
Zebrafish as a Model to Evaluate a CRISPR/Cas9-Based Exon Excision Approach as a Future Treatment Option for EYS-Associated Retinitis Pigmentosa

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
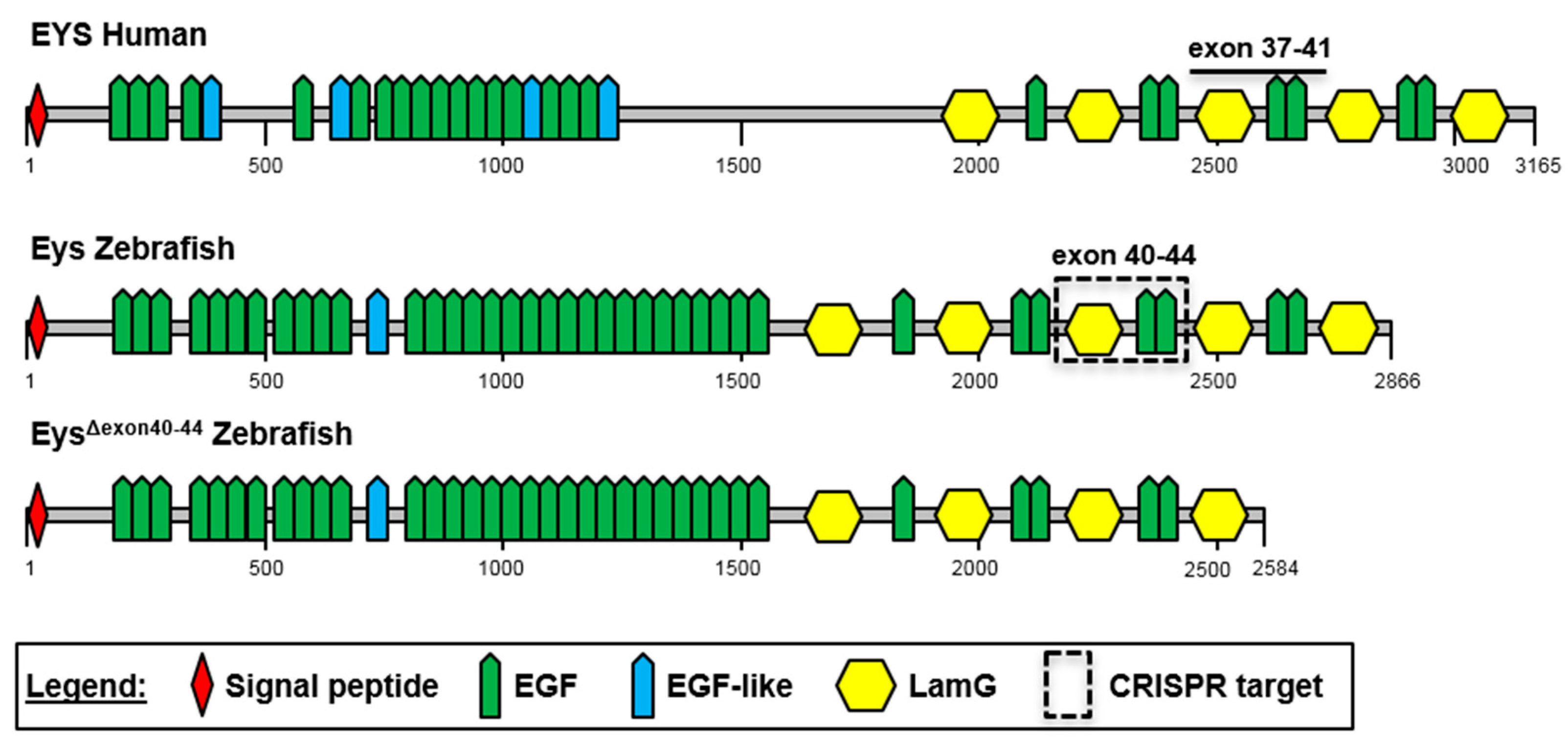
2.1. The C-Terminal Region of EYS Is Highly Conserved between Zebrafish and Man
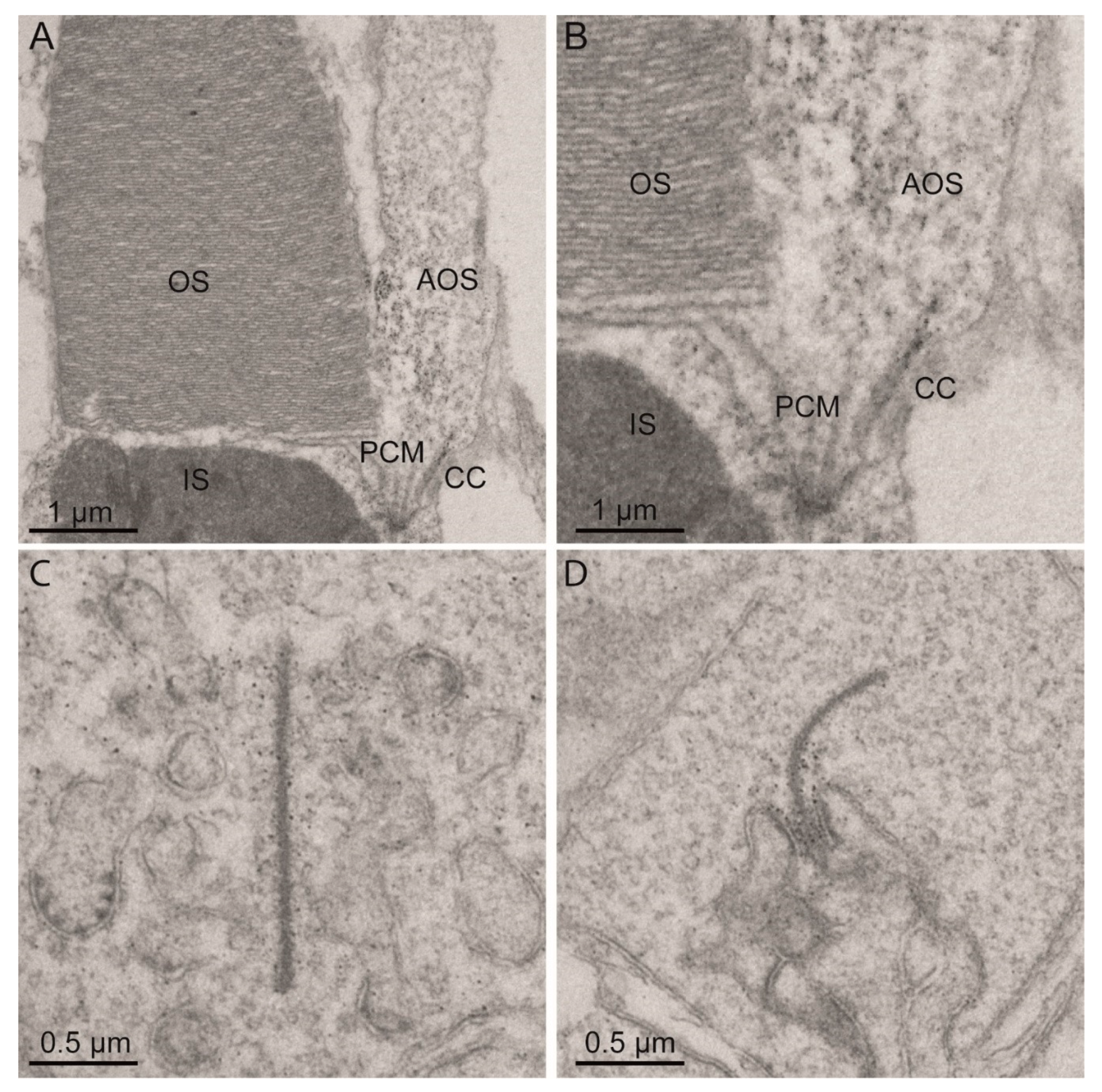
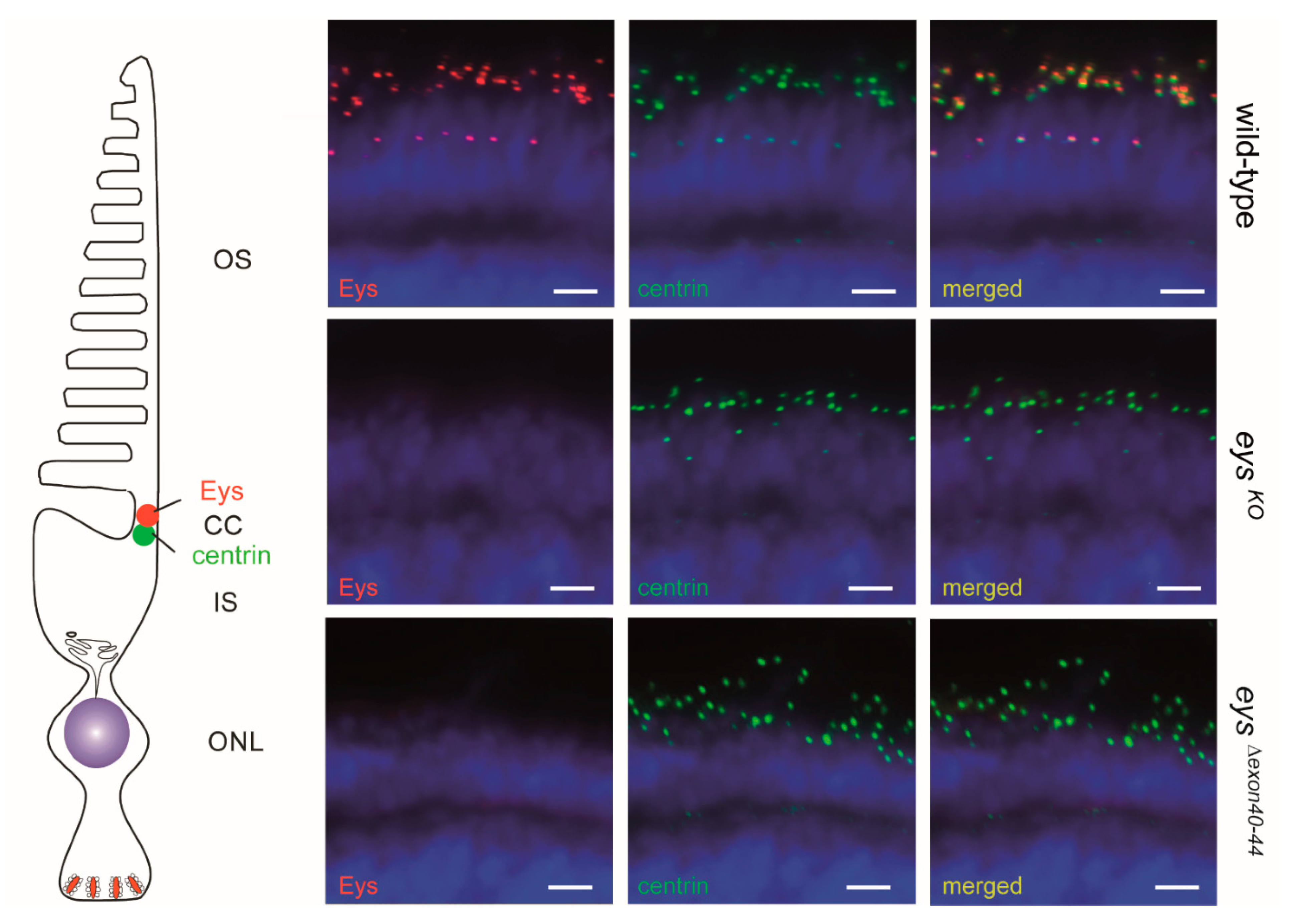
2.2. Eys Is Present at the Periciliary Membrane, Accessory Outer Segment, Connecting Cilium and Ribbon Synapse of Zebrafish Photoreceptor Cells
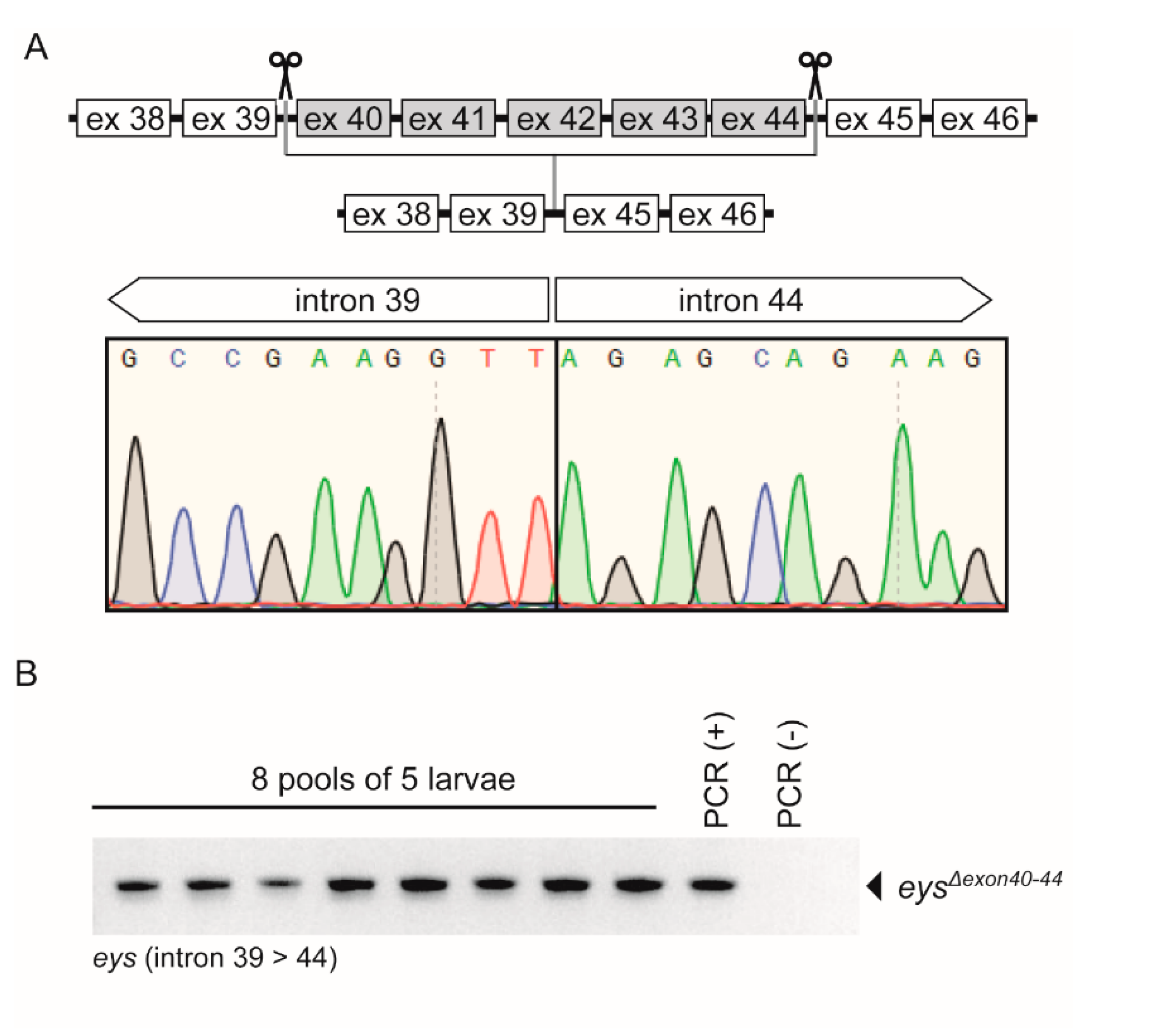
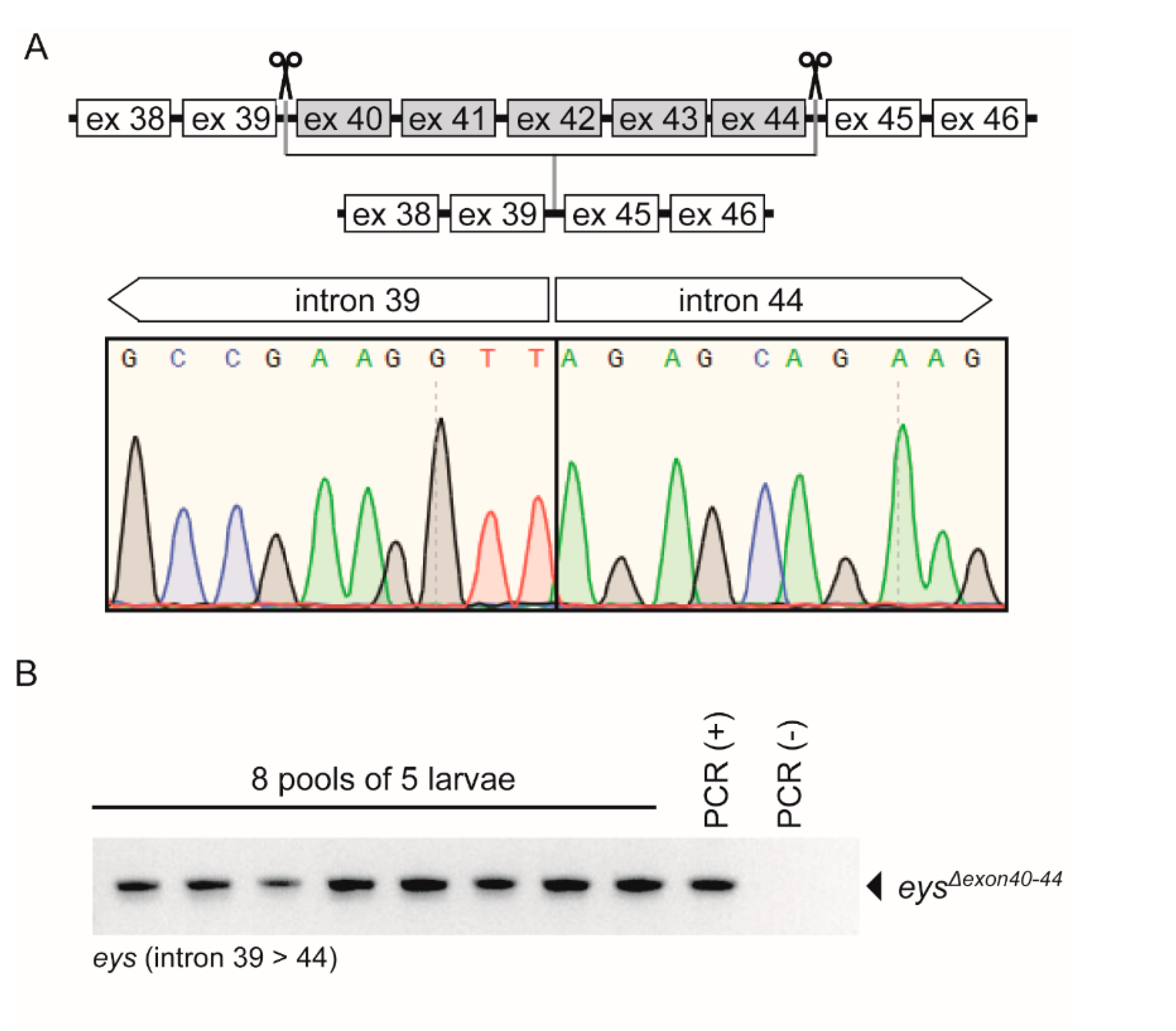
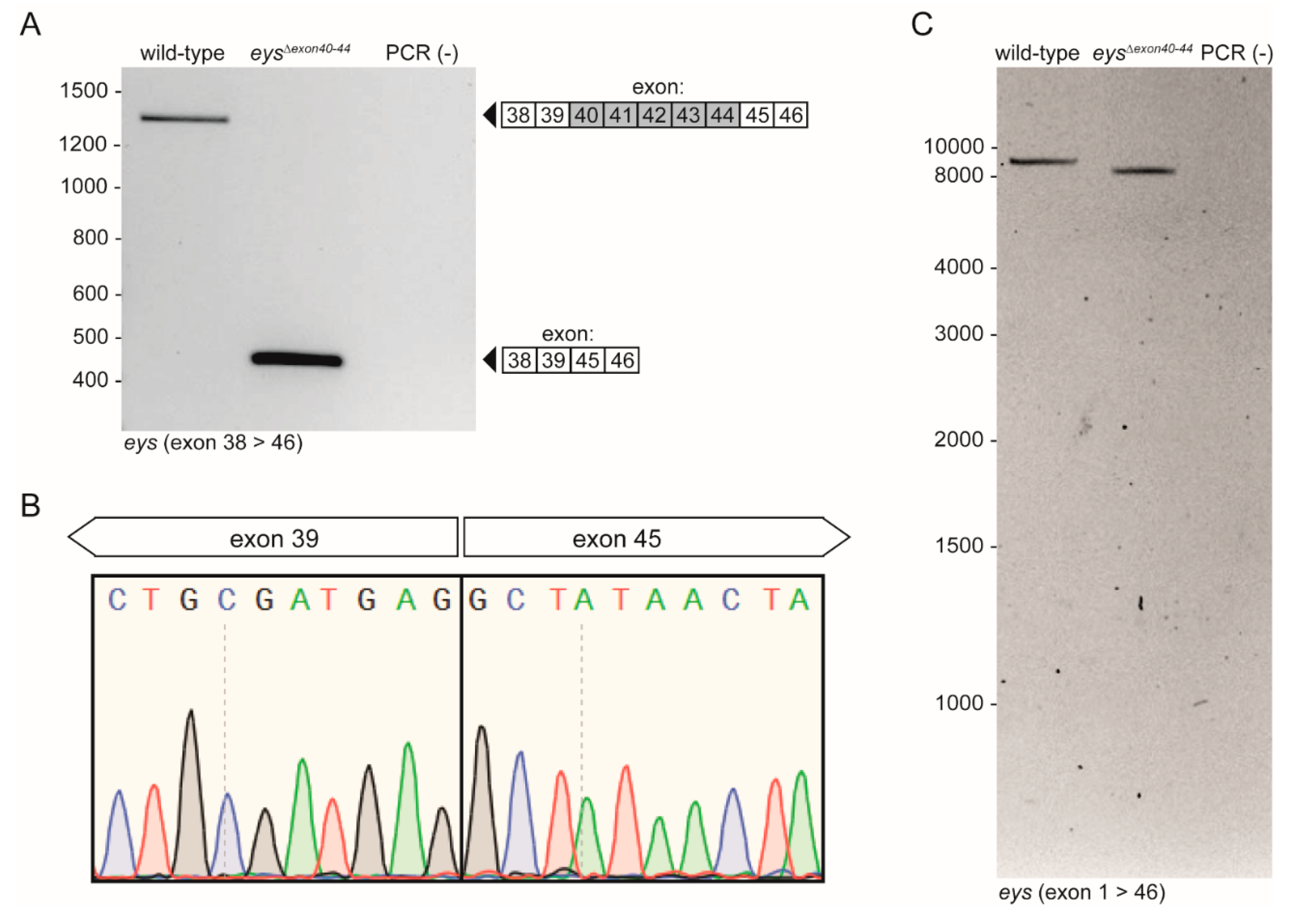
2.3. Generation of the eysΔexon40-44 Zebrafish Line Using CRISPR/Cas9 Technology
2.4. eysΔexon40-44 Transcript Analysis
2.5. Eys Is Absent from the Retina of 5 dpf eysΔexon40-44 Zebrafish
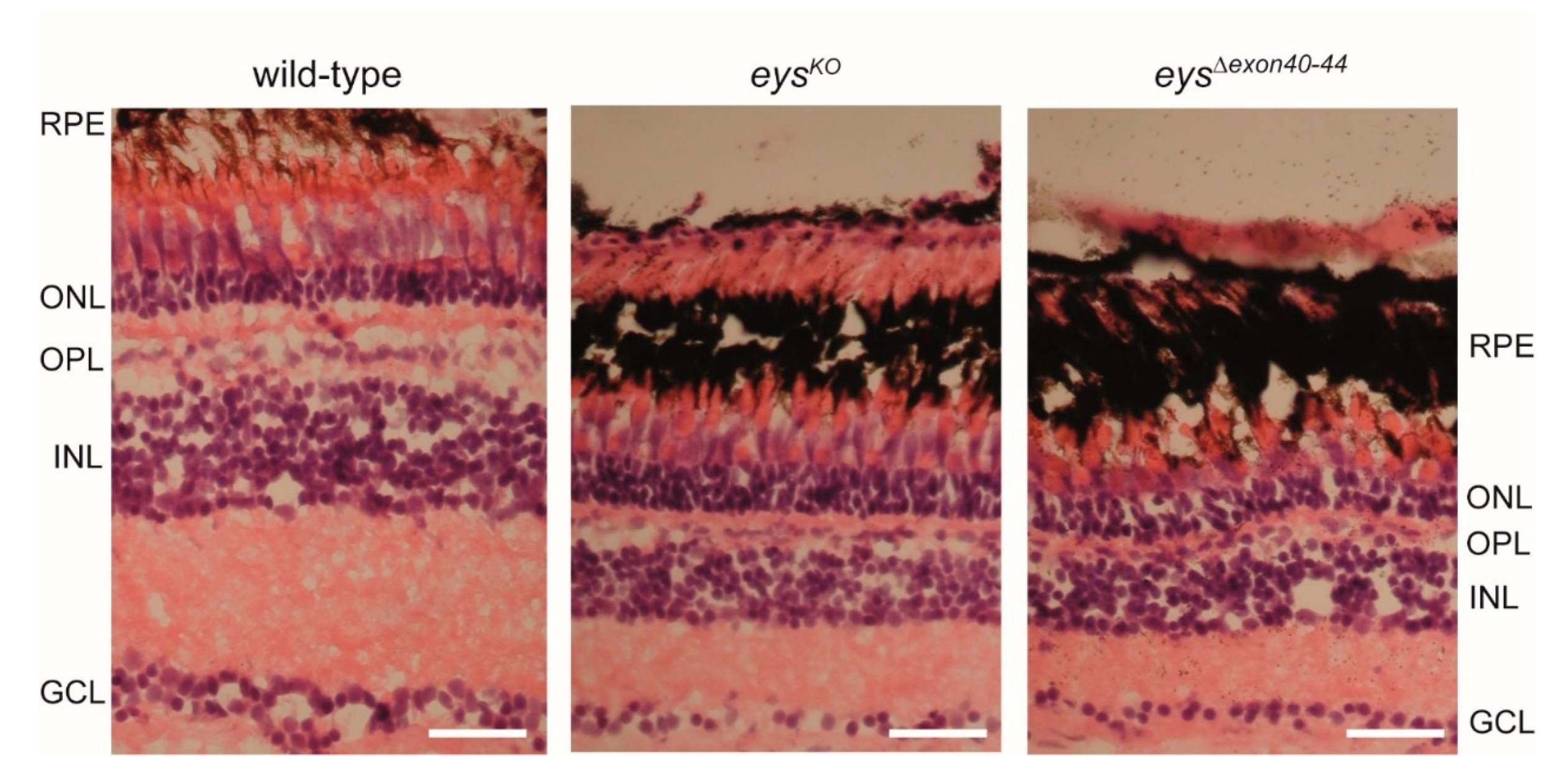
2.6. Disturbed Retinal Morphology and Disorganization of Photoreceptor Outer Segments in Adult eysΔexon40-44 Zebrafish
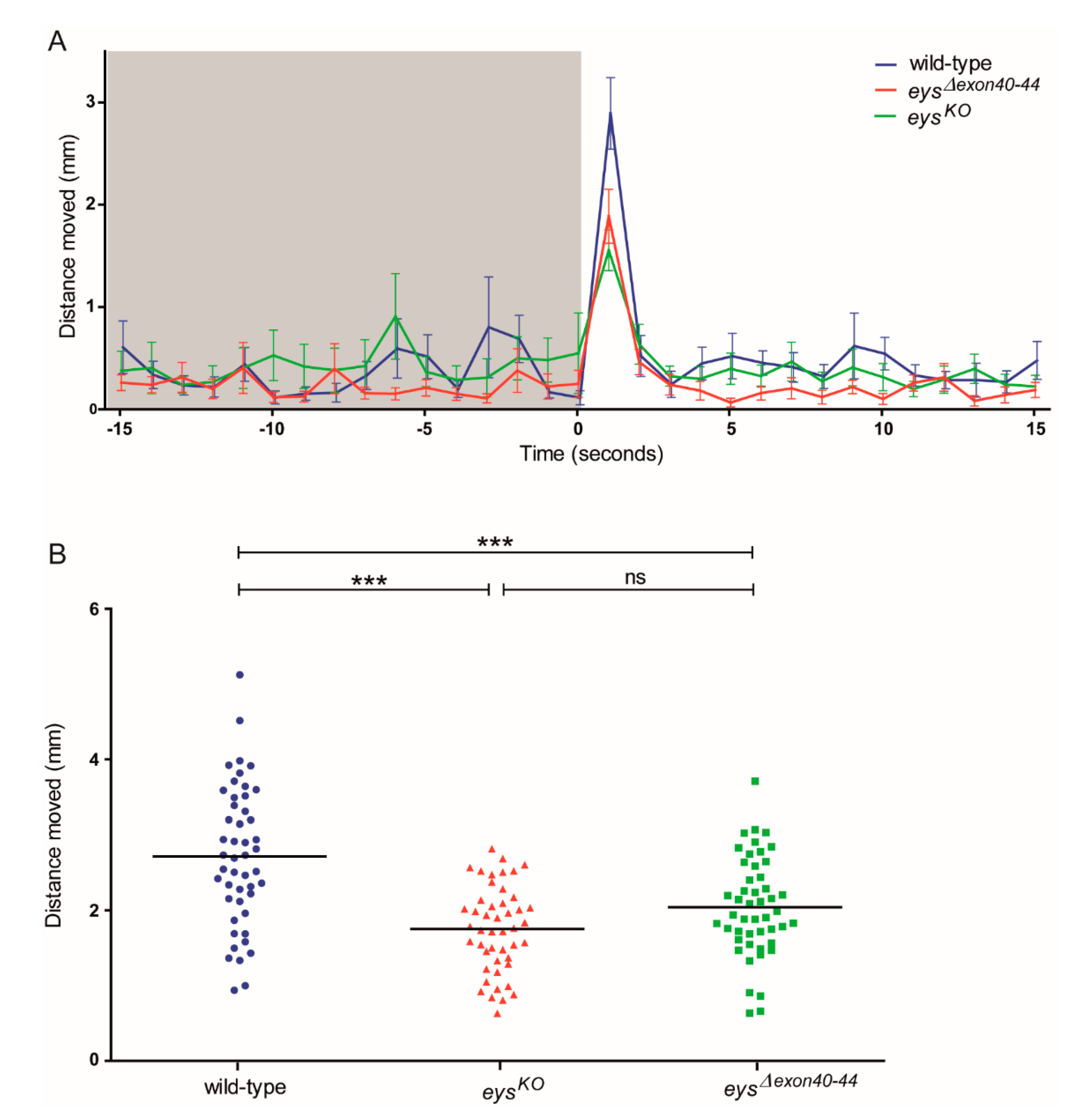
2.7. Impaired Locomotor Activity of eysΔexon40-44 Zebrafish in Response to Light
3. Discussion
4. Materials and Methods
4.1. Zebrafish Ethics, Maintenance and Husbandry
4.2. Multiple Sequence Alignment
4.3. Fixation and Pre-Embedding Labeling for Immunoelectron Microscopy
4.4. CRISPR/Cas9 Genome-Editing Design
4.5. Microinjections
4.6. Genotyping
4.7. Transcript Analysis
4.8. Targeted Transcript Sequencing
4.9. Immunohistochemistry and Histology
4.10. Visual Motor Response Assay (VMR)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verbakel, S.K.; van Huet, R.A.; Boon, C.; Hollander, A.I.D.; Collin, R.W.; Klaver, C.C.; Hoyng, C.B.; Roepman, R.; Klevering, B.J. Non-syndromic retinitis pigmentosa. Prog. Retin. Eye Res. 2018, 66, 157–186. [Google Scholar] [CrossRef] [PubMed]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef]
- Dulla, K.; Slijkerman, R.; van Diepen, H.C.; Albert, S.; Dona, M.; Beumer, W.; Turunen, J.J.; Chan, H.L.; Schulkens, I.A.; Vorthoren, L.; et al. Antisense oligonucleotide-based treatment of retinitis pigmentosa caused by USH2A exon 13 mutations. Mol. Ther. 2021, 29, 2441–2455. [Google Scholar] [CrossRef]
