
Optimizing the Expression of Human Dopamine Receptors in Escherichia coli


Abstract
:1. Introduction
2. Results and Discussion
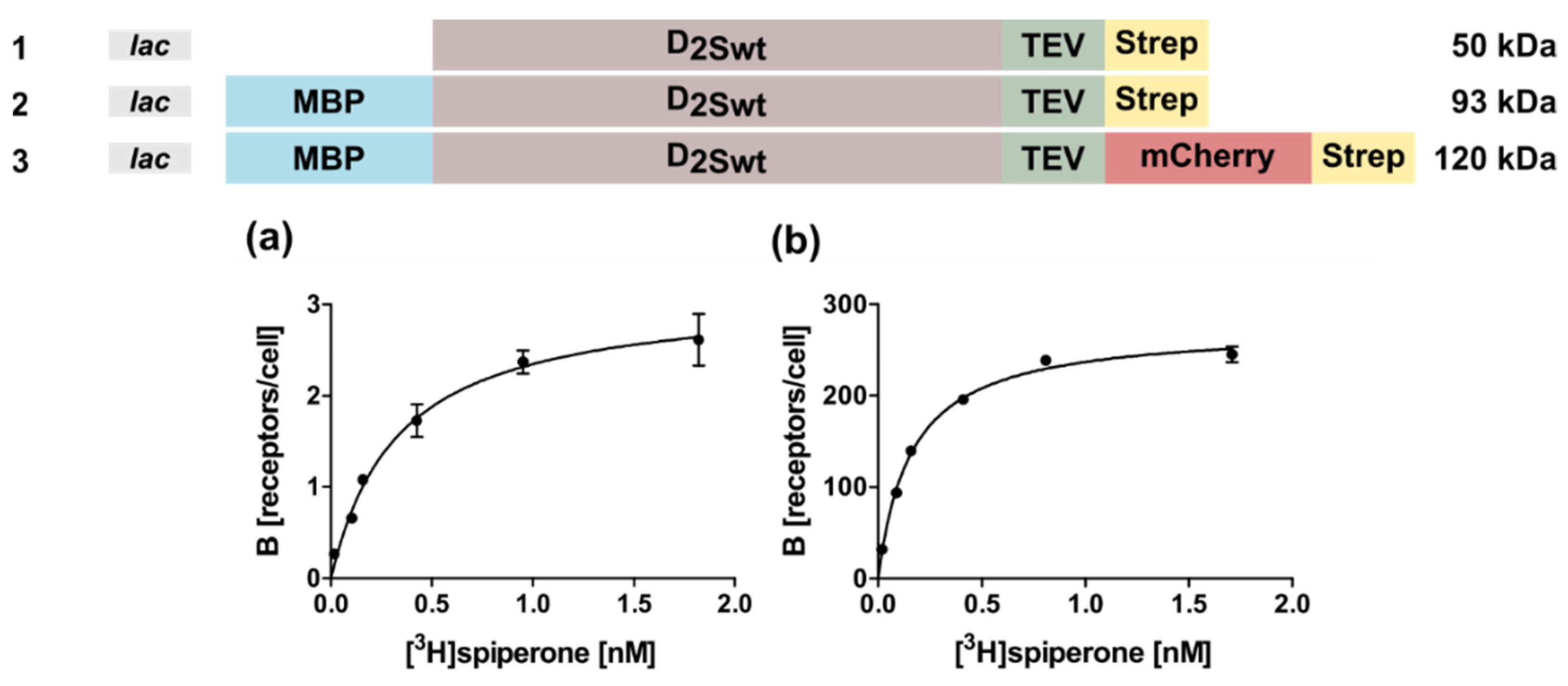
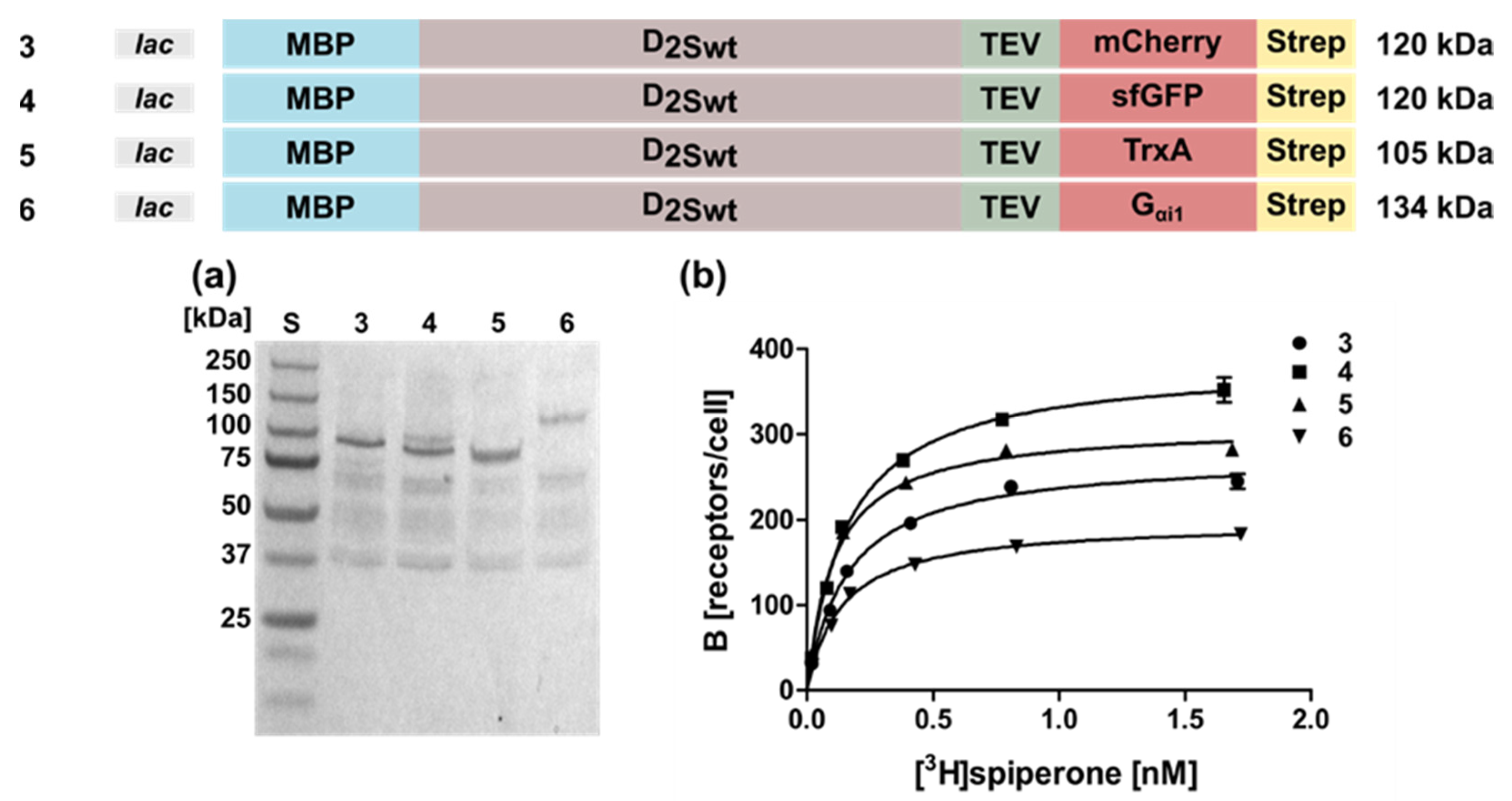
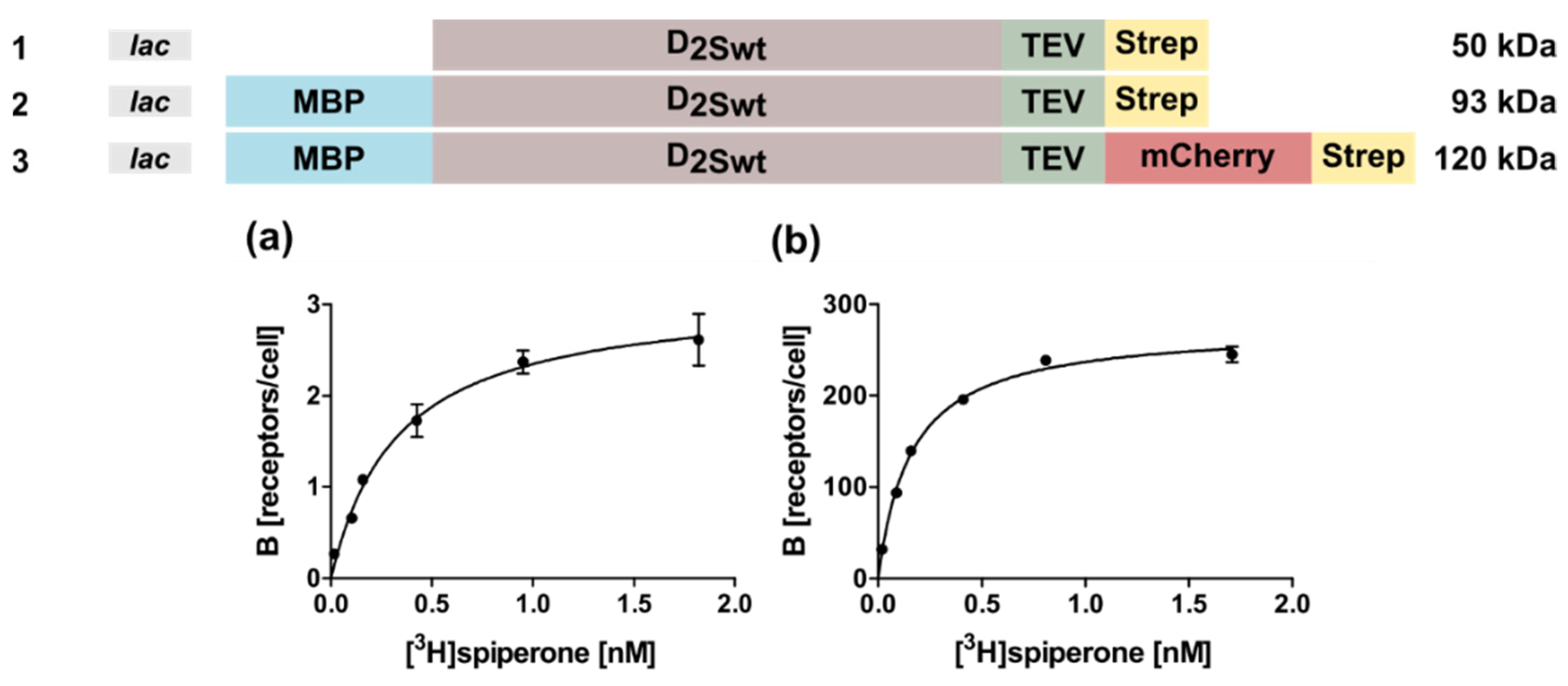
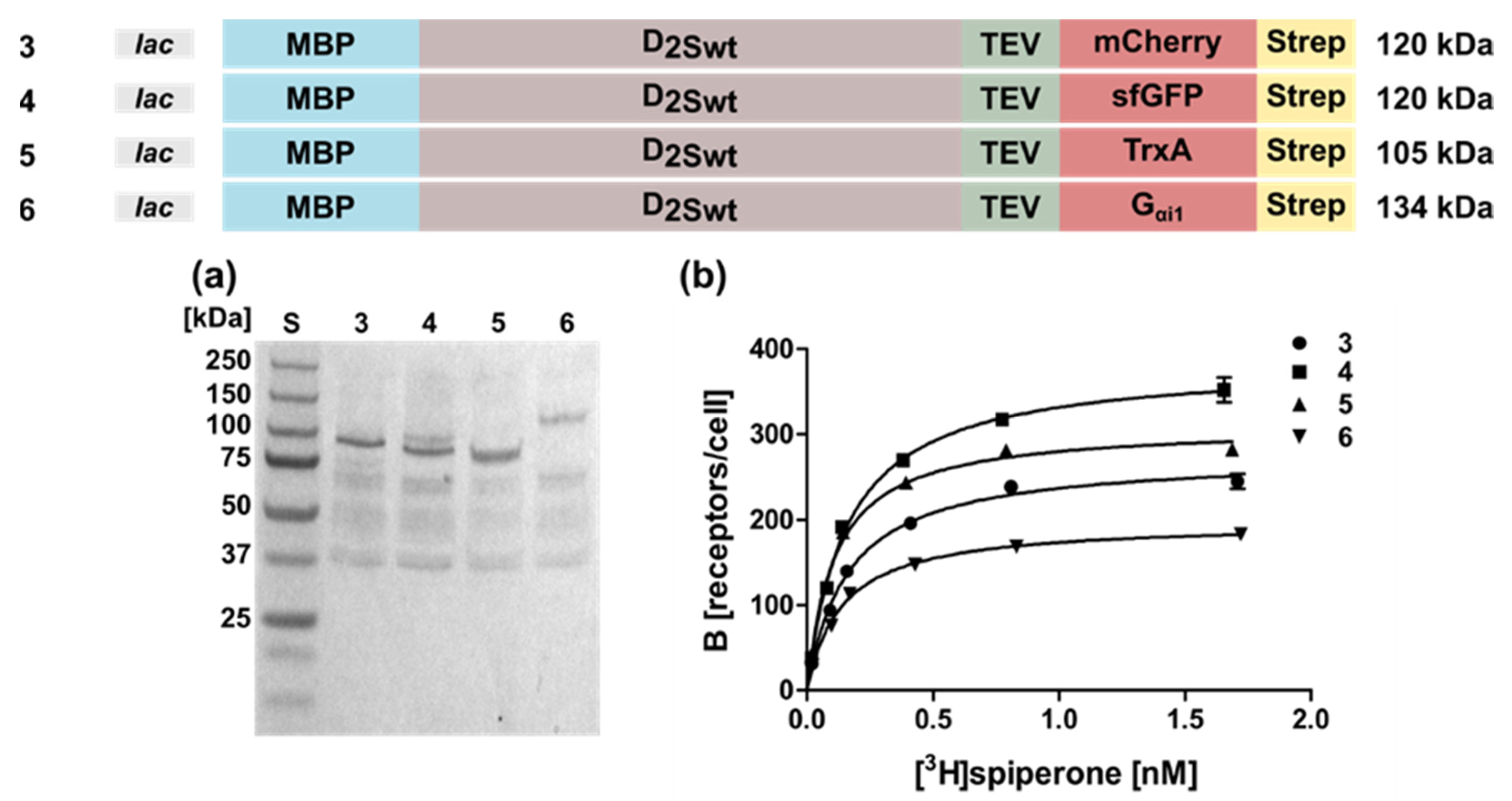
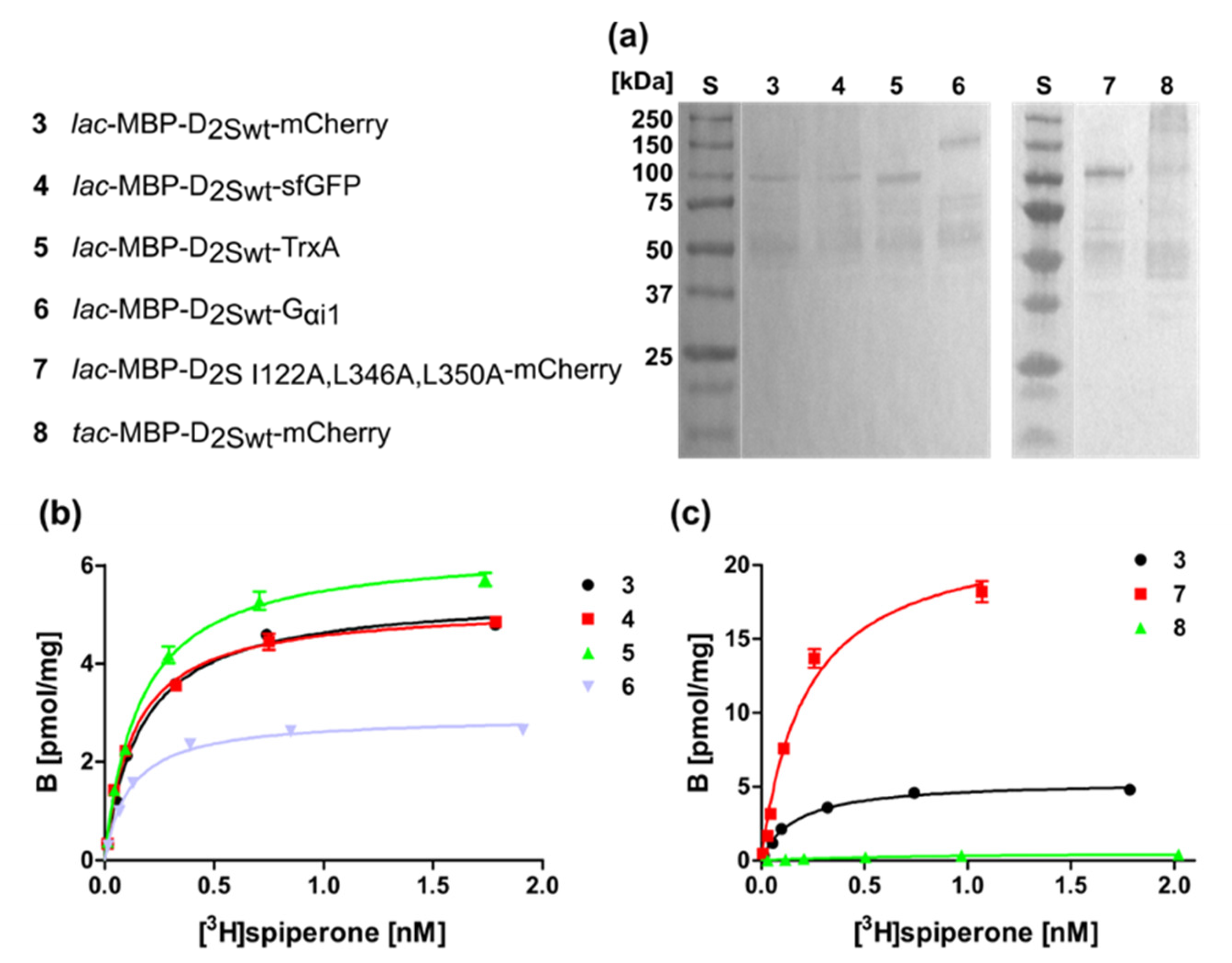
2.1. Expression of the Human D2S Receptor in E. coli Using N- and C-Terminal Fusion Proteins
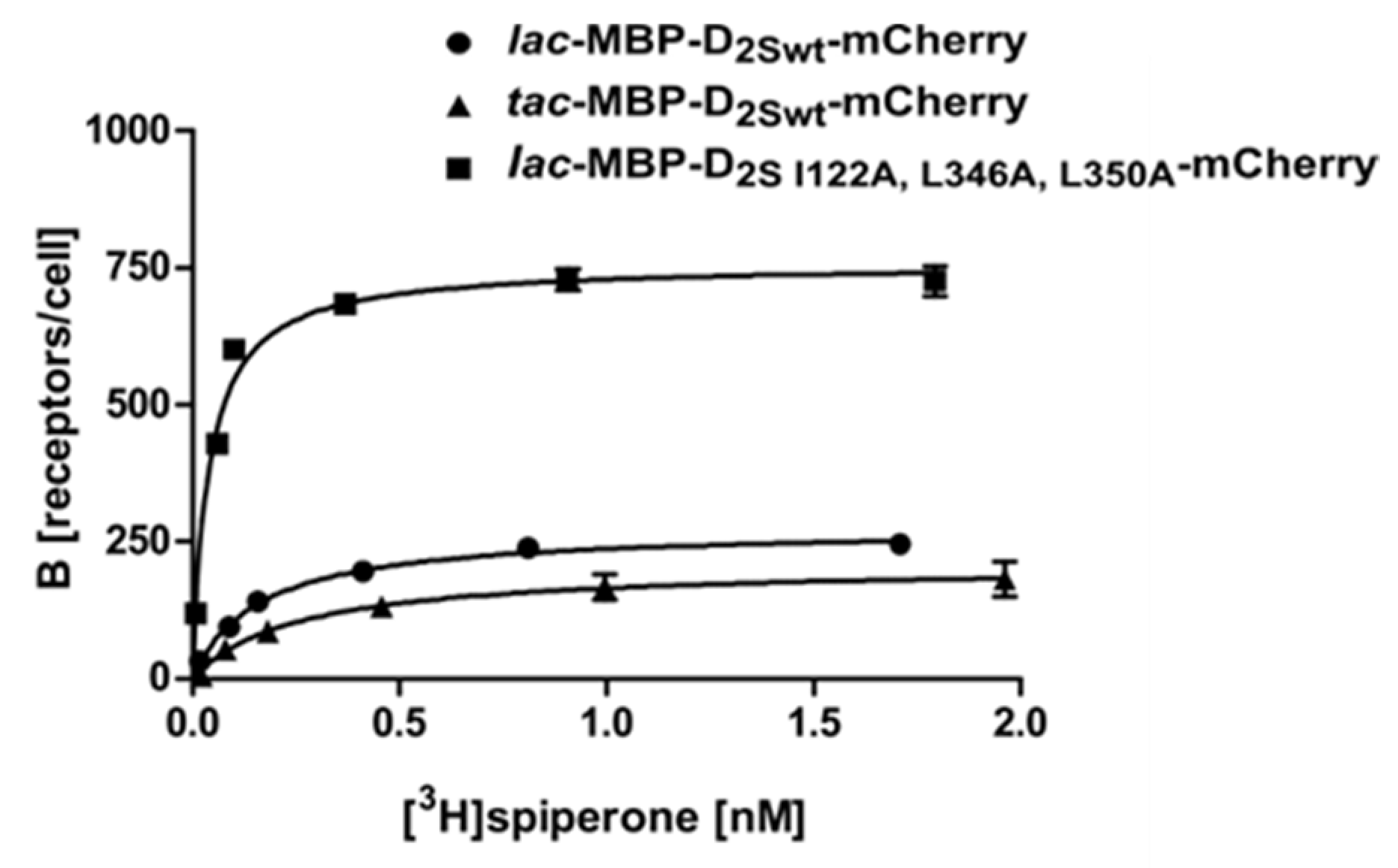
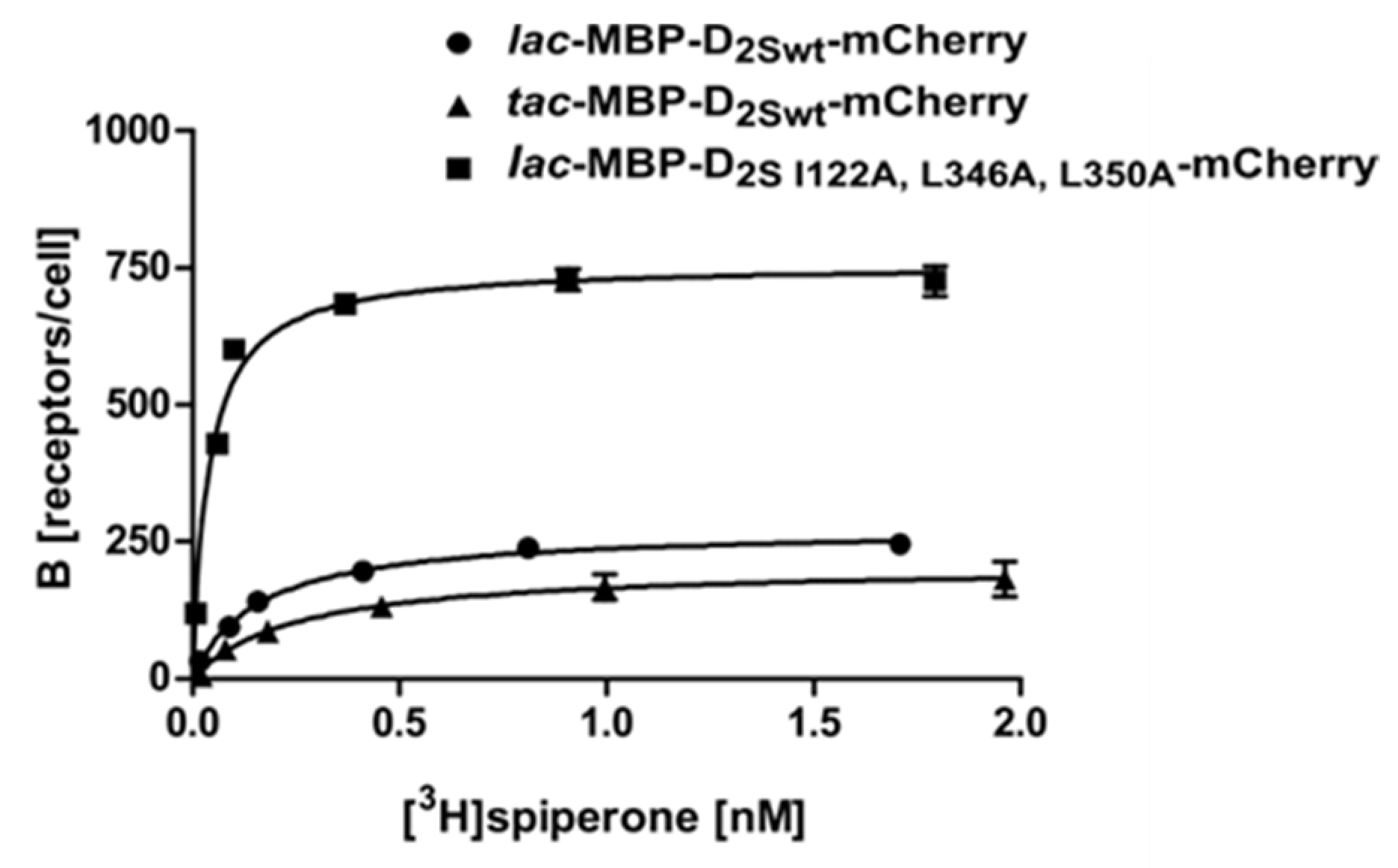
2.2. Use of the Tac Promoter and Introduction of Thermostabilizing Mutations
2.3. Isolation and Analysis of Membrane Homogenates after Expression of D2S
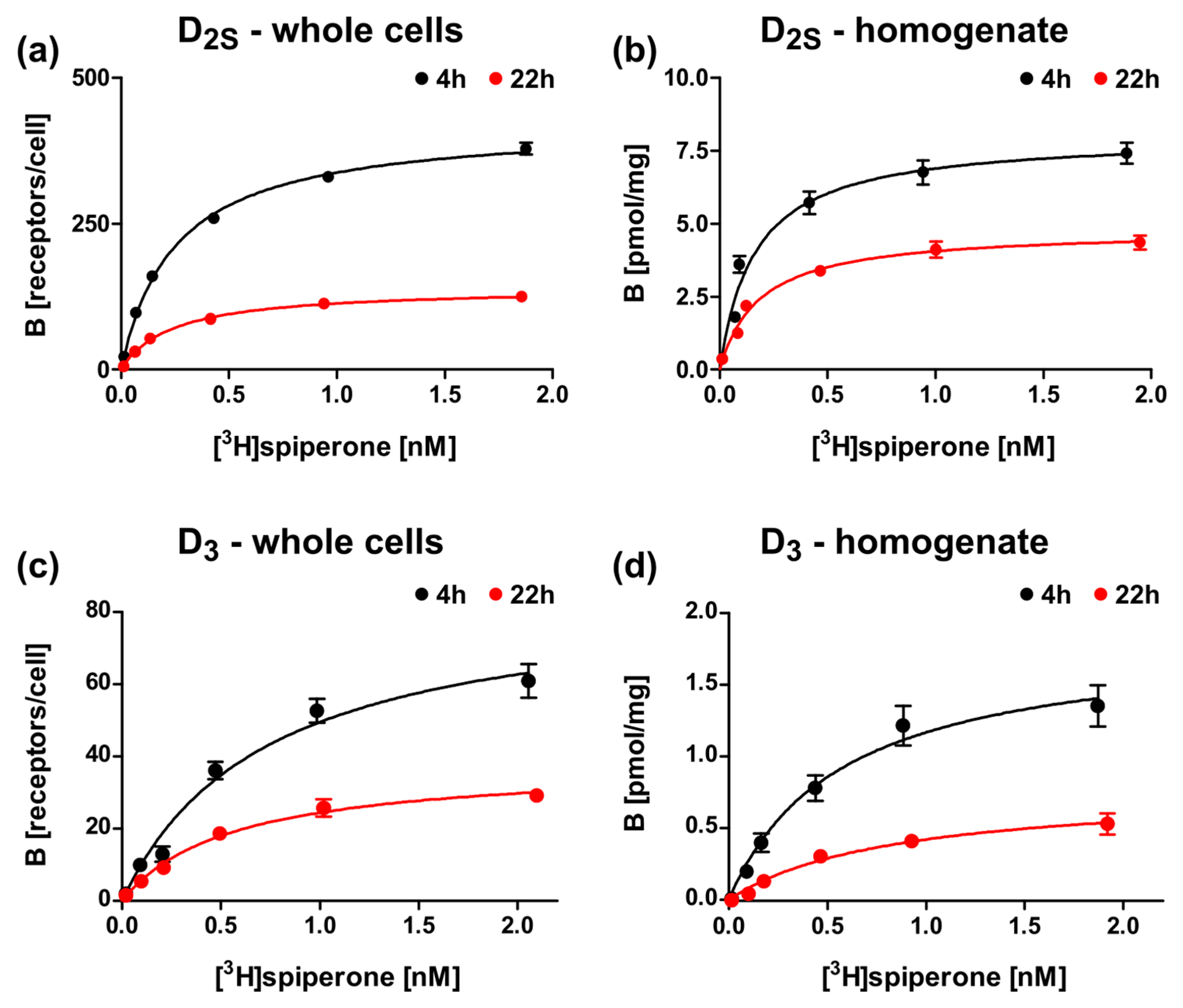
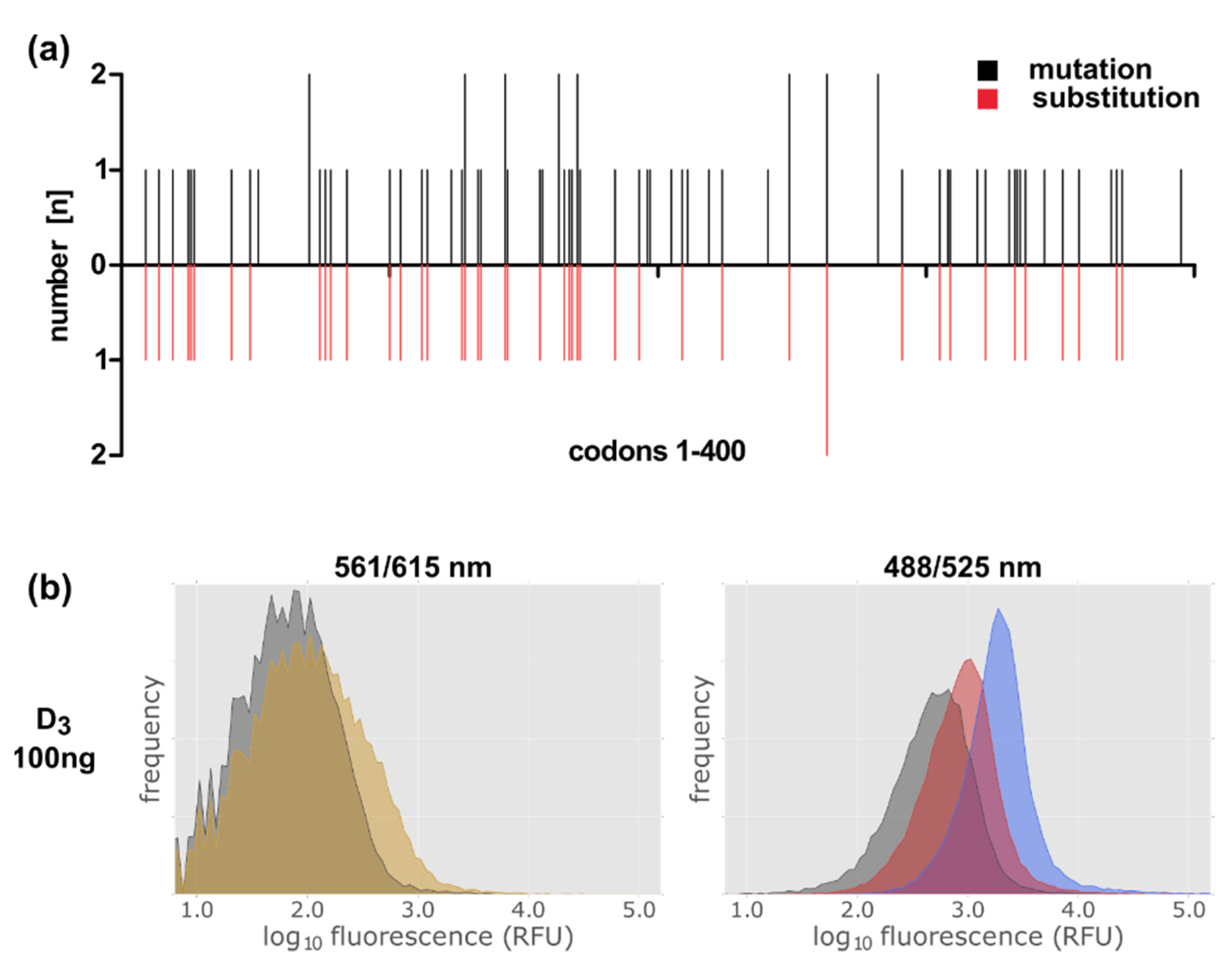
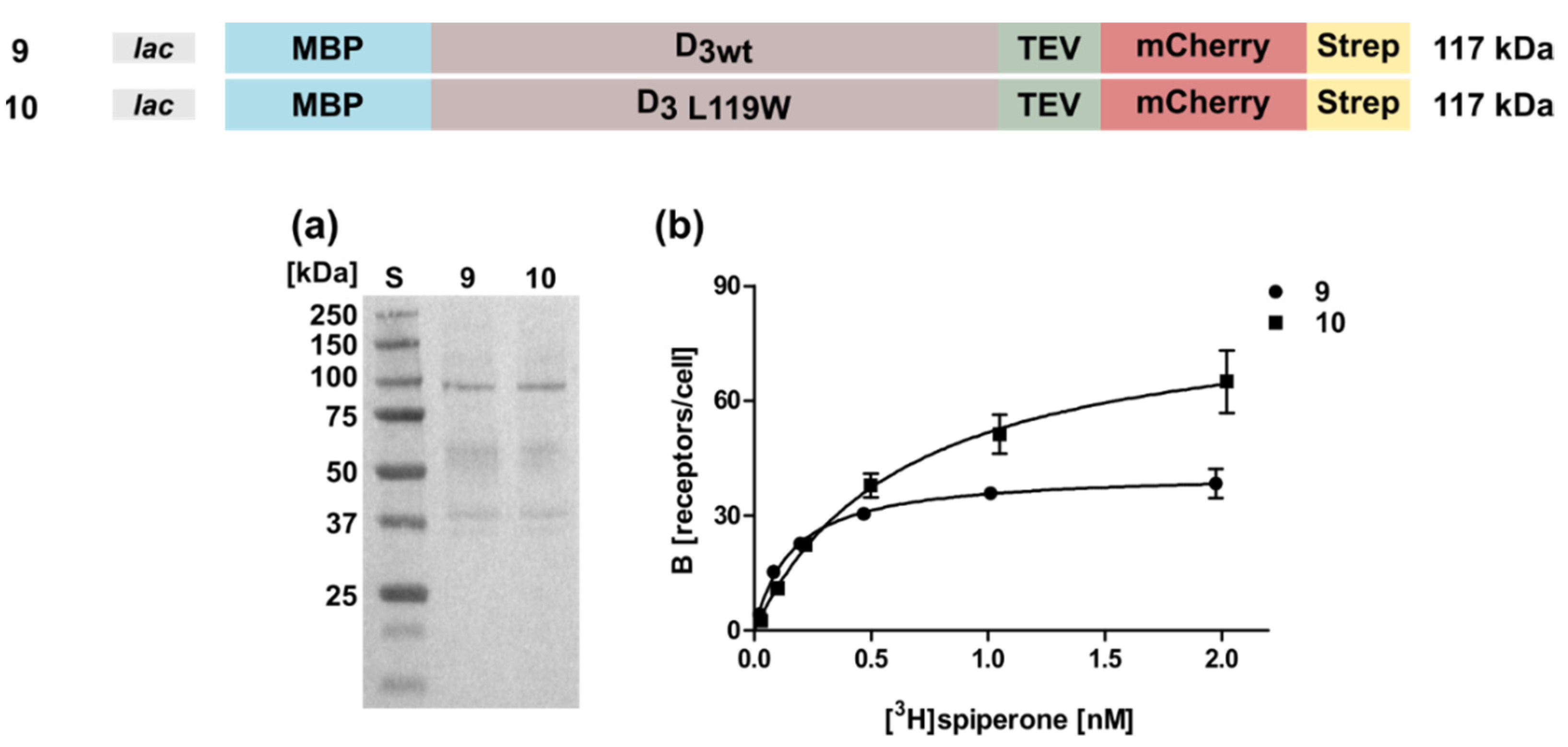
2.4. Expression of the D3 Receptor in the E. coli System Is Less Efficient
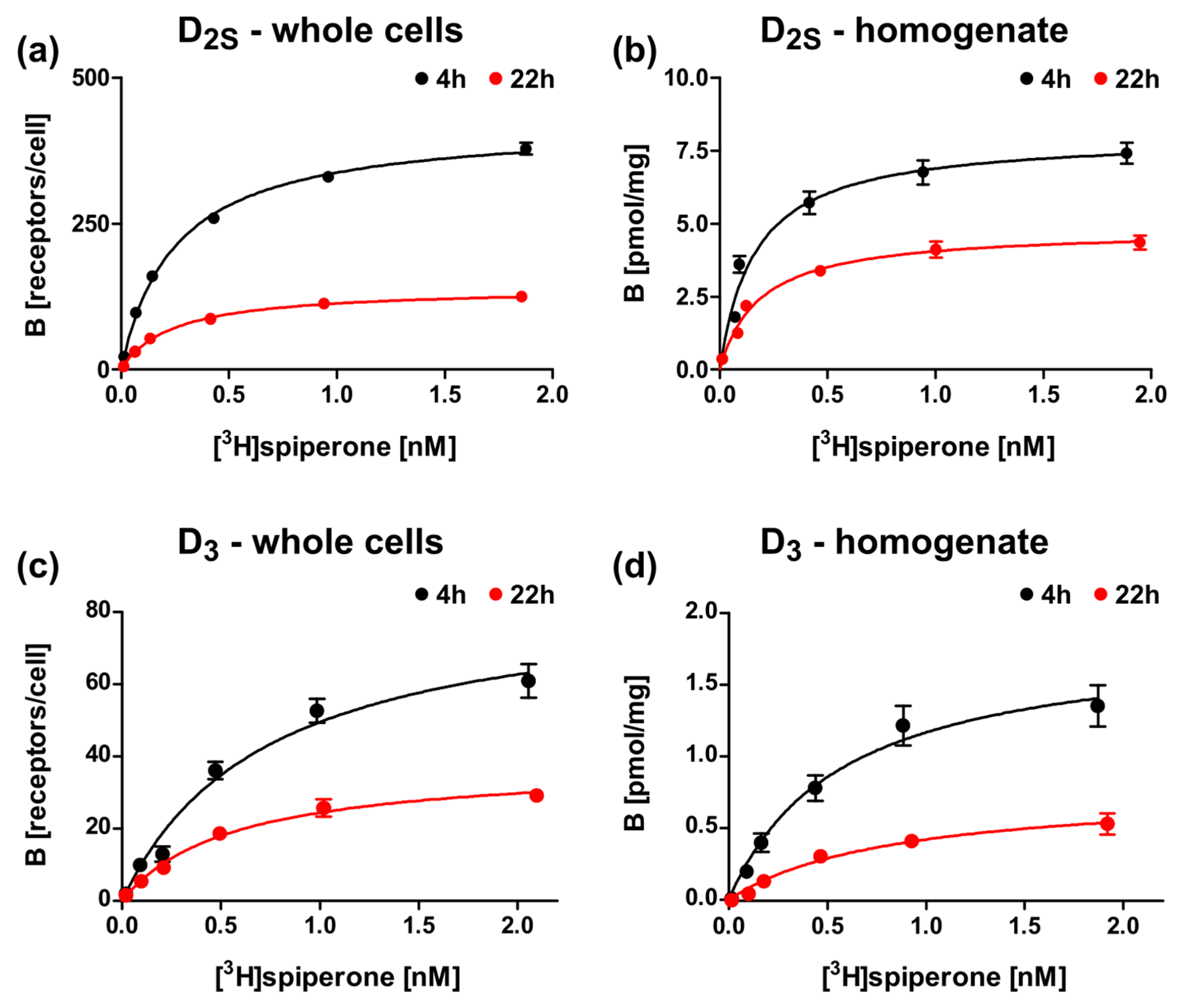
2.5. Optimization of the Induction Times
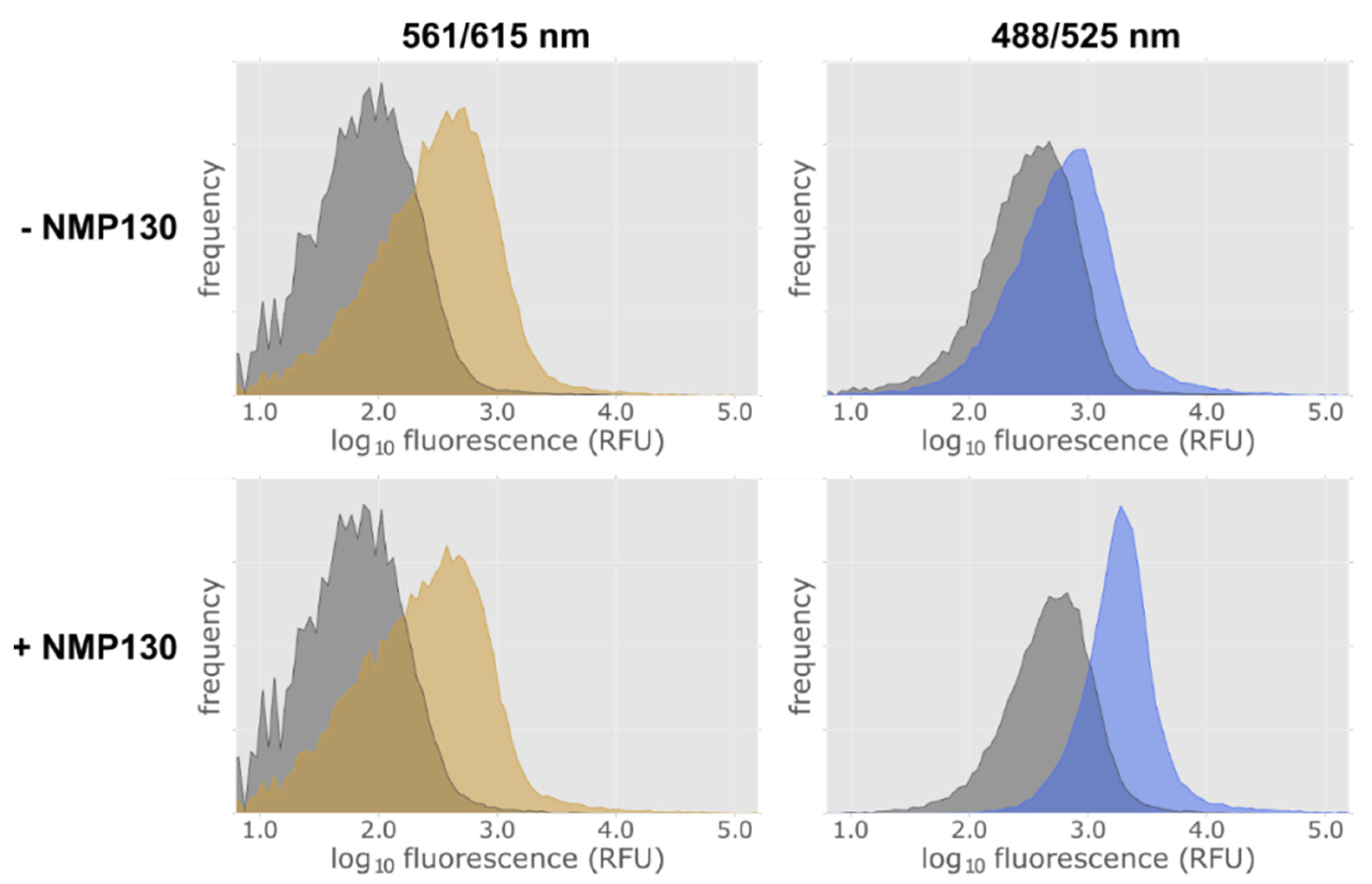
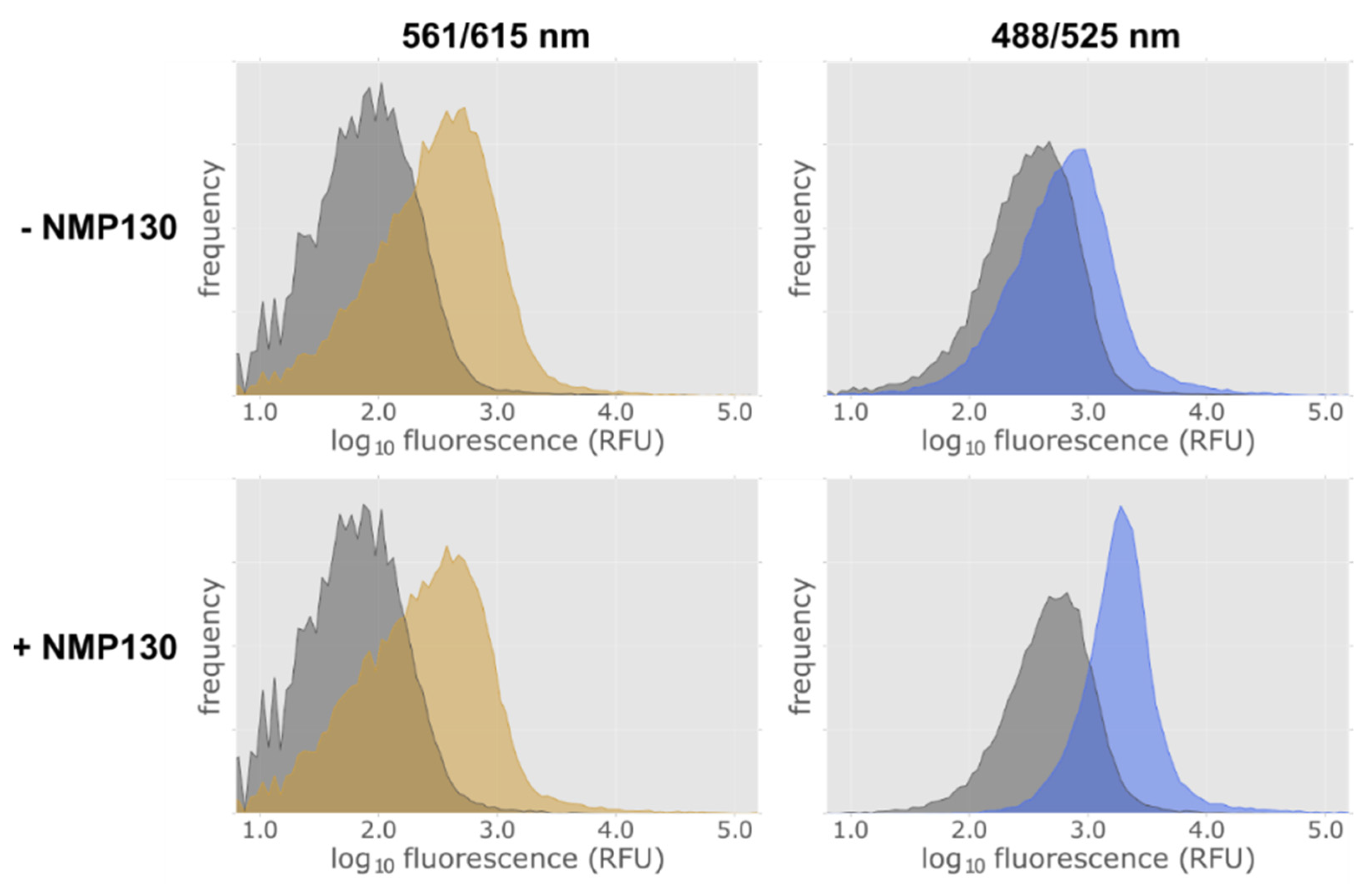
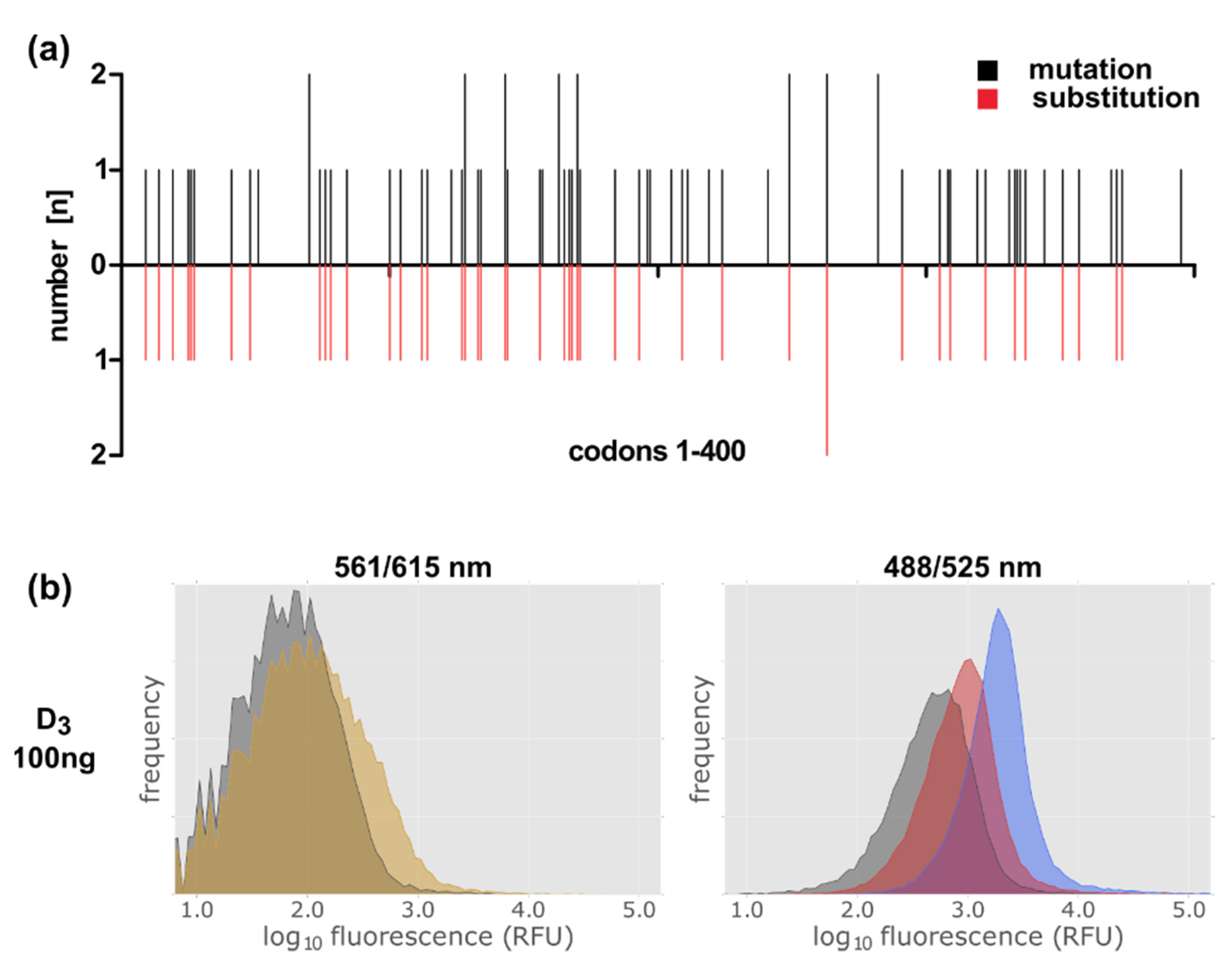
2.6. Establishment of a Method to Select Functional Thermostable Dopamine Receptor Variants in E. coli
3. Materials and Methods
3.1. Gene Cloning and Plasmid Preparation
3.2. Heterologous Gene Expression in E. coli
3.3. Preparation of Membrane Homogenate
3.4. Western Blot Analysis
3.5. Radioligand Binding Assay
3.6. Fluorescent Activated Cell Sorting
3.7. Library Construction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wacker, D.; Stevens, R.C.; Roth, B.L. How ligands illuminate GPCR molecular pharmacology. Cell 2017, 170, 414–427. [Google Scholar] [CrossRef] [Green Version]
