
Bioactive Peptides as Potential Nutraceuticals for Diabetes Therapy: A Comprehensive Review


Abstract
:1. Introduction
2. Functional Foods and Bioactive Peptides
3. Current Diabetes Targets
4. Peptide Drugs and Diabetes
5. Antidiabetic Peptides from Food Sources
5.1. Animal Products
5.1.1. Dairy Foods

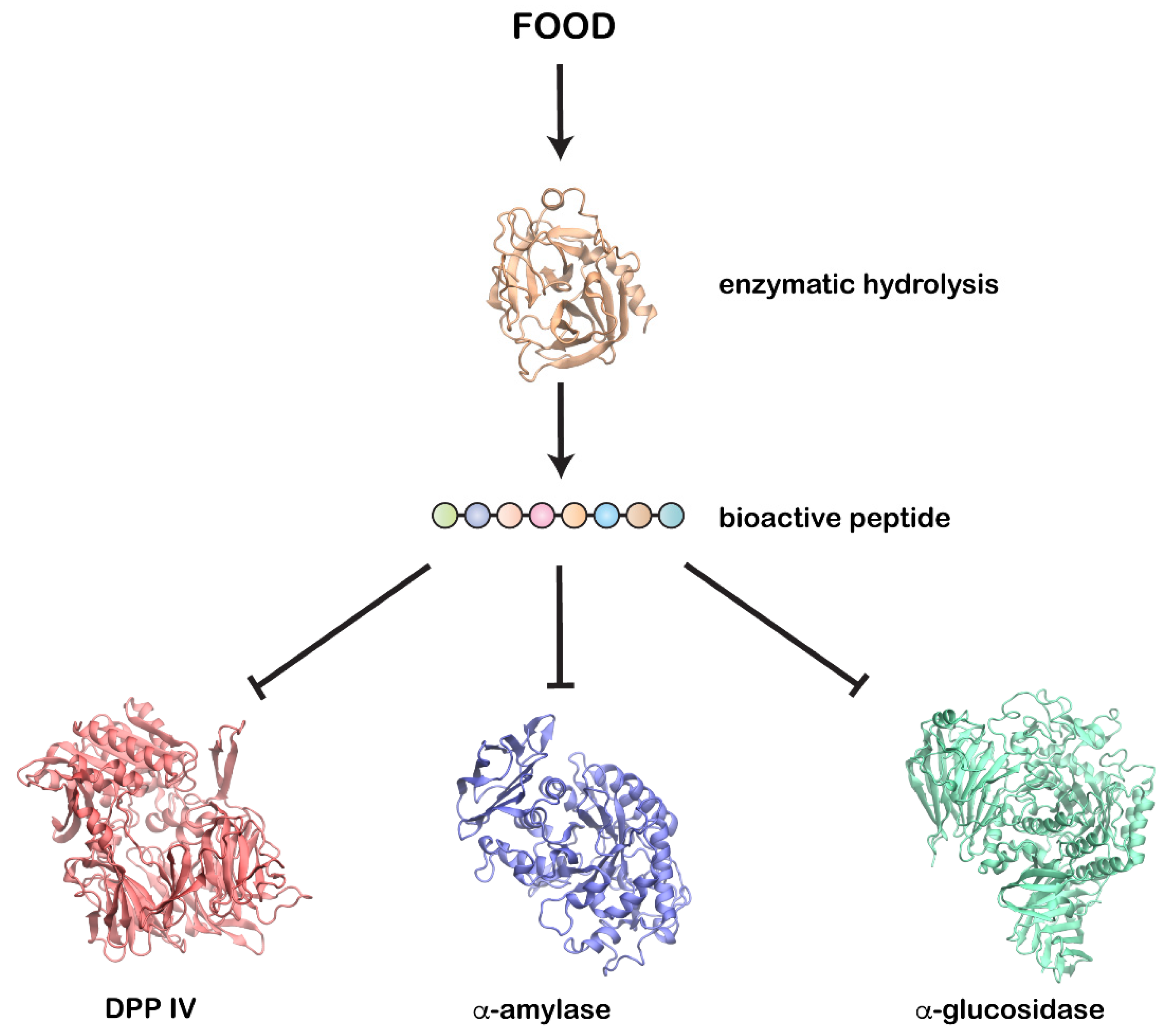{kind=link}
| Source | Enzyme(s) Used | Substrate | Protein Hydrolysate | Peptide(s) Identified | Mechanism of Action | IC50 | Reference |
|---|---|---|---|---|---|---|---|
| Cow milk | Trypsin | Milk protein | Milk protein hydrolysates | Reduced blood plasma glucose level in vivo | [52] | ||
| Donkey milk | Pepsin and pancreatin | Milk protein hydrolysates | Improved damaged β-cells viability in vitro, reduced blood glucose level and improved insulin resistance in vivo | [53] | |||
| Goat milk | Flavourzyme | Casein | Casein hydrolysate | SDIPNPIGSE | Ameliorated insulin resistance in HepG2 cells | [54] | |
| NPWDQVKR | |||||||
| SLSSSEESITH | |||||||
| QEPVLGPVRGPFP | |||||||
| Goat milk | Trypsin and chymotrypsin | Casein | Casein hydrolysate | MHQPPQPL | Inhibited DPP IV in vitro | 350.41 ± 4.1 μM | [55] |
| SPTVMFPPQSVL | 676.31 ± 12.6 μM | ||||||
| VMFPPQSVL | Nd | ||||||
| INNQFLPYPY | 40.08 ± 5.0 μM | ||||||
| AWPQYL | Nd | ||||||
| Camel milk | Alcalase, bromelain, and papain | Milk proteins | Milk protein hydrolysates | MPSKPPLL | Inhibited pancreatic α-amylase in vitro | [56] | |
| KDLWDDFKGL | |||||||
| Camel milk | Trypsin | Milk proteins | Milk protein hydrolysates | FLQY | Inhibited DPP IV in vitro | >1000 μM | [57] |
| FQLGASPY | >1000 μM | ||||||
| ILDKEGIDY | 347.8 ± 42.8 μM | ||||||
| ILELA | 721.1 ± 46.3 μM | ||||||
| LLQLEAIR | 177.8 ± 22.2 μM | ||||||
| LPVP | 87.0 ± 3.2 μM | ||||||
| LQALHQGQIV | >1000 μM | ||||||
| MPVQA | 93.3 ± 8.0 μM | ||||||
| SPVVPF | 214.1 ± 16.7 μM | ||||||
| Gouda cheese | Microbial fermentation | Water-soluble extracts (WSEs) | LPQNIPPL | Reduced glucose level in vivo and inhibited DPP IV in vivo and in vitro | 46 μM | [60] | |
| LPQ | 82 μM | ||||||
| VPITPTL | 110 μM | ||||||
| VPITPT | 130 μM | ||||||
| Egg | Alcalase | Egg white protein | Egg white protein hydrolysates | RVPSLM | Inhibited α-glucosidase in vitro | 23.07 μmol/L | [61] |
| TPSPR | 40.02 μmol/L | ||||||
| DLQGK | >150.0 μmol/L | ||||||
| AGLAPY | >150.0 μmol/L | ||||||
| RVPSL | >150.0 μmol/L | ||||||
| DHPFLF | >150.0 μmol/L | ||||||
| HAEIN | >150.0 μmol/L | ||||||
| QIGLF | >150.0 μmol/L | ||||||
| Egg | Pepsin | Egg yolk protein | Egg yolk protein hydrolysates | YINQMPQKSRE, | Inhibited DPP IV in vitro | 222.8 μg/mL | [62] |
| VTGRFAGHPAAQ | Inhibited α-glucosidase in vitro | 365.4 μg/mL | |||||
| Turtle egg | Trypsin, pepsin, α-chymotrypsin, thermolysin, and GI enzyme | Egg yolk protein | Egg yolk protein hydrolysates | LPSW | Inhibited DPP IV in vitro and in silico | 289.2 ± 11.85 μM | [63] |
| WLQL | 269.7 ± 15.91 μM | ||||||
| LPLF | 463.6 ± 5.52 μM | ||||||
| VPGLAL | >2000 | ||||||
| LVGLPL | 432.5 ± 40.31 μM | ||||||
| Chicken egg | Pepsin and trypsin | Myosin and lysozyme | ADF | Inhibited DPP IV in vitro and in silico | 16.83 mM | [64] | |
| MIR | 4.86 mM | ||||||
| FGR | 46.2 mM | ||||||
| Cooked meat (pork, beef, chicken, and turkey) | Amylase, pepsin and trypsin | Meat proteins | Meat protein hydrolysates | IPI | Inhibited DPP IV in vitro | 3.5 μmol/L | [65] |
| WL | 44 μmol/L | ||||||
| LPL | 241 μmol/L | ||||||
| WI | 89 μmol/L | ||||||
| FL | 400 μmol/L | ||||||
| LW | 993 μmol/L | ||||||
| Cooked meat (Pork, chicken and turkey) | VL | 74 μmol/L | |||||
| WM | 243 μmol/L | ||||||
| ML | 91 μmol/L | ||||||
| Cooked meat (Pork and beef) | IP | 150 μmol/L | |||||
| LP | 712 μmol/L | ||||||
| AL | 882 μmol/L | ||||||
| Cooked meat (Chicken and turkey) | FP | 363 μmol/L | |||||
| Cooked meat (Pork) | IPM | 70 μmol/L | |||||
| Dry cured ham | KA | Inhibited DPP IV in vitro | 6.27 ± 0.59 mM | [66] | |||
| AA | 9.40 ± 0.10 mM | ||||||
| GP | 9.69 ± 0.49 mM | ||||||
| PL | >10 mM | ||||||
| AAATP | 6.47 ± 0.20 mM | ||||||
| AAAAG | 8.13 ± 0.48 mM | ||||||
| ALGGA | >10 mM | ||||||
| LVSGM | >10 mM | ||||||
| Chicken feet | Neutrase and protamex | Feet protein | Feet protein hydrolysates | Stimulated GLP-1 release in vivo and inhibited DPP IV in vitro | 4.42 ± 0.1 μL | [67] | |
| Chicken by product | Flavourzyme and corolase | Mechanical chicken deboning residue | Protein hydrolysates | TL | Promoted glucose uptake and inhibited DPP IV in vitro | [68] | |
| HT | |||||||
| LA | |||||||
| LADVEVDLL | |||||||
| LL | |||||||
| ETGKGEDGE | |||||||
| FL | |||||||
| LFFSMLLML | |||||||
| LF |
5.1.2. Eggs
5.1.3. Meat Products
5.2. Fish Products
5.3. Plant Products
5.3.1. Pulses and Legumes
5.3.2. Cereals and Pseudocereals
5.3.3. Plant Seeds and Nuts
| Source | Enzyme(s) Used | Substrate | Proteins Hydrolysate | Peptide(s) Identified | Mechanism of Action | IC50 | Reference |
|---|---|---|---|---|---|---|---|
