
Action of Multiple Rice β-Glucosidases on Abscisic Acid Glucose Ester

 and
and 
Abstract
1. Introduction
2. Results
2.1. Hydrolysis of ABA-GE by Rice Enzymes Belonging to GH1 Family
2.2. Kinetic Analysis of Os4BGlu12 and Os4BGlu13 Hydrolysis of ABA-GE
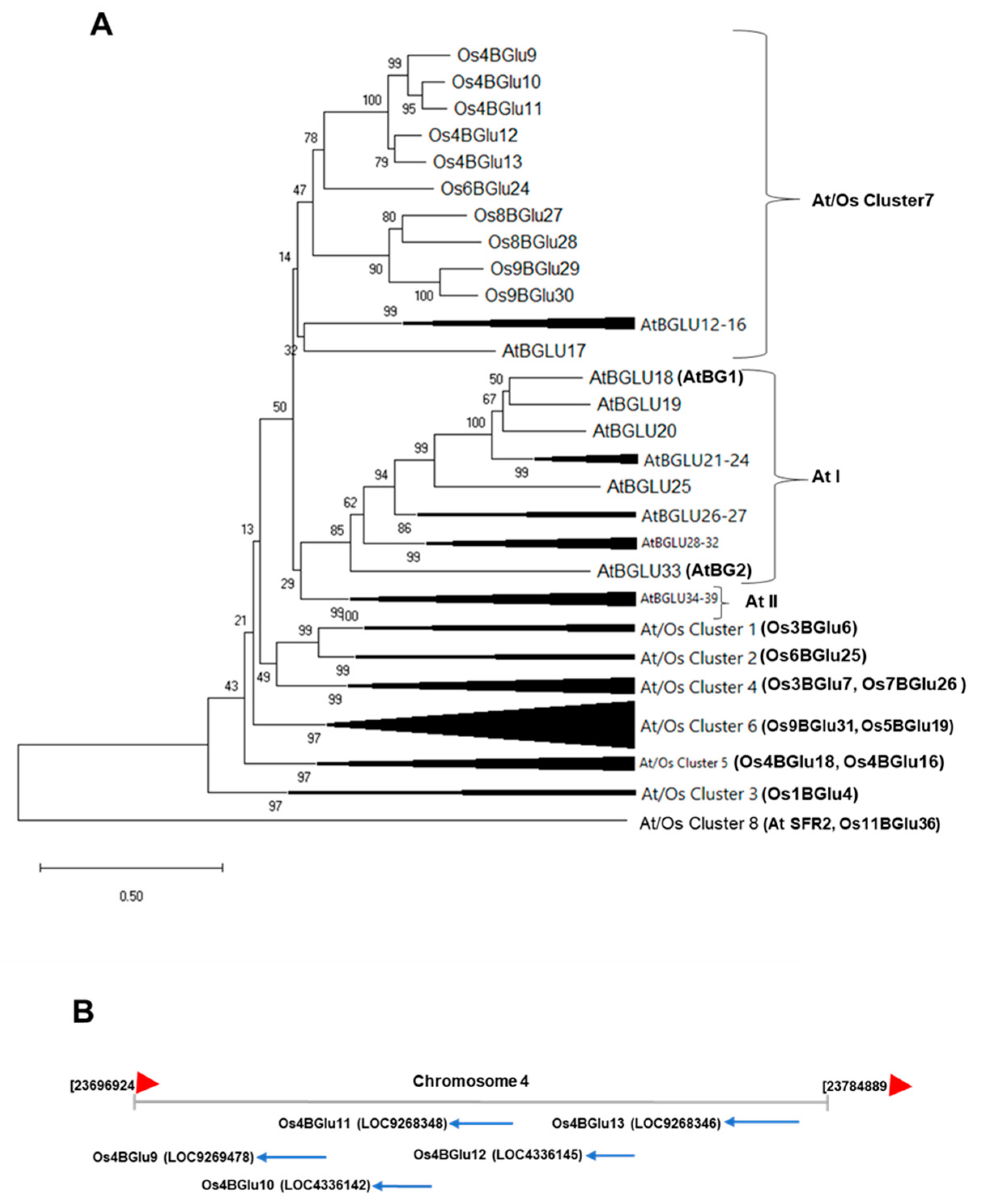
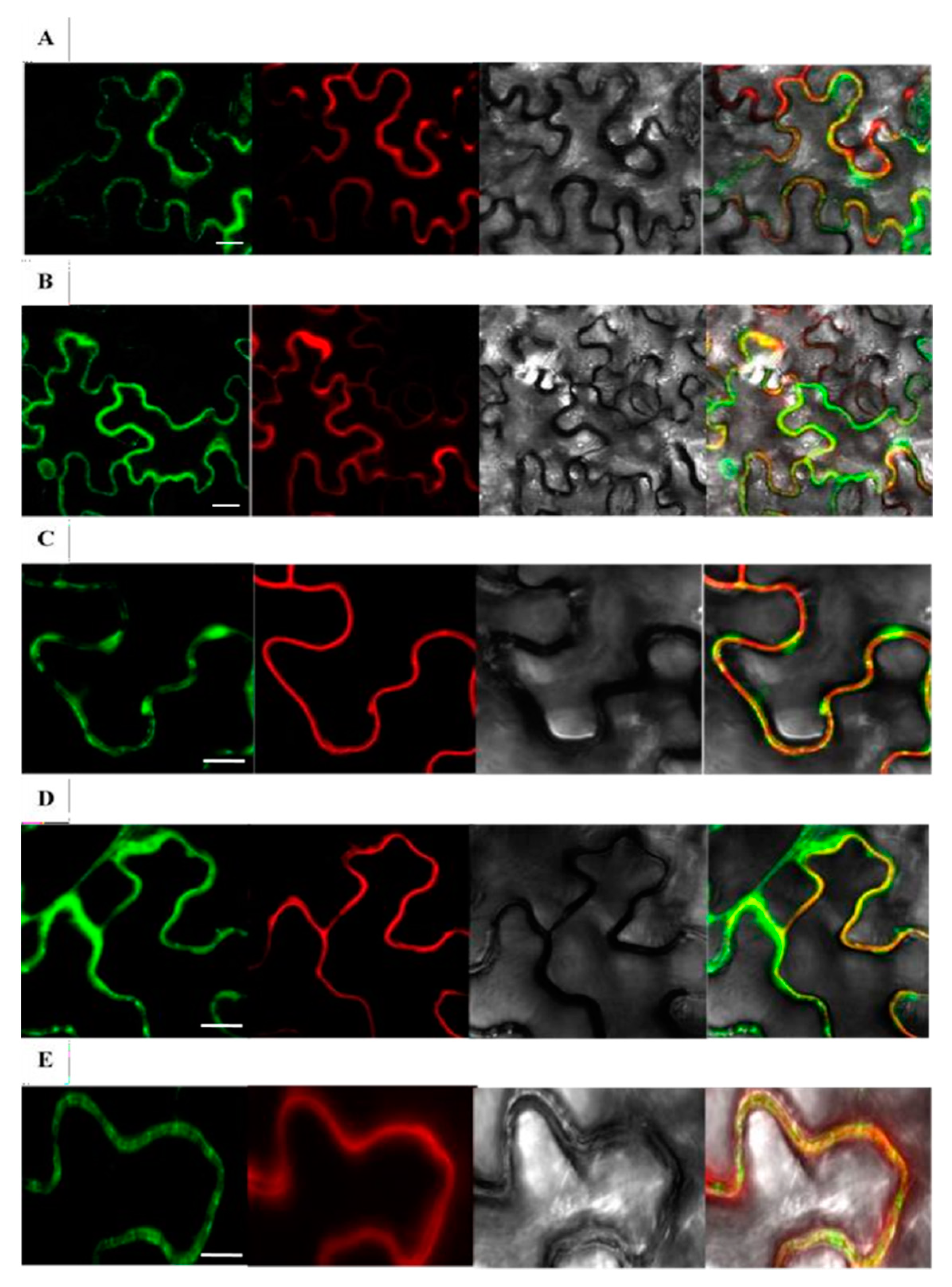
2.3. Subcellular Localization Examination of Rice β-Glucosidases Os4BGlu9, Os4BGlu10, Os4BGlu11, Os4BGlu12 and Os4BGlu13
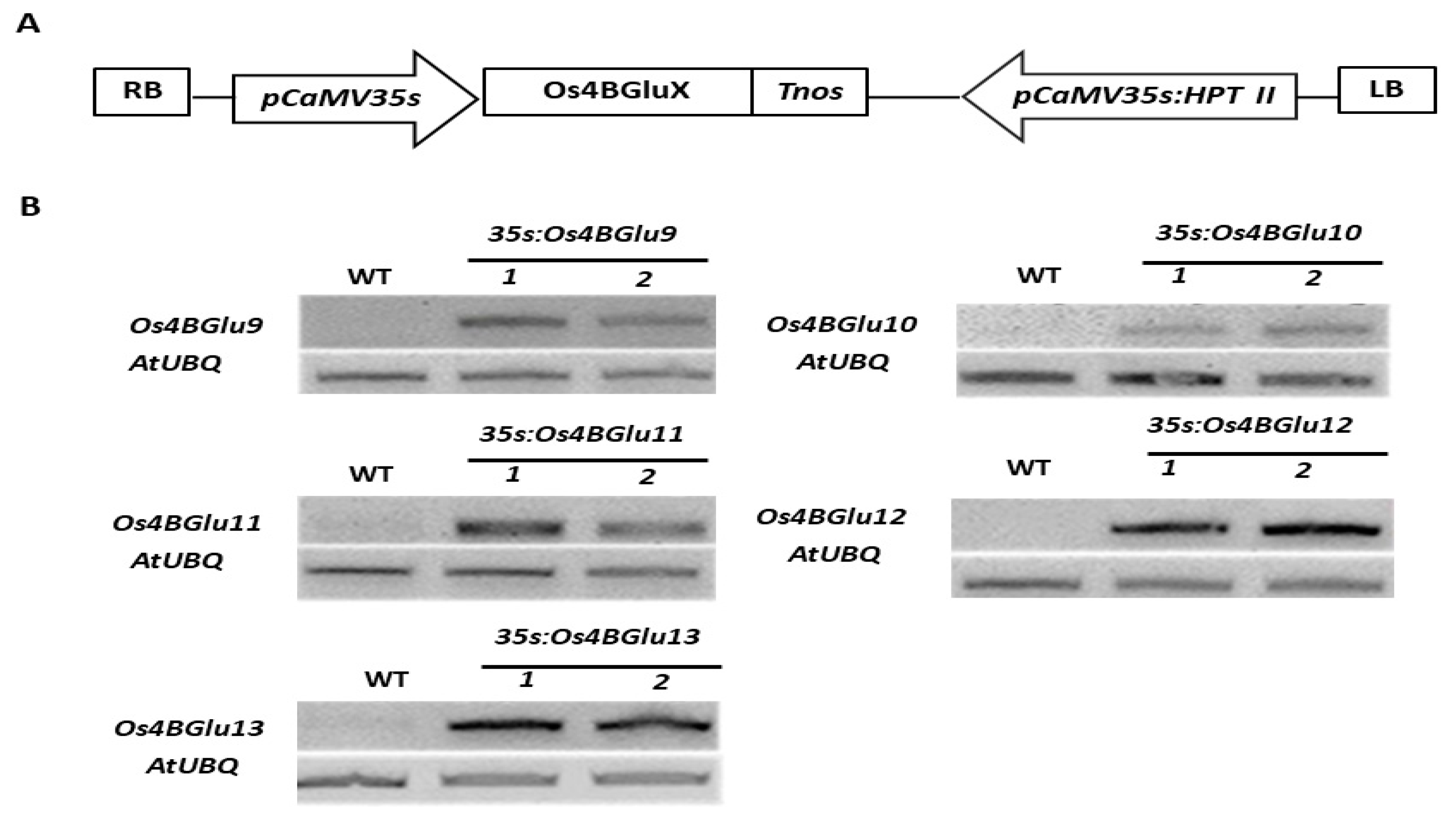
2.4. Generation of Transgenic Arabidopsis Plants Expressing Rice β-Glucosidases
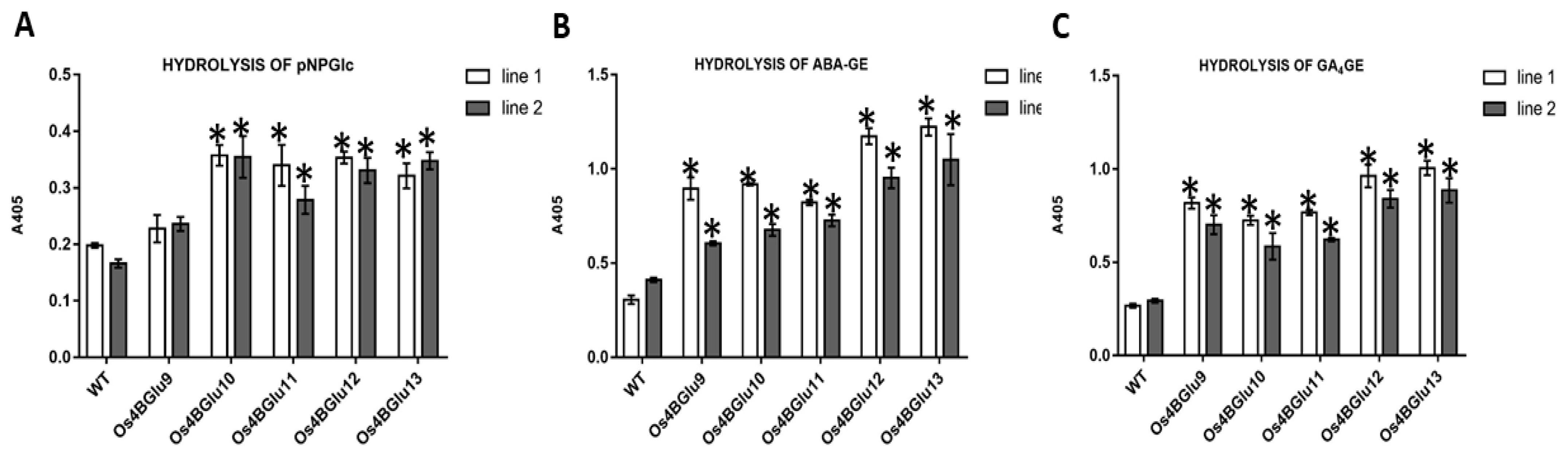
2.5. In Vitro Hydrolysis of pNPGlc, ABA-GE and GA4-GE by Extracts of Transgenic Arabidopsis Expressing Rice β-Glucosidases
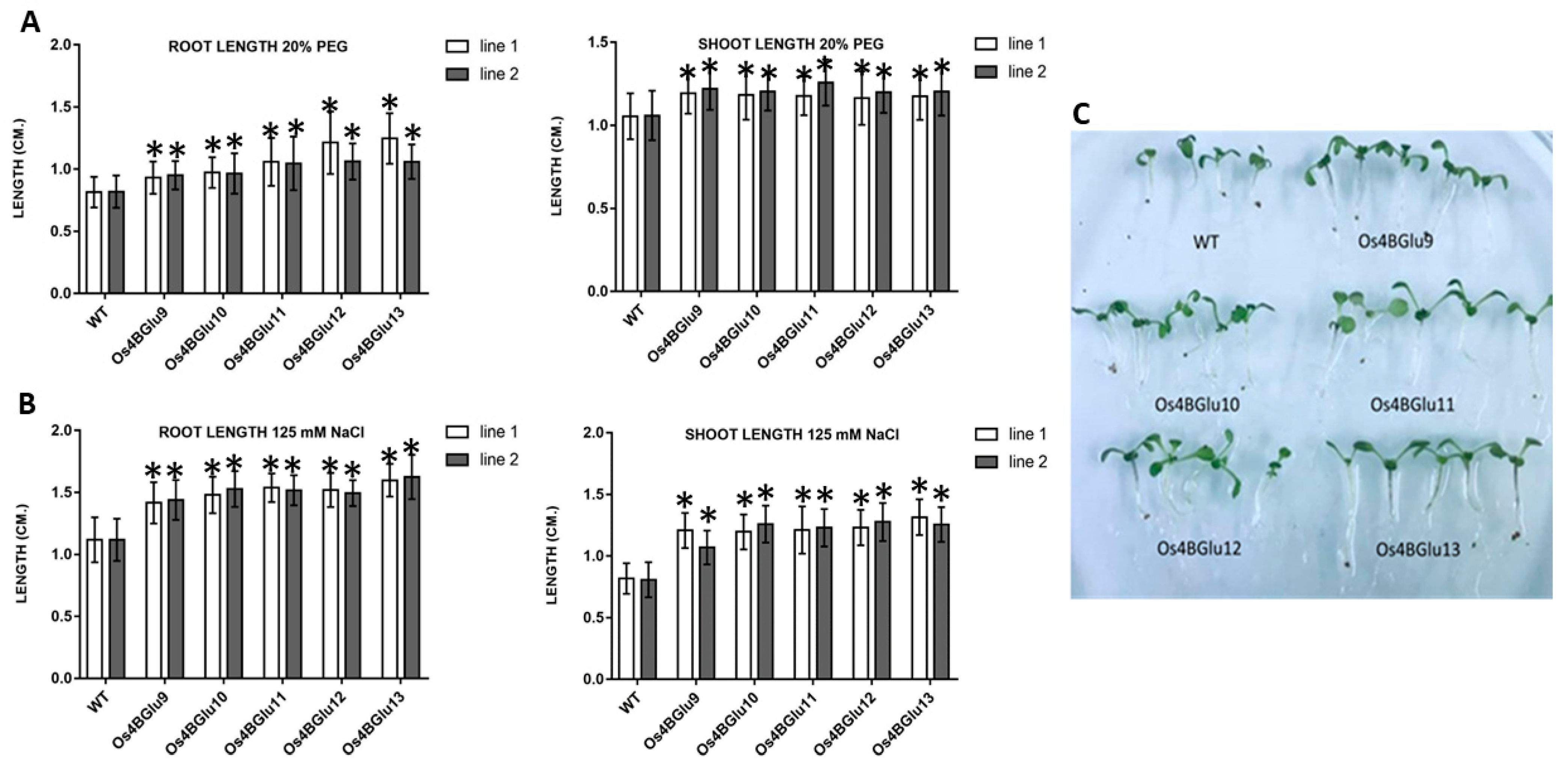
2.6. Growth of Arabidopsis Seedlings Expressing Rice β-Glucosidases upon Stress Treatments
3. Discussion
4. Materials and Methods
4.1. Production of ABA-GE by Os9BGlu31 Transglucosidase
4.2. The Hydrolysis Activity of Rice GH 1 Enzymes toward ABA-GE
4.3. Kinetic Study of Os4BGlu12 and Os4BGlu13 with ABA-GE
4.4. Subcellular Localization of Os4BGlu-GFP Fusion Proteins in Plant Cells
4.5. Construction of Plant Expression Vectors and Arabidopsis Transformation
4.6. RNA Isolation and RT-PCR Analysis
4.7. Growth and Treatments of Transgenic Arabidopsis
4.8. Extraction of Total Protein from Transgenic Arabidopsis and Determination of β-Glucosidase Activity
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Ketudat Cairns, J.R.; Esen, A. β-Glucosidases. Cell. Mol. Life Sci. 2010, 67, 3389–3405. [Google Scholar] [CrossRef]