- Sriram, K.; Insel, P.A. G Protein-Coupled Receptors as Targets for Approved Drugs: How Many Targets and How Many Drugs? Mol. Pharmacol. 2018, 93, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Latorraca, N.R.; Venkatakrishnan, A.J.; Dror, R.O. GPCR Dynamics: Structures in Motion. Chem. Rev. 2017, 117, 139–155. [Google Scholar] [CrossRef]
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and dynamics of GPCR signaling complexes. Nat. Struct. Mol. Biol. 2018, 25, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [Green Version]
- Vallone, D.; Picetti, R.; Borrelli, E. Structure and function of dopamine receptors. Neurosci. Biobehav. Rev. 2000, 24, 125–132. [Google Scholar] [CrossRef]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.-G.; Schiöth, H.B. The G-Protein-Coupled Receptors in the Human Genome Form Five Main Families. Phylogenetic Analysis, Paralogon Groups, and Fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef] [Green Version]
- Congreve, M.; de Graaf, C.; Swain, N.A.; Tate, C.G. Impact of GPCR Structures on Drug Discovery. Cell 2020, 181, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Sarramegna, V.; Talmont, F.; Demange, P.; Milon, A. Heterologous expression of G-protein-coupled receptors: Comparison of expression systems from the standpoint of large-scale production and purification. Cell. Mol. Life Sci. 2003, 60, 1529–1546. [Google Scholar] [CrossRef] [PubMed]
- Franke, B.; Opitz, C.; Isogai, S.; Grahl, A.; Delgado, L.; Gossert, A.D.; Grzesiek, S. Production of isotope-labeled proteins in insect cells for NMR. J. Biomol. NMR 2018, 71, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opitz, C.; Isogai, S.; Grzesiek, S. An economic approach to efficient isotope labeling in insect cells using homemade 15N-, 13C- and 2H-labeled yeast extracts. J. Biomol. NMR 2015, 62, 373–385. [Google Scholar] [CrossRef]
