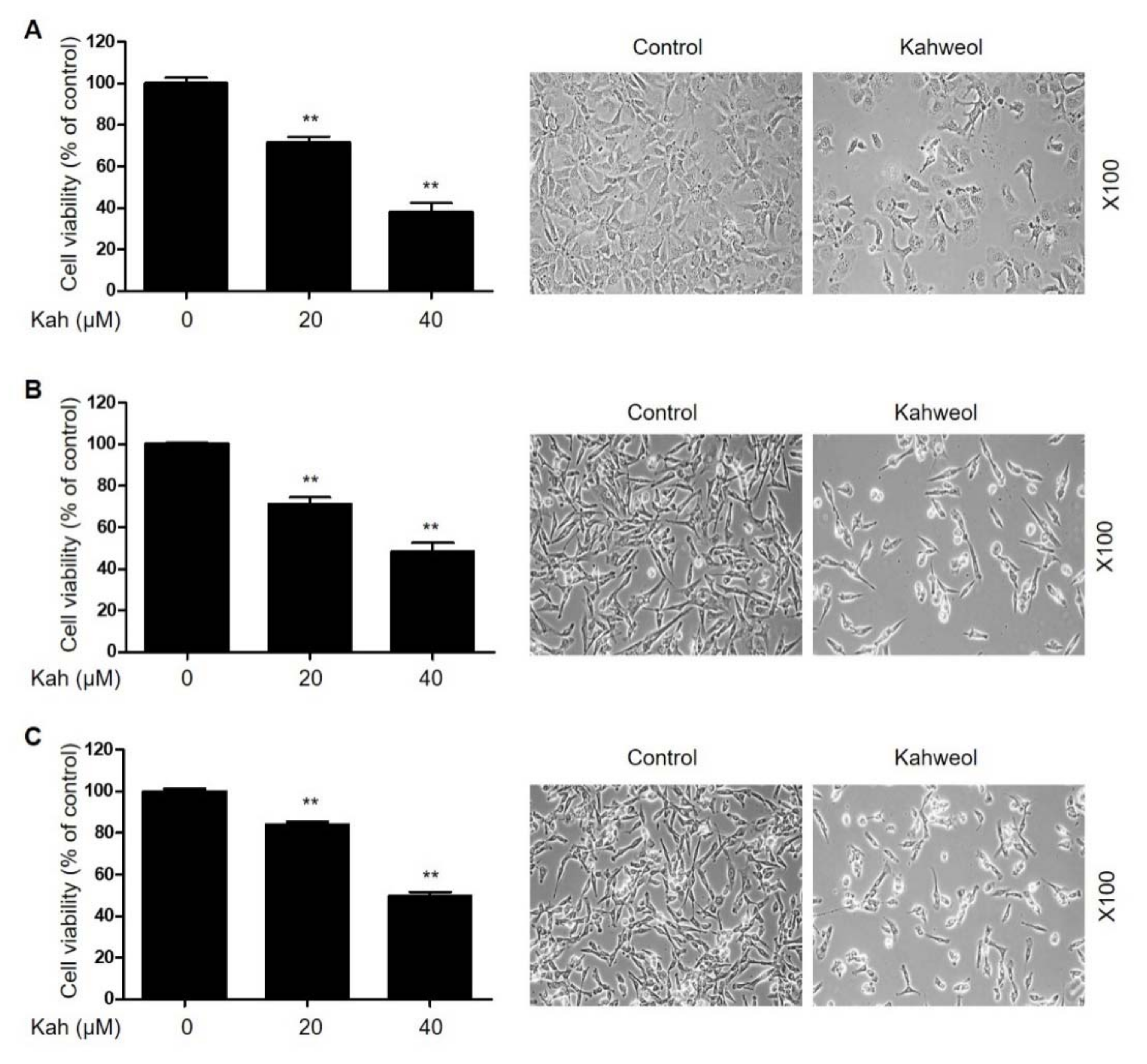
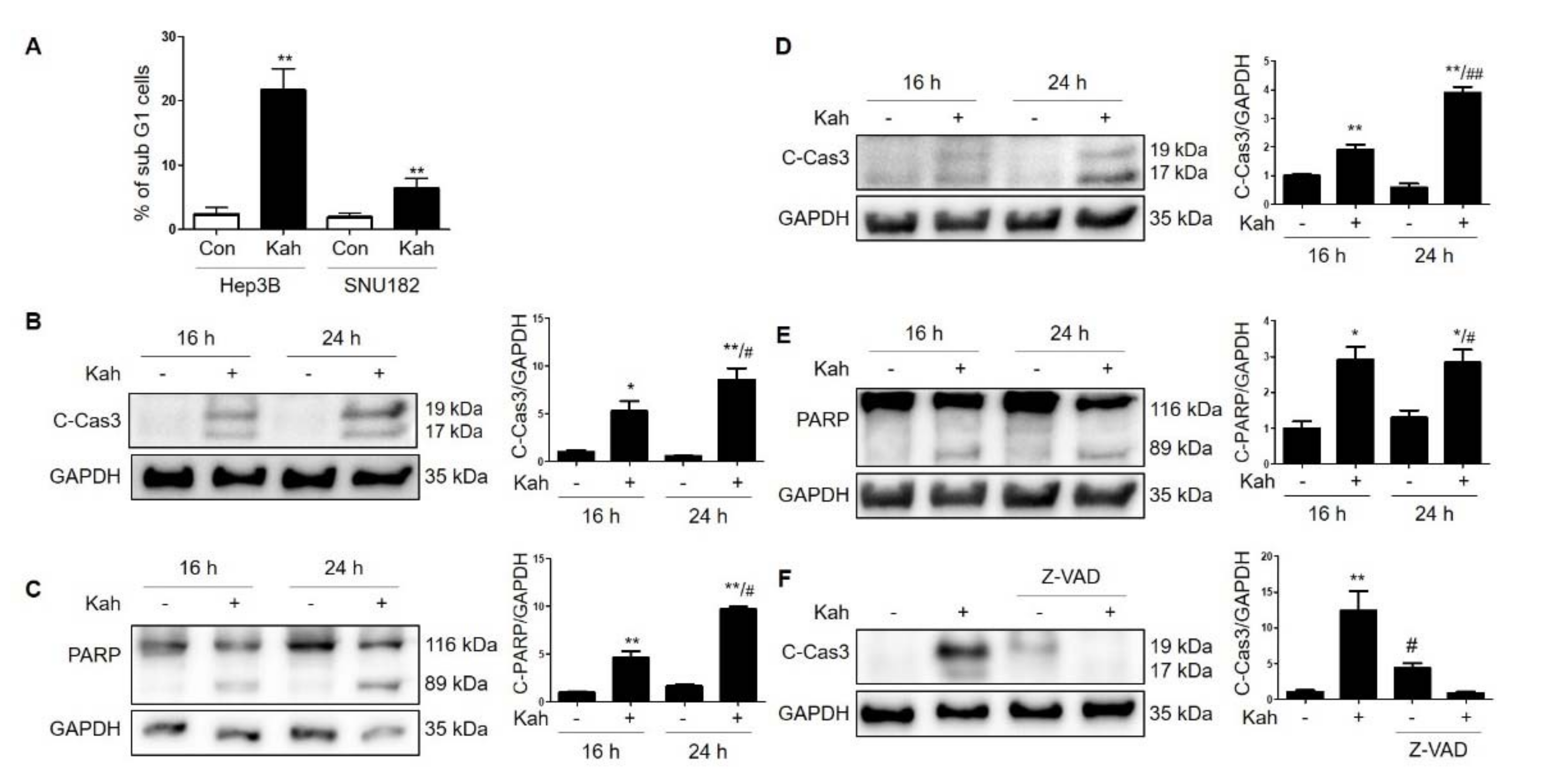
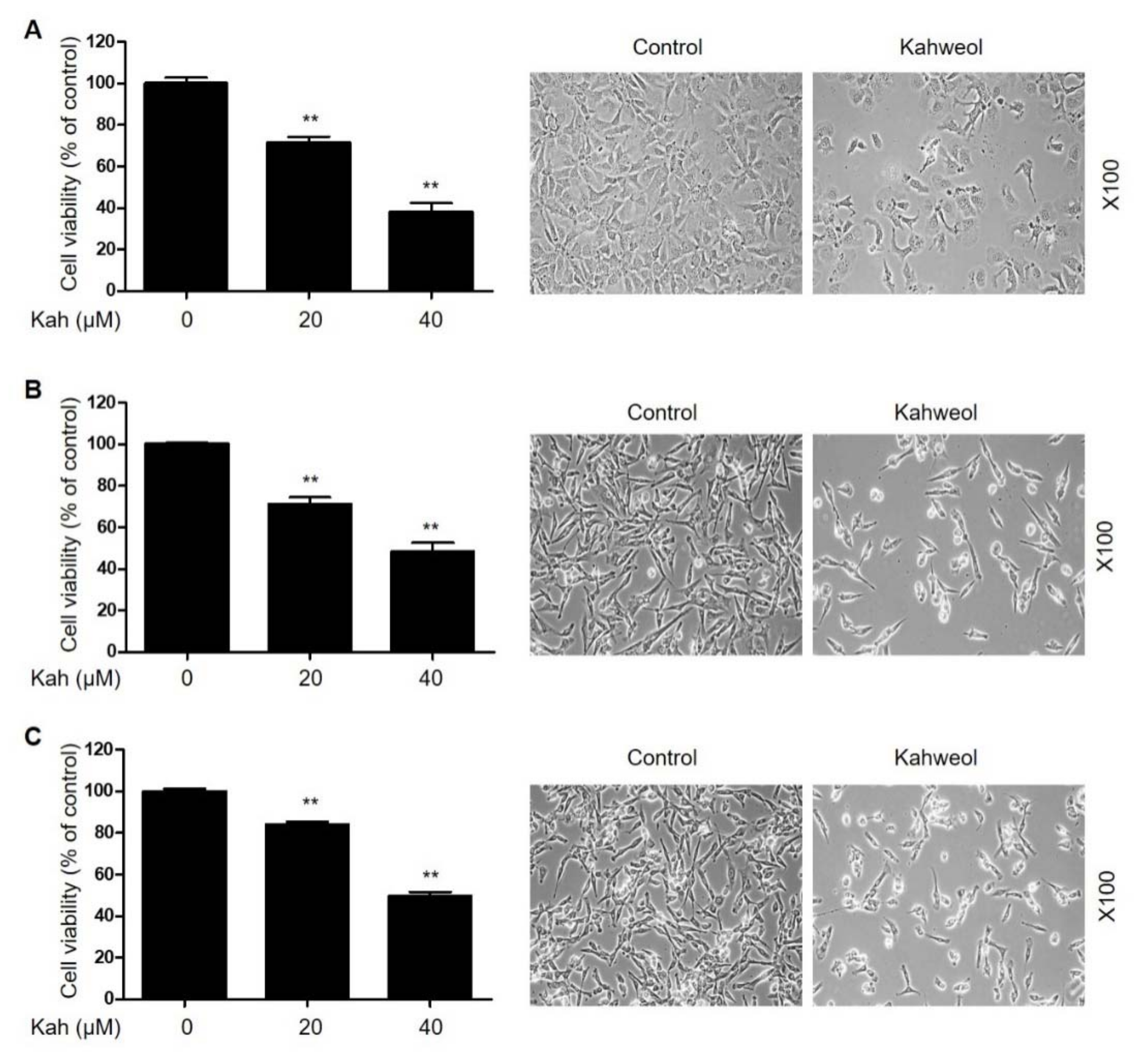
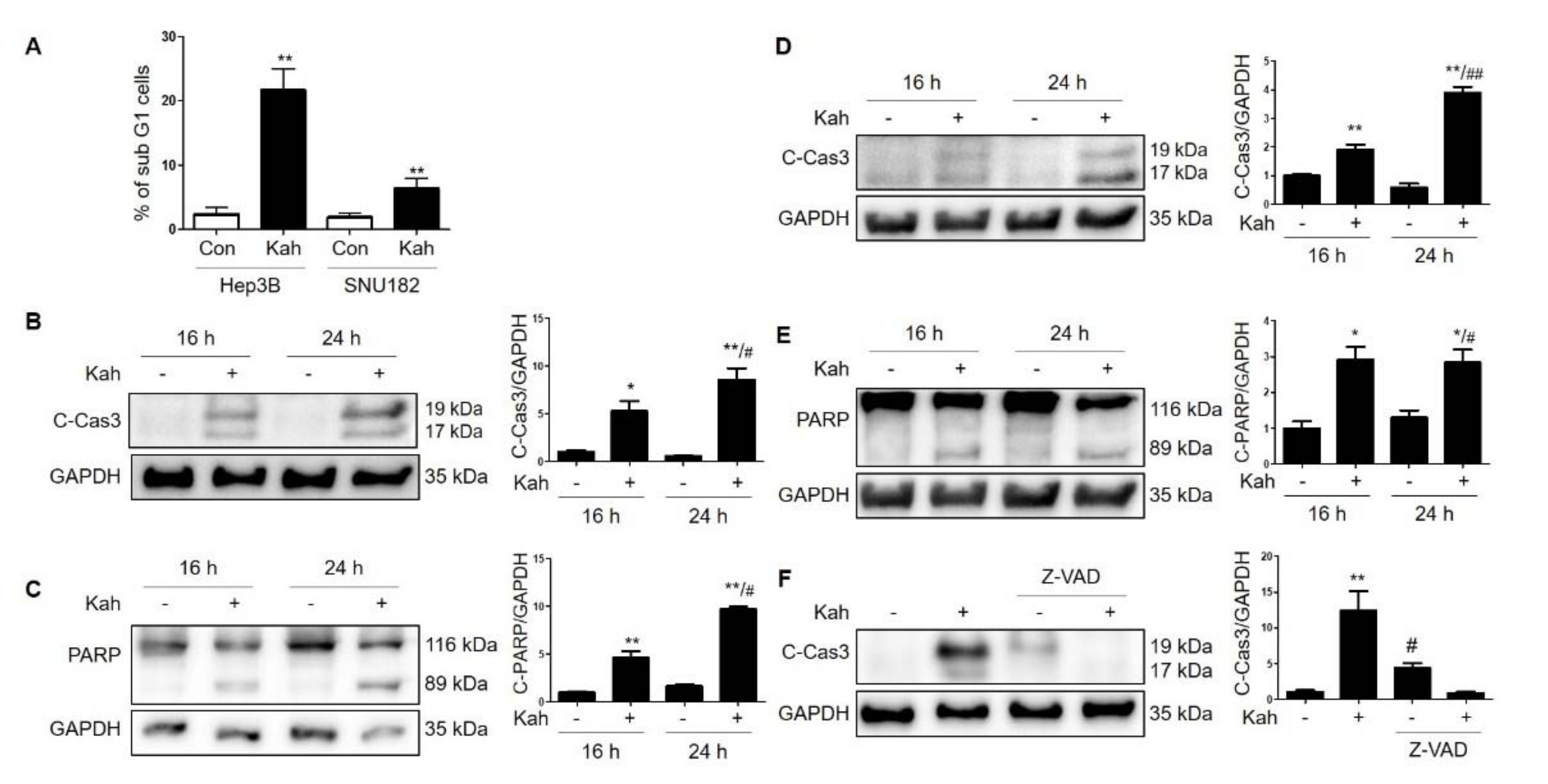
2.1. KahweolInhibits Cell Proliferation and Induces Apoptosis in HCC Cells
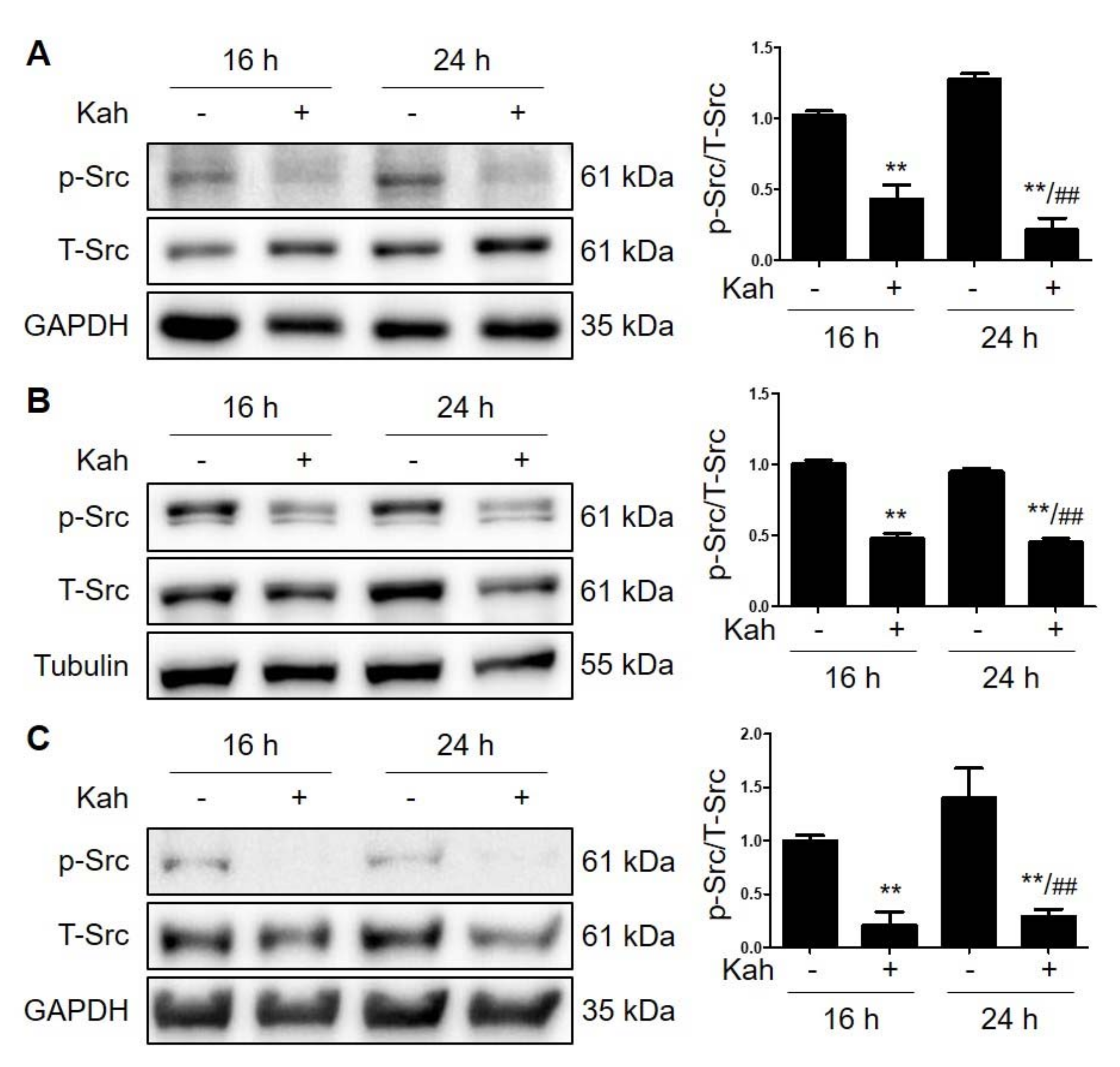
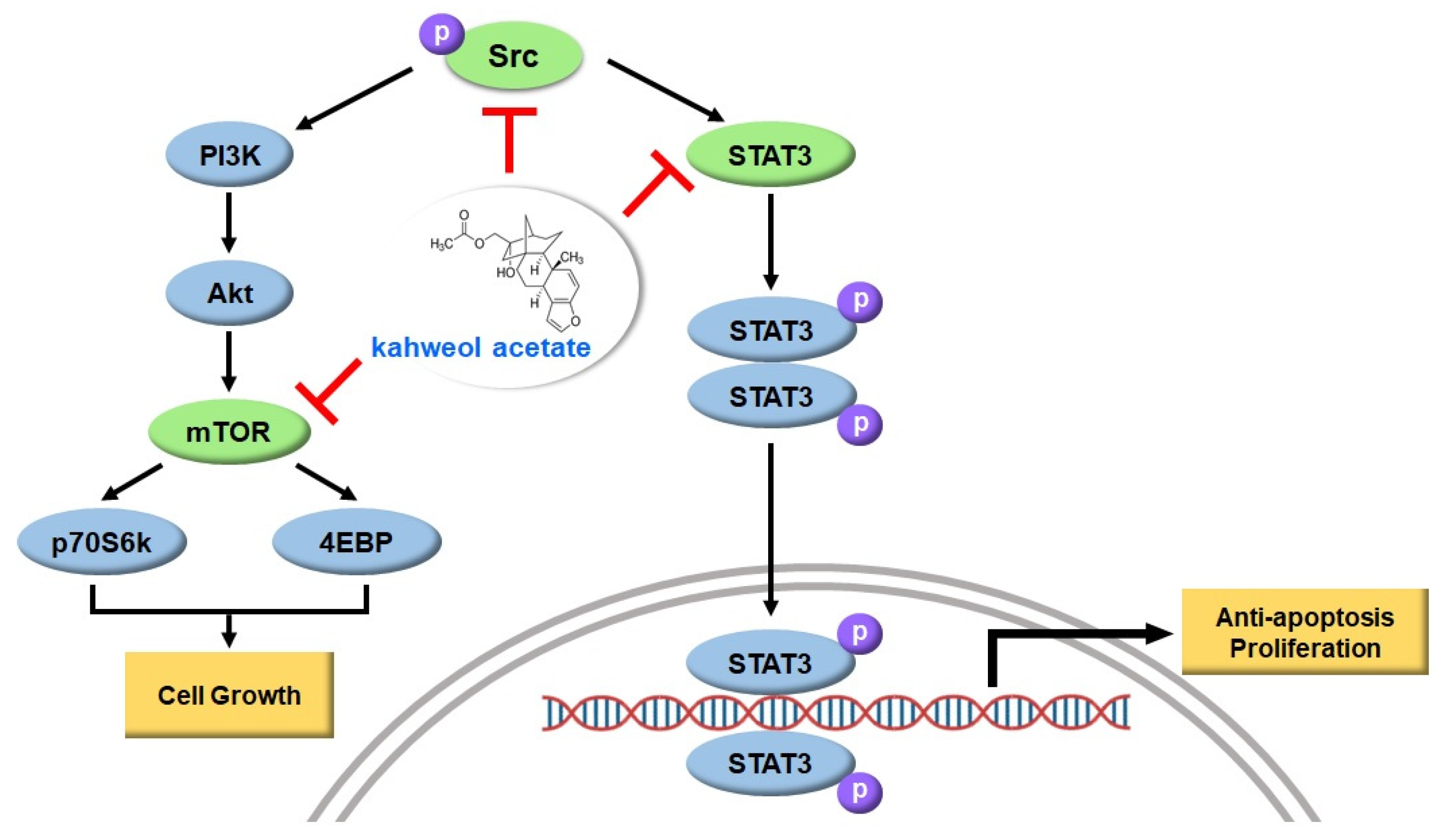
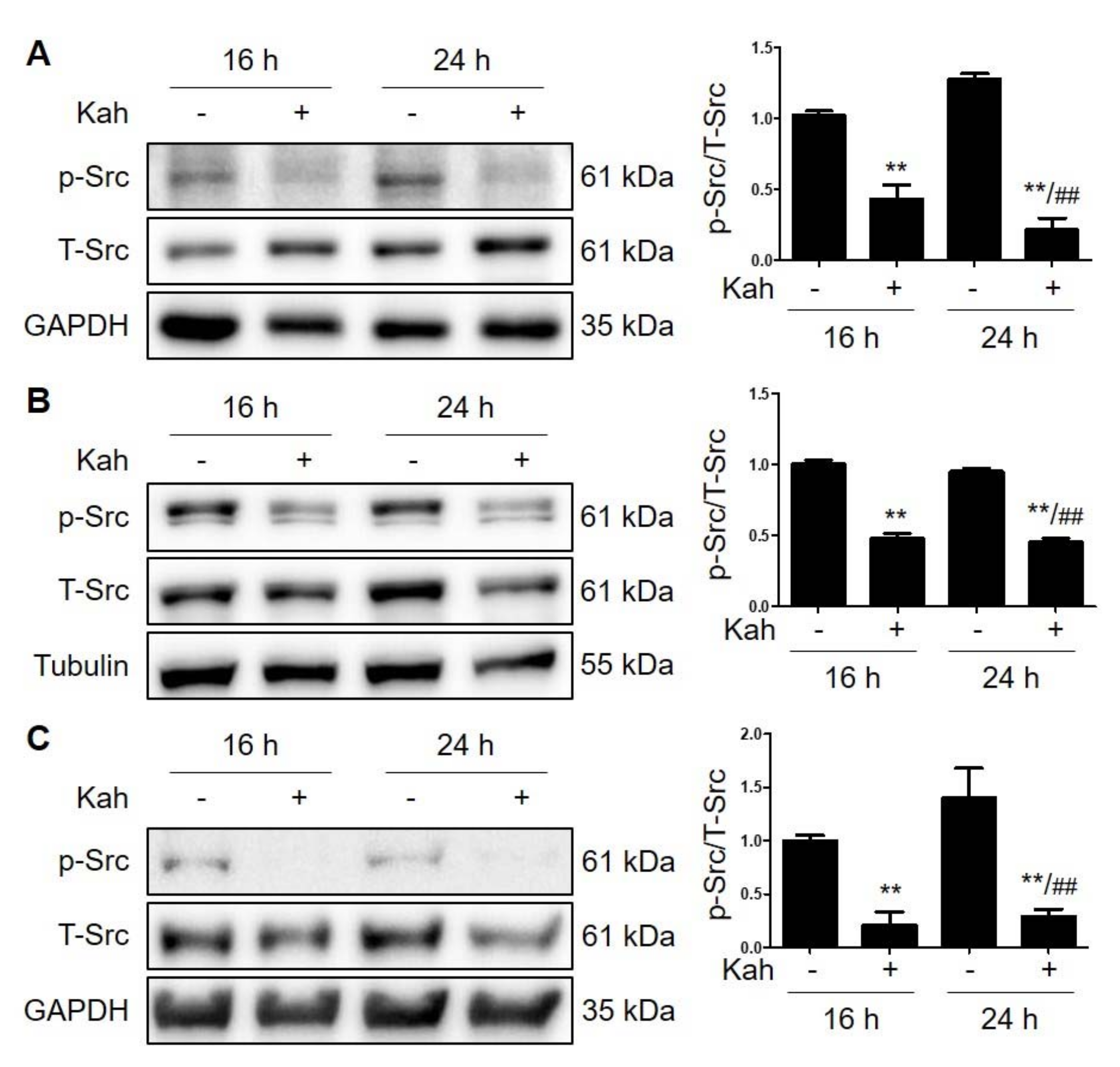
2.2. Kahweol Attenuates Src Phosphorylation in HCC Cells
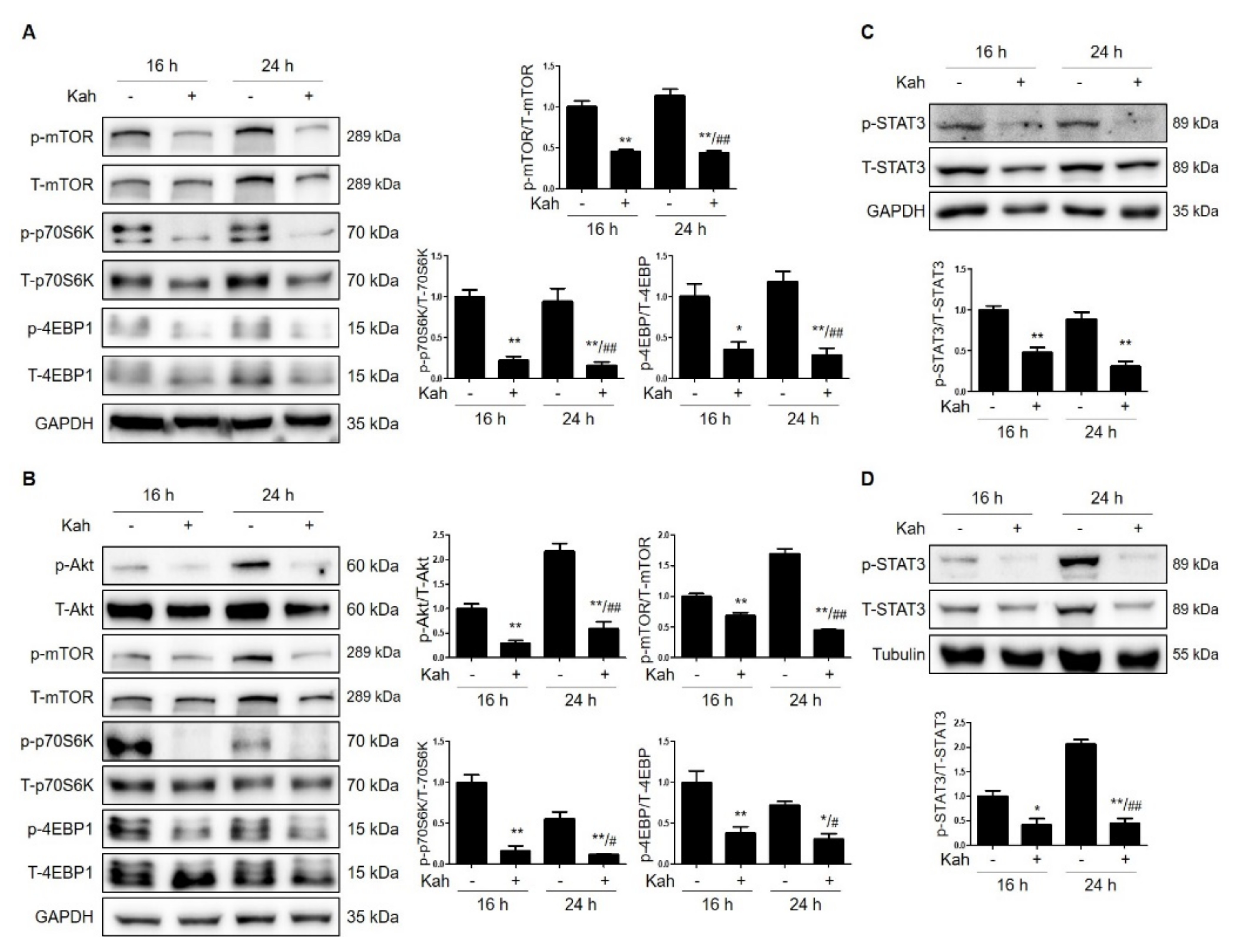
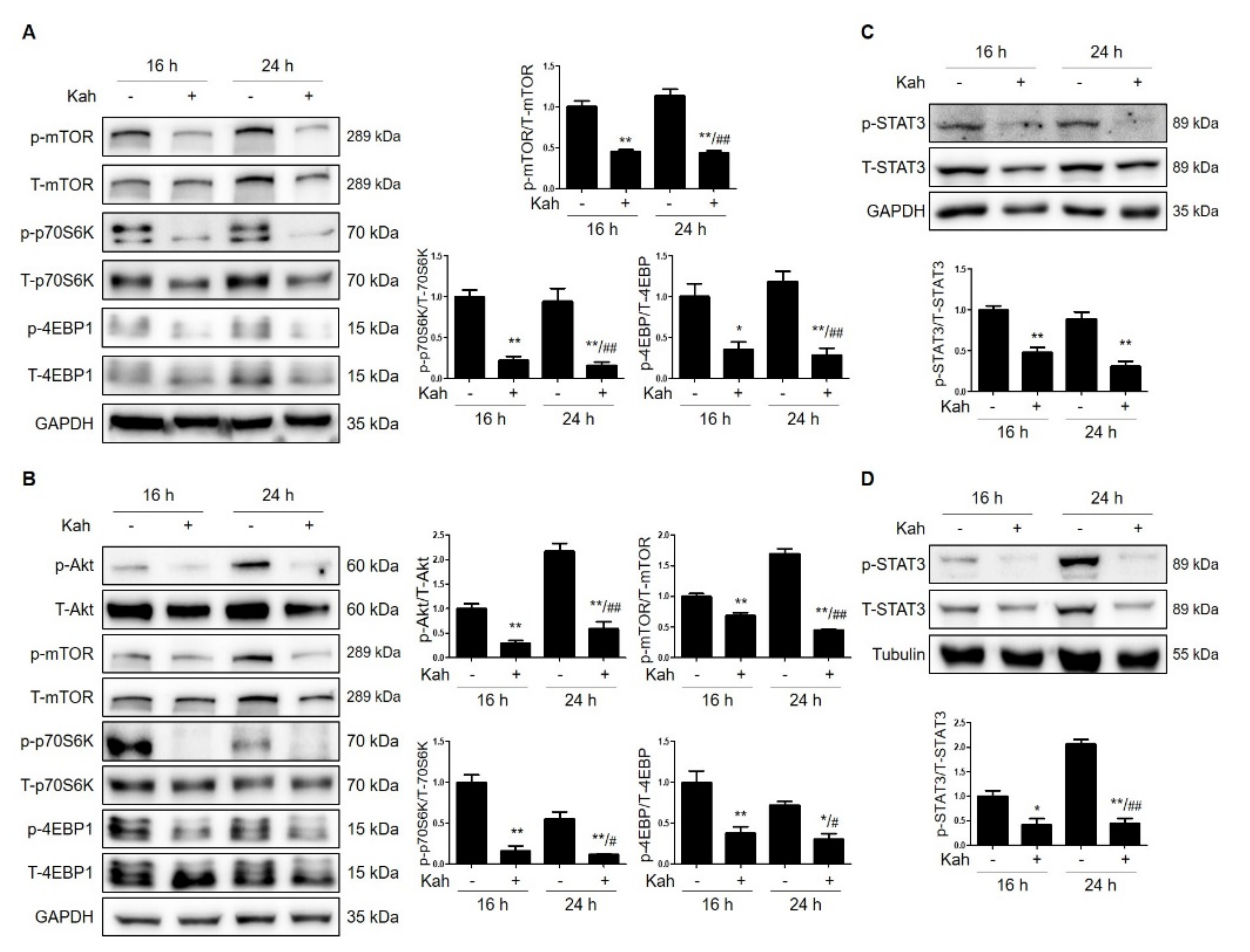
2.3. Kahweol Attenuates mTOR Activation in HCC Cells
2.4. Kahweol Attenuates STAT3 Phosphorylation in HCC Cells
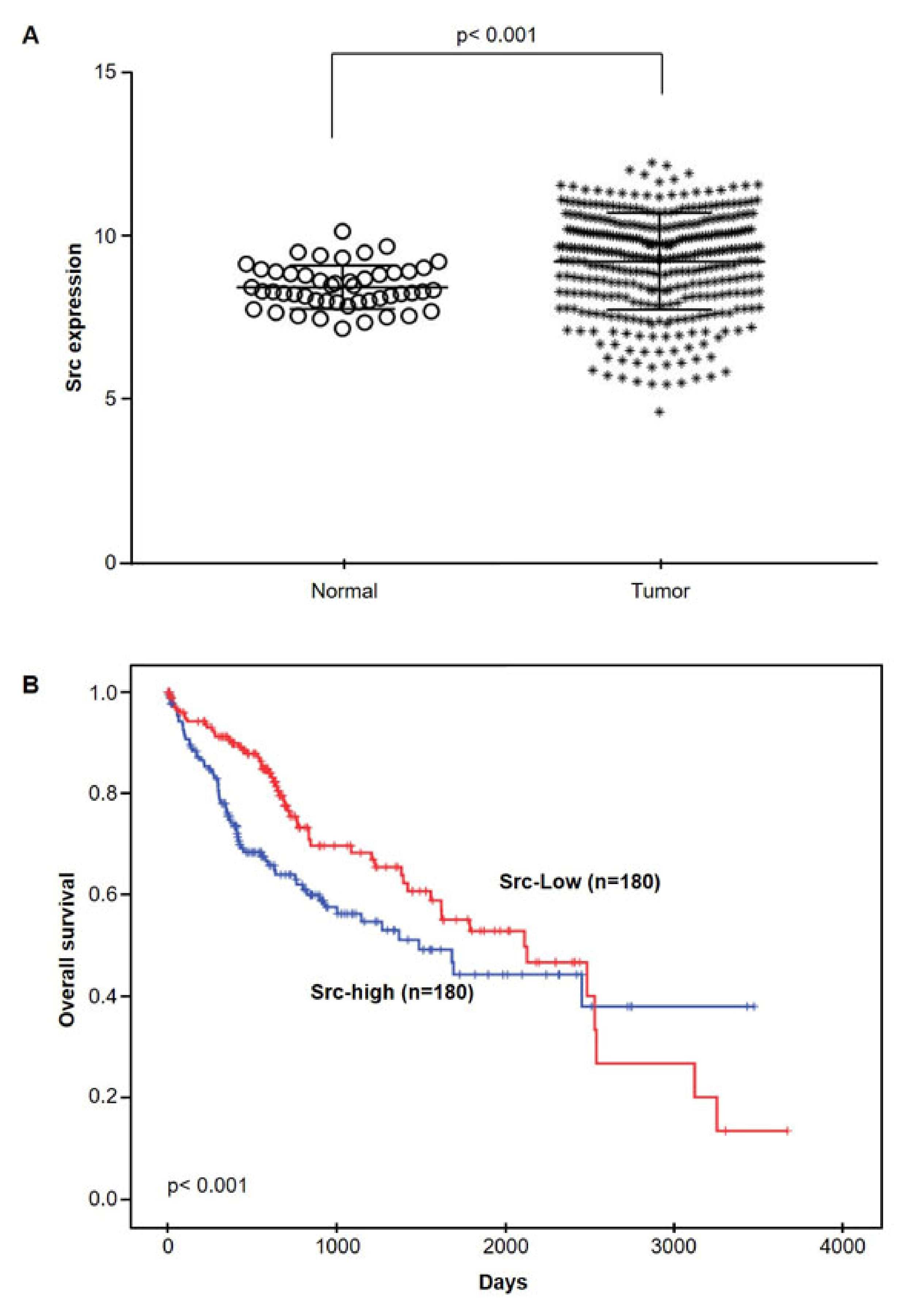
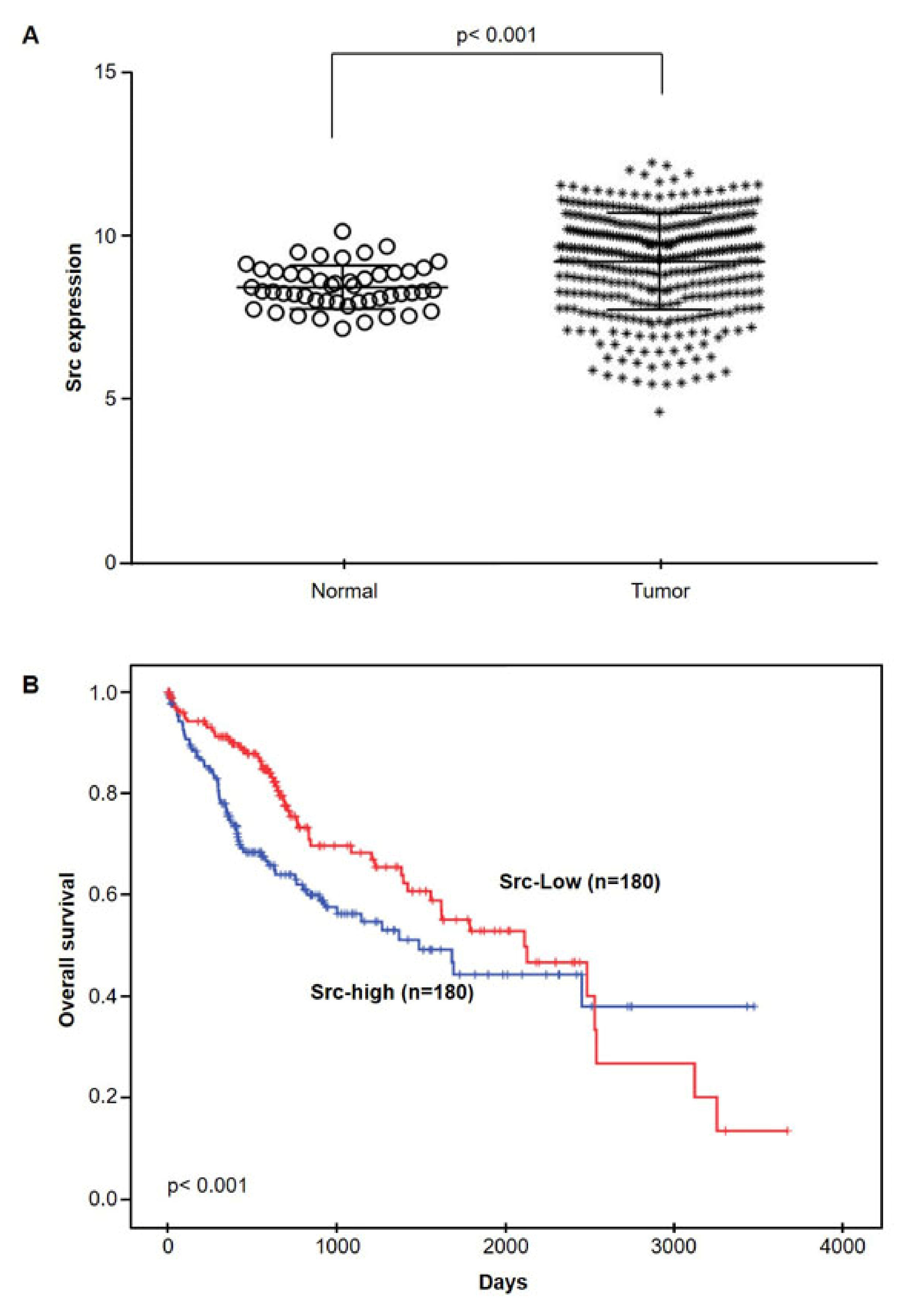
2.5. Src mRNA Expressions in HCC by TCGA database
3. Discussion
4. Materials and Methods
4.1. The Cancer Genome Atlas (TCGA) Data Analysis
4.2. Chemicals
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Flow Cytometry Analysis
4.6. Western Blot Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Fujiwara, N.; Friedman, S.L.; Goossens, N.; Hoshida, Y. Risk factors and prevention of hepatocellular carcinoma in the era of precision medicine. J. Hepatol. 2018, 68, 526–549. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.; Stepanova, M.; Ong, J.P.; Jacobson, I.M.; Bugianesi, E.; Duseja, A.; Eguchi, Y.; Wong, V.W.; Negro, F.; Yilmaz, Y.; et al. Nonalcoholic Steatohepatitis Is the Fastest Growing Cause of Hepatocellular Carcinoma in Liver Transplant Candidates. Clin. Gastroenterol. Hepatol. 2019, 17, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Myers, S.; Neyroud-Caspar, I.; Spahr, L.; Gkouvatsos, K.; Fournier, E.; Giostra, E.; Magini, G.; Frossard, J.-L.; Bascaron, M.-E.; Vernaz, N.; et al. NAFLD and MAFLD as emerging causes of HCC: A populational study. JHEP Rep. 2021, 3, 100231. [Google Scholar] [CrossRef]
- Yang, W.-S.; Zeng, X.-F.; Liu, Z.-N.; Zhao, Q.-H.; Tan, Y.-T.; Gao, J.; Li, H.-L.; Xiang, Y.-B. Diet and liver cancer risk: A narrative review of epidemiological evidence. Br. J. Nutr. 2020, 124, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Butt, M.S.; Sultan, M.T. Coffee and its consumption: Benefits and risks. Crit. Rev. Food Sci. Nutr. 2011, 51, 363–373. [Google Scholar] [CrossRef]
- Coffee consumption and health: Umbrella review of meta-analyses of multiple health outcomes. BMJ 2018, 360, k194. [CrossRef] [Green Version]
- Bravi, F.; Tavani, A.; Bosetti, C.; Boffetta, P.; La Vecchia, C. Coffee and the risk of hepatocellular carcinoma and chronic liver disease: A systematic review and meta-analysis of prospective studies. Eur. J. Cancer Prev. 2017, 26, 368–377. [Google Scholar] [CrossRef]
