
Selection of Highly Proliferative and Multipotent Meniscus Progenitors through Differential Adhesion to Fibronectin: A Novel Approach in Meniscus Tissue Engineering

 , , ,
, , , 
Abstract
1. Introduction
2. Results
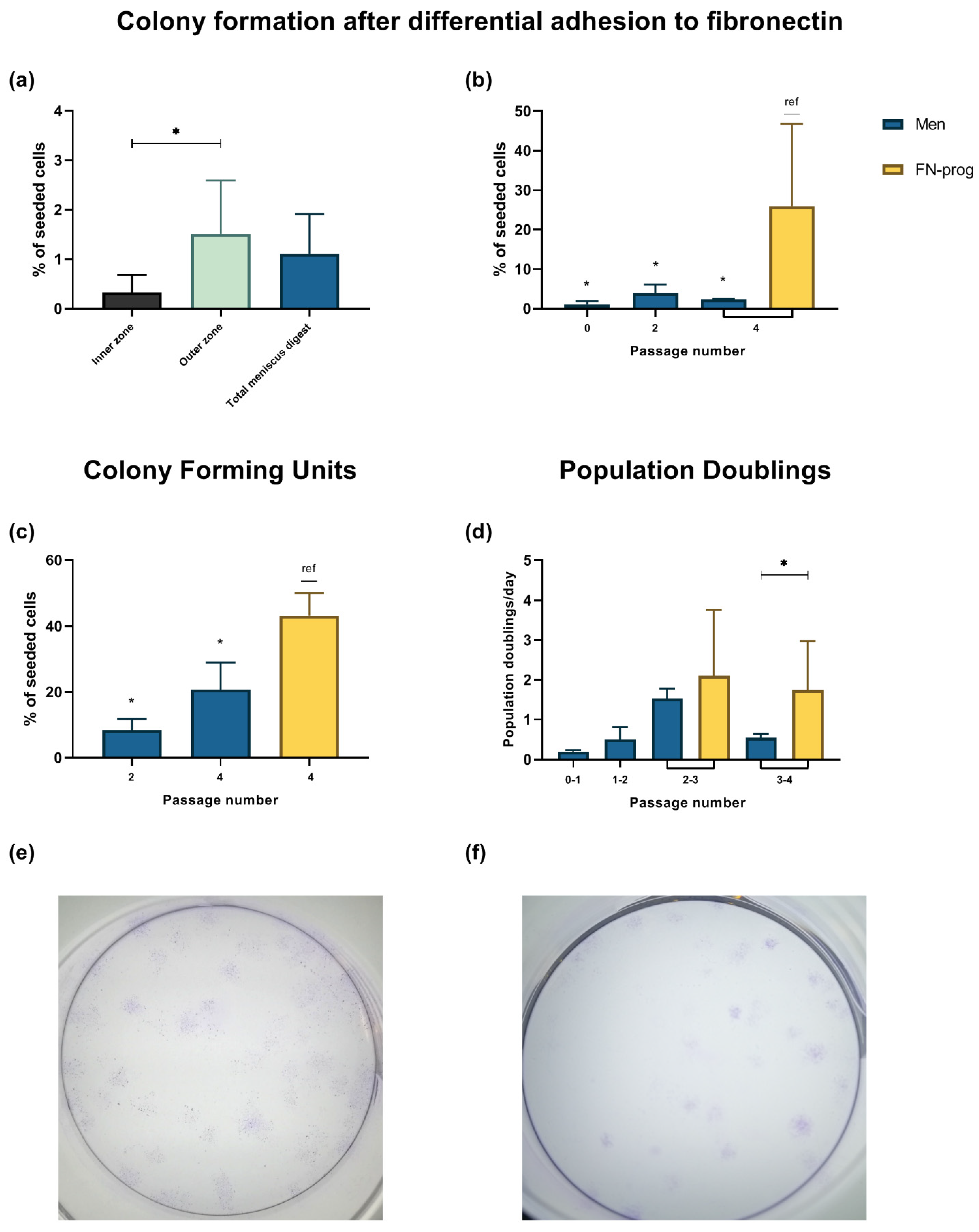
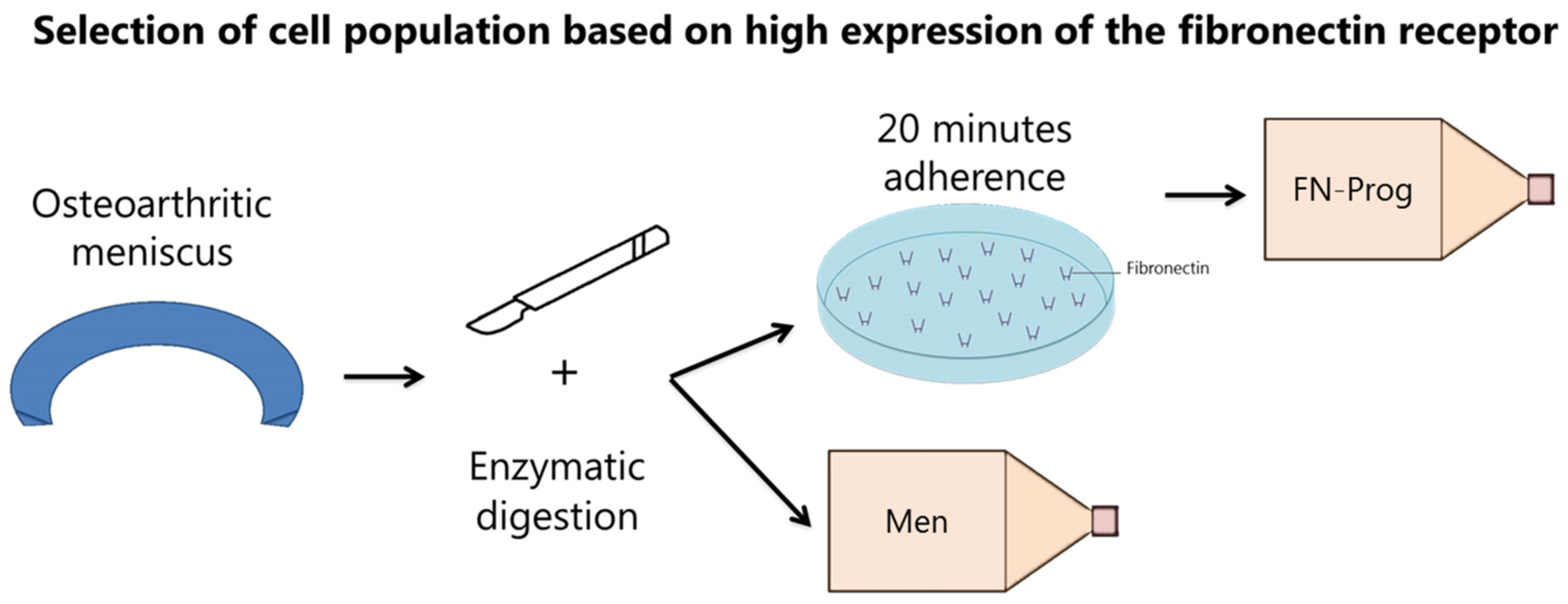
2.1. Selection of a Clonogenic Population from the Meniscus Inner and Outer Zone
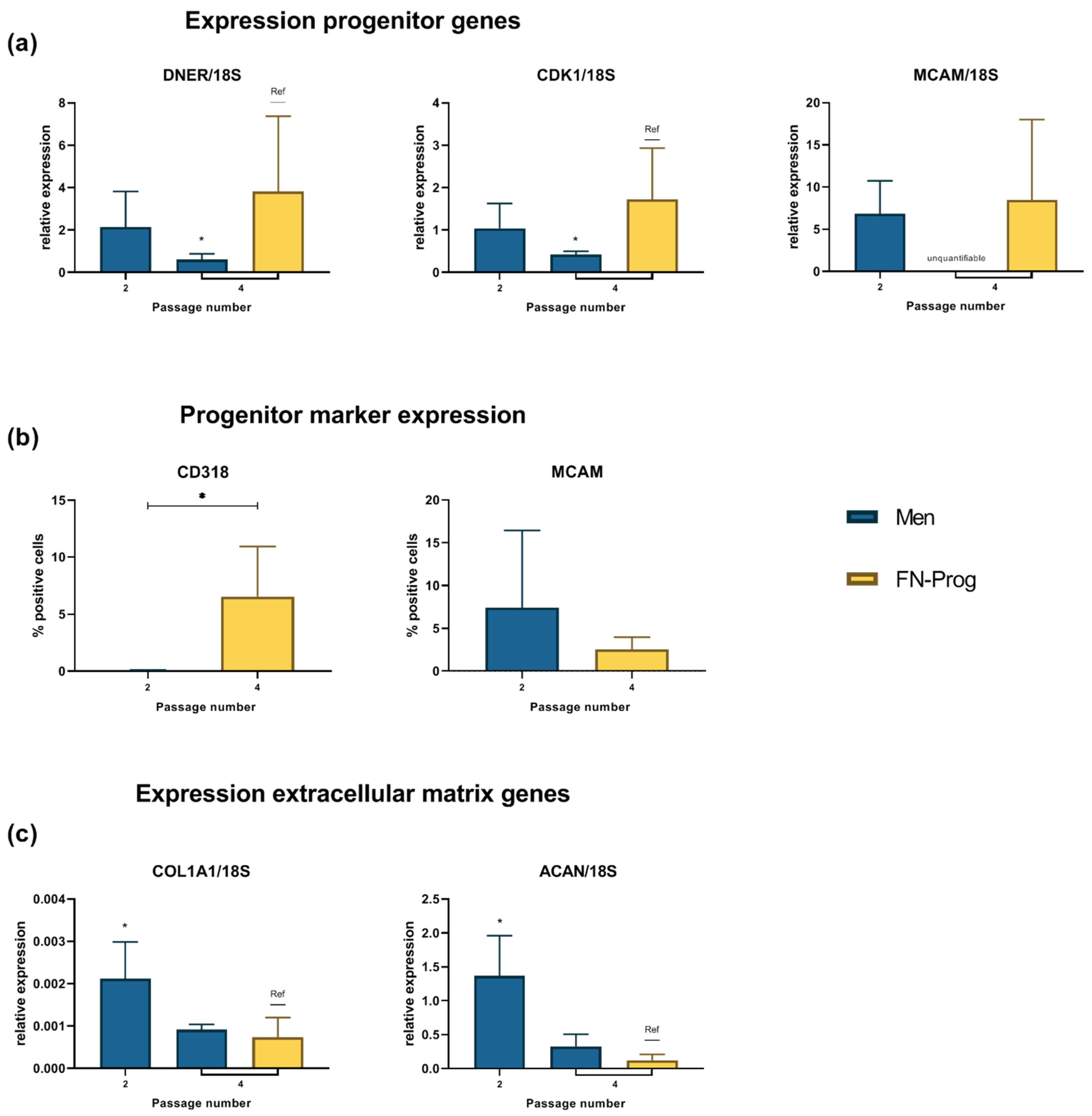
2.2. Expression of MSC Markers
2.3. Multilineage Potential
2.4. Expression Profile after Monolayer Expansion
2.5. Chondropermissive/Redifferentiation Culture
3. Discussion
3.1. Limitations
3.2. Implications
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. Colony Formation
4.3. Multilineage Differentiation
4.4. Expression of MSC Markers
4.5. Chondropermissive Cultures/ Redifferentiation
4.6. Gene Expression
4.7. Release and Deposition of Glycosaminoglycans and Collagen
4.8. Histology and Immunohistochemistry
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDevitt, C.A.; Webber, R.J. The ultrastructure and biochemistry of meniscal cartilage. Clin. Orthop. Relat. Res. 1990, 252, 8–18. [Google Scholar] [CrossRef]
- Verdonk, P.C.M.; Forsyth, R.G.; Wang, J.; Almqvist, K.F.; Verdonk, R.; Veys, E.M.; Verbruggen, G. Characterisation of human knee meniscus cell phenotype. Osteoarthr. Cartil. 2005, 13, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Arnoczky, S.P.; Warren, R.F. Microvasculature of the human meniscus. Am. J. Sports Med. 1982, 10, 90–95. [Google Scholar] [CrossRef]
- King, D. The healing of semilunar cartilages. Clin. Orthop. Relat. Res. 1990, 252, 4–7. [Google Scholar] [CrossRef]
- Starke, C.; Kopf, S.; Petersen, W.; Becker, R.; Stärke, C.; Kopf, S.; Petersen, W.; Becker, R. Meniscal repair. Arthroscopy 2009, 25, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Espejo-Reina, A.; Aguilera, J.; Espejo-Reina, M.J.; Espejo-Reina, M.P.; Espejo-Baena, A. One-Third of Meniscal Tears Are Repairable: An Epidemiological Study Evaluating Meniscal Tear Patterns in Stable and Unstable Knees. Arthrosc. J. Arthrosc. Relat. Surg. 2019, 35, 857–863. [Google Scholar] [CrossRef]