- Werner, K.; Richter, C.; Klein-Seetharaman, J.; Schwalbe, H. Isotope labeling of mammalian GPCRs in HEK293 cells and characterization of the C-terminus of bovine rhodopsin by high resolution liquid NMR spectroscopy. J. Biomol. NMR 2008, 40, 49–53. [Google Scholar] [CrossRef]
- Heydenreich, F.M.; Vuckovic, Z.; Matkovic, M.; Veprintsev, D.B. Stabilization of G protein-coupled receptors by point mutations. Front. Pharmacol. 2015, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Baneres, J.-L.; Martin, A.; Hullot, P.; Girard, J.-P.; Rossi, J.-C.; Parello, J. Structure-based analysis of GPCR function: Conformational adaptation of both agonist and receptor upon leukotriene B4 binding to recombinant BLT1. J. Mol. Biol. 2003, 329, 801–814. [Google Scholar] [CrossRef]
- Ferré, G.; Louet, M.; Saurel, O.; Delort, B.; Czaplicki, G.; M’Kadmi, C.; Damian, M.; Renault, P.; Cantel, S.; Gavara, L.; et al. Structure and dynamics of G protein-coupled receptor-bound ghrelin reveal the critical role of the octanoyl chain. Proc. Natl. Acad. Sci. USA 2019, 116, 17525–17530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodevski, I.; Plückthun, A. Evolution of three human GPCRs for higher expression and stability. J. Mol. Biol. 2011, 408, 599–615. [Google Scholar] [CrossRef]
- Grisshammer, R.; Duckworth, R.; Henderson, R. Expression of a rat neurotensin receptor in Escherichia coli. Biochem. J. 1993, 295, 571–576. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, C.A.; Dodevski, I.; Kenig, M.; Dudli, S.; Mohr, A.; Hermans, E.; Plückthun, A. Directed evolution of a G protein-coupled receptor for expression, stability, and binding selectivity. Proc. Natl. Acad. Sci. USA 2008, 105, 14808–14813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skretas, G.; Georgiou, G. Genetic Analysis of G Protein-Coupled Receptor Expression in Escherichia coli. Biotechnol. Bioeng. 2009, 102, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, J.; Grisshammer, R. Purification of a rat neurotensin receptor expressed in Escherichia coli. Biochem. J. 1996, 317 Pt 3, 891–899. [Google Scholar] [CrossRef] [Green Version]
- Weiss, H.M.; Grisshammer, R. Purification and characterization of the human adenosine A2a receptor functionally expressed in Escherichia coli. Eur. J. Biochem. 2002, 269, 82–92. [Google Scholar] [CrossRef]
- Yeliseev, A.A.; Wong, K.K.; Soubias, O.; Gawrisch, K. Expression of human peripheral cannabinoid receptor for structural studies. Protein Sci. 2005, 14, 2638–2653. [Google Scholar] [CrossRef] [Green Version]
- Egloff, P.; Hillenbrand, M.; Klenk, C.; Batyuk, A.; Heine, P.; Balada, S.; Schlinkmann, K.M.; Scott, D.J.; Schütz, M.; Plückthun, A. Structure of signaling-competent neurotensin receptor 1 obtained by directed evolution in Escherichia coli. Proc. Natl. Acad. Sci. USA 2014, 111, E655–E662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, M.; Deluigi, M.; Pantić, M.; Vacca, S.; Baumann, C.; Scott, D.J.; Plückthun, A.; Zerbe, O. Optimizing the α1B-adrenergic receptor for solution NMR studies. Biochim. Biophys. Acta (BBA) Biomembr. 2020, 1862, 183354. [Google Scholar] [CrossRef]