- Ketudat Cairns, J.R.; Mahong, B.; Baiya, S.; Jeon, J.S. β-Glucosidases: Multitasking, moonlighting or simply misunderstood? Plant Sci. 2015, 241, 246–259. [Google Scholar] [CrossRef]
- Vishal, B.; Kumar, P.P. Regulation of seed germination and abiotic stresses by gibberellins and abscisic acid. Front. Plant Sci. 2018, 9, 838. [Google Scholar] [CrossRef]
- Ma, Y.; Cao, J.; He, J.; Chen, Q.; Li, X.; Yang, Y. Molecular mechanism for the regulation of ABA Homeostasis During plant development and stress responses. Int. J. Mol. Sci. 2018, 19, 3643. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. Drought, ozone, ABA and ethylene: New insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef]
- Piotrowska, A.; Bajguz, A. Conjugates of abscisic acid, brassinosteroids, ethylene, gibberellins, and jasmonates. Phytochemistry 2011, 72, 2097–2112. [Google Scholar] [CrossRef]
- Bray, E.A.; Zeevaart, J.A. The compartmentation of abscisic acid and beta-d-glucopyranosyl abscisate in mesophyll cells. Plant Physiol. 1985, 79, 719–722. [Google Scholar] [CrossRef]
- Lehmann, H.; Glund, K. Abscisic acid metabolism -vacuolar/extravacuolar distribution of metabolites. Planta 1986, 168, 559–562. [Google Scholar] [CrossRef]
- Liu, Z.; Yan, J.P.; Li, D.K.; Luo, Q.; Yan, Q.; Liu, Z.B.; Ye, L.M.; Wang, J.M.; Li, X.F.; Yang, Y. UDP-glucosyltransferase71c5, a major glucosyltransferase, mediates abscisic acid homeostasis in Arabidopsis. Plant Physiol. 2015, 167, 1659–1670. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Lee, K.H.; Dong, T.; Jeong, J.C.; Jin, J.B.; Kanno, Y.; Kim, D.H.; Kim, S.Y.; Seo, M.; Bressan, R.A.; et al. A vacuolar β-glucosidase homolog that possesses glucose-conjugated abscisic acid hydrolyzing activity plays an important role in osmotic stress responses in Arabidopsis. Plant Cell 2012, 24, 2184–2199. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J.; Sauter, A.; Wichert, K.; Messdaghi, D.; Hartung, W. Extracellular beta-glucosidase activity in barley involved in the hydrolysis of ABA glucose conjugate in leaves. J. Exp. Bot. 2000, 51, 937–944. [Google Scholar] [CrossRef]
- Lee, K.H.; Piao, H.L.; Kim, H.Y.; Choi, S.M.; Jiang, F.; Hartung, W.; Hwang, I.; Kwak, J.M.; Lee, I.J.; Hwang, I. Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid. Cell 2006, 126, 1109–1120. [Google Scholar] [CrossRef]
- Ondzighi-Assoume, C.A.; Chakraborty, S.; Harris, J.M. Environmental nitrate stimulates abscisic acid accumulation in Arabidopsis root tips by releasing it from inactive stores. Plant Cell 2016, 28, 729–745. [Google Scholar] [CrossRef]
- Han, Y.J.; Cho, K.C.; Hwang, O.J.; Choi, Y.S.; Shin, A.Y.; Hwang, I.; Kim, J.I. Overexpression of an Arabidopsis β-glucosidase gene enhances drought resistance with dwarf phenotype in creeping bentgrass. Plant Cell Rep. 2012, 31, 1677–1686. [Google Scholar] [CrossRef]
- Opassiri, R.; Pomthong, B.; Onkoksoong, T.; Akiyama, T.; Esen, A.; Ketudat Cairns, J.R. Analysis of rice glycosyl hydrolase family 1 and expression of Os4bglu12 beta-glucosidase. BMC Plant Biol. 2006, 6, 33. [Google Scholar] [CrossRef]
- Opassiri, R.; Ketudat Cairns, J.R.; Akiyama, T.; Wara-Aswapati, O.; Svasti, J.; Esen, A. Characterization of a rice β-glucosidase highly expressed in flower and germinating shoot. Plant Sci. 2003, 165, 627–638. [Google Scholar] [CrossRef]
