
The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment


Abstract
1. Introduction
2. CB2R Expression and Inducible Feature
3. Drug Resistance in Epilepsy
4. Cannabinoid System as a Potential Therapeutic Target for Treating Epilepsy
4.1. Endocannabinoid System
4.2. Cannabinoids’ Effects on Epilepsy
4.3. CB2R Effects on Preclinical Epilepsy
4.4. CB2R Mediated Anti-Epileptic Effects through a Reduction of Neural Excitability and Synchronization
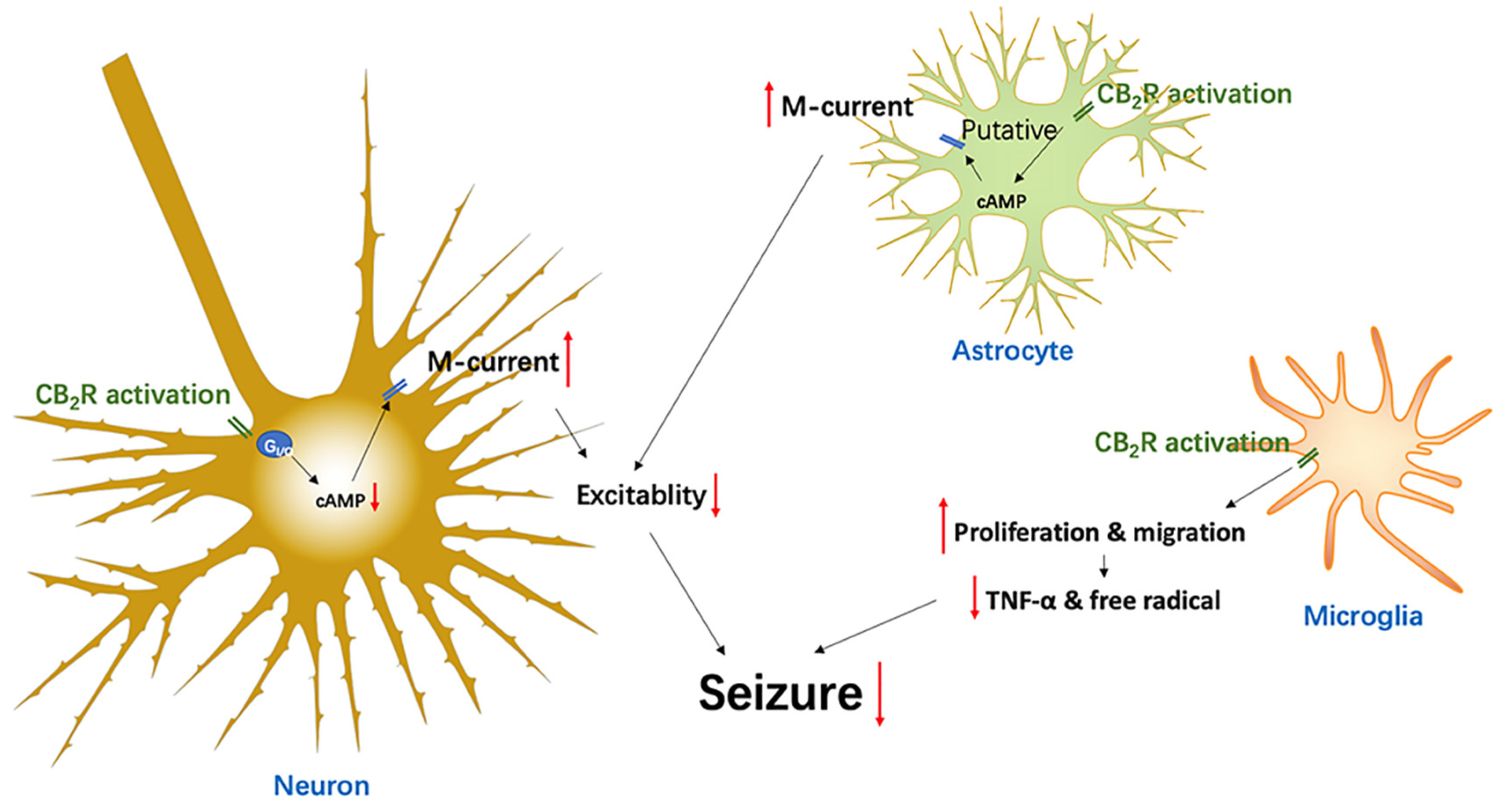
4.5. Glia CB2R-Mediated Anti-Epileptic Effects via Inflammation and Excitability
5. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Espinosa-Jovel, C.; Toledano, R.; Aledo-Serrano, A.; Garcia-Morales, I.; Gil-Nagel, A. Epidemiological profile of epilepsy in low income populations. Seizure 2018, 56, 67–72. [Google Scholar] [CrossRef]
- Rezaei, M.; Sadeghian, A.; Roohi, N.; Shojaei, A.; Mirnajafi-Zadeh, J. Epilepsy and dopaminergic system. Physiol. Pharmacol. 2017, 21, 1–14. [Google Scholar]
- Devinsky, O.; Vezzani, A.; O’Brien, T.J.; Jette, N.; Scheffer, I.E.; de Curtis, M.; Perucca, P. Epilepsy. Nat. Rev. Dis. Primers 2018, 4, 18024. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory pathways as treatment targets and biomarkers in epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef]
- Wallace, M.J.; Blair, R.E.; Falenski, K.W.; Martin, B.R.; DeLorenzo, R.J. The endogenous cannabinoid system regulates seizure frequency and duration in a model of temporal lobe epilepsy. J. Pharm. Exp. 2003, 307, 129–137. [Google Scholar] [CrossRef]
- Deshpande, L.S.; Sombati, S.; Blair, R.E.; Carter, D.S.; Martin, B.R.; DeLorenzo, R.J. Cannabinoid CB1 receptor antagonists cause status epilepticus-like activity in the hippocampal neuronal culture model of acquired epilepsy. Neurosci. Lett. 2007, 411, 11–16. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kozan, R.; Ayyildiz, M.; Agar, E. The effects of intracerebroventricular AM-251, a CB1-receptor antagonist, and ACEA, a CB1-receptor agonist, on penicillin-induced epileptiform activity in rats. Epilepsia 2009, 50, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Cakil, D.; Yildirim, M.; Ayyildiz, M.; Agar, E. The effect of co-administration of the NMDA blocker with agonist and antagonist of CB1-receptor on penicillin-induced epileptiform activity in rats. Epilepsy Res. 2011, 93, 128–137. [Google Scholar] [CrossRef]
- Capasso, A. Do Cannabinoids Confer Neuroprotection Against Epilepsy? An Overview. Open Neurol. J. 2017, 11, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Neale, M. Efficacy and safety of cannabis for treating children with refractory epilepsy. Nurs. Child Young People 2017, 29, 32–37. [Google Scholar] [CrossRef]
- De Caro, C.; Leo, A.; Citraro, R.; De Sarro, C.; Russo, R.; Calignano, A.; Russo, E. The potential role of cannabinoids in epilepsy treatment. Expert Rev. Neurother. 2017, 17, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, Y.; Yamazaki, M.; Uchigashima, M.; Kobayashi, K.; Watanabe, M.; Sakimura, K.; Kano, M. Crucial Roles of the Endocannabinoid 2-Arachidonoylglycerol in the Suppression of Epileptic Seizures. Cell Rep. 2016, 16, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Bie, B.; Wu, J.; Foss, J.F.; Naguib, M. An overview of the cannabinoid type 2 receptor system and its therapeutic potential. Curr. Opin. Anaesthesiol. 2018, 31, 407–414. [Google Scholar] [CrossRef]
