
Overexpression of ERAP2N in Human Trophoblast Cells Promotes Cell Death

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Sample Identification
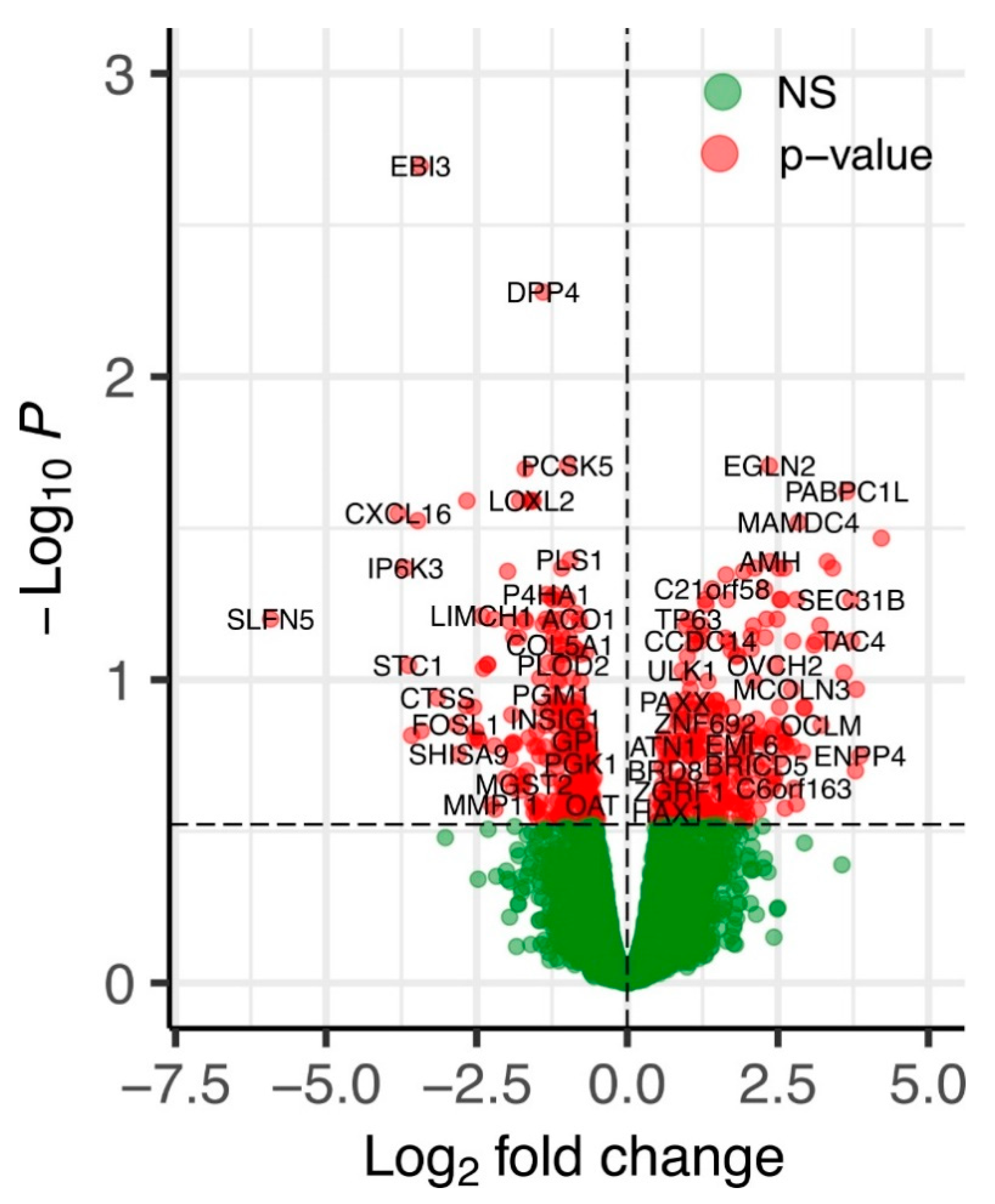
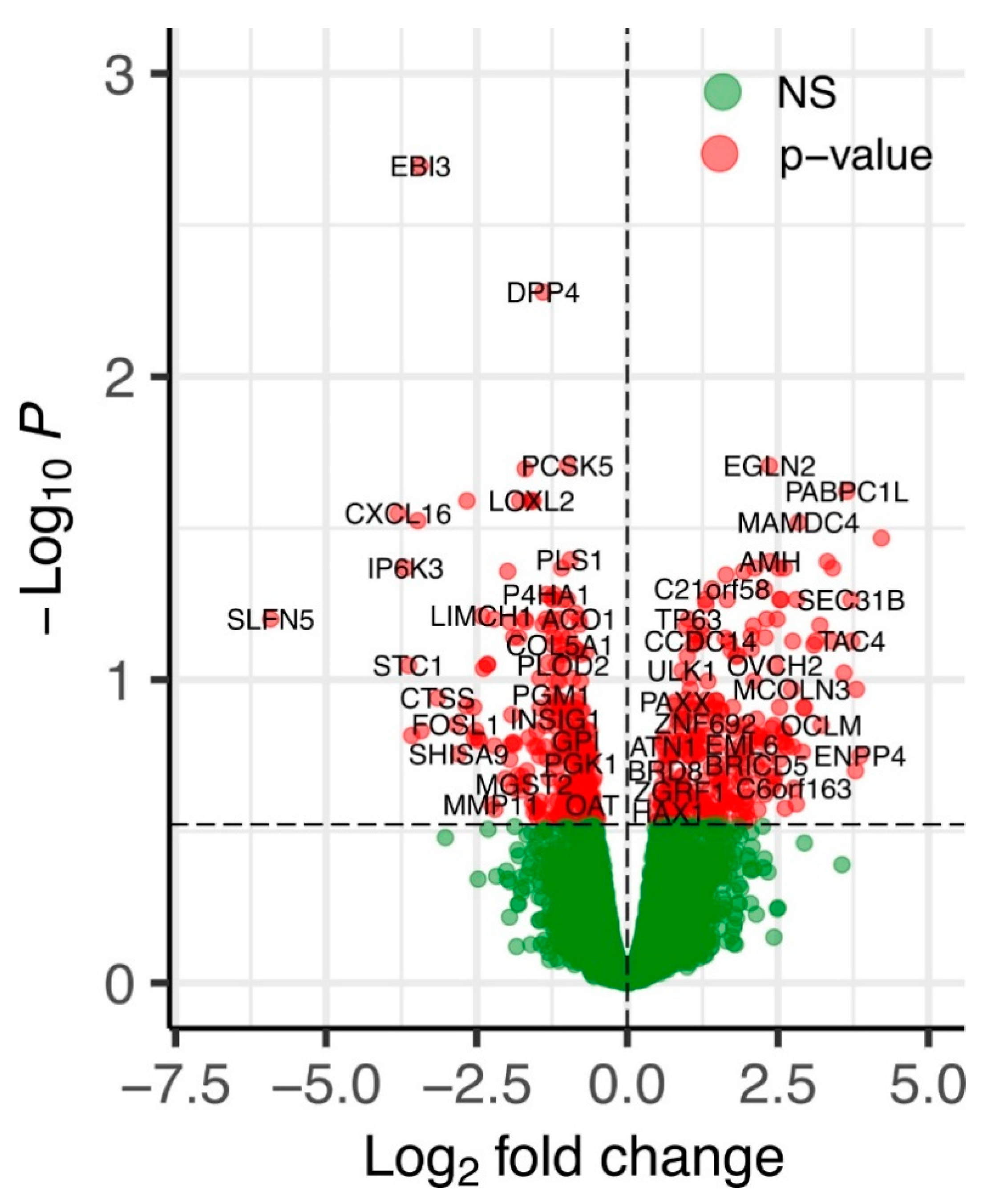