- Opassiri, R.; Maneesan, J.; Akiyama, T.; Pomthong, B.; Jin, S.; Kimura, A.; Cairns, J. Rice Os4BGlu12 is a wound-induced β-glucosidase that hydrolyzes cell wall-β-glucan-derived oligosaccharides and glycosides. Plant Sci. 2010, 179, 273–280. [Google Scholar] [CrossRef]
- Luang, S.; Cho, J.I.; Mahong, B.; Opassiri, R.; Akiyama, T.; Phasai, K.; Komvongsa, J.; Sasaki, N.; Hua, Y.L.; Matsuba, Y.; et al. Rice Os9BGlu31 is a transglucosidase with the capacity to equilibrate phenylpropanoid, flavonoid, and phytohormone glycoconjugates. J. Biol. Chem. 2013, 288, 10111–10123. [Google Scholar] [CrossRef]
- Wakuta, S.; Hamada, S.; Ito, H.; Matsuura, H.; Nabeta, K.; Matsui, H. Identification of a beta-glucosidase hydrolyzing tuberonic acid glucoside in rice (Oryza sativa L.). Phytochemistry 2010, 71, 1280–1288. [Google Scholar] [CrossRef]
- Himeno, N.; Saburi, W.; Wakuta, S.; Takeda, R.; Matsuura, H.; Nabeta, K.; Sansenya, S.; Ketudat Cairns, J.R.; Mori, H.; Imai, R.; et al. Identification of rice β-glucosidase with high hydrolytic activity towards salicylic acid β-D-glucoside. Biosci. Biotechnol. Biochem. 2013, 77, 934–939. [Google Scholar] [CrossRef]
- Wakuta, S.; Hamada, S.; Ito, H.; Imai, R.; Mori, H.; Matsuura, H.; Nabeta, K.; Matsui, H. Comparison of Enzymatic Properties and Gene Expression Profiles of Two Tuberonic Acid Glucoside β-Glucosidases from Oryza sativa L. J. Appl. Glycosci. 2011, 58, 67–70. [Google Scholar] [CrossRef]
- Hua, Y.; Ekkhara, W.; Sansenya, S.; Srisomsap, C.; Roytrakul, S.; Saburi, W.; Takeda, R.; Matsuura, H.; Mori, H.; Ketudat Cairns, J.R. Identification of rice Os4BGlu13 as a β-glucosidase which hydrolyzes gibberellin A4 1-O-β-d-glucosyl ester, in addition to tuberonic acid glucoside and salicylic acid derivative glucosides. Arch. Biochem. Biophys. 