- Xu, C.; Zhao, J. A meta-analysis comparing meniscal repair with meniscectomy in the treatment of meniscal tears: The more meniscus, the better outcome? Knee Surg. Sports Traumatol. Arthrosc. 2013, 23, 164–170. [Google Scholar] [CrossRef]
- Englund, M.; Roos, E.M.; Lohmander, L.S. Impact of type of meniscal tear on radiographic and symptomatic knee osteoarthritis: A sixteen-year followup of meniscectomy with matched controls. Arthritis Rheum. 2003, 48, 2178–2187. [Google Scholar] [CrossRef]
- Shimomura, K.; Hamamoto, S.; Hart, D.A.; Yoshikawa, H.; Nakamura, N. Meniscal repair and regeneration: Current strategies and future perspectives. J. Clin. Orthop. Trauma 2018, 9, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Korpershoek, J.V.; De Windt, T.S.; Hagmeijer, M.H.; Vonk, L.A.; Saris, D.B.F. Cell-Based Meniscus Repair and Regeneration: At the Brink of Clinical Translation? Orthop. J. Sports Med. 2017, 5, 232596711769013. [Google Scholar] [CrossRef]
- Mueller, M.B.; Tuan, R.S. Functional characterization of hypertrophy in chondrogenesis of human mesenchymal stem cells. Arthritis Rheum. 2008, 58, 1377–1388. [Google Scholar] [CrossRef]
- Ding, Z.; Huang, H. Mesenchymal stem cells in rabbit meniscus and bone marrow exhibit a similar feature but a heterogeneous multi-differentiation potential: Superiority of meniscus as a cell source for meniscus repair Evolutionary developmental biology and morphology. BMC Musculoskelet. Disord. 2015, 16, 65. [Google Scholar] [CrossRef] [PubMed]
- Vangsness, C.T.; Farr, J.; Boyd, J.; Dellaero, D.T.; Mills, C.R.; LeRoux-Williams, M. Adult human mesenchymal stem cells delivered via intra-articular injection to the knee following partial medial meniscectomy A Randomized, Double-Blind, Controlled Study. J. Bone Jt. Surg. Ser. A 2014, 96, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal stem cells: Time to change the name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef]
