
Aversive Learning Deficits and Depressive-Like Behaviors Are Accompanied by an Increase in Oxidative Stress in a Rat Model of Fetal Alcohol Spectrum Disorders: The Protective Effect of Rapamycin

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
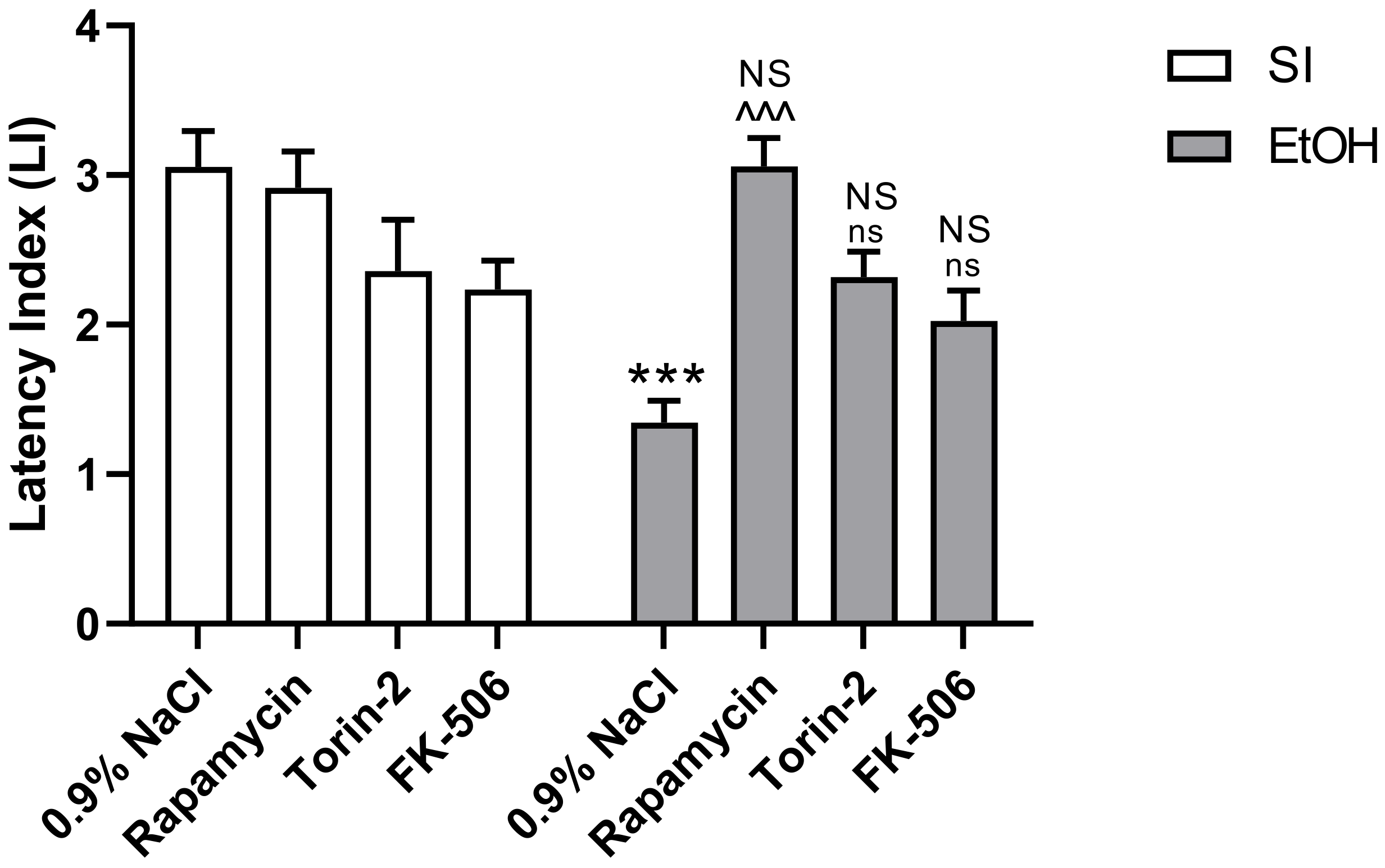
2.1. Influence of Rapamycin, Torin-2, and FK-506 Pre-Treatment before Ethanol Administration during PND4–9 on Fear Learning and Memory as Measured in the PA Test in Adult (PND60/61) Male Rats