2.2. An Overview of Differentially Expressed Genes in ERAP2N-Positive vs. ERAP2N-Negative Trophoblast Cells
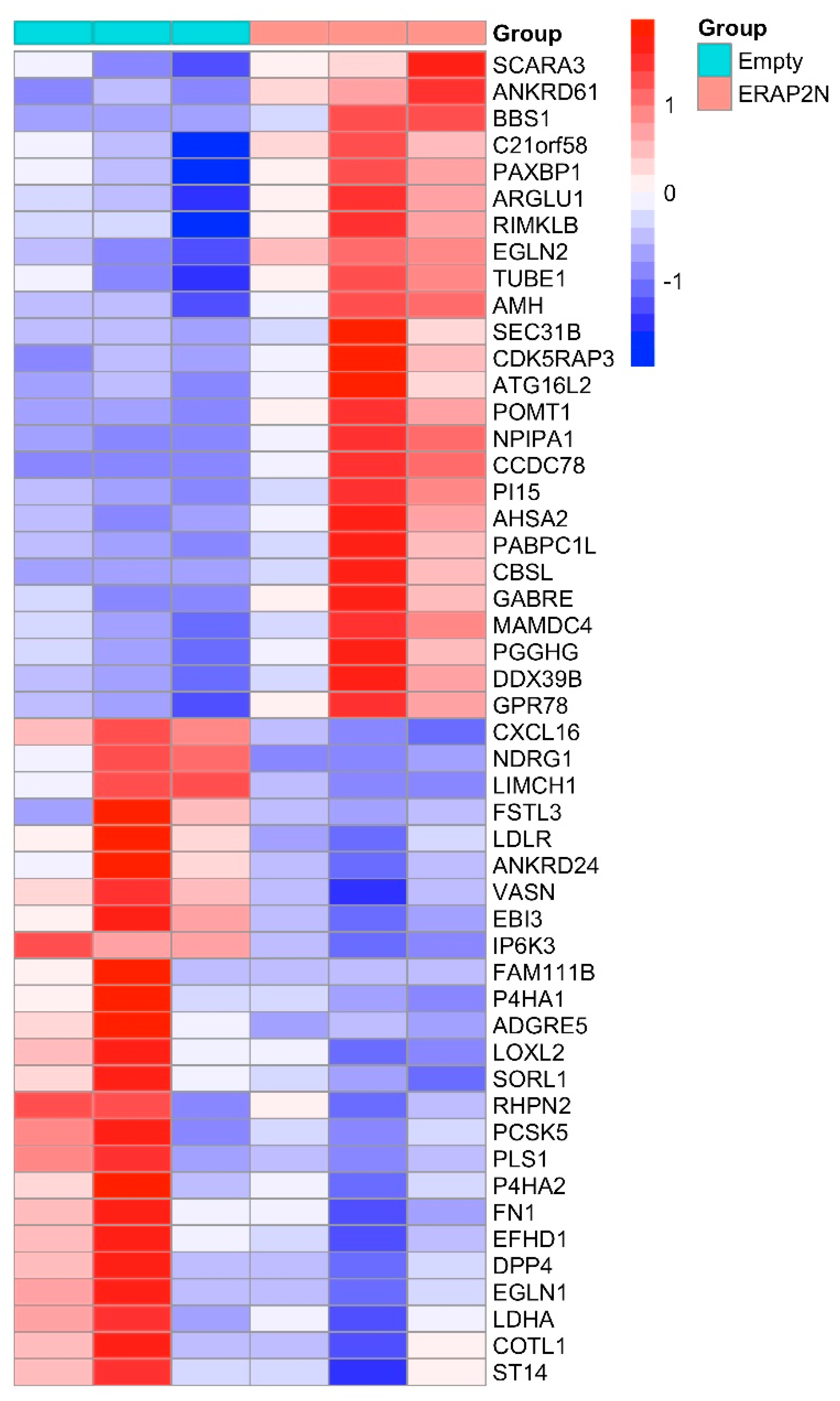
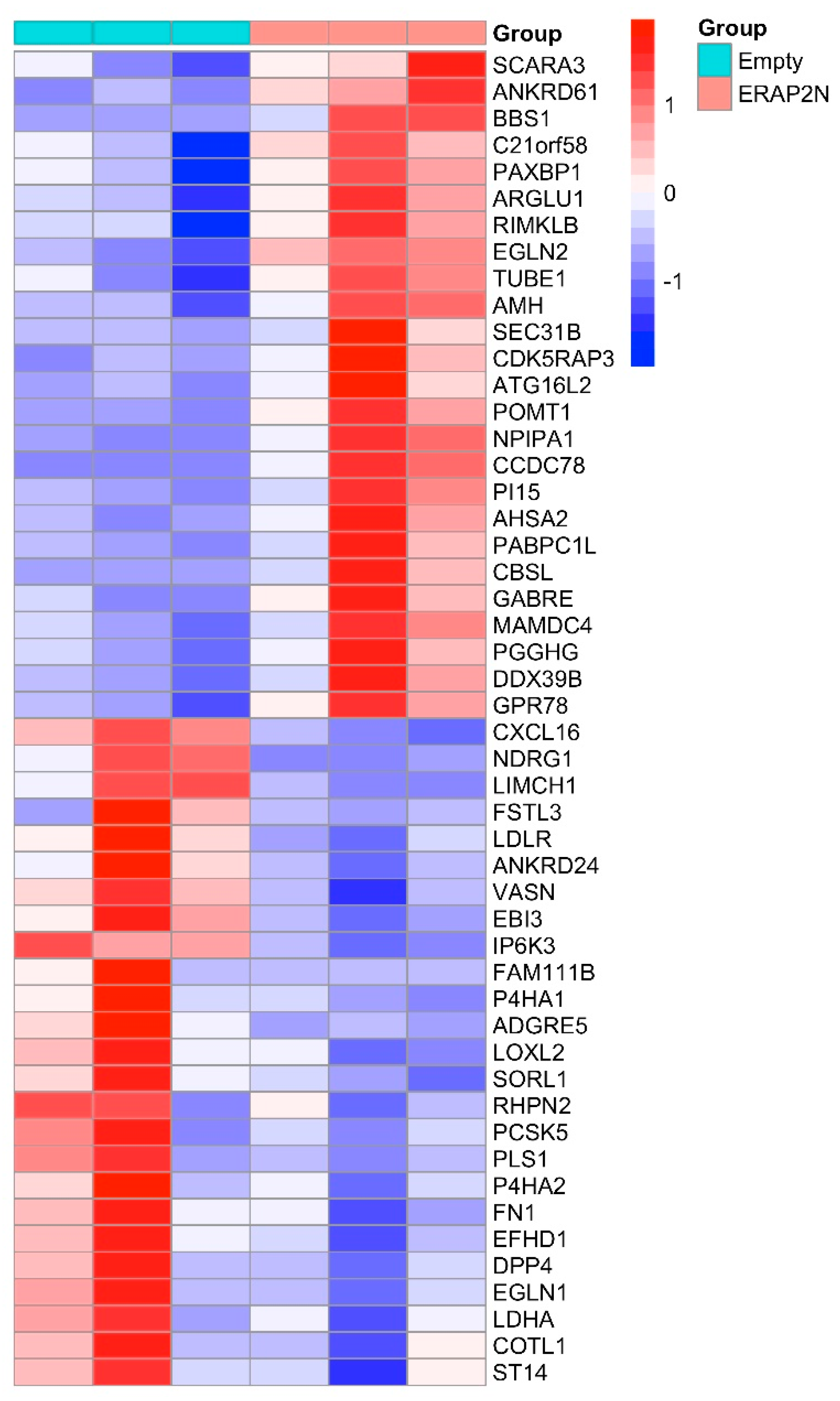
2.3. The Heatmap of the Top 50 Most Significantly Differentially Expressed Genes in