- Vanhauwe, J.F.; Josson, K.; Luyten, W.H.; Driessen, A.J.; Leysen, J.E. G-protein sensitivity of ligand binding to human dopamine D2 and D3 receptors expressed in Escherichia coli: Clues for a constrained D3receptor structure. J. Pharmacol. Exp. Ther. 2000, 295, 274–283. [Google Scholar]
- De Mei, C.; Ramos, M.; Iitaka, C.; Borrelli, E. Getting specialized: Presynaptic and postsynaptic dopamine D2 receptors. Curr. Opin. Pharmacol. 2009, 9, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Toso, R.; Sommer, B.; Ewert, M.; Herb, A.; Pritchett, D.B.; Bach, A.; Shivers, B.D.; Seeburg, P.H. The dopamine D2 receptor: Two molecular forms generated by alternative splicing. EMBO J. 1989, 8, 4025–4034. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Che, T.; Levit, A.; Shoichet, B.K.; Wacker, D.; Roth, B.L. Structure of the D2 dopamine receptor bound to the atypical antipsychotic drug risperidone. Nature 2018, 555, 269–273. [Google Scholar] [CrossRef]
- Klenk, C.; Ehrenmann, J.; Schütz, M.; Plückthun, A. A generic selection system for improved expression and thermostability of G protein-coupled receptors by directed evolution. Sci. Rep. 2016, 6, 21294. [Google Scholar] [CrossRef] [Green Version]
- Schlinkmann, K.M.; Plückthun, A. Directed evolution of G-protein-coupled receptors for high functional expression and detergent stability. Methods Enzymol. 2013, 520, 67–97. [Google Scholar] [CrossRef] [PubMed]
- Chien, E.Y.T.; Liu, W.; Zhao, Q.; Katritch, V.; Han, G.W.; Hanson, M.A.; Shi, L.; Newman, A.H.; Javitch, J.A.; Cherezov, V.; et al. Structure of the human dopamine D3 receptor in complex with a D2/D3 selective antagonist. Science 2010, 330, 1091–1095. [Google Scholar] [CrossRef] [Green Version]
- Allikalt, A.; Purkayastha, N.; Flad, K.; Schmidt, M.F.; Tabor, A.; Gmeiner, P.; Hübner, H.; Weikert, D. Fluorescent ligands for dopamine D2/D3 receptors. Sci. Rep. 2020, 10, 21842. [Google Scholar] [CrossRef]