- Alsalameh, S.; Amin, R.; Gemba, T.; Lotz, M. Identification of Mesenchymal Progenitor Cells in Normal and Osteoarthritic Human Articular Cartilage. Arthritis Rheum. 2004, 50, 1522–1532. [Google Scholar] [CrossRef]
- Williams, R.; Khan, I.M.; Richardson, K.; Nelson, L.; McCarthy, H.E.; Analbelsi, T.; Singhrao, S.K.; Dowthwaite, G.P.; Jones, R.E.; Baird, D.M.; et al. Identification and clonal characterisation of a progenitor cell sub-population in normal human articular cartilage. PLoS ONE 2010, 5, e13246. [Google Scholar] [CrossRef] [PubMed]
- Dowthwaite, G.P.; Bishop, J.C.; Redman, S.N.; Khan, I.M.; Rooney, P.; Evans, D.J.R.; Haughton, L.; Bayram, Z.; Boyer, S.; Thomson, B.; et al. The surface of articular cartilage contains a progenitor cell populations. J. Cell Sci. 2004, 117, 889–897. [Google Scholar] [CrossRef]
- Barbero, A.; Ploegert, S.; Heberer, M.; Martin, I. Plasticity of clonal populations of dedifferentiated adult human articular chondrocytes. Arthritis Rheum. 2003, 48, 1315–1325. [Google Scholar] [CrossRef]
- Jiang, Y.; Cai, Y.; Zhang, W.; Hu, C.; Tong, T.; Lu, P.; Zhang, S.; Neculai, D.; Tuan, R.S.; Ouyang, H.W.; et al. Human Cartilage-Derived Progenitor Cells From Committed Chondrocytes for Efficient Cartilage Repair and Regeneration. Stem Cells Transl. Med. 2016, 5, 733–744. [Google Scholar] [CrossRef]
- McCarthy, H.E.; Bara, J.J.; Brakspear, K.; Singhrao, S.K.; Archer, C.W. The comparison of equine articular cartilage progenitor cells and bone marrow-derived stromal cells as potential cell sources for cartilage repair in the horse. Vet. J. 2012, 192, 345–351. [Google Scholar] [CrossRef]
- Huang, H.; Wang, S.; Gui, J.; Shen, H. A study to identify and characterize the stem/progenitor cell in rabbit meniscus. Cytotechnology 2016, 68, 2083–2103. [Google Scholar] [CrossRef][Green Version]