2015, 583, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, S.; Dong, Y.; Ren, R.; Chen, D.; Chen, X. Chloroplastic Os3BGlu6 contributes significantly to cellular ABA pools and impacts drought tolerance and photosynthesis in rice. New Phytol. 2020, 226, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Sansenya, S.; Saetang, C.; Wakuta, S.; Ketudat Cairns, J.R. Enzymatic and structural characterization of hydrolysis of gibberellin A4 glucosyl ester by a rice β-D-glucosidase. Arch. Biochem. Biophys. 2013, 537, 39–48. [Google Scholar] [CrossRef]
- Akiyama, T.; Kaku, H.; Shibuya, N. A cell wall-bound beta-glucosidase from germinated rice: Purification and properties. Phytochemistry 1998, 48, 49–54. [Google Scholar] [CrossRef]
- Chen, T.T.; Liu, F.F.; Xiao, D.W.; Jiang, X.Y.; Li, P.; Zhao, S.M.; Hou, B.K.; Li, Y.J. The Arabidopsis UDP-glycosyltransferase75B1, conjugates abscisic acid and affects plant response to abiotic stresses. Plant Mol. Biol. 2020, 102, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, L.; Forde, B.G.; Davies, W.J. The biphasic root growth response to abscisic acid in Arabidopsis involves interaction with ethylene and auxin signalling pathways. Front. Plant Sci. 2017, 8, 1493. [Google Scholar] [CrossRef]
- Tardieu, F.; Parent, B.; Simonneau, T. Control of leaf growth by abscisic acid: Hydraulic or non-hydraulic processes? Plant Cell Environ. 2010, 33, 636–647. [Google Scholar] [CrossRef]