| Soy bean | ASCNGVCSPFEMPPCGSSACRC IPVGLVVGYCRHPSG (aglycin) | Enhanced insulin receptor signalling pathway and enhanced glucose uptake in vivo | [85] | ||||
| Soy bean | VSCNGVCSPFEMPPCGSSACR CIPYGLVVGNCRHPSG (vglycin) | Enhanced insulin signalling by activating the insulin receptor (IR)/Akt signalling pathway | [86,87] | ||||
| Restored impaired insulin signalling, glucose tolerance and pancreatic function in vivo | |||||||
| Soy bean | β-conglycinin | YPFVV (soymorphin-5) | Improved glucose and lipid metabolism via activation of the adiponectin and PPARα system- in vivo | [88] | |||
| Soy bean | Pepsin and pancreatin | Germinated soybeans | Protein hydrolysates | Inhibited DPP IV, α-amylase and α-glucosidase in vitro | DPP-IV inhibition- 1.49 mg/mL) | [89] | |
| α-amylase inhibition- 1.70 mg/mL | |||||||
| Maltase and sucrase activities of α-glucosidase 3.73 and 2.90 mg/mL | |||||||
| Lupin bean | LTFPGSAED | Inhibited DPP IV in situ | 207.5 µM | [90] | |||
| Soy bean | IAVPTGV | 223.2 µM | |||||
| Fermented soy bean - Natto | Water soluble extract | KL | Inhibited DPP IV in vitro | 41.40 ± 2.68 μg/mL | [96] | ||
| LA | 598.02 ± 18.35 μg/mL | ||||||
| Black bean | Proteinase K, pepsin, trypsin, papain, alcalase, flavourzyme, themolysin, and chymotrypsin | Protein hydrolysates | EGLELLLLLLAG | Inhibited DPP IV in vitro and in silico | [97] | ||
| AKSPLF | |||||||
| FEELN | |||||||
| TTGGKGGK | Inhibited α-glucosidase in vitro and in silico | ||||||
| AKSPLF | |||||||
| WEVM | Inhibited α-amylase in vitro and in silico | ||||||
| Pinto beans | Protamex | Protein hydrolysates | PPHMLP | Inhibited α-amylase in vitro | 1.97 mg/mL | [98,99] | |
| PLPTGAGF | 8.96 mg/mL | ||||||
| PPHMGGP | 14.63 mg/mL | ||||||
| PLPLHMLP | 18.45 mg/mL | ||||||
| LSSLEMGSLGALFVCM | 20.56 mg/mL | ||||||
| Common beans | Pepsin, and pancreatin | Protein hydrolysates | Lowered blood glucose level by inhibiting α-glucosidase | [100] | |||
| Bambara bean | Alcalase and thermolysin | Protein hydrolysates | Inhibited DPP IV in vitro | 1.73 mg/mL | [101] | ||
| Cow pea | Peptides | Activated the insulin signalling pathway in vivo | [102] | ||||
| Chickpea | Pepsin, pancreatin and bromelain | Protein hydrolysates | PHPATSGGGL | Inhibited DPP IV in vitro | 245 μg/mL | [103] | |
| YVDGSGTPLT | |||||||
| Oats | Pepsin, trypsin, pancreatin, and alcalase | Globulin | LQAFEPLR | Inhibited DPP IV in vitro | 103.5 μM | [106] | |
| EFLLAGNNK | |||||||
| Wheat | Debitrase HYW20 | Gluten | Gluten hydrolysates | VPL WL | Inhibited DPP IV in vitro | [107] | |
| Quinoa | Pepsin and pancreatin | Protein hydrolysates | GEHGSDGNV | Inhibited DPP IV and α-glucosidase in vitro | [109] | ||
| IQAEGGLT | Inhibited DPP IV and α-glucosidase in vitro | ||||||
| DKKYPK | Inhibited α-glucosidase in vitro | ||||||
| Quinoa | Papain, ficin and bromelain | Seed storage proteins | MAF | Inhibited DPPIV in vitro | [110] | ||
| NMF | |||||||
| HPF | |||||||
| MCG | |||||||
| Amaranthus | Alcalase | Albumin and globulin | Protein hydrolysates | Inhibited DPP IV in vitro and in vivo | 0.12 ± 0.01 mg/mL | [111] | |
| Amaranthus | Pepsin and pancreatin | Protein hydrolysates | FLISCLL | Inhibited α-amylase and DPP IV in vitro | [112] | ||
| SVFDEELS | |||||||
| DFIILE | |||||||
| NRPET | |||||||
| HVIKPPS | |||||||
| Walnut | Alcalase, Trypsin, Neutrase, Protamex and Flavourzyme | Protein hydrolysates | LPLLR | Inhibited both α-glucosidase and α-amylase in vitro | [115,116] | ||
| Cumin seeds | Protamex | Seed protein hydrolysates | FFRSKLLSDGAAAAKGALLPQYW | Inhibited porcine pancreatic α-amylase in vitro | [119] | ||
| Basil seeds | Alcalase | Seed protein hydrolysates | ACGNLPRMC | Inhibited α-amylase activity in vitro | [118] | ||
| ACNLPRMC | |||||||
| AGCGCEAMFAGA |
6. Structural Characteristics of Antidiabetic Peptides from Food
7. Oral Bioavailability and Stability of Peptide Drugs
8. Future Prospects of Peptide Drugs
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.-F. Global, regional, and national burden and trend of diabetes in 195 countries and territories: An analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Classification and diagnosis of diabetes: Standards of medical care in diabetes—2020. Diabetes Care 2020, 43, S14–S31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the international diabetes federation diabetes atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bragg, F.; Holmes, M.V.; Iona, A.; Guo, Y.; Du, H.; Chen, Y.; Bian, Z.; Yang, L.; Herrington, W.; Bennett, D.; et al. Association between diabetes and cause-specific mortality in rural and urban areas of China. JAMA 2017, 317, 280–289. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Diabetes Fact Sheet. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 10 June 2021).
- Banu, S.; Bhowmick, A. Therapeutic targets of type 2 diabetes: An overview. MOJ Drug Des. Dev. Ther. 2017, 1, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Padhi, S.; Nayak, A.K.; Behera, A. Type II diabetes mellitus: A review on recent drug based therapeutics. Biomed. Pharmacother. 2020, 131, 110708. [Google Scholar] [CrossRef] [PubMed]