- Sun, H.; Wen, X.; Li, H.; Wu, P.; Gu, M.; Zhao, X.; Zhang, Z.; Hu, S.; Mao, G.; Ma, R.; et al. Single-cell RNA-seq analysis identifies meniscus progenitors and reveals the progression of meniscus degeneration. Ann. Rheum. Dis. 2019, 79, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Chahla, J.; Papalamprou, A.; Chan, V.; Arabi, Y.; Salehi, K.; Nelson, T.J.; Limpisvasti, O.; Mandelbaum, B.R.; Tawackoli, W.; Metzger, M.F.; et al. Assessing the Resident Progenitor Cell Population and the Vascularity of the Adult Human Meniscus. Arthrosc. J. Arthrosc. Relat. Surg. 2021, 37, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, H.; Schminke, B.; Bode, C.; Roth, M.; Albert, J.; von der Heyde, S.; Rosen, V.; Miosge, N. Human migratory meniscus progenitor cells are controlled via the TGF-(beta) pathway. Stem Cell Rep. 2014, 3, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Crisan, M.; Corselli, M.; Chen, W.C.W.; Péault, B. Perivascular cells for regenerative medicine. J. Cell. Mol. Med. 2012, 16, 2851–2860. [Google Scholar] [CrossRef] [PubMed]
- Osawa, A.; Harner, C.D.; Gharaibeh, B.; Matsumoto, T.; Mifune, Y.; Kopf, S.; Ingham, S.J.M.; Schreiber, V.; Usas, A.; Huard, J. The use of blood vessel-derived stem cells for meniscal regeneration and repair. Med. Sci. Sports Exerc. 2013, 45, 813–823. [Google Scholar] [CrossRef]
- Seol, D.; Zhou, C.; Brouillette, M.J.; Song, I.; Yu, Y.; Choe, H.H.; Lehman, A.D.; Jang, K.W.; Fredericks, D.C.; Laughlin, B.J.; et al. Characteristics of meniscus progenitor cells migrated from injured meniscus. J. Orthop. Res. 2017, 35, 1966–1972. [Google Scholar] [CrossRef][Green Version]
- Shen, W.; Chen, J.; Zhu, T.; Chen, L.; Zhang, W.; Fang, Z.; Heng, B.C.; Yin, Z.; Chen, X.; Junfeng, J.; et al. Intra-articular injection of human meniscus stem/progenitor cells promotes meniscus regeneration and ameliorates osteoarthritis through stromal cell-derived factor-1/CXCR4-mediated homing. Stem Cells Transl. Med. 2014, 3, 387–394. [Google Scholar] [CrossRef]