- Gross, G.; Jaccaud, E.; Huggett, A.C. Analysis of the content of the diterpenes cafestol and kahweol in coffee brews. Food Chem. Toxicol. 1997, 35, 547–554. [Google Scholar] [CrossRef]
- Cárdenas, C.; Quesada, A.R.; Medina, M.A. Correction: Anti-Angiogenic and Anti-Inflammatory Properties of Kahweol, a Coffee Diterpene. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Seo, H.Y.; Kim, M.K.; Lee, S.H.; Hwang, J.S.; Park, K.G.; Jang, B.K. Kahweol Ameliorates the Liver Inflammation through the Inhibition of NF-κB and STAT3 Activation in Primary Kupffer Cells and Primary Hepatocytes. Nutrients 2018, 10, 863. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Q.; Bi, L.; Ren, Y.; Song, S.; Wang, Q.; Wang, Y.S. Advances in studies of tyrosine kinase inhibitors and their acquired resistance. Mol. Cancer 2018, 17, 36. [Google Scholar] [CrossRef]
- Brown, M.T.; Cooper, J.A. Regulation, substrates and functions of src. Biochim. Biophys. Acta 1996, 1287, 121–149. [Google Scholar] [CrossRef]
- Mathew, S.; George, S.P.; Wang, Y.; Siddiqui, M.R.; Srinivasan, K.; Tan, L.; Khurana, S. Potential molecular mechanism for c-Src kinase-mediated regulation of intestinal cell migration. J. Biol. Chem. 2008, 283, 22709–22722. [Google Scholar] [CrossRef] [Green Version]