- Daliri, E.; Oh, D.; Lee, B. Bioactive peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Thrasher, J. Pharmacologic management of type 2 diabetes mellitus: Available therapies. Am. J. Med. 2017, 130, S4–S17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylow, L.; Kleinert, M.; Richter, E.A.; Jensen, T.E. Exercise-stimulated glucose uptake—regulation and implications for glycaemic control. Nat. Rev. Endocrinol. 2017, 13, 133–148. [Google Scholar] [CrossRef]
- Forouhi, N.G.; Misra, A.; Mohan, V.; Taylor, R.; Yancy, W. Dietary and nutritional approaches for prevention and management of type 2 diabetes. BMJ 2018, 361, k2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, S.H.; Hamdy, O.; Mohan, V.; Hu, F.B. Prevention and management of type 2 diabetes: Dietary components and nutritional strategies. Lancet 2014, 383, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Iavarone, F.; Desiderio, C.; Vitali, A.; Messana, I.; Martelli, C.; Castagnola, M.; Cabras, T. Cryptides: Latent peptides everywhere. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 246–263. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, P.; Gandía, M.; Garrigues, S.; Marcos, J.F. Improving health-promoting effects of food-derived bioactive peptides through rational design and oral delivery strategies. Nutrients 2019, 11, 2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Aslam, M.Z.; Aslam, M.S.; Firdos, S.; Ghous, G.; Firdos, G.; Hongfei, Z.; Bolin, Z. Role of bioactive peptides in reducing the severity of hypertension with the inhibition of ACE. Int. J. Pept. Res. Ther. 2019, 25, 1639–1649. [Google Scholar] [CrossRef]
- Tyagi, A.; Daliri, E.B.-M.; Kwami Ofosu, F.; Yeon, S.-J.; Oh, D.-H. Food-derived opioid peptides in human health: A review. IJMS 2020, 21, 8825. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhao, J.; Yang, R.; Zhao, W. Bioactive peptides with antidiabetic properties: A review. Int. J. Food Sci. Technol. 2019, 54, 1909–1919. [Google Scholar] [CrossRef] [Green Version]
- Muttenthaler, M.; King, G.F.; Adams, D.J.; Alewood, P.F. Trends in peptide drug discovery. Nat. Rev. Drug Discov. 2021, 20, 309–325. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Antidiabetic food-derived peptides for functional feeding: Production, functionality and In Vivo evidences. Foods 2020, 9, 983. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, T.; Inoue, Y.; Arima, J.; Kumagai, Y.; Usuki, H.; Kawakami, K.; Kimura, M.; Mukaihara, T. Production of dipeptidyl peptidase IV inhibitory peptides from defatted rice bran. Food Chem. 2012, 134, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Lindström, J.; Virtanen, S.M. Functional foods and prevention of diabetes. In Functional Foods; Elsevier: Amsterdam, The Netherlands, 2011; pp. 261–276. ISBN 978-1-84569-690-0. [Google Scholar]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, Z.F.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazcano-Perez, F.; Roman-Gonzalez, S.A.; Sanchez-Puig, N.; Arreguin- Espinosa, R. Bioactive peptides from marine organisms: A short overview. Protein Pept. Lett. 2012, 19, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.P.; Goswami, A.; Kalia, K.; Kate, A.S. Plant-derived bioactive peptides: A treatment to cure diabetes. Int. J. Pept. Res. Ther. 2020, 26, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Chai, K.F.; Voo, A.Y.H.; Chen, W.N. Bioactive peptides from food fermentation: A comprehensive review of their sources, bioactivities, applications, and future development. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3825–3885. [Google Scholar] [CrossRef] [PubMed]
- Raveschot, C.; Cudennec, B.; Coutte, F.; Flahaut, C.; Fremont, M.; Drider, D.; Dhulster, P. Production of bioactive peptides by lactobacillus species: From gene to application. Front. Microbiol. 2018, 9, 2354. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Aronoff, S.L.; Berkowitz, K.; Shreiner, B.; Want, L. Glucose metabolism and regulation: Beyond insulin and glucagon. Diabetes Spectr. 2004, 17, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. IJMS 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Vecchio, I.; Tornali, C.; Bragazzi, N.L.; Martini, M. The discovery of insulin: An important milestone in the history of medicine. Front. Endocrinol. 2018, 9, 613. [Google Scholar] [CrossRef] [Green Version]
- Van de Laar, F.A.; Lucassen, P.L.; Akkermans, R.P.; van de Lisdonk, E.H.; Rutten, G.E.; van Weel, C. Glucosidase inhibitors for patients with type 2 diabetes: Results from a cochrane systematic review and meta-analysis. Diabetes Care 2005, 28, 154–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashary, R.; Vyas, M.; Nayak, S.K.; Suttee, A.; Verma, S.; Narang, R.; Khatik, G.L. An insight of alpha-amylase inhibitors as a valuable tool in the management of type 2 diabetes mellitus. CDR 2020, 16, 117–136. [Google Scholar] [CrossRef] [PubMed]