- El-Aziz, M.M.A.; O’Driscoll, C.A.; Kaye, R.S.; Barragan, I.; El-Ashry, M.F.; Borrego, S.; Gil, G.A.; Pang, C.P.; Webster, A.R.; Bhattacharya, S.S. Identification of novel mutations in the ortholog of drosophila eyes shut gene (EYS) causing autosomal recessive retinitis pigmentosa. Investig. Opthalmol. Vis. Sci. 2010, 51, 4266–4272. [Google Scholar] [CrossRef] [Green Version]
- Barragan, I.; Borrego, S.; Pieras, J.I.; del Pozo, M.G.; Santoyo, J.; Ayuso, C.; Baiget, M.; Millan, J.M.; Mena, M.; El-Aziz, M.M.A.; et al. Mutation spectrum of EYS in Spanish patients with autosomal recessive retinitis pigmentosa. Hum. Mutat. 2010, 31, E1772–E1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littink, K.W.; Born, L.I.V.D.; Koenekoop, R.K.; Collin, R.W.; Zonneveld, M.N.; Blokland, E.A.; Khan, H.; Theelen, T.; Hoyng, C.B.; Cremers, F.P.; et al. Mutations in the Eys gene account for approximately 5% of autosomal recessive retinitis pigmentosa and cause a fairly homogeneous phenotype. Ophthalmology 2010, 117, 2026–2033.e7. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Maeda, A.; Hirami, Y.; Ishigami, C.; Kosugi, S.; Mandai, M.; Kurimoto, Y.; Takahashi, M. Retinitis pigmentosa with EYS mutations is the most prevalent inherited retinal dystrophy in Japanese populations. J. Ophthalmol. 2015, 2015, 819760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandah-Rozenfeld, D.; Littink, K.W.; Ben-Yosef, T.; Strom, T.M.; Chowers, I.; Collin, R.W.J.; Hollander, A.I.D.; Born, L.I.V.D.; Zonneveld, M.N.; Merin, S.; et al. Novel null mutations in the Eys gene are a frequent cause of autosomal recessive retinitis pigmentosa in the Israeli population. Investig. Opthalmology Vis. Sci. 2010, 51, 4387–4394. [Google Scholar] [CrossRef] [PubMed]
- Audo, I.; Sahel, J.A.; Mohand-Saïd, S.; Lancelot, M.E.; Antonio, A.; Moskova-Doumanova, V.; Nandrot, E.F.; Doumanov, J.; Barragan, I.; Antinolo, G. EYS is a major gene for rod-cone dystrophies in France. Hum. Mutat. 2010, 31, E1406–E1435. [Google Scholar] [CrossRef] [Green Version]
