
Extreme Enlargement of the Inverted Repeat Region in the Plastid Genomes of Diatoms from the Genus Climaconeis

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Morphology
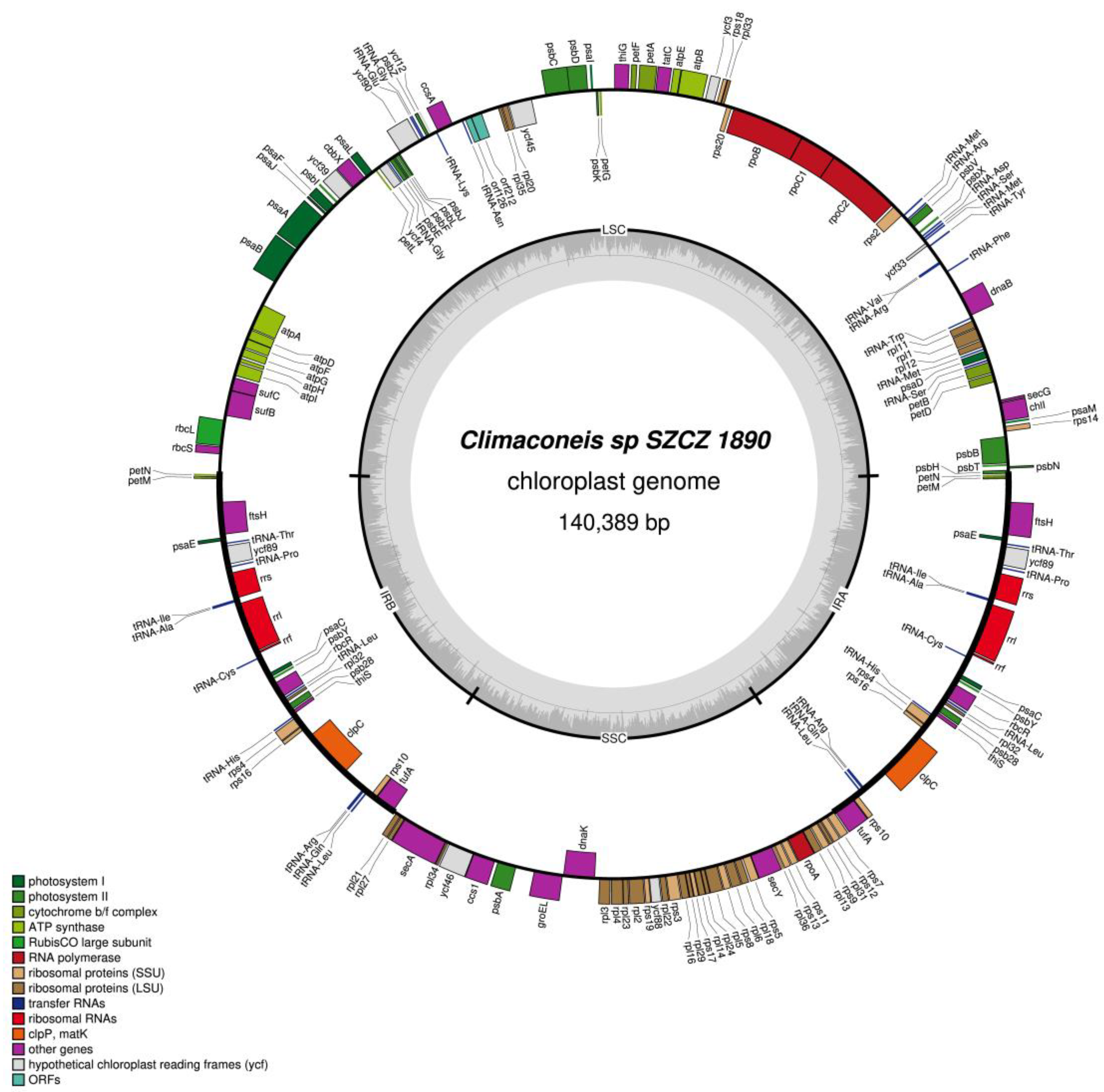
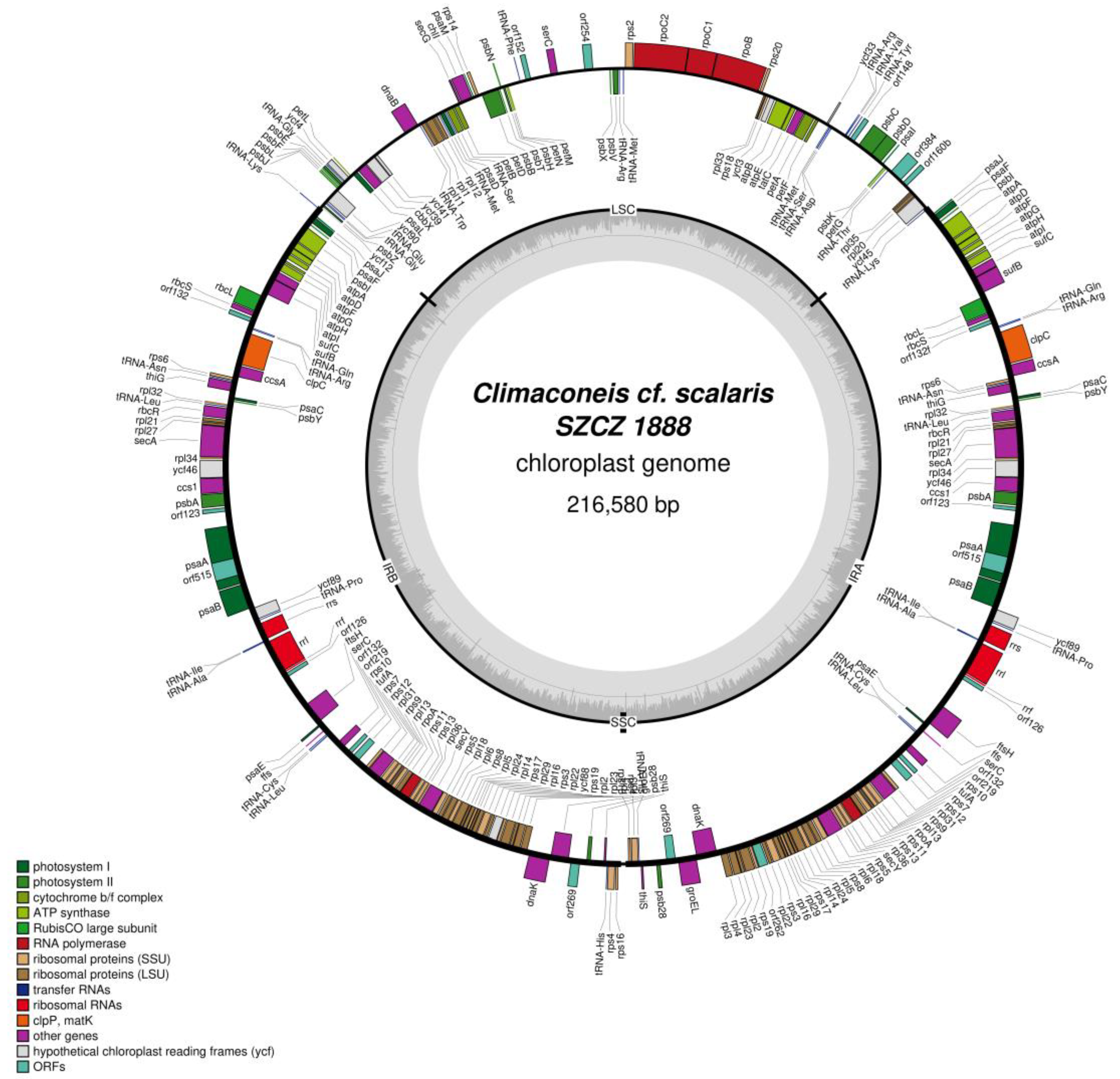
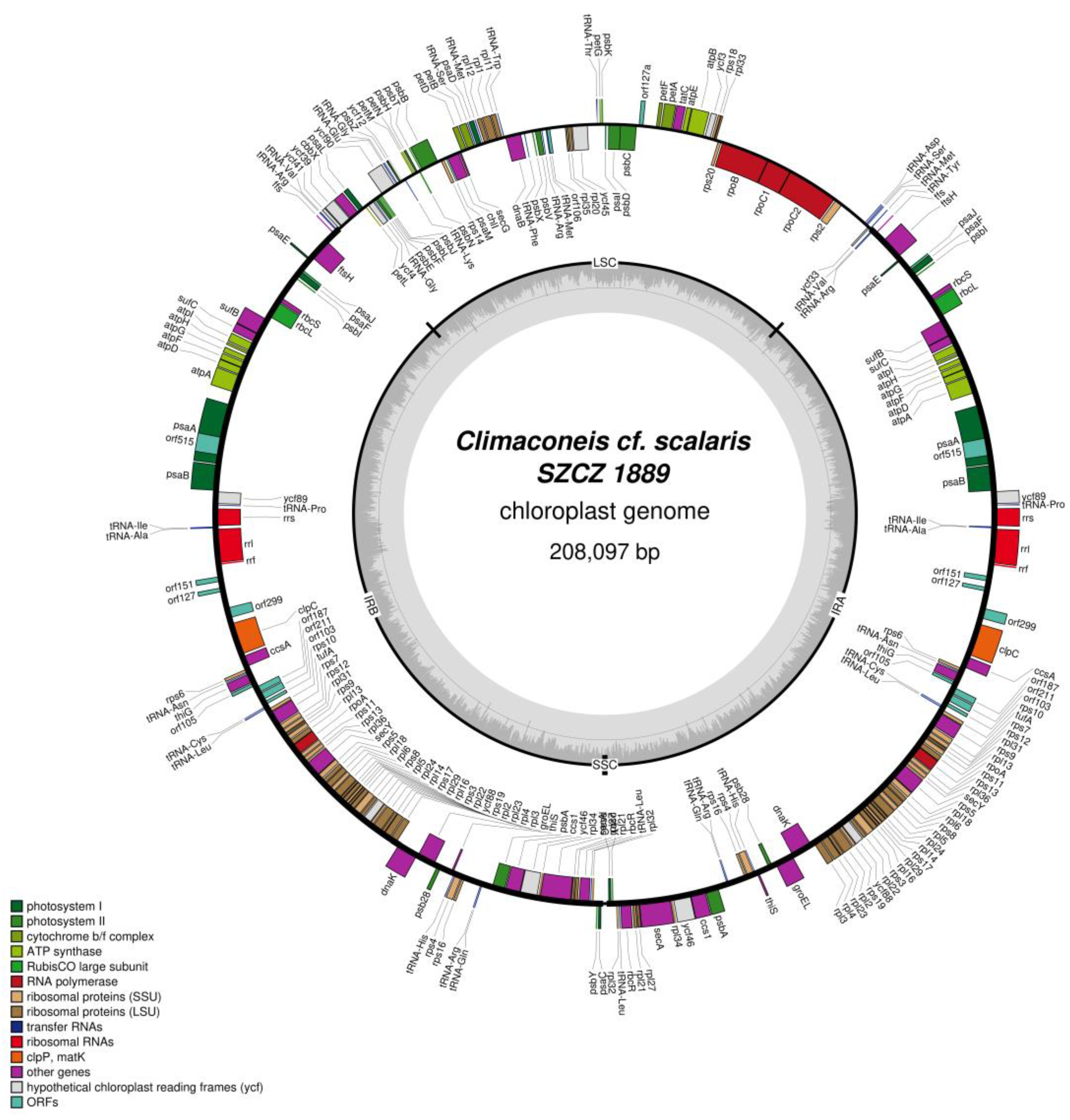
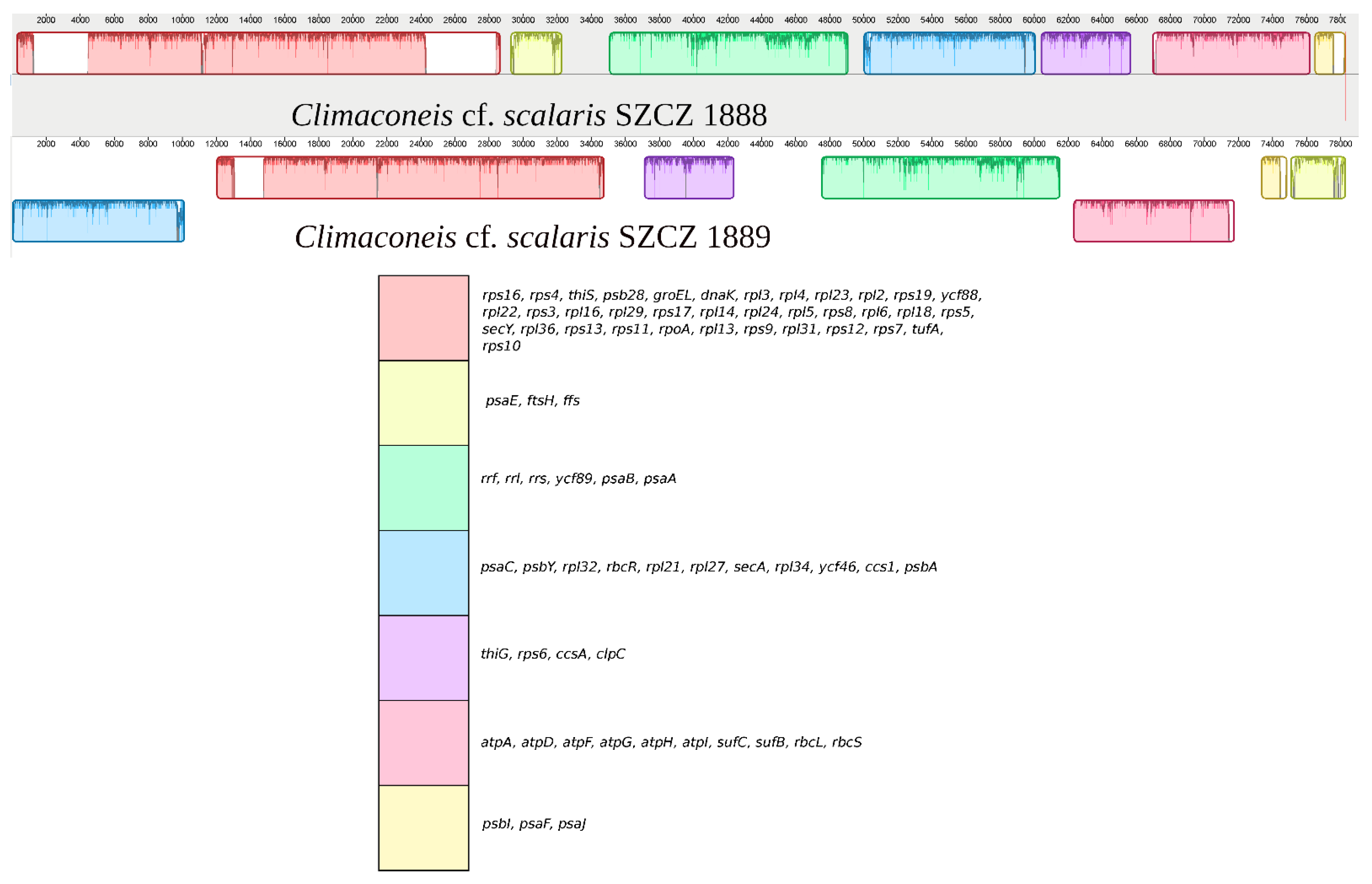
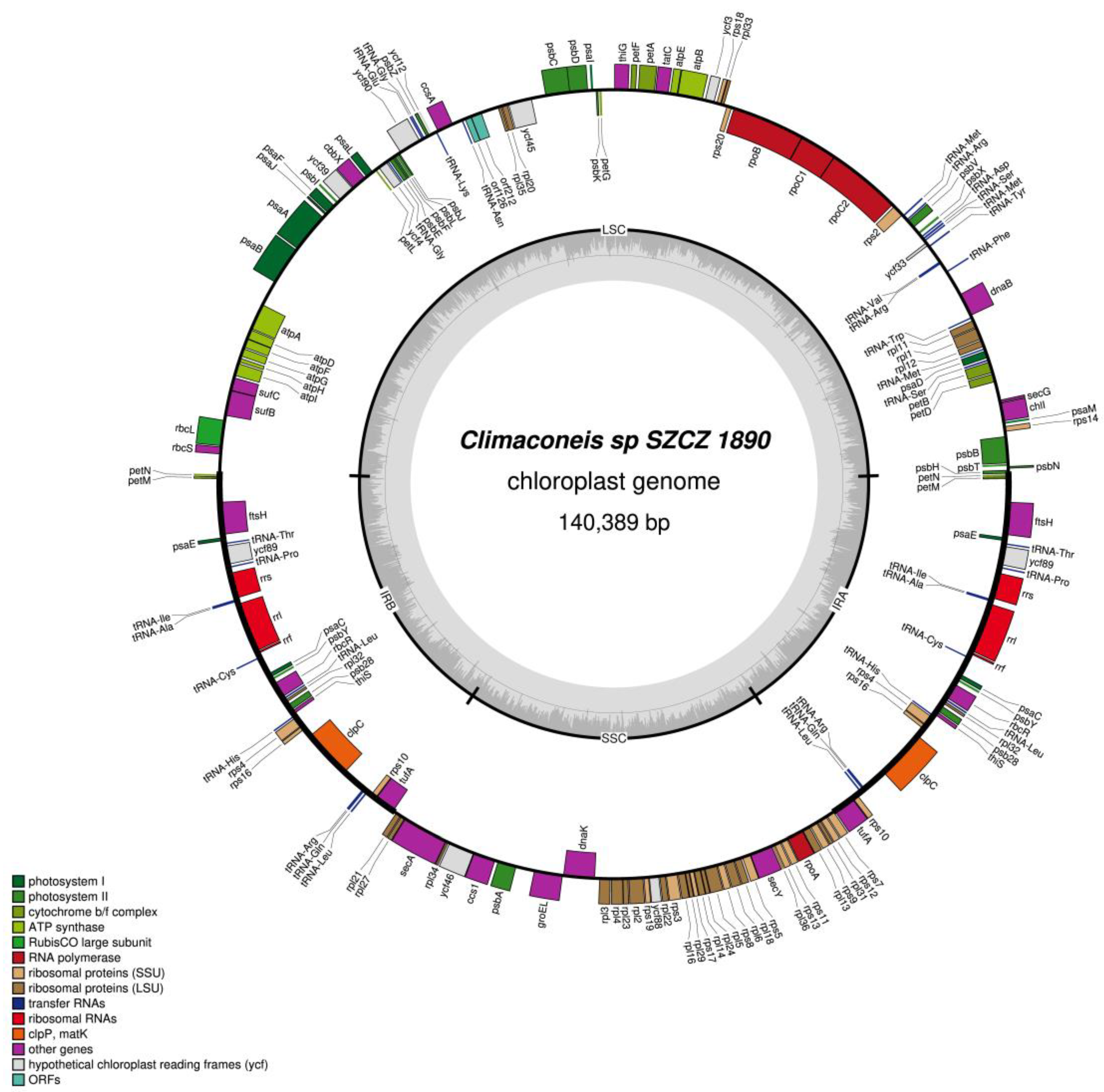
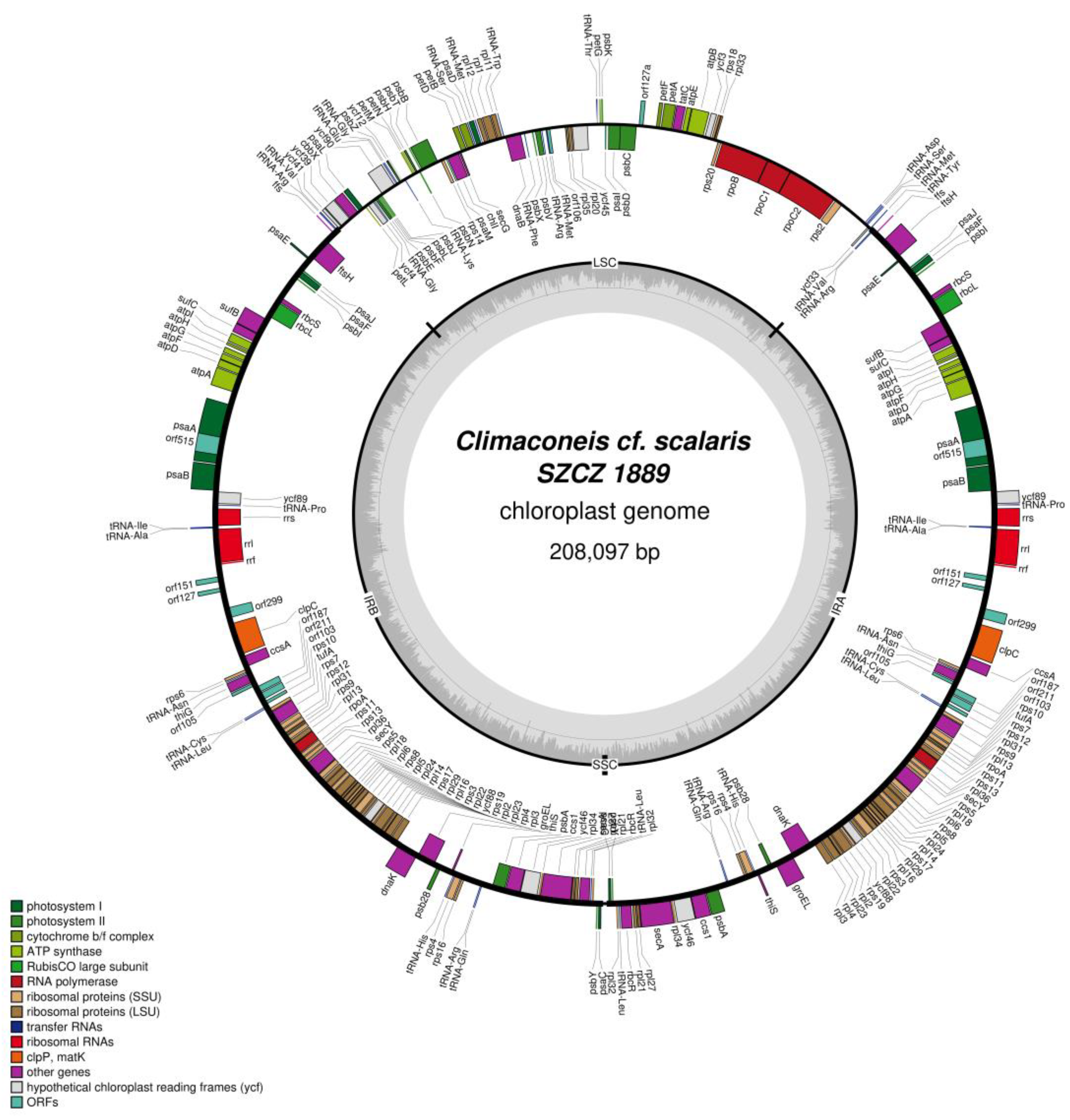
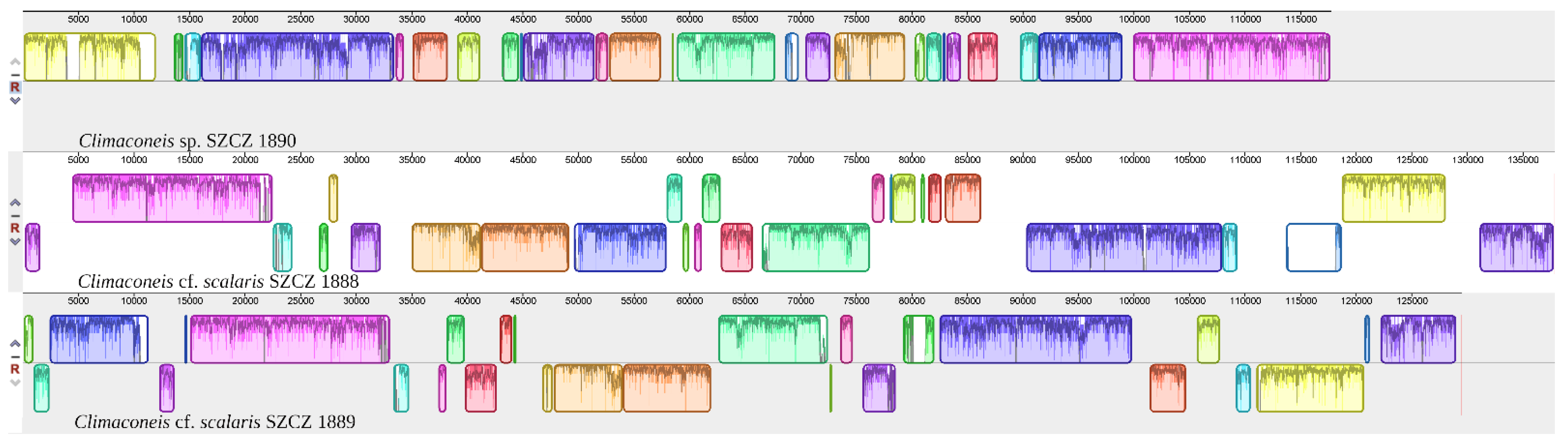
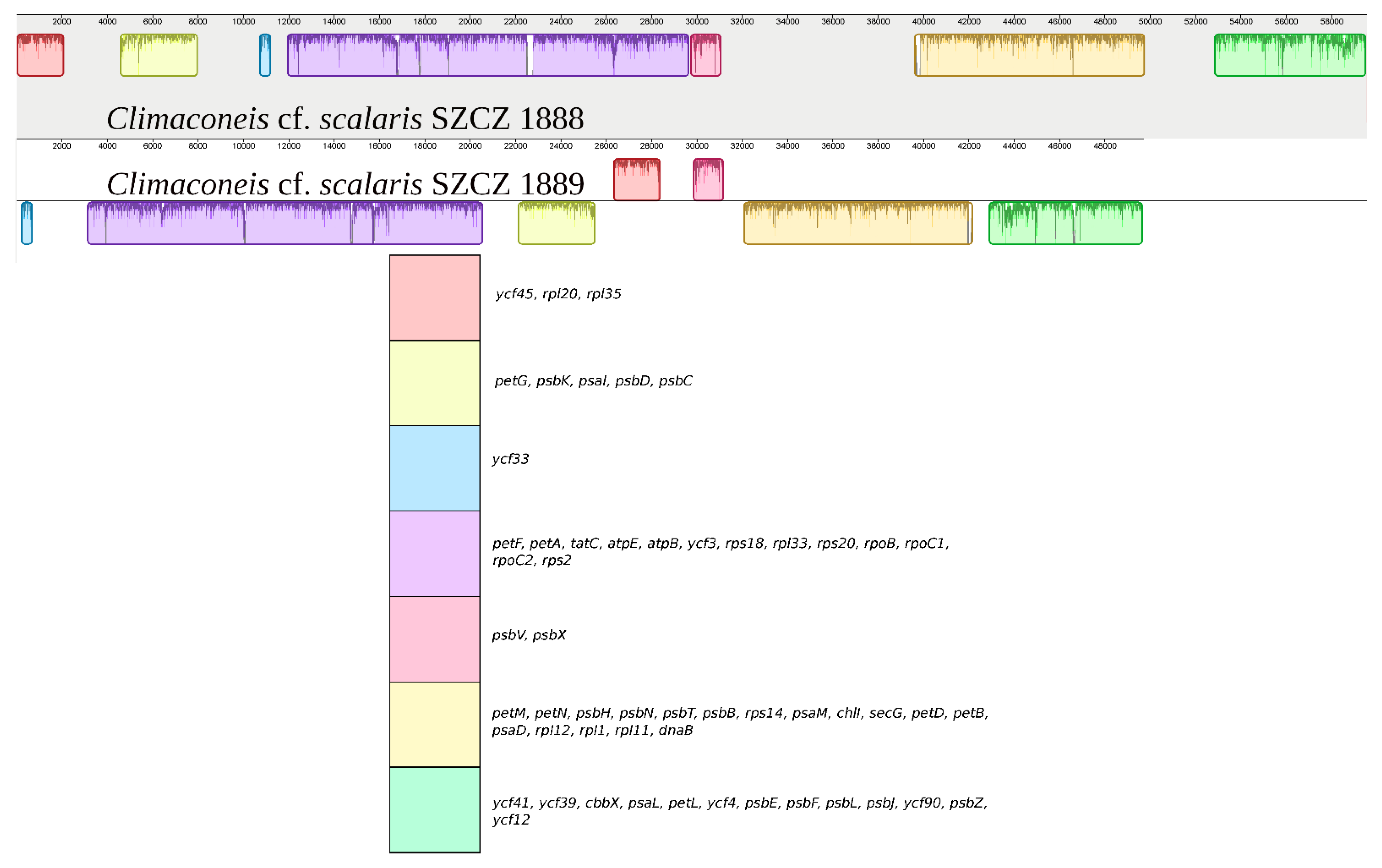
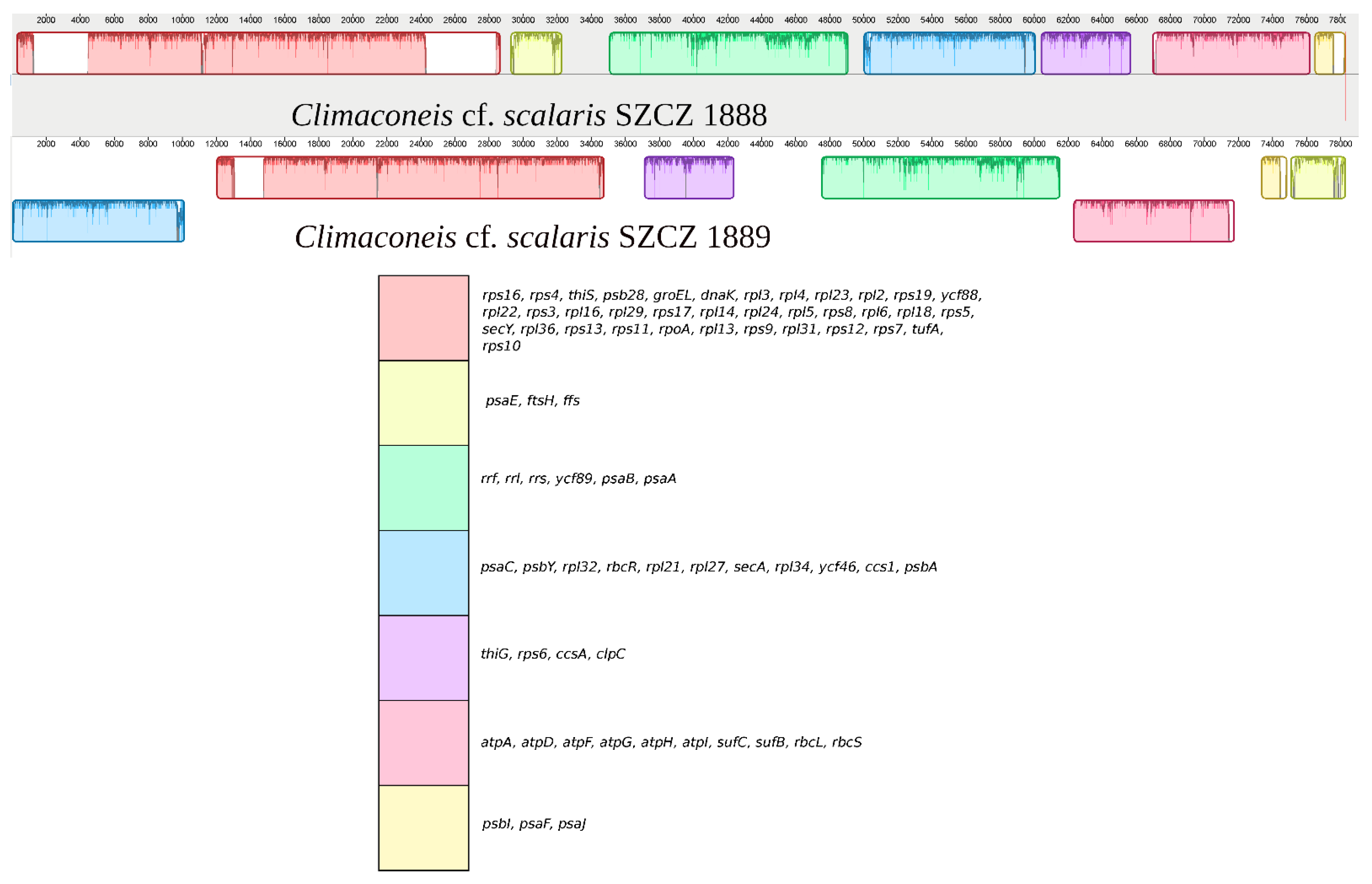
2.2. Structure of the Sequenced Plastid Genomes
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Scanning Electron Microscopy
4.3. DNA Extraction, Sequencing and Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Amato, A.; Kooistra, W.H.; Ghiron, J.H.; Mann, D.G.; Pröschold, T.; Montresor, M. Reproductive isolation among sympatric cryptic species in marine diatoms. Protist 2007, 158, 193–207. [Google Scholar] [CrossRef]
- Beszteri, B.; Acs, E.; Medlin, L.K. Ribosomal DNA sequence variation among sympatric strains of the Cyclotella meneghiniana complex (Bacillariophyceae) reveals cryptic diversity. Protist 2005, 156, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarska, I.; Mather, L.A.; Luddington, I.A.; Muise, F.; Ehrman, J.M. Cryptic diversity in a cosmopolitan diatom known as Asterionellopsis glacialis (Fragilariaceae): Implications for ecology, biogeography, and taxonomy. Am. J. Bot. 2014, 101, 267–286. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, N.; Bates, S.S.; Baugh, K.A.; Bill, B.D.; Connell, L.B.; Léger, C.; Trainer, V.L. Cryptic and pseudo-cryptic diversity in diatoms—With descriptions of Pseudo-nitzschia hasleana sp. nov. and P. fryxelliana sp. nov. J. Phycol. 2012, 48, 436–454. [Google Scholar] [CrossRef] [PubMed]
- Pinseel, E.; Kulichová, J.; Scharfen, V.; Urbánková, P.; Van de Vijver, B.; Vyverman, W. Extensive cryptic diversity in the terrestrial diatom Pinnularia borealis (Bacillariophyceae). Protist 2019, 170, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Poulícková, A.; Veselá, J.; Neustupa, J.; Skaloud, P. Pseudocryptic diversity versus cosmopolitanism in diatoms: A case study on Navicula cryptocephala Kütz. (Bacillariophyceae) and morphologically similar taxa. Protist 2010, 161, 353–369. [Google Scholar] [CrossRef]