- Kalra, S.; Bhutani, J. Alpha-glucosidase Inhibitors. In Diabetology: Type 2 Diabetes Mellitus; Jaypee Brothers Medical Publishers (P) Ltd.: New Delhi, India, 2014; p. 55. ISBN 978-93-5152-001-6. [Google Scholar]
- Kawamori, R.; Tajima, N.; Iwamoto, Y.; Kashiwagi, A.; Shimamoto, K.; Kaku, K. Voglibose for prevention of type 2 diabetes mellitus: A randomised, double-blind trial in japanese individuals with impaired glucose tolerance. Lancet 2009, 373, 1607–1614. [Google Scholar] [CrossRef]
- Kazakos, K. Incretin effect: GLP-1, GIP, DPP4. Diabetes Res. Clin. Pract. 2011, 93, S32–S36. [Google Scholar] [CrossRef]
- Gilbert, M.P.; Pratley, R.E. GLP-1 analogs and DPP-4 inhibitors in type 2 diabetes therapy: Review of head-to-head clinical trials. Front. Endocrinol. 2020, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Chao, E.C.; Henry, R.R. SGLT2 inhibition—a novel strategy for diabetes treatment. Nat. Rev. Drug Discov. 2010, 9, 551–559. [Google Scholar] [CrossRef]
- Shorr, R.I.; Ray, W.A.; Daugherty, J.R.; Griffin, M.R. Individual sulfonylureas and serious hypoglycemia in older people. J. Am. Geriatr. Soc. 1996, 44, 751–755. [Google Scholar] [CrossRef]
- Kokil, G.R.; Veedu, R.N.; Ramm, G.A.; Prins, J.B.; Parekh, H.S. Type 2 diabetes mellitus: Limitations of conventional therapies and intervention with nucleic acid-based therapeutics. Chem. Rev. 2015, 115, 4719–4743. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorganic Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef]
- De la Torre, B.G.; Albericio, F. Peptide therapeutics 2.0. Molecules 2020, 25, 2293. [Google Scholar] [CrossRef] [PubMed]
- Furman, B.L. The Development of byetta (Exenatide) from the venom of the gila monster as an anti-diabetic agent. Toxicon 2012, 59, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Wajcberg, E. Amatur amarah liraglutide in the management of type 2 diabetes. Drug Des. Dev. Ther. 2010, 4, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, D.P.; Mohapatra, S.; Misra, S.; Sahu, P.S. Milk derived bioactive peptides and their impact on human health-a review. Saudi J. Biol. Sci. 2015, 23, 577–583. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, R.; Mills, S.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from casein and whey proteins. In Milk and Dairy Products as Functional Foods; Kanekanian, A., Ed.; John Wiley & Sons, Ltd: Chichester, UK, 2014; pp. 23–54. ISBN 978-1-118-63505-6. [Google Scholar]
- Vargas-Bello-Pérez, E.; Márquez-Hernández, R.I.; Hernández-Castellano, L.E. Bioactive Peptides from milk: Animal determinants and their implications in human health. J. Dairy Res. 2019, 86, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Lucarini, M. Bioactive peptides in milk: From encrypted sequences to nutraceutical aspects. Beverages 2017, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, M.I.; Awad, S.; Wahba, A.; El Attar, A.; Yousef, M.I.; Zedan, M. In Vivo anti-diabetic and biological activities of milk protein and milk protein hydrolyaste. Adv. Dairy Res. 2016, 4. [Google Scholar] [CrossRef]
- Li, Y.; Fan, Y.; Shaikh, A.S.; Wang, Z.; Wang, D.; Tan, H. Dezhou donkey (Equus asinus) milk a potential treatment strategy for type 2 diabetes. J. Ethnopharmacol. 2020, 246, 112221. [Google Scholar] [CrossRef]
- Gong, H.; Gao, J.; Wang, Y.; Luo, Q.W.; Guo, K.R.; Ren, F.Z.; Mao, X.Y. Identification of novel peptides from goat milk casein that ameliorate high-glucose-induced insulin resistance in HepG2 cells. J. Dairy Sci. 2020, 103, 4907–4918. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, R.; Ma, H.; Chen, S. Isolation and identification of dipeptidyl peptidase IV-inhibitory peptides from trypsin/chymotrypsin-treated goat milk casein hydrolysates by 2D-TLC and LC–MS/MS. J. Agric. Food Chem. 2015, 63, 8819–8828. [Google Scholar] [CrossRef] [PubMed]
- Mudgil, P.; Kamal, H.; Yuen, G.C.; Maqsood, S. Characterization and identification of novel antidiabetic and anti-obesity peptides from camel milk protein hydrolysates. Food Chem. 2018, 259, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Susceptibility of milk protein-derived peptides to dipeptidyl peptidase IV (DPP-IV) hydrolysis. Food Chem. 2014, 145, 845–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baspinar, B.; Güldaş, M. Traditional plain yogurt: A therapeutic food for metabolic syndrome? Crit. Rev. Food Sci. Nutr. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Hayes, H.E.; Stead, D.; Raikos, V. Incorporating salal berry (Gaultheria Shallon) and blackcurrant (Ribes Nigrum) pomace in yogurt for the development of a beverage with antidiabetic properties. Heliyon 2018, 4, e00875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Yu, Z.; Yin, Y.; Zhao, W.; Yu, Y.; Liu, B.; Liu, J.; Chen, F. Novel peptides derived from egg white protein inhibiting alpha-glucosidase. Food Chem. 2011, 129, 1376–1382. [Google Scholar] [CrossRef]