- Collin, R.W.; Littink, K.W.; Klevering, B.J.; Born, L.I.V.D.; Koenekoop, R.K.; Zonneveld, M.N.; Blokland, E.A.; Strom, T.M.; Hoyng, C.B.; Hollander, A.I.D.; et al. Identification of a 2 Mb Human Ortholog of Drosophila eyes shut/spacemaker that Is Mutated in Patients with Retinitis Pigmentosa. Am. J. Hum. Genet. 2008, 83, 594–603. [Google Scholar] [CrossRef] [Green Version]
- El-Aziz, M.M.A.; Barragan, I.; O’Driscoll, C.; Goodstadt, L.; Prigmore, E.; Borrego, S.; Mena, M.; Pieras, J.I.; El-Ashry, M.F.; Abu Safieh, L.; et al. EYS, encoding an ortholog of Drosophila spacemaker, is mutated in autosomal recessive retinitis pigmentosa. Nat. Genet. 2008, 40, 1285–1287. [Google Scholar] [CrossRef] [Green Version]
- Alfano, G.; Kruczek, P.M.; Shah, A.Z.; Kramarz, B.; Jeffery, G.; Zelhof, A.C.; Bhattacharya, S.S. EYS Is a Protein Associated with the Ciliary Axoneme in Rods and Cones. PLoS ONE 2016, 11, e0166397. [Google Scholar] [CrossRef]
- Tucker, B.A.; Mullins, R.F.; Streb, L.M.; Anfinson, K.; Eyestone, M.E.; Kaalberg, E.; Riker, M.J.; Drack, A.V.; Braun, T.A.; Stone, E.M. Patient-specific iPSC-derived photoreceptor precursor cells as a means to investigate retinitis pigmentosa. Elife 2013, 2, e00824. [Google Scholar] [CrossRef] [PubMed]
- Slijkerman, R.W.; Song, F.; Astuti, G.D.; Huynen, M.A.; van Wijk, E.; Stieger, K.; Collin, R.W. The pros and cons of vertebrate animal models for functional and therapeutic research on inherited retinal dystrophies. Prog. Retin. Eye Res. 2015, 48, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Hu, X.; Liu, F.; Soares, D.; Liu, X.; Yu, S.; Gao, M.; Han, S.; Qin, Y.; Li, C.; et al. Ablation of EYS in zebrafish causes mislocalisation of outer segment proteins, F-actin disruption and cone-rod dystrophy. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, Y.; Li, J.; Natale, B.N.; Cao, S.; Wang, D.; Amack, J.D.; Hu, H. Eyes shut homolog is required for maintaining the ciliary pocket and survival of photoreceptors in zebrafish. Biol. Open 2016, 5, 1662–1673. [Google Scholar] [CrossRef] [Green Version]
- Messchaert, M.; Dona, M.; Broekman, S.; Peters, T.; Corral-Serrano, J.C.; Slijkerman, R.W.N.; Van Wijk, E.; Collin, R.W.J. Eyes shut homolog is important for the maintenance of photoreceptor morphology and visual function in zebrafish. PLoS ONE 2018, 13, e0200789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaran, N.; Michaelides, M.; Smith, A.J.; Ali, R.; Bainbridge, J.W.B. Retinal gene therapy. Br. Med Bull. 2018, 126, 13–25. [Google Scholar] [CrossRef]