- Sharp, R.E.; LeNoble, M.E. ABA, ethylene and the control of shoot and root growth under water stress. J. Exp. Bot. 2002, 53, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Li, D.; Zhen, C.; Chen, D.; Chen, X. Specific roles of Os4BGlu10, Os6BGlu24, and Os9BGlu33 in seed germination, root elongation, and drought tolerance in rice. Planta 2019, 249, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic acid and gibberellins antagonistically mediate plant development and abiotic stress responses. Front. Plant Sci. 2018, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, L.; Cheng, C.; Ren, Z.; Xu, S.; Li, X. GAI Functions in the plant response to dehydration stress in Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 819. [Google Scholar] [CrossRef]
- Rouyi, C.; Baiya, S.; Lee, S.K.; Mahong, B.; Jeon, J.S.; Ketudat-Cairns, J.R.; Ketudat-Cairns, M. Recombinant expression and characterization of the cytoplasmic rice β-glucosidase Os1BGlu4. PLoS ONE 2014, 9, e96712. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Akiyama, T.; Opassiri, R.; Kuaprasert, B.; Cairns, J.K. Structural and enzymatic characterization of Os3BGlu6, a rice beta-glucosidase hydrolyzing hydrophobic glycosides and (1→3)- and (1→2)-linked disaccharides. Plant Physiol. 2009, 151, 47–58. [Google Scholar] [CrossRef]
- Baiya, S.; Hua, Y.; Ekkhara, W.; Ketudat Cairns, J.R. Expression and enzymatic properties of rice (Oryza sativa L.) monolignol β-glucosidases. Plant Sci. 2014, 227, 101–109. [Google Scholar] [CrossRef]
- Kuntothom, T.; Luang, S.; Harvey, A.J.; Fincher, G.B.; Opassiri, R.; Hrmova, M.; Ketudat Cairns, J.R. Rice family GH1 glycoside hydrolases with beta-D-glucosidase and beta-D-mannosidase activities. Arch. Biochem. Biophys. 2009, 491, 85–95. [Google Scholar] [CrossRef]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Von Schaewen, A.; Rips, S.; Jeong, I.S.; Koiwa, H. Arabidopsis thaliana KORRIGAN1 protein: N-glycan modification, localization, and function in cellulose biosynthesis and osmotic stress responses. Plant Signal. Behav. 2015, 10, e1024397. [Google Scholar] [CrossRef][Green Version]