- Souffreau, C.; Vanormelingen, P.; Van de Vijver, B.; Isheva, T.; Verleyen, E.; Sabbe, K.; Vyverman, W. Molecular evidence for distinct Antarctic lineages in the cosmopolitan terrestrial diatoms Pinnularia borealis and Hantzschia amphioxys. Protist 2013, 164, 101–115. [Google Scholar] [CrossRef]
- Grünow, A. Diatomeen auf sargassum von Honduras, gesammelt von Linding. Hedwigia 1867, 6, 17–32. [Google Scholar]
- Van Heurck, H. A Treatise on the Diatomaceae; Baxter, W.W.E., Translator; Wesley & Son: London, UK, 1896. [Google Scholar]
- Guiry, M.D.; Guiry, G.M.; AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 27 May 2021).
- Cox, E.J. Taxonomic studies on the diatom genus Navicula Bory. IV. Climaconeis grun., a genus including Okedenia inflexa (Bréb.) eulenst. ex de toniand members of Navicula sect. Johnsonieae sensu hustedt. Brit. Phycol. J. 1982, 17, 147–168. [Google Scholar] [CrossRef]
- De Brébisson, L.A. Considerations sur les Diatomées et Essai D’une Classification des Genres et des Espèces Appartenant à Cette Famille; Par A de Brébisson, Auteur de la Flore de Normandie, etc.; Brée l’Ainée Imprimeur-Libraire: Meilhac, France, 1838. [Google Scholar]
- Merschkowsky, C. On Okedenia Eul. Ann. Mag. Nat. Hist. 1901, 8, 415–423. [Google Scholar] [CrossRef]