- Segawa, Y.; Muneta, T.; Makino, H.; Nimura, A.; Mochizuki, T.; Ju, Y.J.; Ezura, Y.; Umezawa, A.; Sekiya, I. Mesenchymal stem cells derived from synovium, meniscus, anterior cruciate ligament, and articular chondrocytes share similar gene expression profiles. J. Orthop. Res. 2009, 27, 435–441. [Google Scholar] [CrossRef]
- Gamer, L.W.; Shi, R.R.; Gendelman, A.; Mathewson, D.; Gamer, J.; Rosen, V. Identification and characterization of adult mouse meniscus stem/progenitor cells. Connect. Tissue Res. 2017, 35, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Pelttari, K.; Winter, A.; Steck, E.; Goetzke, K.; Hennig, T.; Ochs, B.G.; Aigner, T.; Richter, W. Premature induction of hypertrophy during in vitro chondrogenesis of human mesenchymal stem cells correlates with calcification and vascular invasion after ectopic transplantation in SCID mice. Arthritis Rheum. 2006, 54, 3254–3266. [Google Scholar] [CrossRef] [PubMed]
- Tallheden, T.; Dennis, J.E.; Lennon, D.P.; Sjogren-Jansson, E.; Caplan, A.I.; Lindahl, A. Phenotypic Plasticity of Human Articular Chondrocytes. J. Bone Jt. Surg. 2003, 85 (Suppl. 2), 93–99. [Google Scholar] [CrossRef]
- Pauli, C.; Grogan, S.P.; Patil, S.; Otsuki, S.; Hasegawa, A.; Koziol, J.; Lotz, M.K. Macroscopic and histopathologic analysis of human knee menisci in aging and osteoarthritis. Osteoarthr. Cartil. 2011, 19, 1132–1141. [Google Scholar] [CrossRef]
- Zha, K.; Li, X.; Yang, Z.; Tian, G.; Sun, Z.; Sui, X.; Dai, Y.; Liu, S.; Guo, Q. Heterogeneity of mesenchymal stem cells in cartilage regeneration: From characterization to application. NPJ Regen. Med. 2021, 6, 14. [Google Scholar] [CrossRef]
- Grau-vorster, M.; Laitinen, A.; Nystedt, J.; Vives, J. HLA-DR expression in clinical-grade bone marrow-derived multipotent mesenchymal stromal cells: A two-site study. Stem Cell Res. Ther. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Mendicino, M.; Bailey, A.M.; Wonnacott, K.; Puri, R.K.; Bauer, S.R. MSC-based product characterization for clinical trials: An FDA perspective. Cell Stem Cell 2014, 14, 141–145. [Google Scholar] [CrossRef]