- Lau, G.M.; Lau, G.M.; Yu, G.-L.; Gelman, I.H.; Gutowski, A.; Hangauer, D.; Fang, J.W.S. Expression of Src and FAK in Hepatocellular Carcinoma and the Effect of Src Inhibitors on Hepatocellular Carcinoma In Vitro. Dig. Dis. Sci. 2009, 54, 1465–1474. [Google Scholar] [CrossRef]
- Makino, T.; Izumi, K.; Hiratsuka, K.; Kano, H.; Shimada, T.; Nakano, T.; Kadomoto, S.; Naito, R.; Iwamoto, H.; Yaegashi, H.; et al. Anti-proliferative and anti-migratory properties of coffee diterpenes kahweol acetate and cafestol in human renal cancer cells. Sci. Rep. 2021, 11, 675. [Google Scholar] [CrossRef]
- Chen, Q.; Zhou, Z.; Shan, L.; Zeng, H.; Hua, Y.; Cai, Z. The importance of Src signaling in sarcoma. Oncol. Lett. 2015, 10, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.; Wu, J.S.; Mao, S.S.; Yu, X.N.; Huang, X.X. Effect of saracatinib on pulmonary metastases from hepatocellular carcinoma. Oncol. Rep. 2016, 36, 1483–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, X.; Liu, L.; Yang, X.; Qian, Q.; Du, B. c-Src promotes the growth and tumorigenesis of hepatocellular carcinoma via the Hippo signaling pathway. Life Sci. 2021, 264, 118711. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Hwang, Y.P.; Jeong, H.G. Kahweol blocks STAT3 phosphorylation and induces apoptosis in human lung adenocarcinoma A549 cells. Toxicol. Lett. 2009, 187, 28–34. [Google Scholar] [CrossRef]
- Cárdenas, C.; Quesada, A.R.; Medina, M.Á. Insights on the antitumor effects of kahweol on human breast cancer: Decreased survival and increased production of reactive oxygen species and cytotoxicity. Biochem. Biophys. Res. Commun. 2014, 447, 452–458. [Google Scholar] [CrossRef]
- Choi, D.W.; Lim, M.S.; Lee, J.W.; Chun, W.; Lee, S.H.; Nam, Y.H.; Park, J.; Choi, D.H.; Kang, C.D.; Lee, S.J.; et al. The Cytotoxicity of Kahweol in HT-29 Human Colorectal Cancer Cells Is Mediated by Apoptosis and Suppression of Heat Shock Protein 70 Expression. Biomol. Ther. 2015, 23, 128–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Um, H.J.; Park, J.W.; Kwon, T.K. Melatonin sensitizes Caki renal cancer cells to kahweol-induced apoptosis through CHOP-mediated up-regulation of PUMA. J. Pineal Res. 2011, 50, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.I.; Jeon, Y.J.; Shim, J.H. Anti-proliferative properties of kahweol in oral squamous cancer through the regulation specificity protein 1. Phytotherapy Res. 2014, 28, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Lee, J.T.; Yang, E.S.; Chang, J.