- Davidovich, O.I.; Davidovich, N.A.; Gastineau, R.; Witkowski, A. Sexual reproduction of the black sea diatom Climaconeis scalaris (Brébisson) E.J. Cox. Mosc. Univ. Biol. Sci. Bull. 2020, 74, 200–206. [Google Scholar] [CrossRef]
- Chepurnov, V.A.; Mann, D.G.; Sabbe, K.; Vyverman, W. Experimental studies on sexual reproduction in diatoms. Int. Rev. Cytol. 2004, 237, 91–154. [Google Scholar] [PubMed]
- Gastineau, R.; Leignel, V.; Jacquette, B.; Hardivillier, Y.; Wulff, A.; Gaudin, P.; Bendahmane, D.; Davidovich, N.A.; Kaczmarska, I.; Mouget, J.L. Inheritance of mitochondrial DNA in the pennate diatom Haslea ostrearia (Naviculaceae) during auxosporulation suggests a uniparental transmission. Protist 2013, 164, 340–351. [Google Scholar] [CrossRef]
- Ruck, E.C.; Nakov, T.; Jansen, R.K.; Theriot, E.C.; Alverson, A.J. Serial gene losses and foreign DNA underlie size and sequence variation in the plastid genomes of diatoms. Genome Biol. Evol. 2014, 6, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastineau, R.; Hansen, G.; Poulin, M.; Lemieux, C.; Turmel, M.; Bardeau, J.-F.; Leignel, V.; Hardivillier, Y.; Morançais, M.; Fleurence, J.; et al. Haslea silbo, a novel cosmopolitan species of blue diatoms. Biology 2021, 10, 328. [Google Scholar] [CrossRef] [PubMed]