ERAP2N-Positive vs. ERAP2N-Negative Trophoblast Cells
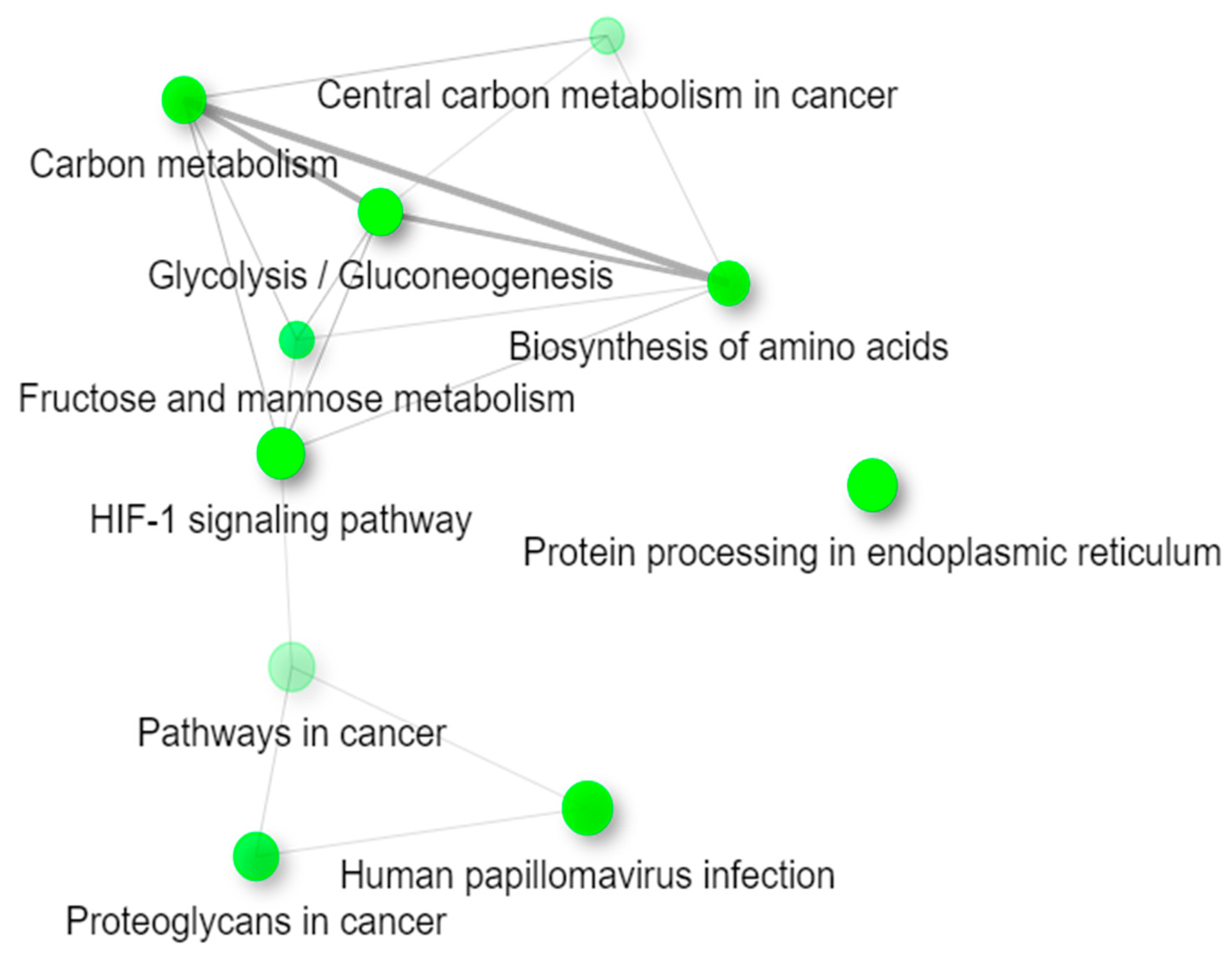
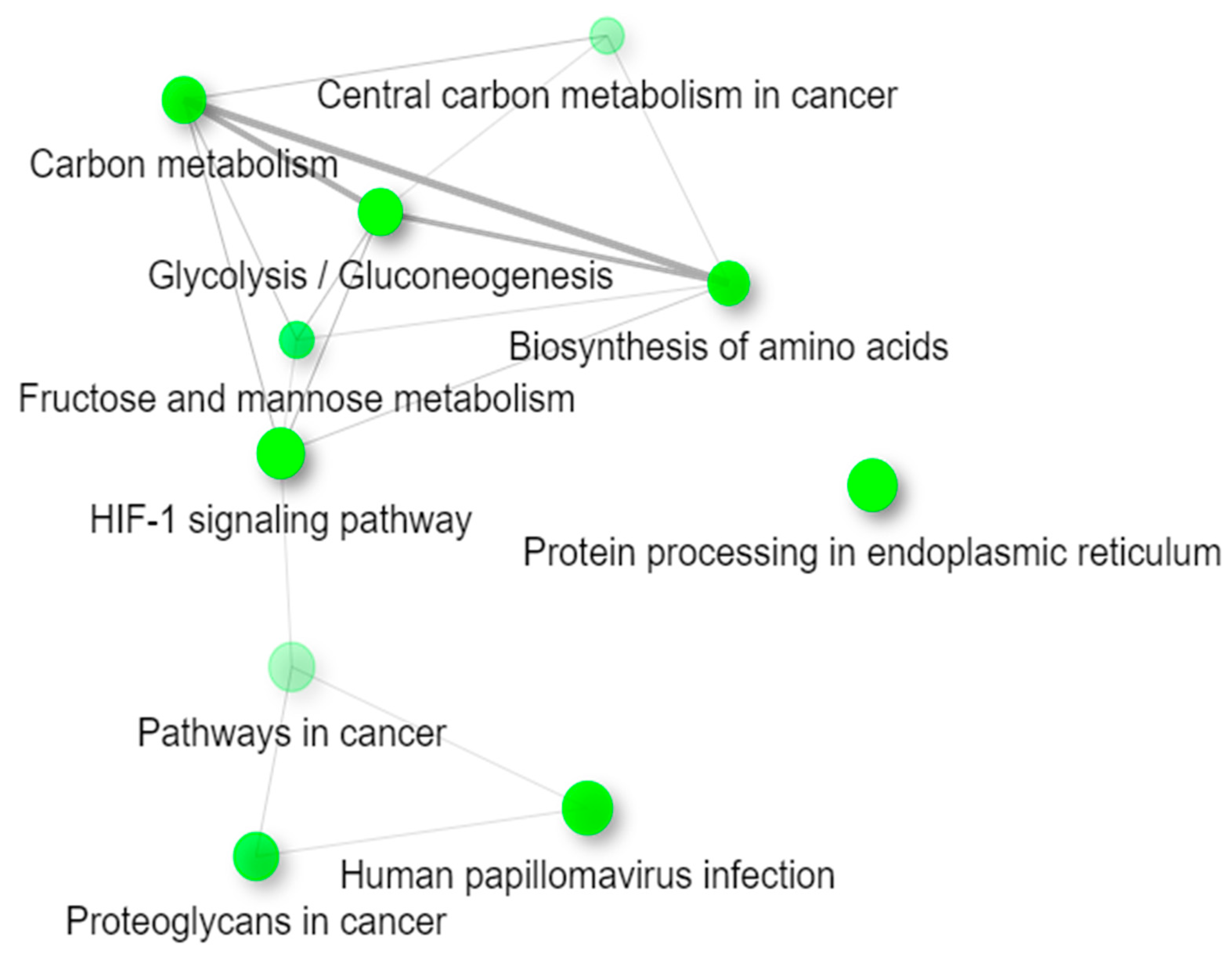
2.4. Functional Enrichment Analysis of KEGG Pathways
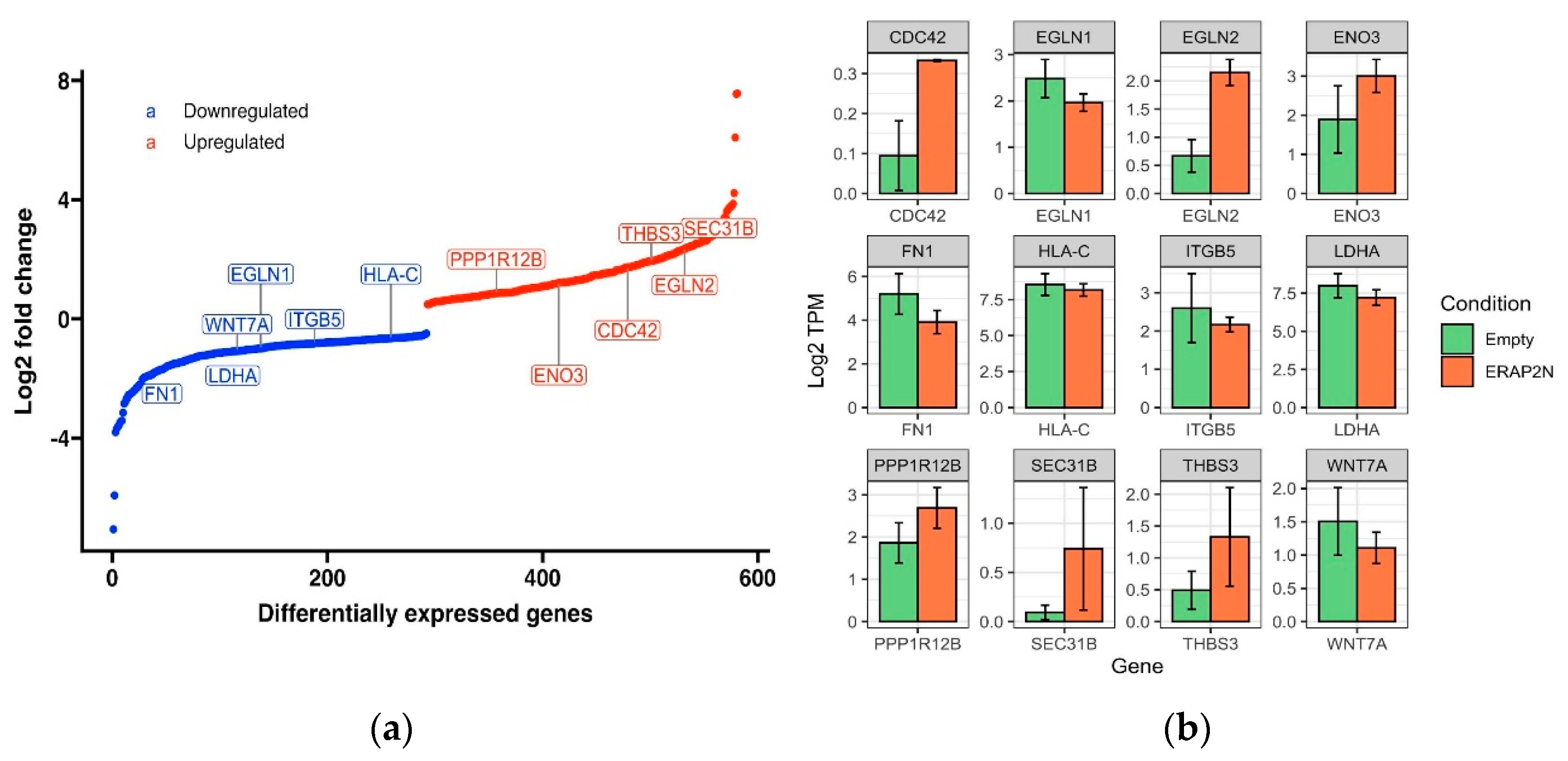