- Zambrowicz, A.; Pokora, M.; Setner, B.; Dąbrowska, A.; Szołtysik, M.; Babij, K.; Szewczuk, Z.; Trziszka, T.; Lubec, G.; Chrzanowska, J. Multifunctional peptides derived from an egg yolk protein hydrolysate: Isolation and characterization. Amino Acids 2015, 47, 369–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nong, N.T.P.; Chen, Y.-K.; Shih, W.-L.; Hsu, J.-L. Characterization of novel dipeptidyl peptidase-iv inhibitory peptides from soft-shelled turtle yolk hydrolysate using orthogonal bioassay-guided fractionations coupled with In Vitro and In Silico study. Pharmaceuticals 2020, 13, 308. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, D.; Yu, Z.; Ding, L.; Liu, J. Novel membrane peptidase inhibitory peptides with activity against angiotensin converting enzyme and dipeptidyl peptidase iv identified from hen eggs. J. Funct. Foods 2020, 64, 103649. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Comparative peptidomic profile and bioactivities of cooked beef, pork, chicken and turkey meat after In Vitro gastro-intestinal digestion. J. Proteom. 2019, 208, 103500. [Google Scholar] [CrossRef]
- Gallego, M.; Aristoy, M.-C.; Toldrá, F. Dipeptidyl peptidase iv inhibitory peptides generated in spanish dry-cured ham. Meat Sci. 2014, 96, 757–761. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Martí, À.; Bravo, F.I.; Serrano, J.; Ardévol, A.; Pinent, M.; Muguerza, B. Antihyperglycemic effect of a chicken feet hydrolysate via the incretin system: DPP-IV-inhibitory activity and GLP-1 release stimulation. Food Funct. 2019, 10, 4062–4070. [Google Scholar] [CrossRef] [PubMed]
- De Cássia Lemos Lima, R.; Berg, R.S.; Rønning, S.B.; Afseth, N.K.; Knutsen, S.H.; Staerk, D.; Wubshet, S.G. Peptides from chicken processing by-product inhibit DPP-IV and promote cellular glucose uptake: Potential ingredients for T2D management. Food Funct. 2019, 10, 1619–1628. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Oey, I.; Bremer, P.; Carne, A.; Silcock, P. Bioactive peptides derived from egg proteins: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 2508–2530. [Google Scholar] [CrossRef] [PubMed]
- Arena, S.; Renzone, G.; Scaloni, A. A multi-approach peptidomic analysis of hen egg white reveals novel putative bioactive molecules. J. Proteom. 2020, 215, 103646. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Liu, R.; Cao, S.; Zhang, W.; Guanghong, Z. Meat protein based bioactive peptides and their potential functional activity: A review. Int. J. Food Sci. Technol. 2019, 54, 1956–1966. [Google Scholar] [CrossRef] [Green Version]
- Kęska, P.; Stadnik, J.; Bąk, O.; Borowski, P. Meat proteins as dipeptidyl peptidase iv inhibitors and glucose uptake stimulating peptides for the management of a type 2 diabetes mellitus In Silico study. Nutrients 2019, 11, 2537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.-C.; Mora, L. Bioactive peptides generated in the processing of dry-cured ham. Food Chem. 2020, 321, 126689. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sánchez, S.M.; Minguela, A.; Prieto-Merino, D.; Zafrilla-Rentero, M.P.; Abellán-Alemán, J.; Montoro-García, S. The effect of regular intake of dry-cured ham rich in bioactive peptides on inflammation, platelet and monocyte activation markers in humans. Nutrients 2017, 9, 321. [Google Scholar] [CrossRef] [Green Version]
- Mora, L.; González-Rogel, D.; Heres, A.; Toldrá, F. Iberian dry-cured ham as a potential source of α-glucosidase-inhibitory peptides. J. Funct. Foods 2020, 67, 103840. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, R.; Chen, X.; Zeng, Z.; Ma, H.; Chen, S. Dipeptidyl peptidase IV-inhibitory peptides derived from silver carp (Hypophthalmichthys molitrix Val.) proteins. J. Agric. Food Chem. 2016, 64, 831–839. [Google Scholar] [CrossRef]
- Hong, H.; Zheng, Y.; Song, S.; Zhang, Y.; Zhang, C.; Liu, J.; Luo, Y. Identification and characterization of DPP-IV inhibitory peptides from silver carp swim bladder hydrolysates. Food Biosci. 2020, 38, 100748. [Google Scholar] [CrossRef]
- Huang, S.-L.; Jao, C.-L.; Ho, K.-P.; Hsu, K.-C. Dipeptidyl-peptidase IV inhibitory activity of peptides derived from tuna cooking juice hydrolysates. Peptides 2012, 35, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.M.; FitzGerald, R.J. Blue whiting (Micromesistius poutassou) muscle protein hydrolysate with in vitro and in vivo antidiabetic properties. J. Funct. Foods 2018, 40, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.M.; FitzGerald, R.J. Atlantic salmon (Salmo salar) co-product-derived protein hydrolysates: A source of antidiabetic peptides. Food Res. Int. 2018, 106, 598–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, C.; Gleddie, S.; Xiao, C.-W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [Green Version]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Technol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Arnoldi, A. Three peptides from soy glycinin modulate glucose metabolism in human hepatic HepG2 cells. Int. J. Mech. Sci. 2015, 16, 27362–27370. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhao, H.; Pan, X.; Orfila, C.; Lu, W.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of A-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef]