-S.; Lee, D.S.; Kim, S.H.; Choi, Y.H.; Park, J.-W.; Kwon, T.K. The coffee diterpene kahweol induces apoptosis in human leukemia U937 cells through down-regulation of Akt phosphorylation and activation of JNK. Apoptosis Int. J. Program. Cell Death. 2009, 14, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Park, G.H.; Song, H.M.; Jeong, J.B. The coffee diterpene kahweol suppresses the cell proliferation by inducing cyclin D1 proteasomal degradation via ERK1/2, JNK and GKS3β-dependent threonine-286 phosphorylation in human colorectal cancer cells. Food Chem. Toxicol. 2016, 95, 142–148. [Google Scholar] [CrossRef]
- Kim, L.C.; Song, L.; Haura, E.B. Src kinases as therapeutic targets for cancer. Nat. Rev. Clin. Oncol. 2009, 6, 587–595. [Google Scholar] [CrossRef]
- Chen, M.-L.; Chai, C.-Y.; Yeh, K.-T.; Wang, S.-N.; Tsai, C.-J.; Yeh, Y.-T.; Yang, S.-F. Crosstalk between activated and inactivated c-Src in hepatocellular carcinoma. Dis. Markers 2011, 30, 325–333. [Google Scholar] [CrossRef]
- Yori, J.L.; Lozada, K.L.; Seachrist, D.D.; Mosley, J.D.; Abdul-Karim, F.W.; Booth, C.N.; Flask, C.A.; Keri, R.A. Combined SFK/mTOR inhibition prevents rapamycin-induced feedback activation of AKT and elicits efficient tumor regression. Cancer Res. 2014, 74, 4762–4771. [Google Scholar] [CrossRef] [Green Version]