- Baiya, S.; Mahong, B.; Lee, S.K.; Jeon, J.S.; Ketudat Cairns, J.R. Demonstration of monolignol β-glucosidase activity of rice Os4BGlu14, Os4BGlu16 and Os4BGlu18 in Arabidopsis thaliana bglu45 mutant. Plant Physiol. Biochem. 2018, 127, 223–230. [Google Scholar] [CrossRef]
- Himmelbach, A.; Zierold, U.; Hensel, G.T.; Riechen, J.; Douchkov, D.; Schweizer, P.; Kumlehn, J. A Set of modular binary vectors for transformation of cereals. Plant Physiol. 2007, 145, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.S.; Lee, S.; Jung, K.H.; Jun, S.H.; Jeong, D.H.; Lee, J.; Kim, C.; Jang, S.; Yang, K.; Nam, J.; et al. T-DNA insertional mutagenesis for functional genomics in rice. Plant J. 2000, 22, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, S.; Satoh, K.; Nagata, T.; Kawagashira, N.; Doi, K.; Kishimoto, N.; Yazaki, J.; Ishikawa, M.; Yamada, H.; Ooka, H.; et al. Collection, mapping, and annotation of over 28,000 cDNA clones from japonica rice. Science 2003, 301, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Oppenheimer, D.G.; Haas, N.; Silflow, C.D.; Snustad, D.P. The beta-tubulin gene family of Arabidopsis thaliana: Preferential accumulation of the beta 1 transcript in roots. Gene 1988, 63, 87–102. [Google Scholar] [CrossRef]
- Van der Weele, C.M.; Spollen, W.G.; Sharp, R.E.; Baskin, T.I. Growth of Arabidopsis thaliana seedlings under water deficit studied by control of water potential in nutrient-agar media. J. Exp. Bot. 2000, 51, 1555–1562. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| β-Glucosidase | Specific Activity (µM·mg−1·min−1) |