- Surre, J.; Saint-Ruf, C.; Collin, V.; Orenga, S.; Ramjeet, M.; Matic, I. Strong increase in the autofluorescence of cells signals struggle for survival. Sci. Rep. 2018, 8, 12088. [Google Scholar] [CrossRef] [PubMed]
- McAllister, G.; Knowles, M.R.; Ward-Booth, S.M.; Sinclair, H.A.; Patel, S.; Marwood, R.; Emms, F.; Patel, S.; Smith, A.; Seabrook, G.R.; et al. Functional Coupling of Human D2, D3, and D4 Dopamine Receptors in HEK293 Cells. J. Recept. Signal Transduct. 1995, 15, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Tian, J. Circular Polymerase Extension Cloning of Complex Gene Libraries and Pathways. PLoS ONE 2009, 4, e6441. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Tian, J. Circular polymerase extension cloning for high-throughput cloning of complex and combinatorial DNA libraries. Nat. Protoc. 2011, 6, 242–251. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Hübner, H.; Haubmann, C.; Utz, W.; Gmeiner, P. Conjugated Enynes as Nonaromatic Catechol Bioisosteres: Synthesis, Binding Experiments, and Computational Studies of Novel Dopamine Receptor Agonists Recognizing Preferentially the D3 Subtype. J. Med. Chem. 2000, 43, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Hübner, H.; Schellhorn, T.; Gienger, M.; Schaab, C.; Kaindl, J.; Leeb, L.; Clark, T.; Möller, D.; Gmeiner, P. Structure-guided development of heterodimer-selective GPCR ligands. Nat. Commun. 2016, 7, 12298. [Google Scholar] [CrossRef] [Green Version]