2.5. Differentially Expressed Genes (DEGs) in ERAP2N-Positive Cells Compared to ERAP2N-Negative Cells
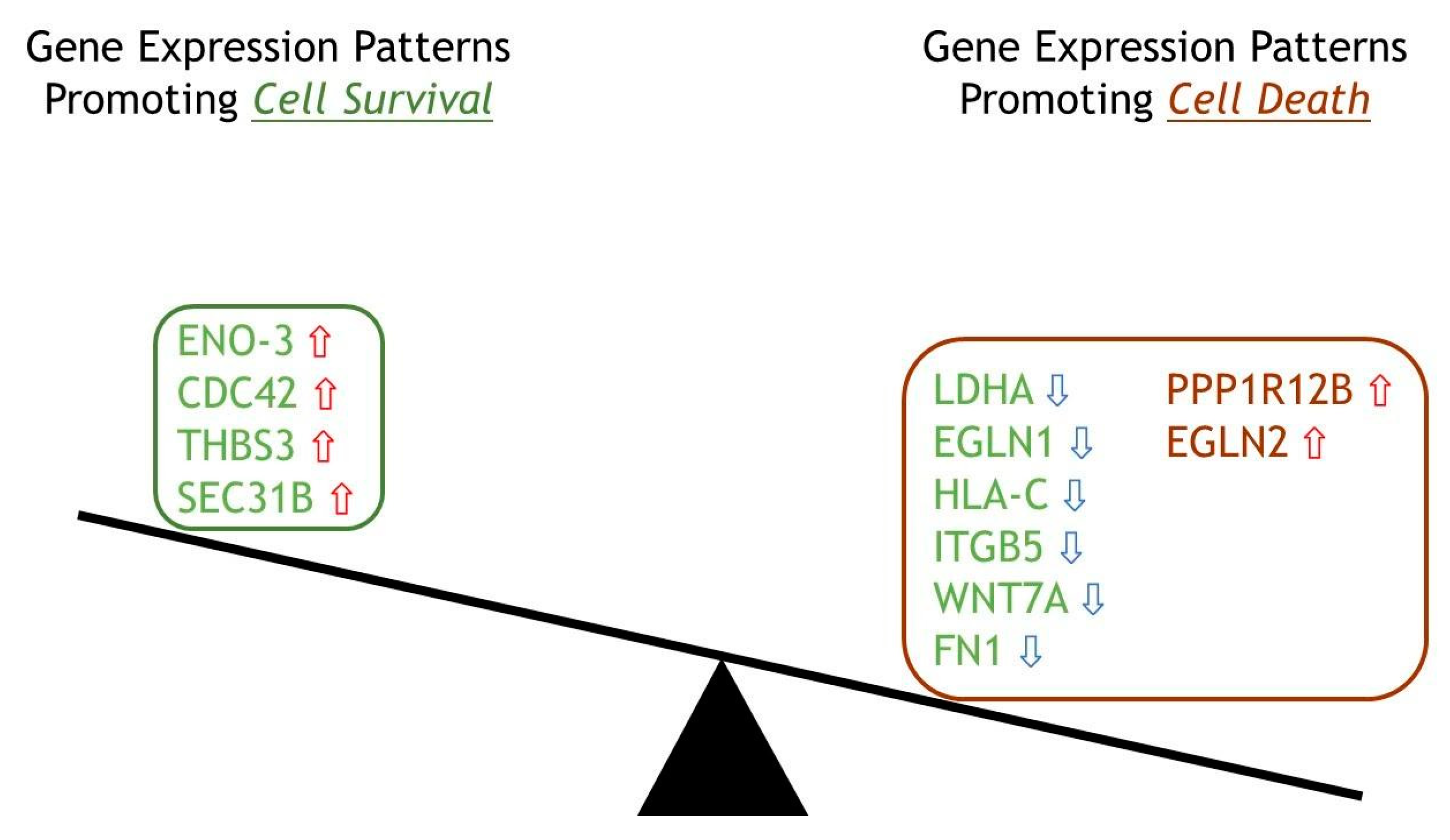
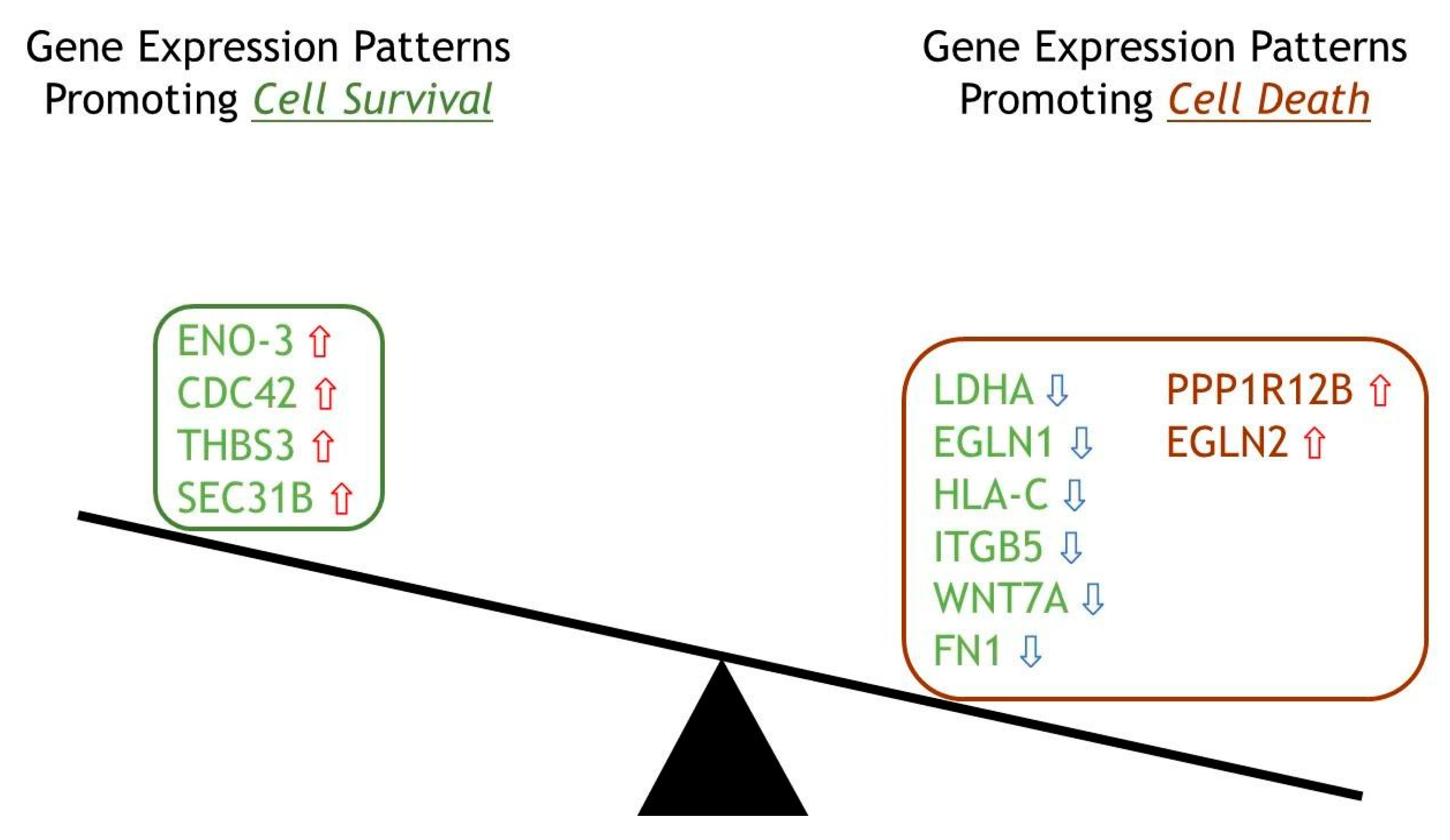
2.6. Top 12 Genes in Cellular Fate Expressed in ERAP2N-Positive Cells
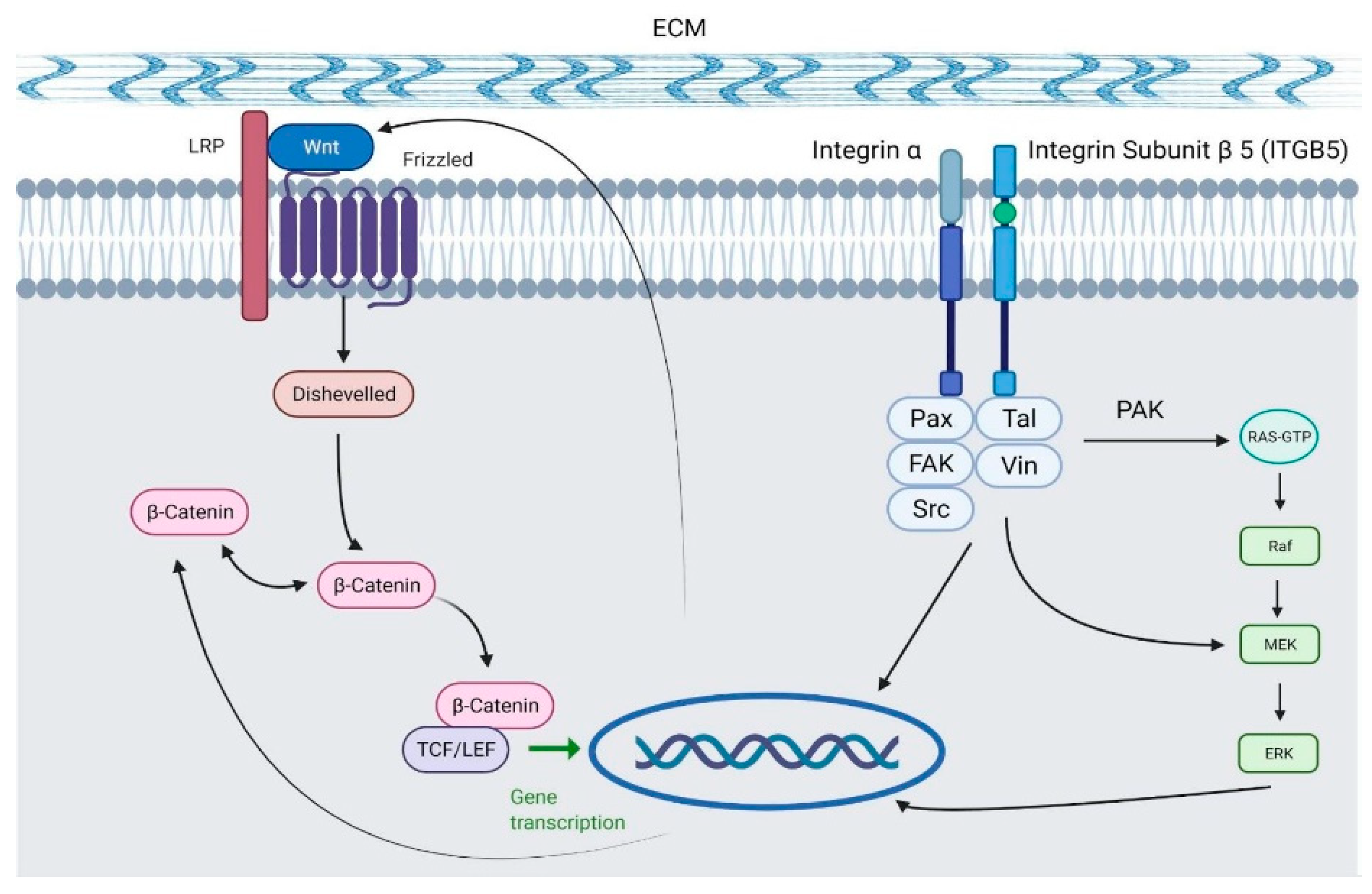
3. Discussion
4. Materials and Methods
4.1. Cell Lines and pcDNA Stable Transfection of JEG-3 Cells
4.2. RNA Sample Collection and Preparation
4.3. Clustering and RNA Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burton, G.J.; Jauniaux, E. Development of the human placenta and fetal heart: Synergic or independent? Front. Physiol. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Human Placenta Project: How Does the Placenta Form? Placent. Dev. Fertil. Full Term 2019, 2–3.