- Lu, J.; Zeng, Y.; Hou, W.; Zhang, S.; Li, L.; Luo, X.; Xi, W.; Chen, Z.; Xiang, M. The soybean peptide aglycin regulates glucose homeostasis in type 2 diabetic mice via IR/IRS1 pathway. J. Nutr. Biochem. 2012, 23, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Feng, J.; Du, Z.; Zhen, H.; Lin, M.; Jia, S.; Li, T.; Huang, X.; Ostenson, C.-G.; Chen, Z. Oral administration of soybean peptide vglycin normalizes fasting glucose and restores impaired pancreatic function in type 2 diabetic wistar rats. J. Nutr. Biochem. 2014, 25, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Tong, Y.; Yan, D.; Jia, S.; Ostenson, C.-G.; Chen, Z. The soybean peptide vglycin preserves the diabetic β-cells through improvement of proliferation and inhibition of apoptosis. Sci. Rep. 2015, 5, 15599. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Muraki, A.; Oie, M.; Kanegawa, N.; Oda, A.; Sawashi, Y.; Kaneko, K.; Yoshikawa, M.; Goto, T.; Takahashi, N.; et al. Soymorphin-5, a soy-derived μ-opioid peptide, decreases glucose and triglyceride levels through activating adiponectin and PPARα systems in diabetic KKAy mice. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E433–E440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Bioactive peptides from germinated soybean with anti-diabetic potential by inhibition of dipeptidyl peptidase-IV, α-amylase, and α-glucosidase enzymes. Int. J. Mol. Sci. 2018, 19, 2883. [Google Scholar] [CrossRef] [Green Version]
- Lammi, C.; Bollati, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Soybean- and lupin-derived peptides inhibit dpp-iv activity on in situ human intestinal caco-2 cells and Ex vivo human serum. Nutrients 2018, 10, 1082. [Google Scholar] [CrossRef] [Green Version]
- Lammi, C.; Arnoldi, A.; Aiello, G. Soybean peptides exert multifunctional bioactivity modulating 3-hydroxy-3-methylglutaryl-coa reductase and dipeptidyl peptidase-IV targets in vitro. J. Agric. Food Chem. 2019, 67, 4824–4830. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Peñas, E.; Frias, J. Bioactive peptides in fermented foods. In Fermented Foods in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2017; pp. 23–47. ISBN 978-0-12-802309-9. [Google Scholar]
- Kwon, D.Y.; Daily, J.W.; Kim, H.J.; Park, S. Antidiabetic effects of fermented soybean products on type 2 diabetes. Nutr. Res. 2010, 30, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tiss, M.; Souiy, Z.; ben Abdeljelil, N.; Njima, M.; Achour, L.; Hamden, K. Fermented soy milk prepared using kefir grains prevents and ameliorates obesity, type 2 diabetes, hyperlipidemia and liver-kidney toxicities in HFFD-rats. J. Funct. Foods 2020, 67, 103869. [Google Scholar] [CrossRef]
- Dong, J.-Y.; Kimura, T.; Ikehara, S.; Cui, M.; Kawanishi, Y.; Ueda, K.; Iso, H.; Yamazaki, S.; Ohya, Y.; Kishi, R.; et al. Soy consumption and incidence of gestational diabetes mellitus: The japan environment and children’s study. Eur. J. Nutr. 2020, 60, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Miyasaka, S.; Tsuji, A.; Tachi, H. Isolation and characterization of peptides with dipeptidyl peptidase IV (DPPIV) inhibitory activity from natto using DPPIV from aspergillus oryzae. Food Chem. 2018, 261, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Mojica, L.; de Mejía, E.G. Optimization of enzymatic production of anti-diabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef]
- Ngoh, Y.-Y.; Gan, C.-Y. Enzyme-assisted extraction and identification of antioxidative and α-amylase inhibitory peptides from pinto beans (Phaseolus vulgaris Cv. Pinto). Food Chem 2016, 190, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.-Y.; Tye, G.J.; Gan, C.-Y. The investigation of α-amylase inhibitory activity of selected pinto bean peptides via preclinical study using AR42J cell. J. Funct. Foods 2017, 35, 641–647. [Google Scholar] [CrossRef]
- Valencia-Mejía, E.; Batista, K.A.; Fernández, J.J.A.; Fernandes, K.F. Antihyperglycemic and hypoglycemic activity of naturally occurring peptides and protein hydrolysates from easy-to-cook and hard-to-cook beans (Phaseolus vulgaris L.). Food Res. Int. 2019, 121, 238–246. [Google Scholar] [CrossRef]
- Mune Mune, M.A.; Minka, S.R.; Henle, T. Investigation on antioxidant, angiotensin converting enzyme and dipeptidyl peptidase iv inhibitory activity of bambara bean protein hydrolysates. Food Chem. 2018, 250, 162–169. [Google Scholar] [CrossRef]
- Uruakp, F. Influence of cowpea (Vigna unguiculata) peptides on insulin resistance. J. Nutr. Heal. Food Sci. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, S.; Luna-Vital, D.; de Mejia, E.G. Identification and comparison of peptides from chickpea protein hydrolysates using either bromelain or gastrointestinal enzymes and their relationship with markers of type 2 diabetes and bitterness. Nutrients 2020, 12, 3843. [Google Scholar] [CrossRef]