- Qian, W.J.; Yin, N.; Gao, F.; Miao, Y.; Li, Q.; Li, F.; Sun, X.H.; Yang, X.L.; Wang, Z. Cannabinoid CB1 and CB2 receptors differentially modulate L- and T-type Ca(2+) channels in rat retinal ganglion cells. Neuropharmacology 2017, 124, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, C.; Sun, T. The Roles of Intracellular Chaperone Proteins, Sigma Receptors, in Parkinson’s Disease (PD) and Major Depressive Disorder (MDD). Front. Pharm. 2019, 10, 528. [Google Scholar] [CrossRef]
- Kreitzer, A.C.; Regehr, W.G. Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto Purkinje cells. Neuron 2001, 29, 717–727. [Google Scholar] [CrossRef]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef]
- Wilson, R.I.; Kunos, G.; Nicoll, R.A. Presynaptic specificity of endocannabinoid signaling in the hippocampus. Neuron 2001, 31, 453–462. [Google Scholar] [CrossRef]
- Devinsky, O.; Patel, A.D.; Cross, J.H.; Villanueva, V.; Wirrell, E.C.; Privitera, M.; Greenwood, S.M.; Roberts, C.; Checketts, D.; VanLandingham, K.E.; et al. Effect of Cannabidiol on Drop Seizures in the Lennox-Gastaut Syndrome. N. Engl. J. Med. 2018, 378, 1888–1897. [Google Scholar] [CrossRef]
- Arslan, G.; Ayyildiz, M.; Agar, E. The interaction between ghrelin and cannabinoid systems in penicillin-induced epileptiform activity in rats. Neuropeptides 2014, 48, 345–352. [Google Scholar] [CrossRef]
- Devinsky, O.; Marsh, E.; Friedman, D.; Thiele, E.; Laux, L.; Sullivan, J.; Miller, I.; Flamini, R.; Wilfong, A.; Filloux, F.; et al. Cannabidiol in patients with treatment-resistant epilepsy: An open-label interventional trial. Lancet Neurol. 2016, 15, 270–278. [Google Scholar] [CrossRef]
- Devinsky, O.; Cross, J.H.; Wright, S. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. N. Engl. J. Med. 2017, 377, 699–700. [Google Scholar] [CrossRef]
- Szaflarski, M.; Hansen, B.; Bebin, E.M.; Szaflarski, J.P. Social correlates of health status, quality of life, and mood states in patients treated with cannabidiol for epilepsy. Epilepsy Behav. 2017, 70, 364–369. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Schatz, A.R.; Lee, M.; Condie, R.B.; Pulaski, J.T.; Kaminski, N.E. Cannabinoid receptors CB1 and CB2: A characterization of expression and adenylate cyclase modulation within the immune system. Toxicol. Appl. Pharm. 1997, 142, 278–287. [Google Scholar] [CrossRef]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef]
- Griffin, G.; Wray, E.J.; Tao, Q.; McAllister, S.D.; Rorrer, W.K.; Aung, M.M.; Martin, B.R.; Abood, M.E. Evaluation of the cannabinoid CB2 receptor-selective antagonist, SR144528: Further evidence for cannabinoid CB2 receptor absence in the rat central nervous system. Eur. J. Pharm. 1999, 377, 117–125. [Google Scholar] [CrossRef]
- Stella, N. Cannabinoid and cannabinoid-like receptors in microglia, astrocytes, and astrocytomas. Glia 2010, 58, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Buriani, A.; Dal Toso, R.; Petrelli, L.; Romanello, S.; Facci, L.; Leon, A. The ALIAmide palmitoylethanolamide and cannabinoids, but not anandamide, are protective in a delayed postglutamate paradigm of excitotoxic death in cerebellar granule neurons. Proc. Natl. Acad. Sci. USA 1996, 93, 3984–3989. [Google Scholar] [CrossRef]
- Lanciego, J.L.; Barroso-Chinea, P.; Rico, A.J.; Conte-Perales, L.; Callen, L.; Roda, E.; Gomez-Bautista, V.; Lopez, I.P.; Lluis, C.; Labandeira-Garcia, J.L.; et al. Expression of the mRNA coding the cannabinoid receptor 2 in the pallidal complex of Macaca fascicularis. J. Psychopharmacol. 2011, 25, 97–104. [Google Scholar] [CrossRef]