- Kwong, L.N.; Davies, M.A. Targeted therapy for melanoma: Rational combinatorial approaches. Oncogene 2014, 33, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, S.; Wankell, M.; Ho, V.; White, R.; Deo, N.; Devine, C.; Dewdney, B.; Bhathal, P.; Govaere, O.; Roskams, T.; et al. Targeting mTOR and Src restricts hepatocellular carcinoma growth in a novel murine liver cancer model. PLoS ONE 2019, 14, e0212860. [Google Scholar] [CrossRef] [PubMed]
- Siveen, K.S.; Sikka, S.; Surana, R.; Dai, X.; Zhang, J.; Kumar, A.P.; Tan, B.K.; Sethi, G.; Bishayee, A. Targeting the STAT3 signaling pathway in cancer: Role of synthetic and natural inhibitors. Biochim. Biophys. Acta 2014, 1845, 136–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miklossy, G.; Hilliard, T.S.; Turkson, J. Therapeutic modulators of STAT signalling for human diseases. Nat. Rev. Drug Discov. 2013, 12, 611–629. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Yao, Z.; Bhuvaneshwar, K.; Gusev, Y.; Kallakury, B.; Yang, S.; Shetty, K.; He, A.R. Transcriptional regulation of STAT3 by SPTBN1 and SMAD3 in HCC through cAMP-response element-binding