- Cavazos, A.; Gonzalez de Mejia, E. Identification of bioactive peptides from cereal storage proteins and their potential role in prevention of chronic diseases. Compr. Rev. Food Sci. Food Saf. 2013, 12, 364–380. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, G.; Mu, H.; Zhang, H.; Chen, Y.Q. The mixture of corn and wheat peptide prevent diabetes in NOD mice. J. Funct. Foods 2019, 56, 163–170. [Google Scholar] [CrossRef]
- Wang, F.; Yu, G.; Zhang, Y.; Zhang, B.; Fan, J. Dipeptidyl peptidase IV inhibitory peptides derived from oat (Avena Sativa L.), buckwheat (Fagopyrum Esculentum), and highland barley (Hordeum Vulgare Trifurcatum (L.) trofim) proteins. J. Agric. Food Chem. 2015, 63, 9543–9549. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Hennemann, M.; Paolella, S.; FitzGerald, R.J. Generation of wheat gluten hydrolysates with dipeptidyl peptidase IV (DPP-IV) Inhibitory properties. Food Funct. 2017, 8, 2249–2257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudgil, P.; Kilari, B.P.; Kamal, H.; Olalere, O.A.; FitzGerald, R.J.; Gan, C.-Y.; Maqsood, S. Multifunctional bioactive peptides derived from quinoa protein hydrolysates: Inhibition of α-glucosidase, dipeptidyl peptidase-IV and angiotensin I converting enzymes. J. Cereal Sci. 2020, 96, 103130. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of Dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Richel, A.; Hao, Y.; Fan, X.; Everaert, N.; Yang, X.; Ren, G. Novel dipeptidyl peptidase-IV and angiotensin-I-converting enzyme inhibitory peptides released from quinoa protein by in silico proteolysis. Food Sci. Nutr. 2020, 8, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorge, S.-S.; Raúl, R.-B.; Isabel, G.-L.; Edith, P.-A.; Bernardo, E.-B.; César, A.-P.; Gerardo, D.-G.; Rubén, R.-R. Dipeptidyl peptidase IV inhibitory activity of protein hydrolyzates from Amaranthus Hypochondriacus L. grain and their influence on postprandial glycemia in streptozotocin-induced diabetic mice. Afr. J. Trad. Compl. Alt. Med. 2015, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Miralles, B.; Hernández-Ledesma, B. Release of multifunctional peptides from kiwicha (Amaranthus caudatus) protein under in vitro gastrointestinal digestion: Multifunctional kiwicha-derived peptides. J. Sci. Food Agric. 2019, 99, 1225–1232. [Google Scholar] [CrossRef] [Green Version]
- Ros, E.; Hu, F.B. Consumption of plant seeds and cardiovascular health: Epidemiological and clinical trial evidence. Circulation 2013, 128, 553–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendall, C.W.C.; Esfahani, A.; Truan, J.; Srichaikul, K.; Jenkins, D.J.A. Health benefits of nuts in prevention and management of diabetes. Asia Pac. J. Clin. Nutr. 2010, 19, 110–116. [Google Scholar] [PubMed]
- Wang, J.; Du, K.; Fang, L.; Liu, C.; Min, W.; Liu, J. Evaluation of the antidiabetic activity of hydrolyzed peptides derived from Juglans mandshurica maxim. Fruits in insulin-resistant HepG2 cells and type 2 diabetic mice. J. Food Biochem. 2018, 42, e12518. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, L.; Liu, C.; Liu, X.; Li, H.; Shi, J.; Li, M.; Min, W. Anti-diabetic effect by walnut (Juglans mandshurica Maxim.)-derived peptide LPLLR through inhibiting α-glucosidase and α-amylase, and alleviating insulin resistance of hepatic HepG2 cells. J. Funct. Foods 2020, 69, 103944. [Google Scholar] [CrossRef]
- Doyen, A.; Udenigwe, C.C.; Mitchell, P.L.; Marette, A.; Aluko, R.E.; Bazinet, L. Anti-diabetic and antihypertensive activities of two flaxseed protein hydrolysate fractions revealed following their simultaneous separation by electrodialysis with ultrafiltration membranes. Food Chem. 2014, 145, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Nurul Hidayatul Afifah, B.S.S.; Gan, C.-Y. Antioxidative and amylase inhibitor peptides from basil seeds. Int. J. Pept. Res. Ther. 2016, 22, 3–10. [Google Scholar] [CrossRef]
- Siow, H.-L.; Gan, C.-Y. Extraction, Identification, and structure–activity relationship of antioxidative and α-amylase inhibitory peptides from cumin seeds (Cuminum cyminum). J. Funct. Foods 2016, 22, 1–12. [Google Scholar] [CrossRef]
- Han, R.; Maycock, J.; Murray, B.S.; Boesch, C. Identification of angiotensin converting enzyme and dipeptidyl peptidase-IV inhibitory peptides derived from oilseed proteins using two integrated bioinformatic approaches. Food Res. Int. 2019, 115, 283–291. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Features of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from dietary proteins. J. Food Biochem. 2019, 43, e12451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arulmozhiraja, S.; Matsuo, N.; Ishitsubo, E.; Okazaki, S.; Shimano, H.; Tokiwa, H. Comparative binding analysis of dipeptidyl peptidase IV (DPP-4) with antidiabetic drugs–an Ab initio fragment molecular orbital study. PLoS ONE 2016, 11, e0166275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.H.; Gaspar, A.R.M. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2018, 91, 370–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otvos, L.; Wade, J.D. Current challenges in peptide-based drug discovery. Front. Chem. 2014, 2, 62. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.D.; Whitehead, K.A.; Mitragotri, S. Materials for oral delivery of proteins and peptides. Nat. Rev. Mater. 2020, 5, 127–148. [Google Scholar] [CrossRef]