- Liu, Q.R.; Pan, C.H.; Hishimoto, A.; Li, C.Y.; Xi, Z.X.; Llorente-Berzal, A.; Viveros, M.P.; Ishiguro, H.; Arinami, T.; Onaivi, E.S.; et al. Species differences in cannabinoid receptor 2 (CNR2 gene): Identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav. 2009, 8, 519–530. [Google Scholar] [CrossRef]
- Lu, Q.; Straiker, A.; Lu, Q.; Maguire, G. Expression of CB2 cannabinoid receptor mRNA in adult rat retina. Vis. Neurosci. 2000, 17, 91–95. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, M.S.; Garcia-Bueno, B.; Zoppi, S.; Leza, J.C.; Manzanares, J. Chronic blockade of cannabinoid CB(2) receptors induces anxiolytic-like actions associated to alterations in GABA(A) receptors. Br. J. Pharm. 2011, 165, 951–964. [Google Scholar] [CrossRef]
- Navarrete, F.; Perez-Ortiz, J.M.; Manzanares, J. Cannabinoid CB(2) receptor-mediated regulation of impulsive-like behaviour in DBA/2 mice. Br. J. Pharm. 2012, 165, 260–273. [Google Scholar] [CrossRef]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 2005, 310, 329–332. [Google Scholar] [CrossRef]
- Viscomi, M.T.; Oddi, S.; Latini, L.; Pasquariello, N.; Florenzano, F.; Bernardi, G.; Molinari, M.; Maccarrone, M. Selective CB2 receptor agonism protects central neurons from remote axotomy-induced apoptosis through the PI3K/Akt pathway. J. Neurosci. 2009, 29, 4564–4570. [Google Scholar] [CrossRef]
- Sherwood, T.A.; Nong, L.; Agudelo, M.; Newton, C.; Widen, R.; Klein, T.W. Identification of transcription start sites and preferential expression of select CB2 transcripts in mouse and human B lymphocytes. J. Neuroimmune Pharm. 2009, 4, 476–488. [Google Scholar] [CrossRef]
- Canseco-Alba, A.; Schanz, N.; Sanabria, B.; Zhao, J.; Lin, Z.; Liu, Q.R.; Onaivi, E.S. Behavioral effects of psychostimulants in mutant mice with cell-type specific deletion of CB2 cannabinoid receptors in dopamine neurons. Behav. Brain Res. 2019, 360, 286–297. [Google Scholar] [CrossRef]
- Agudo, J.; Martin, M.; Roca, C.; Molas, M.; Bura, A.S.; Zimmer, A.; Bosch, F.; Maldonado, R. Deficiency of CB2 cannabinoid receptor in mice improves insulin sensitivity but increases food intake and obesity with age. Diabetologia 2010, 53, 2629–2640. [Google Scholar] [CrossRef]
- Ignatowska-Jankowska, B.; Jankowski, M.M.; Swiergiel, A.H. Cannabidiol decreases body weight gain in rats: Involvement of CB2 receptors. Neurosci. Lett. 2011, 490, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Emadi, L.; Jonaidi, H.; Hosseini Amir Abad, E. The role of central CB2 cannabinoid receptors on food intake in neonatal chicks. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2011, 197, 1143–1147. [Google Scholar] [CrossRef]
- Flake, N.M.; Zweifel, L.S. Behavioral effects of pulp exposure in mice lacking cannabinoid receptor 2. J. Endod. 2012, 38, 86–90. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, M.S.; Manzanares, J. Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 2011, 25, 111–120. [Google Scholar] [CrossRef]
- Hwang, E.S.; Kim, H.B.; Lee, S.; Kim, M.J.; Kim, K.J.; Han, G.; Han, S.Y.; Lee, E.A.; Yoon, J.H.; Kim, D.O.; et al. Antidepressant-like effects of beta-caryophyllene on restraint plus stress-induced depression. Behav. Brain Res. 2020, 380, 112439. [Google Scholar] [CrossRef]
- Ortega-Alvaro, A.; Aracil-Fernandez, A.; Garcia-Gutierrez, M.S.; Navarrete, F.; Manzanares, J. Deletion of CB2 cannabinoid receptor induces schizophrenia-related behaviors in mice. Neuropsychopharmacology 2011, 36, 1489–1504. [Google Scholar] [CrossRef]