- Kole, R.; Krieg, A.M. Exon skipping therapy for Duchenne muscular dystrophy. Adv. Drug Deliv. Rev. 2015, 87, 104–107. [Google Scholar] [CrossRef]
- Rutten, J.W.; Dauwerse, H.G.; Peters, D.J.; Goldfarb, A.; Venselaar, H.; Haffner, C.; van Ommen, G.-J.B.; Aartsma-Rus, A.M.; Lesnik Oberstein, S.A. Therapeutic NOTCH3 cysteine correction in CADASIL using exon skipping. Vitr. Proof Concept. Brain 2016, 139, 1123–1135. [Google Scholar]
- Hodel, C.; Niklaus, S.; Heidemann, M.; Klooster, J.; Kamermans, M.; Biehlmaier, O.; Gesemann, M.; Neuhauss, S.C. Myosin VIIA is a Marker for the Cone Accessory Outer Segment in Zebrafish. Anat. Rec. 2014, 297, 1777–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modrzejewski, D.; Hartung, F.; Lehnert, H.; Sprink, T.; Kohl, C.; Keilwagen, J.; Wilhelm, R. Which Factors Affect the Occurrence of Off-Target Effects Caused by the Use of CRISPR/Cas: A Systematic Review in Plants. Front. Plant Sci. 2020, 11, 1838. [Google Scholar] [CrossRef]
- Venkatraman, P.; Carmer, R.; Pang, C.P.; Zhang, M.; Leung, Y.F. Understanding the contribution of photoreceptors to the Visual Motor Response. Investig. Ophthalmol. Vis. Sci. 2015, 56, 998. [Google Scholar]
- Liu, Y.; Carmer, R.; Zhang, G.; Venkatraman, P.; Brown, S.A.; Pang, C.-P.; Zhang, M.; Ma, P.; Leung, Y.F. Statistical Analysis of Zebrafish Locomotor Response. PLoS ONE 2015, 10, e0139521. [Google Scholar] [CrossRef] [Green Version]
- Iwanami, M.; Oshikawa, M.; Nishida, T.; Nakadomari, S.; Kato, S. High prevalence of mutations in the EYS gene in Japanese patients with autosomal recessive retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Iwanami, M.; Oishi, A.; Ogino, K.; Seko, Y.; Nishida-Shimizu, T.; Yoshimura, N.; Kato, S. Five major sequence variants and copy number variants in the EYS gene account for one-third of Japanese patients with autosomal recessive and simplex retinitis pigmentosa. Mol. Vis. 2019, 25, 766–779. [Google Scholar]
- Yacob, A.; Wise, C.; Kunz, Y.W. The accessory outer segment of rods and cones in the retina of the guppy, Poecilia reticulata P. (teleostei). Cell Tissue Res. 1977, 177, 181–193. [Google Scholar] [CrossRef]