- Polchert, D.; Sobinsky, J.; Douglas, G.W.; Kidd, M.; Moadsiri, A.; Reina, E.; Genrich, K.; Mehrotra, S.; Setty, S.; Smith, B.; et al. IFN-γ activation of mesenchymal stem cells for treatment and prevention of graft versus host disease. Eur. J. Immunol. 2008, 38, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Dighe, P.A.; Viswanathan, P.; Mruthunjaya, A.K.; Seetharam, R.N. Effect of bFGF on HLA-DR expression of human bone marrow-derived mesenchymal stem cells. J. Stem Cells 2013, 8, 43–57. [Google Scholar]
- Li, Y.; Zhou, J.; Yang, X.; Jiang, Y.; Gui, J. Intermittent hydrostatic pressure maintains and enhances the chondrogenic differentiation of cartilage progenitor cells cultivated in alginate beads. Dev. Growth Differ. 2016, 58, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Ustunel, I.; Ozenci, A.M.; Sahin, Z.; Ozbey, O.; Acar, N.; Tanriover, G.; Celik-Ozenci, C.; Demir, R. The immunohistochemical localization of notch receptors and ligands in human articular cartilage, chondroprogenitor culture and ultrastructural characteristics of these progenitor cells. Acta Histochem. 2008, 110, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Vinod, E.; Kachroo, U.; Rebekah, G.; Yadav, B.K.; Ramasamy, B. Characterization of human articular chondrocytes and chondroprogenitors derived from non-diseased and osteoarthritic knee joints to assess superiority for cell-based therapy. Acta Histochem. 2020, 122, 151588. [Google Scholar] [CrossRef]
- Kachroo, U.; Ramasamy, B.; Vinod, E. Evaluation of CD49e as a distinguishing marker for human articular cartilage derived chondroprogenitors. Knee 2020, 27, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Kalnievl, M.; Krystev, D. Immunohistochemical Study of the Distribution of Fibronectin in Some Zones of the Meniscus. 101–106. Available online: https://www.researchgate.net/profile/Nikolay-Dimitrov-11/publication/283121594_Normal_morphology_of_biologically_active_point_BAPST36_rat/links/562ba3de08aef25a2441ca03/Normal-morphology-of-biologically-active-point-BAP-ST36-rat.pdf#page=103 (accessed on 9 August 2021).