- Xi, Z.X.; Peng, X.Q.; Li, X.; Song, R.; Zhang, H.Y.; Liu, Q.R.; Yang, H.J.; Bi, G.H.; Li, J.; Gardner, E.L. Brain cannabinoid CB(2) receptors modulate cocaine’s actions in mice. Nat. Neurosci. 2011, 14, 1160–1166. [Google Scholar] [CrossRef]
- Navarrete, F.; Rodriguez-Arias, M.; Martin-Garcia, E.; Navarro, D.; Garcia-Gutierrez, M.S.; Aguilar, M.A.; Aracil-Fernandez, A.; Berbel, P.; Minarro, J.; Maldonado, R.; et al. Role of CB2 Cannabinoid Receptors in the Rewarding, Reinforcing, and Physical Effects of Nicotine. Neuropsychopharmacology 2013, 38, 2515–2524. [Google Scholar] [CrossRef]
- Ortega-Alvaro, A.; Ternianov, A.; Aracil-Fernandez, A.; Navarrete, F.; Garcia-Gutierrez, M.S.; Manzanares, J. Role of cannabinoid CB receptor in the reinforcing actions of ethanol. Addict. Biol. 2013, 20, 43–55. [Google Scholar] [CrossRef]
- Miller, L.K.; Devi, L.A. The highs and lows of cannabinoid receptor expression in disease: Mechanisms and their therapeutic implications. Pharm. Rev. 2011, 63, 461–470. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Functional expression of brain neuronal CB2 cannabinoid receptors are involved in the effects of drugs of abuse and in depression. Ann. N. Y. Acad. Sci. 2008, 1139, 434–449. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gu, S.; Liu, Q.R. CNS effects of CB2 cannabinoid receptors: Beyond neuro-immuno-cannabinoid activity. J. Psychopharmacol. 2012, 26, 92–103. [Google Scholar] [CrossRef]
- Spyridakos, D.; Papadogkonaki, S.; Dionysopoulou, S.; Mastrodimou, N.; Polioudaki, H.; Thermos, K. Effect of acute and subchronic administration of (R)-WIN55,212-2 induced neuroprotection and anti inflammatory actions in rat retina: CB1 and CB2 receptor involvement. Neurochem. Int. 2021, 142, 104907. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Gao, F.; Larsen, B.; Gao, M.; Luo, Z.; Chen, D.; Ma, X.; Qiu, S.; Zhou, Y.; Xie, J.; et al. Mechanisms of cannabinoid CB2 receptor-mediated reduction of dopamine neuronal excitability in mouse ventral tegmental area. EBioMedicine 2019, 42, 225–237. [Google Scholar] [CrossRef]
- Loscher, W.; Potschka, H.; Sisodiya, S.M.; Vezzani, A. Drug Resistance in Epilepsy: Clinical Impact, Potential Mechanisms, and New Innovative Treatment Options. Pharm. Rev. 2020, 72, 606–638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, X.; Peng, A.; Lai, W.; He, S.; Qiu, X.; Zou, X.; Chen, L. Predictors of drug-resistance in epilepsy with auditory features. Epilepsy Res. 2020, 164, 106353. [Google Scholar] [CrossRef]
- Kolker, S. Metabolism of amino acid neurotransmitters: The synaptic disorder underlying inherited metabolic diseases. J. Inherit. Metab. Dis. 2018, 41, 1055–1063. [Google Scholar] [CrossRef]
- Epi25 Collaborative. Ultra-Rare Genetic Variation in the Epilepsies: A Whole-Exome Sequencing Study of 17,606 Individuals. Am. J. Hum. Genet. 2019, 105, 267–282. [Google Scholar] [CrossRef]
- Iannone, L.F.; Preda, A.; Blottiere, H.M.; Clarke, G.; Albani, D.; Belcastro, V.; Carotenuto, M.; Cattaneo, A.; Citraro, R.; Ferraris, C.; et al. Microbiota-gut brain axis involvement in neuropsychiatric disorders. Expert Rev. Neurother. 2019, 19, 1037–1050. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 174, 497. [Google Scholar] [CrossRef]
- Li, Y.; Tymchuk, S.; Barry, J.; Muppidi, S.; Le, S. Antibody Prevalence in Epilepsy before Surgery (APES) in drug-resistant focal epilepsy. Epilepsia 2021, 62, 720–728. [Google Scholar] [CrossRef]