- Burnside, B.; Wang, E.; Pagh-Roehl, K.; Rey, H. Retinomotor movements in isolated teleost retinal cone inner-outer segment preparations (CIS-COS): Effects of light, dark and dopamine. Exp. Eye Res. 1993, 57, 709–722. [Google Scholar] [CrossRef]
- Dona, M.; Slijkerman, R.; Lerner, K.; Broekman, S.; Wegner, J.; Howat, T.; Peters, T.; Hetterschijt, L.; Boon, N.; de Vrieze, E.; et al. Usherin defects lead to early-onset retinal dysfunction in zebrafish. Exp. Eye Res. 2018, 173, 148–159. [Google Scholar] [CrossRef]
- Richard, M.; Roepman, R.; Aartsen, W.M.; Van Rossum, A.G.; Hollander, A.I.D.; Knust, E.; Wijnholds, J.; Cremers, F.P. Towards understanding CRUMBS function in retinal dystrophies. Hum. Mol. Genet. 2006, 15, R235–R243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maerker, T.; Van Wijk, E.; Overlack, N.; Kersten, F.F.; McGee, J.; Goldmann, T.; Sehn, E.; Roepman, R.; Walsh, E.J.; Kremer, H.; et al. A novel Usher protein network at the periciliary reloading point between molecular transport machineries in vertebrate photoreceptor cells. Hum. Mol. Genet. 2007, 17, 71–86. [Google Scholar] [CrossRef]
- Wouters, M.A.; Rigoutsos, I.; Chu, C.K.; Feng, L.L.; Sparrow, D.; Dunwoodie, S. Evolution of distinct EGF domains with specific functions. Protein Sci. 2005, 14, 1091–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tisi, D.; Talts, J.F.; Timpl, R.; Hohenester, E. Structure of the C-terminal laminin G-like domain pair of the laminin α2 chain harbouring binding sites for α-dystroglycan and heparin. EMBO J. 2000, 19, 1432–1440. [Google Scholar] [CrossRef]
- Russell, S.; Bennett, J.; Wellman, J.A.; Chung, D.C.; Yu, Z.-F.; Tillman, A.; Wittes, J.; Pappas, J.; Elci, O.; McCague, S.; et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65 -mediated inherited retinal dystrophy: A randomised, controlled, open-label, phase 3 trial. Lancet 2017, 390, 849–860. [Google Scholar] [CrossRef]