- Han, K.Y.; Maciszewski, K.; Graf, L.; Yang, J.H.; Andersen, R.A.; Karnkowska, A.; Yoon, H.S. Dictyochophyceae plastid genomes reveal unusual variability in their organization. J. Phycol. 2019, 55, 1166–1180. [Google Scholar] [CrossRef]
- Muñoz-Gómez, S.A.; Mejía-Franco, F.G.; Durnin, K.; Colp, M.; Grisdale, C.J.; Archibald, J.M.; Slamovits, C.H. The new red algal subphylum proteorhodophytina comprises the largest and most divergent plastid genomes known. Curr. Biol. 2017, 27, 1677–1684. [Google Scholar] [CrossRef] [Green Version]
- Lemieux, C.; Vincent, A.T.; Labarre, A.; Otis, C.; Turmel, M. Chloroplast phylogenomic analysis of chlorophyte green algae identifies a novel lineage sister to the Sphaeropleales (Chlorophyceae). BMC Evol. Biol. 2015, 15, 264. [Google Scholar] [CrossRef] [Green Version]
- Cox, E.J. Studies on the diatom genus Navicula Bory. Navicula scopulorum Bréb. and a further comment on the genus Berkeleya grev. Brit. Phycol. J. 1979, 14, 161–174. [Google Scholar] [CrossRef]
- Yu, M.; Ashworth, M.P.; Hajrah, N.H.; Khiyami, M.A.; Sabir, M.J.; Alhebshi, A.M.; Al-Malki, A.L.; Sabir, J.S.M.; Theriot, E.C.; Jansen, R.K. Evolution of the plastid genomes in diatoms. Adv. Bot. Res. 2018, 85, 129–155. [Google Scholar]