- Kim, S.-M.; Kim, J.-S. A Review of Mechanisms of Implantation. Dev. Reprod. 2017, 21, 351–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Than, N.G.; Hahn, S.; Rossi, S.W.; Szekeres-Bartho, J. Editorial: Fetal-Maternal Immune Interactions in Pregnancy. Front. Immunol. 2019, 10, 1–4. [Google Scholar] [CrossRef]
- Ehrentraut, S.; Sauss, K.; Neumeister, R.; Luley, L.; Oettel, A.; Fettke, F.; Costa, S.-D.; Langwisch, S.; Zenclussen, A.; Schumacher, A.; et al. Human Miscarriage Is Associated with Dysregulations in Peripheral Blood-Derived Myeloid Dendritic Cell Subsets. Front. Immunol. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shmeleva, E.V.; Colucci, F. Maternal natural killer cells at the intersection between reproduction and mucosal immunity. Mucosal Immunol. 2021, 1–15. [Google Scholar] [CrossRef]
- Fiorillo, M.T.; van Endert, P.M.; Bouvier, M.; López De Castro, J.A. How ERAP1 and ERAP2 Shape the Peptidomes of Disease-Associated MHC-I Proteins. Front. Immunol. 2018, 9, 2463. [Google Scholar]
- Hill, L.D.; Hilliard, D.D.; York, T.P.; Srinivas, S.; Kusanovic, J.P.; Gomez, R.; Elovitz, M.A.; Romero, R.; Strauss, J.F. Fetal ERAP2 Variation is Associated with Preeclampsia in African Americans in A Case-Control Study. BMC Med. Genet. 2011. Available online: http://www.biomedcentral.com/1471-2350/12/64 (accessed on 1 July 2021). [CrossRef] [Green Version]
- Johnson, M.P.; Roten, L.T.; Dyer, T.D.; East, C.; Forsmo, S.; Blangero, J.; Brennecke, S.; Austgulen, R.; Moses, E. The ERAP2 gene is associated with preeclampsia in Australian and Norwegian populations. Hum. Genet. 2009, 126, 655–666. [Google Scholar] [CrossRef] [Green Version]
- Vanhille, D.L.; Hill, L.D.; Hilliard, D.D.; Lee, E.D.; Teves, M.E.; Srinivas, S.; Kusanovic, J.P.; Gómez, R.; Stratikos, E.; Elovitz, M.; et al. A novel ERAP2 haplotype structure in a Chilean population: Implications for ERAP2 protein expression and preeclampsia risk. Mol. Genet. Genom. Med. 2013, 1, 98–107. [Google Scholar] [CrossRef]
- Evnouchidou, I.; Birtley, J.; Seregin, S.; Papakyriakou, A.; Zervoudi, E.; Samiotaki, M.; Panayotou, G.; Giastas, P.; Petrakis, O.; Georgiadis, D.; et al. Altered Antigen Processing Induces a Specificity Switch That Leads to in Endoplasmic Reticulum Aminopeptidase 2 A Common Single Nucleotide Polymorphism. J. Immunol. 2021, 189, 2383–2392. [Google Scholar] [CrossRef]
- Lee, E.D. Endoplasmic Reticulum Aminopeptidase 2, a common immunological link to adverse pregnancy outcomes and cancer clearance? Placenta 2017, 56, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Andrés, A.M.; Dennis, M.Y.; Kretzschmar, W.W.; Cannons, J.L.; Lee-Lin, S.-Q. Balancing Selection Maintains a Form of ERAP2 that Undergoes Nonsense-Mediated Decay and Affects Antigen Presentation. PLoS Genet. 2010, 6, e1001157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plácido, A.; Pereira, C.M.F.; Duarte, A.; Candeias, E.; Correia, S.; dos Santos, R.X.C.; Carvalho, C.; Cardoso, S.M.; Oliveira, C.; Moreira, P.; et al. The role of endoplasmic reticulum in amyloid precursor protein processing and trafficking: Implications for Alzheimer’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 1444–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sa, S.; Wong, L.; Mccloskey, K.E. Combinatorial Fibronectin and Laminin Signaling Promote Highly Efficient Cardiac Differentiation of Human Embryonic Stem Cells. BioRes. Open Access 2014, 3, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Warthan, M.D.; Washington, S.L.; Franzese, S.E.; Ramus, R.M.; Kim, K.-R.; York, T.P.; Stratikos, E.; Strauss, J.F.; Lee, E.D. The role of endoplasmic reticulum aminopeptidase 2 in modulating immune detection of choriocarcinoma. Biol. Reprod. 2018, 98, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Dunlap, K.A.; Filant, J.; Hayashi, K.; Rucker, E.B.; Song, G.; Deng, J.M.; Behringer, R.R.; DeMayo, F.J.; Lydon, J.; Jeong, J.-W.; et al. Postnatal Deletion of Wnt7a Inhibits Uterine Gland Morphogenesis and Compromises Adult Fertility in Mice 1. Biol. Reprod. 2011, 85, 386–396. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, S.; King, M.L.; Ran, S.; Okuda, H.; Ii, J.A.M.; McAsey, M.E.; Sugino, N.; Brard, L.; Watabe, K.; Hayashi, K. WNT7A Regulates Tumor Growth and Progression in Ovarian Cancer through the WNT/b-Catenin Pathway. Mol. Cancer Res. 2012, 10, 469–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haseeb, M.; Hassan Pirzada, R.; Ul Ain, Q.; Choi, S. Wnt Signaling in the Regulation of Immune Cell and Cancer Therapeutics. Cells 2019, 8, 1380. [Google Scholar] [CrossRef] [Green Version]
- Ding, C.; Tang, W.; Wu, H.; Fan, X.; Luo, J.; Feng, J.; Wen, K.; Wu, G. The PEAK1-PPP1R12B axis inhibits tumor growth and metastasis by regulating Grb2/PI3K/Akt signalling in colorectal cancer. Cancer Lett. 2019, 442, 383–395. [Google Scholar] [CrossRef]
- Price, C.; Gill, S.; Ho, Z.V.; Davidson, S.M.; Merkel, E.; McFarland, J.M.; Leung, L.; Tang, A.; Kost-Alimova, M.; Tsherniak, A.; et al. Molecular Cell Biology Genome-Wide Interrogation of Human Cancers Identifies EGLN1 Dependency in Clear Cell Ovarian Cancers. Cancer Res. 2019, 79, 2564–2579. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Liu, C.; Zhang, T.N.; Zhu, Y.W.; Dong, X.; Xue, P. Down-regulation of FN1 inhibits colorectal carcinogenesis by suppressing proliferation, migration, and invasion. J. Cell. Biochem. 2018, 119, 4717–4728. [Google Scholar] [CrossRef] [PubMed]
- Soikkeli, J.; Podlasz, P.; Yin, M.; Nummela, P.; Jahkola, T.; Virolainen, S.; Krogerus, L.; Heikkilä, P.; von Smitten, K.; Saksela, O.; et al. Metastatic outgrowth encompasses COL-I, FN1, and POSTN up-regulation and assembly to fibrillar networks regulating cell adhesion, migration, and growth. Am. J. Pathol. 2010, 177, 387–403. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Zhang, Y.; He, J.; Zang, Z.; Zhou, Z.; Pei, X.; Zheng, X.; Zhang, W.; Yang, H.; Li, S.; et al. Lactate Dehydrogenase A Promotes the Invasion and Proliferation of Pituitary Adenoma. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Xiong, Y.; Qiao, T.; Li, X.; Jia, L.; Han, Y. Lactate dehydrogenase A: A key player in carcinogenesis and potential target in cancer therapy. Cancer Med. 2018, 7, 6124–6136. [Google Scholar] [CrossRef] [Green Version]
- Contini, P.; Ghio, M.; Poggi, A.; Filaci, G.; Induveri, F.; Ferrone, S.; Puppo, F. Soluble HLA-A,-B,-C and-G Molecules Induce Apoptosis in T and NK CD8 + Cells and Inhibit Cytotoxic T Cell Activity through CD8 Ligation. Eur. J. Immunol. 2003, 33, 125–134. [Google Scholar] [CrossRef]
- Hiby, S.E.; Walker, J.; O’Shaughnessy, K.M.; Redman, C.W.; Carrington, M.; Trowsdale, J.; Moffett, A. Combinations of Maternal KIR and Fetal HLA-C Genes Influence the Risk of Preeclampsia and Reproductive Success. J. Exp. Med. 2004, 200, 957–965. [Google Scholar] [CrossRef]
- The Mechanisms of β-Catenin on Keloid Fibroblast Cells Proliferation and Apoptosis. Available online: https://www.semanticscholar.org/paper/The-mechanisms-of-%CE%B2-catenin-on-keloid-fibroblast-Chen-Yu/5387841c19eb188b07740da750976375dabcf5b3 (accessed on 1 July 2021).