- Maguire, A.M.; Russell, S.; Wellman, J.A.; Chung, D.C.; Yu, Z.-F.; Tillman, A.; Wittes, J.; Pappas, J.; Elci, O.; Marshall, K.A. Efficacy, safety, and durability of voretigene neparvovec-rzyl in RPE65 mutation-associated inherited retinal dystrophy: Results of phase 1 and 3 trials. Ophthalmology 2019, 126, 1273–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemayel, M.C.; Bhatwadekar, A.D.; Ciulla, T. RNA therapeutics for retinal diseases. Expert Opin. Biol. Ther. 2021, 21, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Lesman, D.; Rodriguez, Y.; Rajakumar, D.; Wein, N. U7 snRNA, a small RNA with a big impact in gene therapy. Hum. Gene Ther. 2021. [Google Scholar] [CrossRef]
- Kuijper, E.C.; Bergsma, A.J.; Pijnappel, W.P.; Aartsma-Rus, A. Opportunities and challenges for antisense oligonucleotide therapies. J. Inherit. Metab. Dis. 2021, 44, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.R.; Drack, A.V.; Cideciyan, A.V.; Jacobson, S.G.; Leroy, B.P.; Pfeifer, W.L.; Dumitrescu, A.V.; Garafolo, A.V.; Ho, A.C.; Van Cauwenbergh, C. Results of a phase 1b/2 trial of intravitreal (IVT) sepofarsen (QR-110) antisense oligonucleotide in Leber congenital amaurosis 10 (LCA10) due to p. Cys998X mutation in the CEP290 gene. Investig. Ophthalmol. Vis. Sci. 2020, 61, 866. [Google Scholar]
- Niks, E.; Aartsma-Rus, A. Exon skipping: A first in class strategy for Duchenne muscular dystrophy. Expert Opin. Biol. Ther. 2016, 17, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Syed, Y.Y. Eteplirsen: First Global Approval. Drugs 2016, 76, 1699–1704. [Google Scholar] [CrossRef]
- Heo, Y.-A. Golodirsen: First Approval. Drugs 2020, 80, 329–333. [Google Scholar] [CrossRef]
- Frank, D.E.; Schnell, F.J.; Akana, C.; El-Husayni, S.H.; Desjardins, C.A.; Morgan, J.; Charleston, J.S.; Sardone, V.; Domingos, J.; Dickson, G.; et al. Increased dystrophin production with golodirsen in patients with Duchenne muscular dystrophy. Neurology 2020, 94, e2270–e2282. [Google Scholar] [CrossRef] [Green Version]
- Mendell, J.R.; Rodino-Klapac, L.R.; Sahenk, Z.; Roush, K.; Bird, L.; Lowes, L.P.; Alfano, L.; Gomez, A.M.; Lewis, S.; Kota, J. Eteplirsen for the treatment of Duchenne muscular dystrophy. Ann. Neurol. 2013, 74, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Mendell, J.R.; Goemans, N.; Lowes, L.P.; Alfano, L.N.; Berry, K.; Shao, J.; Kaye, E.M.; Mercuri, E.; Group, E.S.; Network, T.F.D.I.; et al. Longitudinal effect of eteplirsen versus historical control on ambulation in D uchenne muscular dystrophy. Ann. Neurol. 2016, 79, 257–271. [Google Scholar] [CrossRef]