- Korbie, D.J.; Mattick, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nat. Protoc. 2008, 3, 1452–1456. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Werner, E.; Holder, A.A.; Hoheisel, J.D. Growth and Storage of YAC Clones in Hogness Freezing Medium. Available online: https://pubmed.ncbi.nlm.nih.gov/9060445/ (accessed on 2 December 2020).
- Fan, L.; Tan, L.; Chen, Z.; Qi, J.; Nie, F.; Luo, Z.; Cheng, J.; Wang, S. Haloperidol bound D2 dopamine receptor structure inspired the discovery of subtype selective ligands. Nat. Commun. 2020, 11, 1074. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Chen, K.-Y.M.; Clark, M.J.; Hijazi, M.; Kumari, P.; Bai, X.-C.; Sunahara, R.K.; Barth, P.; Rosenbaum, D.M. Structure of a D2 dopamine receptor-G-protein complex in a lipid membrane. Nature 2020, 584, 125–129. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct Number | Construct Name | * KD [nM] | * Bmax [Receptors/Cell] |
|---|---|---|---|
| 2 | lac-MBP-D2Swt | 0.31 ± 0.11 | 6.5 ± 2.2 |
| 3 | lac-MBP-D2Swt-mCherry | 0.14 ± 0.04 | 270 ± 12 |
| 4 | lac-MBP-D2Swt-sfGFP | 0.13 ± 0.04 | 330 ± 21 |
| 5 | lac-MBP-D2Swt-TrxA | 0.11 ± 0.03 | 280 ± 10 |
| 6 | lac-MBP-D2Swt-Gαi1 | 0.14 ± 0.01 | 190 ± 18 |
| 7 | lac-MBP-D2S- I122A,L346A,L350A-mCherry | 0.11 ± 0.04 | 740 ± 38 |
| 8 | tac-MBP-D2Swt-mCherry | 0.23 ± 0.03 | 190 ± 33 |
| 9 | lac-MBP-D3wt-mCherry | 0.52 ± 0.24 | 46 ± 3 |
| 10 | lac-MBP-D3 L119W-mCherry | 1.0 ± 0.1 | 69 ± 8 |
| Construct Number | Construct Name | * KD [nM] | * Bmax [Pmol/mg] |
|---|---|---|---|
| 3 | lac-MBP-D2Swt-mCherry | 0.11 ± 0.09 | 3.4 ± 2.9 |
| 4 | lac-MBP-D2Swt-sfGFP | 0.10 ± 0.04 | 3.3 ± 2.7 |
| 5 | lac-MBP-D2Swt-TrxA | 0.12 ± 0.06 | 4.1 ± 3.3 |
| 6 | lac-MBP-D2Swt-Gαi1 | 0.090 ± 0.05 | 2.0 ± 1.4 |
| 7 | lac-MBP-D2S-I122A,L346A,L350A-mCherry | 0.16 ± 0.09 | 13 ± 10 |