- Walton, D.; Spencer, D.C.; Nevitt, S.J.; Michael, B.D. Transcranial magnetic stimulation for the treatment of epilepsy. Cochrane Database Syst. Rev. 2021, 4, CD011025. [Google Scholar]
- Fox, K.C.R.; Shi, L.; Baek, S.; Raccah, O.; Foster, B.L.; Saha, S.; Margulies, D.S.; Kucyi, A.; Parvizi, J. Intrinsic network architecture predicts the effects elicited by intracranial electrical stimulation of the human brain. Nat. Hum. Behav. 2020, 4, 1039–1052. [Google Scholar] [CrossRef]
- Lim, Z.; Wong, K.; Downs, J.; Bebbington, K.; Demarest, S.; Leonard, H. Vagus nerve stimulation for the treatment of refractory epilepsy in the CDKL5 Deficiency Disorder. Epilepsy Res. 2018, 146, 36–40. [Google Scholar] [CrossRef]
- Giordano, C.; Marchio, M.; Timofeeva, E.; Biagini, G. Neuroactive peptides as putative mediators of antiepileptic ketogenic diets. Front. Neurol. 2014, 5, 63. [Google Scholar] [CrossRef]
- Schonhofen, P.; Bristot, I.J.; Crippa, J.A.; Hallak, J.E.C.; Zuardi, A.W.; Parsons, R.B.; Klamt, F. Cannabinoid-Based Therapies and Brain Development: Potential Harmful Effect of Early Modulation of the Endocannabinoid System. CNS Drugs 2018, 32, 697–712. [Google Scholar] [CrossRef]
- Senn, L.; Cannazza, G.; Biagini, G. Receptors and Channels Possibly Mediating the Effects of Phytocannabinoids on Seizures and Epilepsy. Pharmaceuticals 2020, 13, 174. [Google Scholar] [CrossRef]
- Kaplan, J.S.; Stella, N.; Catterall, W.A.; Westenbroek, R.E. Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, 11229–11234. [Google Scholar] [CrossRef]
- Kwan Cheung, K.A.; Peiris, H.; Wallace, G.; Holland, O.J.; Mitchell, M.D. The interplay between the endocannabinoid system, epilepsy and cannabinoids. Int. J. Mol. Sci. 2019, 20, 6079. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, J.; Huang, S.; Zhu, W.; Wang, Y.; Chen, O.; Xue, J. Neuroprotective effects of isoliquiritigenin against cognitive impairment via suppression of synaptic dysfunction, neuronal injury, and neuroinflammation in rats with kainic acid-induced seizures. Int. Immunopharmacol. 2019, 72, 358–366. [Google Scholar] [CrossRef]
- Haj-Mirzaian, A.; Ramezanzadeh, K.; Afshari, K.; Mousapour, P.; Abbasi, N.; Haj-Mirzaian, A.; Nikbakhsh, R.; Haddadi, N.-S.; Dehpour, A.R. Activation of ATP-sensitive K-channel promotes the anticonvulsant properties of cannabinoid receptor agonist through mitochondrial ATP level reduction. Epilepsy Behav. 2019, 93, 1–6. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, Z.; Jin, B.; Qin, X.; Zhu, X.; Sun, J.; Huo, L.; Wang, R.; Shi, Y.; Jia, Z. Cannabidiol inhibits febrile seizure by modulating AMPA receptor kinetics through its interaction with the N-terminal domain of GluA1/GluA2. Pharmacol. Res. 2020, 161, 105128. [Google Scholar] [CrossRef]
- Bialer, M.; Perucca, E. Does cannabidiol have antiseizure activity independent of its interactions with clobazam? An appraisal of the evidence from randomized controlled trials. Epilepsia 2020, 61, 1082–1089. [Google Scholar] [CrossRef]
- Tham, M.; Yilmaz, O.; Alaverdashvili, M.; Kelly, M.E.; Denovan-Wright, E.M.; Laprairie, R.B. Allosteric and orthosteric pharmacology of cannabidiol and cannabidiol-dimethylheptyl at the type 1 and type 2 cannabinoid receptors. Br. J. Pharmacol. 2019, 176, 1455–1469. [Google Scholar] [CrossRef]