|---|---|
| Os4BGlu13 | 1.63 × 10−3 ± 0.005 |
| Os4BGlu12 | 1.54 × 10−3 ± 0.027 |
| Os1BGlu4 | 1.31 × 10−3 ± 0.019 |
| Os3BGlu7 | 1.18 × 10−3 ± 0.040 |
| Os4BGlu18 | 1.16 × 10−3 ± 0.026 |
| Barley βII | 0.57 × 10−3 ± 0.008 |
| Os3BGlu6 | 0.39 × 10−3 ± 0.004 |
| Os7BGlu26 | 0.11 × 10−3 ± 0.009 |
| Os9BGlu31 | 0.06 × 10−3 ± 0.004 |
| β-Glucosidase | KM (mM) | kcat (s−1) | kcat/KM (mM−1s−1) |
|---|---|---|---|
| Os4BGlu12 | 10.9 ± 0.9 | 7.50 ± 0.0002 | 0.689 |
| Os4BGlu13 | 1.66 ± 0.09 | 20.6 ± 0.5 | 12.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kongdin, M.; Mahong, B.; Lee, S.-K.; Shim, S.-H.; Jeon, J.-S.; Ketudat Cairns, J.R. Action of Multiple Rice β-Glucosidases on Abscisic Acid Glucose Ester. Int. J. Mol. Sci. 2021, 22, 7593. https://doi.org/10.3390/ijms22147593
Kongdin M, Mahong B, Lee S-K, Shim S-H, Jeon J-S, Ketudat Cairns JR. Action of Multiple Rice β-Glucosidases on Abscisic Acid Glucose Ester. International Journal of Molecular Sciences. 2021; 22(14):7593. https://doi.org/10.3390/ijms22147593
Chicago/Turabian StyleKongdin, Manatchanok, Bancha Mahong, Sang-Kyu Lee, Su-Hyeon Shim, Jong-Seong Jeon, and James R. Ketudat Cairns. 2021. "Action of Multiple Rice β-Glucosidases on Abscisic Acid Glucose Ester" International Journal of Molecular Sciences 22, no. 14: 7593. https://doi.org/10.3390/ijms22147593
APA StyleKongdin, M., Mahong, B., Lee, S.-K., Shim, S.-H., Jeon, J.-S., & Ketudat Cairns, J. R. (2021). Action of Multiple Rice β-Glucosidases on Abscisic Acid Glucose Ester. International Journal of Molecular Sciences, 22(14), 7593. https://doi.org/10.3390/ijms22147593