- Ruck, E.C.; Linard, S.R.; Nakov, T.; Theriot, E.C.; Alverson, A.J. Hoarding and horizontal transfer led to an expanded gene and intron repertoire in the plastid genome of the diatom, Toxarium undulatum (Bacillariophyta). Curr. Genet. 2017, 63, 499–507. [Google Scholar] [CrossRef]
- Turmel, M.; de Cambiaire, J.C.; Otis, C.; Lemieux, C. Distinctive architecture of the chloroplast genome in the chlorodendrophycean green algae Scherffelia dubia and Tetraselmis sp. CCMP 881. PLoS ONE 2016, 11, e0148934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Li, H.; Sahu, S.K.; Xu, Y.; Wang, H.; Liang, H.; Wang, S. The complete chloroplast genome of Tetraselmis desikacharyi (Chlorodendrophyceae) and phylogenetic analysis. Mitochondrial DNA Part B 2019, 4, 1692–1693. [Google Scholar] [CrossRef] [Green Version]
- Gastineau, R.; Yılmaz, E.; Solak, C.N.; Lemieux, C.; Turmel, M.; Witkowski, A. Complete chloroplast genome of the mixotrophic chrysophyte Poterioochromonas malhamensis (Ochromonadales, Synurophyceae) from Van Lake in Eastern Anatolia. Mitochondrial DNA Part B, In press.