- Aartsma-Rus, A.; Janson, A.A.; Kaman, W.E.; Bremmer-Bout, M.; van Ommen, G.-J.B.; Dunnen, J.T.D.; van Deutekom, J.C. Antisense-Induced Multiexon Skipping for Duchenne Muscular Dystrophy Makes More Sense. Am. J. Hum. Genet. 2004, 74, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Pendse, N.D.; Lamas, V.; Pawlyk, B.S.; Maeder, M.L.; Chen, Z.-Y.; Pierce, E.A.; Liu, Q. In Vivo Assessment of Potential Therapeutic Approaches for USH2A-Associated Diseases. In Retinal Degenerative Diseases; Springer: Berlin, Germany, 2019; Volume 1185, pp. 91–96. [Google Scholar] [CrossRef]
- Rose, A.B. The effect of intron location on intron-mediated enhancement of gene expression in Arabidopsis. Plant J. 2004, 40, 744–751. [Google Scholar] [CrossRef]
- Shaul, O. How introns enhance gene expression. Int. J. Biochem. Cell Biol. 2017, 91, 145–155. [Google Scholar] [CrossRef]
- Erdmann-Pham, D.D.; Duc, K.D.; Song, Y.S. The Key Parameters that Govern Translation Efficiency. Cell Syst. 2020, 10, 183–192.e6. [Google Scholar] [CrossRef] [Green Version]
- Messchaert, M.; Haer-Wigman, L.; Khan, M.I.; Cremers, F.P.; Collin, R.W. EYS mutation update: In silico assessment of 271 reported and 26 novel variants in patients with retinitis pigmentosa. Hum. Mutat. 2018, 39, 177–186. [Google Scholar] [CrossRef]
- Bremer, J.; van der Heijden, E.H.; Eichhorn, D.S.; Meijer, R.; Lemmink, H.H.; Scheffer, H.; Sinke, R.J.; Jonkman, M.F.; Pasmooij, A.M.; Akker, P.C.V.D. Natural Exon Skipping Sets the Stage for Exon Skipping as Therapy for Dystrophic Epidermolysis Bullosa. Mol. Ther. Nucleic Acids 2019, 18, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish, 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Sedmak, T.; Sehn, E.; Wolfrum, U. Immunoelectron Microscopy of Vesicle Transport to the Primary Cilium of Photoreceptor Cells. Micropatterning Cell Biol. 2009, 94, 259–272. [Google Scholar] [CrossRef]