- Laprairie, R.; Bagher, A.; Kelly, M.; Denovan-Wright, E. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, B.; Zheng, F.; Lu, S.; Li, Y.; Xiong, Y.; Yang, Q.; Yang, Y.; Fu, P.; Xiao, F.; et al. The Phosphodiesterase 10A Inhibitor PF-2545920 Enhances Hippocampal Excitability and Seizure Activity Involving the Upregulation of GluA1 and NR2A in Post-synaptic Densities. Front. Mol. Neurosci. 2017, 10, 100. [Google Scholar] [CrossRef]
- Cassano, T.; Calcagnini, S.; Pace, L.; De Marco, F.; Romano, A.; Gaetani, S. Cannabinoid Receptor 2 Signaling in Neurodegenerative Disorders: From Pathogenesis to a Promising Therapeutic Target. Front. Neurosci. 2017, 11, 30. [Google Scholar] [CrossRef]
- Xin, Q.; Xu, F.; Taylor, D.H.; Zhao, J.F.; Wu, J. The impact of cannabinoid type 2 receptors (CB2Rs) in neuroprotection against neurological disorders. Acta Pharm. Sin. 2020, 41, 1507–1518. [Google Scholar] [CrossRef]
- Wu, J. Cannabis, cannabinoid receptors, and endocannabinoid system: Yesterday, today, and tomorrow. Acta Pharm. Sin. 2019, 40, 297–299. [Google Scholar] [CrossRef]
- Aghaei, I.; Rostampour, M.; Shabani, M.; Naderi, N.; Motamedi, F.; Babaei, P.; Khakpour-Taleghani, B. Palmitoylethanolamide attenuates PTZ-induced seizures through CB1 and CB2 receptors. Epilepsy Res. 2015, 117, 23–28. [Google Scholar] [CrossRef]
- Huizenga, M.N.; Wicker, E.; Beck, V.C.; Forcelli, P.A. Anticonvulsant effect of cannabinoid receptor agonists in models of seizures in developing rats. Epilepsia 2017, 58, 1593–1602. [Google Scholar] [CrossRef]
- Rowley, S.; Sun, X.; Lima, I.V.; Tavenier, A.; de Oliveira, A.C.P.; Dey, S.K.; Danzer, S.C. Cannabinoid receptor 1/2 double-knockout mice develop epilepsy. Epilepsia 2017, 58, e162–e166. [Google Scholar] [CrossRef]
- Li, Y.; Kim, J. Distinct roles of neuronal and microglial CB2 cannabinoid receptors in the mouse hippocampus. Neuroscience 2017, 363, 11–25. [Google Scholar] [CrossRef]
- Rizzo, V.; Carletti, F.; Gambino, G.; Schiera, G.; Cannizzaro, C.; Ferraro, G.; Sardo, P. Role of CB2 receptors and cGMP pathway on the cannabinoid-dependent antiepileptic effects in an in vivo model of partial epilepsy. Epilepsy Res. 2014, 108, 1711–1718. [Google Scholar] [CrossRef]
- Shapiro, L.; Wong, J.C.; Escayg, A. Reduced cannabinoid 2 receptor activity increases susceptibility to induced seizures in mice. Epilepsia 2019, 60, 2359–2369. [Google Scholar] [CrossRef]
- da Silva Oliveira, G.L.; da Silva, J.; Dos Santos, C.L.d.S.A.P.; Feitosa, C.M.; de Castro Almeida, F.R. Anticonvulsant, Anxiolytic and Antidepressant Properties of the beta-caryophyllene in Swiss Mice: Involvement of Benzodiazepine-GABAAergic, Serotonergic and Nitrergic Systems. Curr. Mol. Pharm. 2021, 14, 36–51. [Google Scholar] [CrossRef]
- de Carvalho, C.R.; Hoeller, A.A.; Franco, P.L.; Martini, A.P.; Soares, F.M.; Lin, K.; Prediger, R.D.; Whalley, B.J.; Walz, R. The cannabinoid CB2 receptor-specific agonist AM1241 increases pentylenetetrazole-induced seizure severity in Wistar rats. Epilepsy Res. 2016, 127, 160–167. [Google Scholar] [CrossRef]
- Xia, K.K.; Shen, J.X.; Huang, Z.B.; Song, H.M.; Gao, M.; Chen, D.J.; Zhang, S.J.; Wu, J. Heterogeneity of cannabinoid ligand-induced modulations in intracellular Ca(2+) signals of mouse pancreatic acinar cells in vitro. Acta Pharm. Sin. 2019, 40, 410–417. [Google Scholar] [CrossRef]
- Cifelli, P.; Grace, A.A. Pilocarpine-induced temporal lobe epilepsy in the rat is associated with increased dopamine neuron activity. Int. J. Neuropsychopharmacol. 2012, 15, 957–964. [Google Scholar] [CrossRef]
- Floresco, S.B.; West, A.R.; Ash, B.; Moore, H.; Grace, A.A. Afferent modulation of dopamine neuron firing differentially regulates tonic and phasic dopamine transmission. Nat. Neurosci. 2003, 6, 968–973. [Google Scholar] [CrossRef]