- Kim, J.I.; Shin, H.; Škaloud, P.; Jung, J.; Yoon, H.S.; Archibald, J.M.; Shin, W. Comparative plastid genomics of Synurophyceae: Inverted repeat dynamics and gene content variation. BMC Evol. Biol. 2019, 19, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ševčíková, T.; Horák, A.; Klimeš, V.; Zbránková, V.; Demir-Hilton, E.; Sudek, S.; Jenkins, J.; Schmutz, J.; Přibyl, P.; Fousek, J.; et al. Updating algal evolutionary relationships through plastid genome sequencing: Did alveolate plastids emerge through endosymbiosis of an ochrophyte? Sci. Rep. 2015, 5, 10134. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.L.; Ruhlman, T.A.; Jansen, R.K. Expansion of inverted repeat does not decrease substitution rates in pelargonium plastid genomes. New Phytol. 2017, 214, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.; Green, P. Consed: A graphical editor for next-generation sequencing. Bioinformatics 2013, 29, 2936–2937. [Google Scholar] [CrossRef] [Green Version]
- Gastineau, R.; Hamedi, C.; Baba Hamed, M.B.; Abi-Ayad, S.-M.E.-A.; Bąk, M.; Lemieux, C.; Turmel, M.; Dobosz, S.; Wróbel, R.J.; Kierzek, A.; et al. Morphological and molecular identification reveals that waters from an isolated oasis in Tamanrasset (extreme south of Algerian sahara) are colonized by opportunistic and pollution-tolerant diatom species. Ecol. Ind. 2021, 121. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW—A suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets, Nucl. Acids Res. 2013, 41, W575–W581. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. ProgressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Length (in μm) | Width (in μm) | Density of Transapical Stria in 10 μm | Density of Longitudinal Stria in 10 μm | |
|---|---|---|---|---|
| Climaconeis cf. scalaris SZCZ 1888 | 47–53 | 5.2–7.0 | 19 | 28 |
| Climaconeis cf. scalaris SZCZ 1889 | 62–66 | 5.0–6.2 | 20 | 28 |
| Climaconeis sp. SZCZ 1890 | 47–64 | 6.2–6.8 | 20 | 24 |
| Climaconeis cf. scalaris SZCZ 1888 | |
|---|---|
| LSC | ycf45, rpl20, rpl35, orf160b, orf384, tRNA-Thr, petG, psbK, psaI, psbD, psbC, orf148, tRNA-Tyr, tRNA-Val, tRNA-Arg, tRNA-Asn, tRNA-Ser, tRNA-Met, ycf33, petF, petA, tatC, atpE, atpB, ycf3, rps18, rpl33, rps20, rpoB, rpoC1, rpoC2, rps2, tRNA-Met, tRNA-Arg, psbV, psbX, orf254, serC, orf152, tRNA-Phe, petM, petN, psbH, psbN, psbT, psbB, rps14, psaM, chlI, secG, petD, petB, tRNA-Ser, psaD, tRNA-Met, rpl12, rpl1, rpl11, tRNA-Trp, dnaB, ycf41, ycf39, cbbX, psaL, petL, ycf4, tRNA-Gly, psbE, psbF, psbL, psbJ, ycf90, tRNA-Glu, tRNA-Gly, psbZ, ycf12 |
| SSC | None |
| IR | tRNA-Lys, psaJ, psaF, psbI, atpaA, atpD, atpF, atpG, atpH, atpI, sufC, sufB, rbcL, rbcS, orf132, tRNA-Gln, tRNA-Arg, clpC, ccsA, rps6, tRNA-Asn, thiG, psaC, psbY, rpl32, tRNA-Leu, rbcR, rpl21, rpl27, secA, rpl34, ycf46, ccs1, psbA, orf123, psaA + intron, psaB, ycf89, tRNA-Pro, rrs, tRNA-Ile, tRNA-Ala, rrl, rrf, orf126, ftsH, psaE, tRNA-Cys, tRNA-Leu, serC, orf132, orf219, rps10, tufA, rps7, rps12, rpl31, rps9, rpl13, rpoA, rps11, rps13, rpl36, secY, rps5, rpl18, rpl6, rps8, rpl5, rpl24, rpl14, rps17, rpl29, rpl16, rps3, rpl22, ycf88, rps19, rpl2, rpl23, rpl4, rpl3, dnaK, groEL, orf269, psb28, thiS, tRNA-His, rps4, rps16, ffs |
| Climaconeis cf. scalaris SZCZ 1889 | |
| LSC | ycf33, tRNA-Met, tRNA-Ser, tRNA-Asp, rps2, rpoC2, rpoC1, rpoB, rps20, rpl33, rps18, ycf3, atpB, atpE, tatC, petA, petF, orf127a, psbC, psbD, psaI, psbK, petG, tRNA-Thr, ycf45, rpl20, rpl35, orf106, tRNA-Met, tRNA-Arg, psbV, psbX, tRNA-Phe, dnaB, tRNA-Trp, rpl11, rpl1, rpl12, tRNA-Met, psaD, tRNA-Ser, petB, petD, secG, chlI, psaM, rps14, psbB, psbT, psbN, psbH, petN, petM, tRNA-Lys, ycf12, psbZ, tRNA-Gly, tRNA-Glu, ycf90, psbJ, psbL, psbF, psbE, tRNA-Gly, ycf4, petL, psaL, cbbX, ycf39, ycf41 |
| SSC | None |