- Zhang, L.Y.; Guo, Q.; Guan, G.F.; Cheng, W.; Cheng, P.; Wu, A.H. Integrin Beta 5 Is a Prognostic Biomarker and Potential Therapeutic Target in Glioblastoma. Front. Oncol. 2019, 9, 904. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Lee, Y.; Je, S.; Chang, S.; Kim, N.; Jeong, E.; Yoon, S. Molecules and Cells Overexpression and Selective Anticancer Efficacy of ENO3 in STK11 Mutant Lung Cancers. Mol. Cells 2019, 42, 804–809. [Google Scholar]
- Qadir, M.I.; Parveen, A.; Ali, M. Cdc42: Role in Cancer Management. Chem. Biol. Drug Des. 2015, 86, 432–439. [Google Scholar] [CrossRef]
- Dalla-Torre, C.A.; Yoshimoto, M.; Lee, C.H.; Joshua, A.M.; de Toledo, S.R.; Petrilli, A.S.; Andrade, J.A.; Chilton-MacNeill, S.; Zielenska, M.; Squire, J.A.; et al. Effects of THBS3, SPARC and SPP1 Expression on Biological Behavior and Survival in Patients with Osteosarcoma. BMC Cancer 2006, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Gourguechon, S.; Wang, C.C.; Li, Z. The G1 Cyclin-dependent Kinase CRK1 in Trypanosoma brucei Regulates Anterograde Protein Transport by Phosphorylating the COPII Subunit Sec31. J. Biol. Chem. 2016, 291, 15527–15539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, A.M.; Enders, A.C.; Pijnenborg, R. The role of invasive trophoblast in implantation and placentation of primates. Phil. Trans. R. Soc. B 2015, 370, 20140070. [Google Scholar] [CrossRef]
- Kiewisz, J.; Kaczmarek, M.M.; Andronowska, A.; Blitek, A.; Ziecik, A.J. Gene expression of WNTs, β-catenin and E-cadherin during the periimplantation period of pregnancy in pigs—Involvement of steroid hormones. Theriogenology 2011, 76, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Saben, J.; Zhong, Y.; McKelvey, S.; Dajani, N.K.; Andres, A.; Badger, T.M.; Gomez-Acevedo, H.; Shankar, K. A comprehensive analysis of the human placenta transcriptome. Placenta 2014, 35, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Mo, H.Q.; Tian, F.J.; Ma, X.L.; Zhang, Y.C.; Zhang, C.X.; Zeng, W.H.; Zhang, Y.; Lin, Y. PDIA3 regulates trophoblast apoptosis and proliferation in preeclampsia via the MDM2/p53 pathway. Reproduction 2020, 160, 293–305. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S. Sequence analysis STAR: Ultrafast universal RNA-seq aligner. Oxford Acad. 2013, 29, 15–21. [Google Scholar]
- Url, S.; Society, R.S. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing Yoav Benjamini; Yosef Hochberg. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. 2013. Available online: http://amp.pharm.mssm.edu/Enrichr (accessed on 1 July 2021).
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathways | p-Value | Adjusted p-Value | Genes |
|---|---|---|---|
| Glycolysis/Gluconeogenesis | 6.66 × 10−8 | 1.75 × 10−5 | GPI; TPI1; PGAM1; ENO1; ENO3; HK2; LDHA; PKM; PGK1; ALDOC; ALDOA; PGM1; PFKP |
| HIF-1 signaling pathway | 3.30 × 10−5 | 3.27 × 10−3 | EGLN1; LDHA; EGLN3; EGLN2; TFRC; STAT3; PGK1; SLC2A1; ENO1; ALDOA; ENO3; HK2 |
| Fructose and mannose metabolism | 3.73 × 10−5 | 3.27 × 10−3 | PFKFB4; TPI1; AKR1B1; ALDOC; ALDOA; HK2; PFKP |
| Protein processing in the endoplasmic reticulum | 9.74 × 10−5 | 6.41 × 10−3 | ERO1A; PDIA3; HSPA5; WFS1; RRBP1; CKAP4; DDOST; PDIA4; HSP90B1; OS9; CALR; P4HB; SEC24D; SEC31B; HSPA1B |
| Antigen processing and presentation | 3.85 × 10−4 | 1.45 × 10−2 | PDIA3; HSPA5; RFX5; HLA-C; CALR; CTSS; HSPA1B; CTSB; TAPBP |
| Central carbon metabolism in cancer | 5.68 × 10−4 | 1.66 × 10−2 | LDHA; PKM; PGAM1; IDH1; SLC2A1; SLC16A3; HK2; PFKP |
| Proteoglycans in cancer | 6.05 × 10−3 | 1.23 × 10−1 | TGFB1; ITGB5; FZD7; STAT3; FN1; WNT7A; ITPR2; CDC42; WNT11; SDC1; ITGA5; EZR; PPP1R12B |
| ECM-receptor interaction | 9.79 × 10−3 | 1.84 × 10−1 | ITGB5; FN1; SDC1; COL9A3; ITGB6; ITGA5; THBS3 |
| Human papillomavirus infection | 1.68 × 10−2 | 2.45 × 10−1 | ITGB5; FZD7; FN1; HLA-C; WNT7A; THBS3; CDC42; SLC9A3R1; PKM; WNT11; HEY1; ATP6V0A4; COL9A3; MAML3; ITGB6; ITGA5; TLR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lospinoso, K.; Dozmorov, M.; El Fawal, N.; Raghu, R.; Chae, W.-J.; Lee, E.D. Overexpression of ERAP2N in Human Trophoblast Cells Promotes Cell Death. Int. J. Mol. Sci. 2021, 22, 8585. https://doi.org/10.3390/ijms22168585
Lospinoso K, Dozmorov M, El Fawal N, Raghu R, Chae W-J, Lee ED. Overexpression of ERAP2N in Human Trophoblast Cells Promotes Cell Death. International Journal of Molecular Sciences. 2021; 22(16):8585. https://doi.org/10.3390/ijms22168585
Chicago/Turabian StyleLospinoso, Kristen, Mikhail Dozmorov, Nadine El Fawal, Rhea Raghu, Wook-Jin Chae, and Eun D. Lee. 2021. "Overexpression of ERAP2N in Human Trophoblast Cells Promotes Cell Death" International Journal of Molecular Sciences 22, no. 16: 8585. https://doi.org/10.3390/ijms22168585
APA StyleLospinoso, K., Dozmorov, M., El Fawal, N., Raghu, R., Chae, W.-J., & Lee, E. D. (2021). Overexpression of ERAP2N in Human Trophoblast Cells Promotes Cell Death. International Journal of Molecular Sciences, 22(16), 8585. https://doi.org/10.3390/ijms22168585