- Ganesh, A.N.; Heusser, C.; Garad, S.; Sánchez-Félix, M.V. Patient-centric design for peptide delivery: Trends in routes of administration and advancement in drug delivery technologies. Med. Drug Discov. 2021, 9, 100079. [Google Scholar] [CrossRef]
- Ibrahim, Y.H.-E.Y.; Regdon, G.; Hamedelniel, E.I.; Sovány, T. Review of recently used techniques and materials to improve the efficiency of orally administered proteins/peptides. DARU J. Pharm. Sci. 2020, 28, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zizzari, A.T.; Pliatsika, D.; Gall, F.M.; Fischer, T.; Riedl, R. New perspectives in oral peptide delivery. Drug Discov. Today 2021, 26, 1097–1105. [Google Scholar] [CrossRef]
- Zhu, Q.; Chen, Z.; Paul, P.K.; Lu, Y.; Wu, W.; Qi, J. Oral delivery of proteins and peptides: Challenges, status quo and future perspectives. Acta Pharm. Sin. B 2021. [Google Scholar] [CrossRef]
- Pudlarz, A.; Szemraj, J. Nanoparticles as carriers of proteins, peptides and other therapeutic molecules. Open Life Sci. 2018, 13, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Rachmiel, M.; Barash, G.; Leshem, A.; Sagi, R.; Doenyas-Barak, K.; Koren, S. OR14-1 Pharmacodynamics, safety, tolerability, and efficacy of oral insulin formulation (Oshadi Icp) among young adults with type 1 diabetes: A summary of clinical studies phases I, Ib, and Ii. J. Endocr. Soc. 2019, 3, OR14-1. [Google Scholar] [CrossRef]
- Redko, B.; Tuchinsky, H.; Segal, T.; Tobi, D.; Luboshits, G.; Ashur-Fabian, O.; Pinhasov, A.; Gerlitz, G.; Gellerman, G. Toward the development of a novel Non-RGD cyclic peptide drug conjugate for treatment of human metastatic melanoma. Oncotarget 2017, 8, 757–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alas, M.; Saghaeidehkordi, A.; Kaur, K. Peptide–drug conjugates with different linkers for cancer therapy. J. Med. Chem. 2021, 64, 216–232. [Google Scholar] [CrossRef]
- Khedkar, A.; Lebovitz, H.; Fleming, A.; Cherrington, A.; Jose, V.; Athalye, S.N.; Vishweswaramurthy, A. Impact of insulin tregopil and its permeation enhancer on pharmacokinetics of metformin in healthy volunteers: Randomized, open-label, placebo-controlled, crossover study. Clin. Transl. Sci. 2019, 12, 276–282. [Google Scholar] [CrossRef]
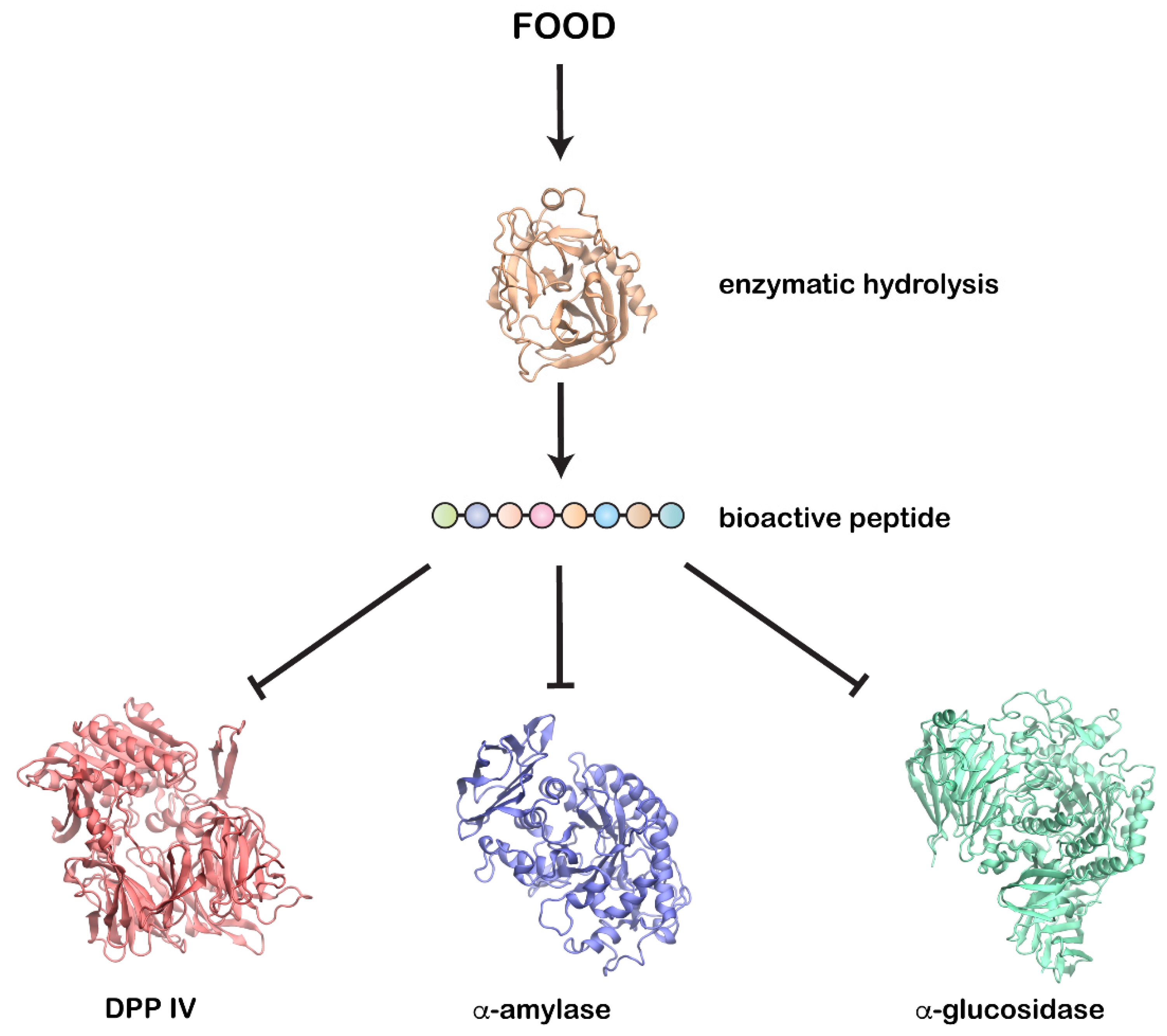
| Source | Enzyme(s) Used | Substrate | Proteins Hydrolysates | Peptide(s) Identified | Mechanism of Action | IC50 | Reference |
|---|---|---|---|---|---|---|---|
| Silver carp | Trypsin, neutrase, alcalase, papain, pepsin, and flavourzyme | Dorsal muscle | Muscle protein hydrolysates | LPIIDI | Inhibited DPP IV in vitro | 105.44 μM | [76] |
| APGPAGP | 229.14 μM | ||||||
| AGPPGPSG | Nd | ||||||
| ALAPSTM | Nd | ||||||
| Silver carp | Papain, bromelain, alcalase 2.4 L, neutrase, and flavourzyme | Swim bladder protein | Protein hydrolysates | WGDEHIPGSPYH | Inhibited DPP IV in vitro and improved glucose uptake in INS cells | 0.35 ± 0.01 mM | [77] |
| IAQPQEKAPDPFRH | 0.81 ± 0.01 mM | ||||||
| IAGPAGPRGPAGPN | 1.17 ± 0.04 mM | ||||||
| VAPEEHPTL | 1.93 ± 0.02 mM | ||||||
| YALPHAI | 2.27 ± 0.04 mM | ||||||
| EPGNPGPAGPA | 2.94 ± 0.01 mM | ||||||
| Tuna | Protease XXIII, orientase 90N | Cooking juice | Cooking juice hydrolysates | PGVGGPLGPIGPCYE | Inhibited DPP IV in vitro | 116.1 μM | [78] |
| CAYQWQRPVDRIR | 78 μM | ||||||
| PACGGFWISGRPG | 96.4 μM | ||||||
| Blue whiting | Alcalase and flavourzyme 500 | Fish meat | Meat protein hydrolysates | Inhibited DPP IV and mediated insulin and GLP-1 release from BRIN-BD11 and GLUTag cells | [79] | ||
| Salmon | Alcalase and Flavourzyme | Skin and trimmings | Protein hydrolysates | Inhibited DPP IV and mediated insulin and GLP-1 release from BRIN-BD11 and GLUTag cells | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antony, P.; Vijayan, R. Bioactive Peptides as Potential Nutraceuticals for Diabetes Therapy: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 9059. https://doi.org/10.3390/ijms22169059
Antony P, Vijayan R. Bioactive Peptides as Potential Nutraceuticals for Diabetes Therapy: A Comprehensive Review. International Journal of Molecular Sciences. 2021; 22(16):9059. https://doi.org/10.3390/ijms22169059
Chicago/Turabian StyleAntony, Priya, and Ranjit Vijayan. 2021. "Bioactive Peptides as Potential Nutraceuticals for Diabetes Therapy: A Comprehensive Review" International Journal of Molecular Sciences 22, no. 16: 9059. https://doi.org/10.3390/ijms22169059
APA StyleAntony, P., & Vijayan, R. (2021). Bioactive Peptides as Potential Nutraceuticals for Diabetes Therapy: A Comprehensive Review. International Journal of Molecular Sciences, 22(16), 9059. https://doi.org/10.3390/ijms22169059