| IR | tRNA-Val, tRNA-Arg, ffs, ftsH, psaE, psaJ, psaF, psbI, rbcS, rbcL, sufB, sufC, atpI, atpH, atpG, atpF, atpD, atpA, psaA + intron, psaB, ycf89, tRNA-Pro, rrs, tRNA-Ile, tRNA-Ala, rrl, rrf, orf151, orf127, orf299, clpC, ccsA, rps6, tRNA-Asn, thiG, orf105, orf187, orf211, orf103, tRNA-Cys, tRNA-Leu, rps10, tufA, rps7, rps12, rpl31, rps9, rpl13, rpoA, rps11, rps13, rpl36, secY, rps5, rpl18, rpl6, rps8, rpl5, rpl24, rpl14, rps17, rpl29, rpl16, rps3, rpl22, ycf88, rps19, rpl2, rpl23, rpl4, rpl3, dnaK, groEL, psb28, thiS, tRNA-His, rps4, rps16, tRNA-Arg, tRNA-Gln, psabA, ccs1, ycf46, rpl34, secA, rpl27, rpl21, rbcR, tRNA-Leu, rpl32, psbY, psaC |
| Climaconeis sp. SZCZ 1890 | |
| LSC | psbH, psbN, psbT, psbB, rps14, psaM, chlI, secG, petD, petB, psaD, tRNA-Met, rpl12, rpl1, rpl11, tRNA-Trp, dnaB, tRNA-Phe, tRNA-Arg, tRNA-Val, tRNA-Tyr, ycf33, tRNA-Met, tRNA-Ser, tRNA-Asp, psbX, psbV, tRNA-Arg, tRNA-Met, rps2, rpoC2, rpoC1, rpoB, rps20, rpl33, rps18, ycf3, atpB, atpE, tatC, petA, petF, thiG, petG, psbK, psaI, psbD, psbC, ycf45, rpl20, rpl35, orf212, orf126, tRNA-Asn, ccsA, tRNA-Lys, ycf12, psbZ, tRNA-Gly, tRNA-Glu, ycf90, psbJ, psbL, psbF, psbE, tRNA-Gly, ycf4, petL, psaL, cbbX, ycf39, psbI, psaF, psaJ, psaA, psaB, atpA, atpD, atpF, atpG, atpH, atpI, sufC, sufB, rbcL, rbcS |
| SSC | rpl21, rpl27, secA, rpl34, ycf46, ccs1, psbA, groEL, dnaK, rpl3, rpl4, rpl23, rpl2, rps19, ycf88, rpl22, rps3, rpl16, rpl29, rps17, rpl14, rl24, rpl5, rps8, rpl6, rpl18, rps5, secY, rpl36, rps13, rps11, rpoA, rpl13, rps9, rpl31, rps12, rps7 |
| IR | petN, petM, ftsH, psaE, tRNA-Thr, ycf89, tRNA-Pro, rrs, tRNA-Ile, tRNA-Ala, rrl, rrf, tRNA-Cys, psaC, psbY, rbcR, tRNA-Leu, rpl32, psb28, thisS, tRNA-His, rps4, rps16, clpC, tRNA-Arg, tRNA-Gln, tRNA-Leu, rps10, tufA |
| Species | Genes and ORFs in the Inverted Repeat |
|---|---|
| Climaconeis sp. SZCZ 1890 | petN, petM, ftsH, psaE, tRNA-Thr, ycf89, tRNA-Pro, rns, tRNA-Ile, tRNA-Ala, rnl, rrf, tRNA-Cys, psaC, psbY, rbcR, tRNA-Leu, rpl32, psb28, thisS, tRNA-His, rps4, rps16, clpC, tRNA-Arg, tRNA-Gln, tRNA-Leu, rps10, tufA |
| Seminavis robusta MH356727 | tRNA-Pro, ycf89, rns, tRNA-Ile, tRNA-Ala, rnl, I-SroI (endonuclease), rrf * |
| Haslea nusantara MH681881 | tRNA-Pro, ycf89, rns, tRNA-Ile, tRNA-Ala, rnl, rrf, psbY |
| Haslea silbo MW645082, MW645084 | tRNA-Pro, ycf89, rns, tRNA-Ile, tRNA-Ala, rnl, rrf, psbY |
| Navicula veneta MT383645 | orf161, tRNA-Pro, ycf89, orf140, orf154, rns, tRNA-Ile, tRNA-Ala, rnl, rrf, orf383 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gastineau, R.; Davidovich, N.A.; Davidovich, O.I.; Lemieux, C.; Turmel, M.; Wróbel, R.J.; Witkowski, A. Extreme Enlargement of the Inverted Repeat Region in the Plastid Genomes of Diatoms from the Genus Climaconeis. Int. J. Mol. Sci. 2021, 22, 7155. https://doi.org/10.3390/ijms22137155
Gastineau R, Davidovich NA, Davidovich OI, Lemieux C, Turmel M, Wróbel RJ, Witkowski A. Extreme Enlargement of the Inverted Repeat Region in the Plastid Genomes of Diatoms from the Genus Climaconeis. International Journal of Molecular Sciences. 2021; 22(13):7155. https://doi.org/10.3390/ijms22137155
Chicago/Turabian StyleGastineau, Romain, Nikolaï A. Davidovich, Olga I. Davidovich, Claude Lemieux, Monique Turmel, Rafał J. Wróbel, and Andrzej Witkowski. 2021. "Extreme Enlargement of the Inverted Repeat Region in the Plastid Genomes of Diatoms from the Genus Climaconeis" International Journal of Molecular Sciences 22, no. 13: 7155. https://doi.org/10.3390/ijms22137155
APA StyleGastineau, R., Davidovich, N. A., Davidovich, O. I., Lemieux, C., Turmel, M., Wróbel, R. J., & Witkowski, A. (2021). Extreme Enlargement of the Inverted Repeat Region in the Plastid Genomes of Diatoms from the Genus Climaconeis. International Journal of Molecular Sciences, 22(13), 7155. https://doi.org/10.3390/ijms22137155