proteins ATF3 and CREB2. Carcinogenesis 2014, 35, 2393–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Zhou, L.; Xie, H.; Wang, W.; Zheng, S. Differences in antiproliferative effect of STAT3 inhibition in HCC cells with versus without HBV expression. Biochem. Biophys. Res. Commun. 2015, 461, 513–518. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, H.-Y.; Lee, S.-H.; Lee, J.-H.; Lee, J.-H.; Jang, B.K.; Kim, M.K. Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 10509. https://doi.org/10.3390/ijms221910509
Seo H-Y, Lee S-H, Lee J-H, Lee J-H, Jang BK, Kim MK. Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway. International Journal of Molecular Sciences. 2021; 22(19):10509. https://doi.org/10.3390/ijms221910509
Chicago/Turabian StyleSeo, Hye-Young, So-Hee Lee, Ji-Ha Lee, Jae-Ho Lee, Byoung Kuk Jang, and Mi Kyung Kim. 2021. "Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway" International Journal of Molecular Sciences 22, no. 19: 10509. https://doi.org/10.3390/ijms221910509
APA StyleSeo, H.-Y., Lee, S.-H., Lee, J.-H., Lee, J.-H., Jang, B. K., & Kim, M. K. (2021). Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway. International Journal of Molecular Sciences, 22(19), 10509. https://doi.org/10.3390/ijms221910509