- Bekkers, J.E.J.; Saris, D.B.F.; Tsuchida, A.I.; Van Rijen, M.H.P.; Dhert, W.J.A.; Creemers, L.B. Chondrogenic potential of articular chondrocytes depends on their original location. Tissue Eng. Part A 2014, 20, 663–671. [Google Scholar] [CrossRef]
- Federa Human Tissue and Medical Research: Code of Conduct for Responsible Use. Available online: https://www.federa.org/code-goed-gebruik (accessed on 29 April 2020).
- Van Diest, P.J. No consent should be needed for using leftover body material for scientific purposes. BMJ 2002, 325, 648–651. [Google Scholar] [CrossRef] [PubMed]
- Neuman, E.R.; Logan, A.M. The determination of hydroxyproline. J. Biol. Chem. 1950, 184, 299–306. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Oligonucleotide Sequence (5′ to 3′) | Product Size (bp) |
|---|---|---|
| 18S | Fw: GTAACCCGTTGAACCCCATT Rv: CCATCCAATCGGTAGTAGCG | 151 |
| ACAN | Fw: CAACTACCCGGCCATCC Rv: GATGGCTCTGTAATGGAACAC | 160 |
| COL2A1 | Fw: AGGGCCAGGATGTCCGGCA Rv: GGGTCCCAGGTTCTCCATCT | 195 |
| COL1A1 | Fw: TCCAACGAGATCGAGATCC Rv: AAGCCGAATTCCTGGTCT | 191 |
| DNER | Fw: AAGGCTATGAAGGTCCCAACT Rv: CTGAGAGCGAGGCAGGATTT | 137 |
| MCAM | Fw: AGCTCCGCGTCTACAAAGC Rv: CTACACAGGTAGCGACCTCC | 102 |
| CDK1 | Fw: AAACTACAGGTCAAGTGGTAGCC Rv: TCCTGCATAAGCACATCCTGA | 148 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korpershoek, J.V.; Rikkers, M.; de Windt, T.S.; Tryfonidou, M.A.; Saris, D.B.F.; Vonk, L.A. Selection of Highly Proliferative and Multipotent Meniscus Progenitors through Differential Adhesion to Fibronectin: A Novel Approach in Meniscus Tissue Engineering. Int. J. Mol. Sci. 2021, 22, 8614. https://doi.org/10.3390/ijms22168614
Korpershoek JV, Rikkers M, de Windt TS, Tryfonidou MA, Saris DBF, Vonk LA. Selection of Highly Proliferative and Multipotent Meniscus Progenitors through Differential Adhesion to Fibronectin: A Novel Approach in Meniscus Tissue Engineering. International Journal of Molecular Sciences. 2021; 22(16):8614. https://doi.org/10.3390/ijms22168614
Chicago/Turabian StyleKorpershoek, Jasmijn V., Margot Rikkers, Tommy S. de Windt, Marianna A. Tryfonidou, Daniel B. F. Saris, and Lucienne A. Vonk. 2021. "Selection of Highly Proliferative and Multipotent Meniscus Progenitors through Differential Adhesion to Fibronectin: A Novel Approach in Meniscus Tissue Engineering" International Journal of Molecular Sciences 22, no. 16: 8614. https://doi.org/10.3390/ijms22168614
APA StyleKorpershoek, J. V., Rikkers, M., de Windt, T. S., Tryfonidou, M. A., Saris, D. B. F., & Vonk, L. A. (2021). Selection of Highly Proliferative and Multipotent Meniscus Progenitors through Differential Adhesion to Fibronectin: A Novel Approach in Meniscus Tissue Engineering. International Journal of Molecular Sciences, 22(16), 8614. https://doi.org/10.3390/ijms22168614