- Ahmadi, M.; Dufour, J.P.; Seifritz, E.; Mirnajafi-Zadeh, J.; Saab, B.J. The PTZ kindling mouse model of epilepsy exhibits exploratory drive deficits and aberrant activity amongst VTA dopamine neurons in both familiar and novel space. Behav. Brain Res. 2017, 330, 1–7. [Google Scholar] [CrossRef]
- Boczek, T.; Yu, Q.; Zhu, Y.; Dodge-Kafka, K.L.; Goldberg, J.L.; Kapiloff, M.S. cAMP at Perinuclear mAKAPα Signalosomes Is Regulated by Local Ca2+ Signaling in Primary Hippocampal Neurons. eNeuro 2021, 8. [Google Scholar] [CrossRef]
- Kjaer, C.; Barzaghi, G.; Bak, L.K.; Goetze, J.P.; Yde, C.W.; Woldbye, D.; Pinborg, L.H.; Jensen, L.J. Transcriptome analysis in patients with temporal lobe epilepsy. Brain 2019, 142, e55. [Google Scholar] [CrossRef] [PubMed]
- Guelfi, S.; Botia, J.A.; Thom, M.; Ramasamy, A.; Perona, M.; Stanyer, L.; Martinian, L.; Trabzuni, D.; Smith, C.; Walker, R.; et al. Transcriptomic and genetic analyses reveal potential causal drivers for intractable partial epilepsy. Brain 2019, 142, 1616–1630. [Google Scholar] [CrossRef]
- Gunaydin, C.; Arslan, G.; Bilge, S.S. Proconvulsant effect of trans-cinnamaldehyde in pentylenetetrazole-induced kindling model of epilepsy: The role of TRPA1 channels. Neurosci. Lett. 2020, 721, 134823. [Google Scholar] [CrossRef]
- Ebrahimi-Ghiri, M.; Khakpai, F. The link between cannabinoids and depression. In The Neuroscience of Depression; Elsevier: Amsterdam, The Netherlands, 2021; pp. 293–300. [Google Scholar]
- Franco, R.; Villa, M.; Morales, P.; Reyes-Resina, I.; Gutierrez-Rodriguez, A.; Jimenez, J.; Jagerovic, N.; Martinez-Orgado, J.; Navarro, G. Increased expression of cannabinoid CB2 and serotonin 5-HT1A heteroreceptor complexes in a model of newborn hypoxic-ischemic brain damage. Neuropharmacology 2019, 152, 58–66. [Google Scholar] [CrossRef]
- Foust, A.J.; Yu, Y.; Popovic, M.; Zecevic, D.; McCormick, D.A. Somatic membrane potential and Kv1 channels control spike repolarization in cortical axon collaterals and presynaptic boutons. J. Neurosci. 2011, 31, 15490–15498. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, J.; Ji, W.; He, Q.; Mao, L.; Shu, Y. A- and D-type potassium currents regulate axonal action potential repolarization in midbrain dopamine neurons. Neuropharmacology 2021, 185, 108399. [Google Scholar] [CrossRef]
- Kohling, R.; Wolfart, J. Potassium Channels in Epilepsy. Cold Spring Harb. Perspect. Med. 2016, 6, a022871. [Google Scholar]
- Blomster, L.V.; Strobaek, D.; Hougaard, C.; Klein, J.; Pinborg, L.H.; Mikkelsen, J.D.; Christophersen, P. Quantification of the functional expression of the Ca(2+) -activated K(+) channel KCa 3.1 on microglia from adult human neocortical tissue. Glia 2016, 64, 2065–2078. [Google Scholar] [CrossRef]
- Brown, D. M-currents: An update. Trends Neurosci. 1988, 11, 294–299. [Google Scholar] [CrossRef]
- Gunthorpe, M.J.; Large, C.H.; Sankar, R. The mechanism of action of retigabine (ezogabine), a first-in-class K+ channel opener for the treatment of epilepsy. Epilepsia 2012, 53, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wolk, B.; Nolan, R.; Scott, C.E.; Kendall, D.A. Characterization of Subtype Selective Cannabinoid CB2 Receptor Agonists as Potential Anti-Inflammatory Agents. Pharmaceuticals 2021, 14, 378. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Cross, J.H.; Laux, L.; Marsh, E.; Miller, I.; Nabbout, R.; Scheffer, I.E.; Thiele, E.A.; Wright, S. Cannabidiol in Dravet Syndrome Study, G. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. N. Engl. J. Med. 2017, 376, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Thiele, E.A.; Marsh, E.D.; French, J.A.; Mazurkiewicz-Beldzinska, M.; Benbadis, S.R.; Joshi, C.; Lyons, P.D.; Taylor, A.; Roberts, C.; Sommerville, K.; et al. Cannabidiol in patients with seizures associated with Lennox-Gastaut syndrome (GWPCARE4): A randomised, double-blind, placebo-controlled phase 3 trial. Lancet 2018, 391, 1085–1096. [Google Scholar] [CrossRef]
- Little, J.P.; Villanueva, E.B.; Klegeris, A. Therapeutic potential of cannabinoids in the treatment of neuroinflammation associated with Parkinson’s disease. Mini Rev. Med. Chem. 2011, 11, 582–590. [Google Scholar] [CrossRef][Green Version]
- Cabral, G.A.; Marciano-Cabral, F. Cannabinoid receptors in microglia of the central nervous system: Immune functional relevance. J. Leukoc. Biol. 2005, 78, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Tang, Y. The central cannabinoid receptor type-2 (CB2) and chronic pain. Int. J. Neurosci. 2017, 127, 812–823. [Google Scholar] [CrossRef]
- Food, U.; Administration, D. FDA Approves First Drug Comprised of an Active Ingredient Derived from Marijuana to Treat Rare, Severe Forms of Epilepsy; U.S. Food & Drugs: Washington, DC, USA, 2018; p. 25. [Google Scholar]


{kind=link}
| Modulation Approaches | Seizure or Epilepsy Model | Response | References |
|---|---|---|---|
| - | PTZ induced seizures in heterozygous and homozygous CB2 knockout mice | Susceptibility | [84] |
| - | Handling-induced and spontaneous seizures | Susceptibility | [81] |
| 2-Arachidonoylglycerol (2-AG) | Kainate induced seizures in CB1R and CB2R double-knockout mice | Suppressed excitatory | [12] |
| CB2 agonist beta caryophyllene | PTZ induced seizures in mice | Anticonvulsant activity | [85] |
| CB2 agonist beta caryophyllene | Induced by kainic acid (KA) seizure in mice | Decreased the seizure scores | [85] |
| CB2 antagonist AM630 | Kainate induced seizures in CB1R knockout mice | Increased seizure susceptibility | [80] |
| CB2 agonist AM1241 | PTZ induced seizures in rats | Increased seizure severity | [86] |
| CB2 agonist HU-308 | DMCM and PTZ induced seizures in rats | No significant effect on seizure severity | [80] |
| CB2R agonist JWH133 | PTZ induced seizures in mice | No significant effect on seizure severity | [84] |
| CB2R antagonist SR144528 | PTZ induced seizures in mice | Increases seizure susceptibility | [84] |
| CB1/2R mixed agonist WIN 55,212-2 | DMCM and PTZ induced seizures in rats | Anticonvulsant effects | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, X.; Zeng, Y.; Wu, J. The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment. Int. J. Mol. Sci. 2021, 22, 8961. https://doi.org/10.3390/ijms22168961
Ji X, Zeng Y, Wu J. The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment. International Journal of Molecular Sciences. 2021; 22(16):8961. https://doi.org/10.3390/ijms22168961
Chicago/Turabian StyleJi, Xiaoyu, Yang Zeng, and Jie Wu. 2021. "The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment" International Journal of Molecular Sciences 22, no. 16: 8961. https://doi.org/10.3390/ijms22168961
APA StyleJi, X., Zeng, Y., & Wu, J. (2021). The CB2 Receptor as a Novel Therapeutic Target for Epilepsy Treatment. International Journal of Molecular Sciences, 22(16), 8961. https://doi.org/10.3390/ijms22168961