| 8 | tac-MBP-D2Swt-mCherry | 0.48 ± 0.39 | 0.23 ± 0.17 |
| Construct Number | Construct Name | Whole Cells | |||
|---|---|---|---|---|---|
| a KD [nM] | a Bmax [receptors/cell] | ||||
| 4 h | 22 h | 4 h | 22 h | ||
| 3 | lac-MBP-D2Swt-mCherry | 0.34 ± 0.12 b | 0.14 ± 0.04 a | 410 ± 21 b | 270 ± 12 a |
| 9 | lac-MBP-D3wt-mCherry | 0.85 ± 0.14 b | 0.52 ± 0.24 a | 110 ± 42 b | 46 ± 3 a |
| Homogenate | |||||
| b KD [nM] | b Bmax [pmol/mg] | ||||
| 4 h | 22 h | 4 h | 22 h | ||
| 3 | lac-MBP-D2Swt-mCherry | 0.20 ± 0.04 b | 0.11 ± 0.09 b | 8.1 ± 0.14 b | 3.4 ± 2.9 b |
| 9 | lac-MBP-D3wt-mCherry | 0.66 ± 0.13 b | 0.50 ± 0.21 b | 2.0 ± 0.21 b | 0.52 ± 0.25 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boritzki, V.; Hübner, H.; Allikalt, A.; Gmeiner, P.; Wöhrl, B.M. Optimizing the Expression of Human Dopamine Receptors in Escherichia coli. Int. J. Mol. Sci. 2021, 22, 8647. https://doi.org/10.3390/ijms22168647
Boritzki V, Hübner H, Allikalt A, Gmeiner P, Wöhrl BM. Optimizing the Expression of Human Dopamine Receptors in Escherichia coli. International Journal of Molecular Sciences. 2021; 22(16):8647. https://doi.org/10.3390/ijms22168647
Chicago/Turabian StyleBoritzki, Vanessa, Harald Hübner, Anni Allikalt, Peter Gmeiner, and Birgitta M. Wöhrl. 2021. "Optimizing the Expression of Human Dopamine Receptors in Escherichia coli" International Journal of Molecular Sciences 22, no. 16: 8647. https://doi.org/10.3390/ijms22168647
APA StyleBoritzki, V., Hübner, H., Allikalt, A., Gmeiner, P., & Wöhrl, B. M. (2021). Optimizing the Expression of Human Dopamine Receptors in Escherichia coli. International Journal of Molecular Sciences, 22(16), 8647. https://doi.org/10.3390/ijms22168647