- Sedmak, T.; Wolfrum, U. Intraflagellar transport molecules in ciliary and nonciliary cells of the retina. J. Cell Biol. 2010, 189, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Montague, T.G.; Cruz, J.M.; Gagnon, J.A.; Church, G.M.; Valen, E. CHOPCHOP: A CRISPR/Cas9 and TALEN web tool for genome editing. Nucleic Acids Res. 2014, 42, W401–W407. [Google Scholar] [CrossRef] [Green Version]
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R.; et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, J.A.; Valen, E.; Thyme, S.; Huang, P.; Ahkmetova, L.; Pauli, A.; Montague, T.G.; Zimmerman, S.; Richter, C.; Schier, A.F. Efficient Mutagenesis by Cas9 Protein-Mediated Oligonucleotide Insertion and Large-Scale Assessment of Single-Guide RNAs. PLoS ONE 2014, 9, e98186. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schellens, R.; de Vrieze, E.; Graave, P.; Broekman, S.; Nagel-Wolfrum, K.; Peters, T.; Kremer, H.; Collin, R.W.J.; van Wijk, E. Zebrafish as a Model to Evaluate a CRISPR/Cas9-Based Exon Excision Approach as a Future Treatment Option for EYS-Associated Retinitis Pigmentosa. Int. J. Mol. Sci. 2021, 22, 9154. https://doi.org/10.3390/ijms22179154
Schellens R, de Vrieze E, Graave P, Broekman S, Nagel-Wolfrum K, Peters T, Kremer H, Collin RWJ, van Wijk E. Zebrafish as a Model to Evaluate a CRISPR/Cas9-Based Exon Excision Approach as a Future Treatment Option for EYS-Associated Retinitis Pigmentosa. International Journal of Molecular Sciences. 2021; 22(17):9154. https://doi.org/10.3390/ijms22179154
Chicago/Turabian StyleSchellens, Renske, Erik de Vrieze, Pam Graave, Sanne Broekman, Kerstin Nagel-Wolfrum, Theo Peters, Hannie Kremer, Rob W. J. Collin, and Erwin van Wijk. 2021. "Zebrafish as a Model to Evaluate a CRISPR/Cas9-Based Exon Excision Approach as a Future Treatment Option for EYS-Associated Retinitis Pigmentosa" International Journal of Molecular Sciences 22, no. 17: 9154. https://doi.org/10.3390/ijms22179154
APA StyleSchellens, R., de Vrieze, E., Graave, P., Broekman, S., Nagel-Wolfrum, K., Peters, T., Kremer, H., Collin, R. W. J., & van Wijk, E. (2021). Zebrafish as a Model to Evaluate a CRISPR/Cas9-Based Exon Excision Approach as a Future Treatment Option for EYS-Associated Retinitis Pigmentosa. International Journal of Molecular Sciences, 22(17), 9154. https://doi.org/10.3390/ijms22179154