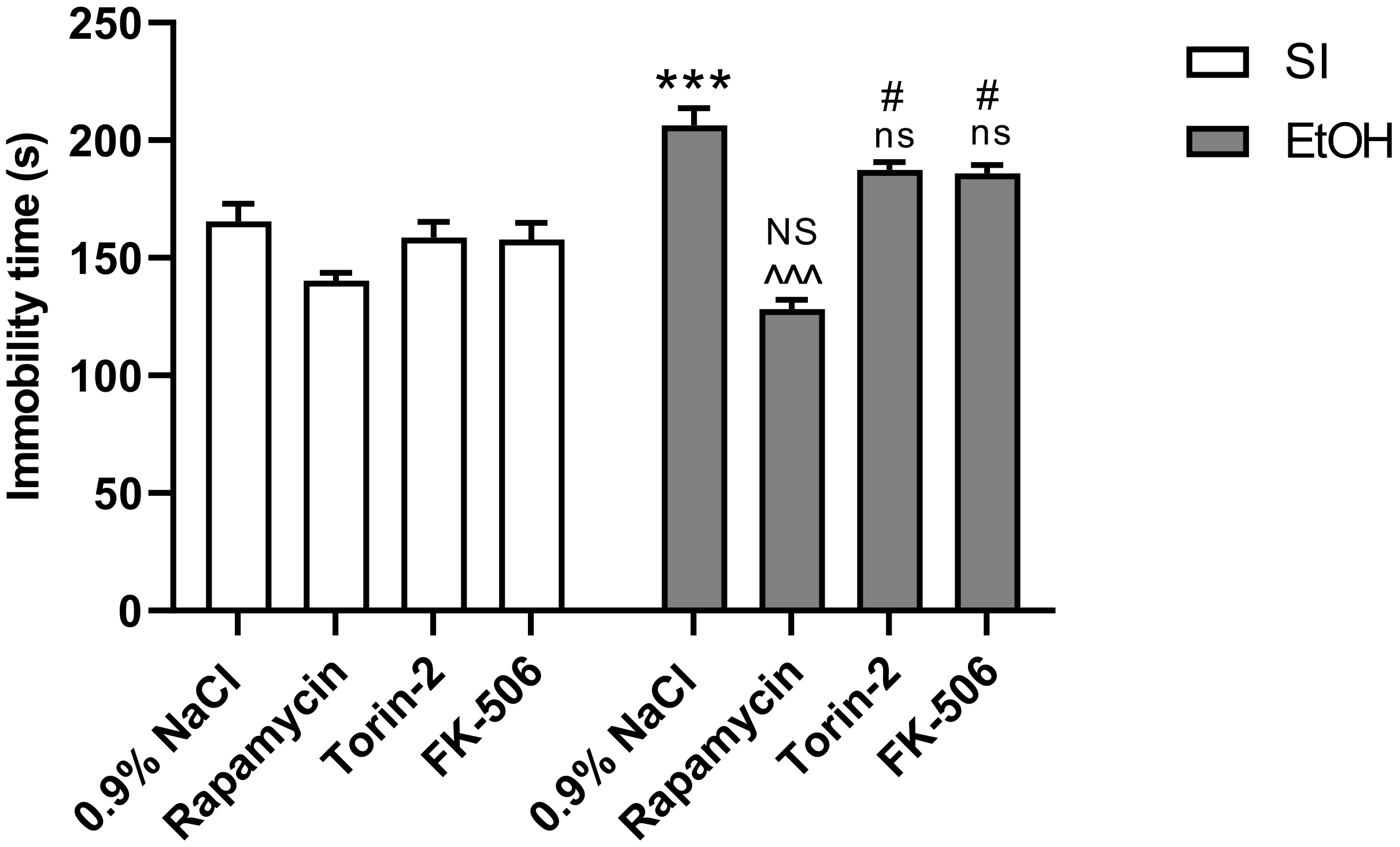
2.2. Influence of Rapamycin, Torin-2, and FK-506 Pre-Treatment before Ethanol Administration during PND4–9 on Depressive-Like Behavior as Measured in the FST Test in Adult (PND65) Male Rats
2.3. Influence of Rapamycin, Torin-2, and FK-506 Pre-Treatment before Ethanol Administration during PND4–9 on Locomotor Activity in Adult (PND59) Male Rats
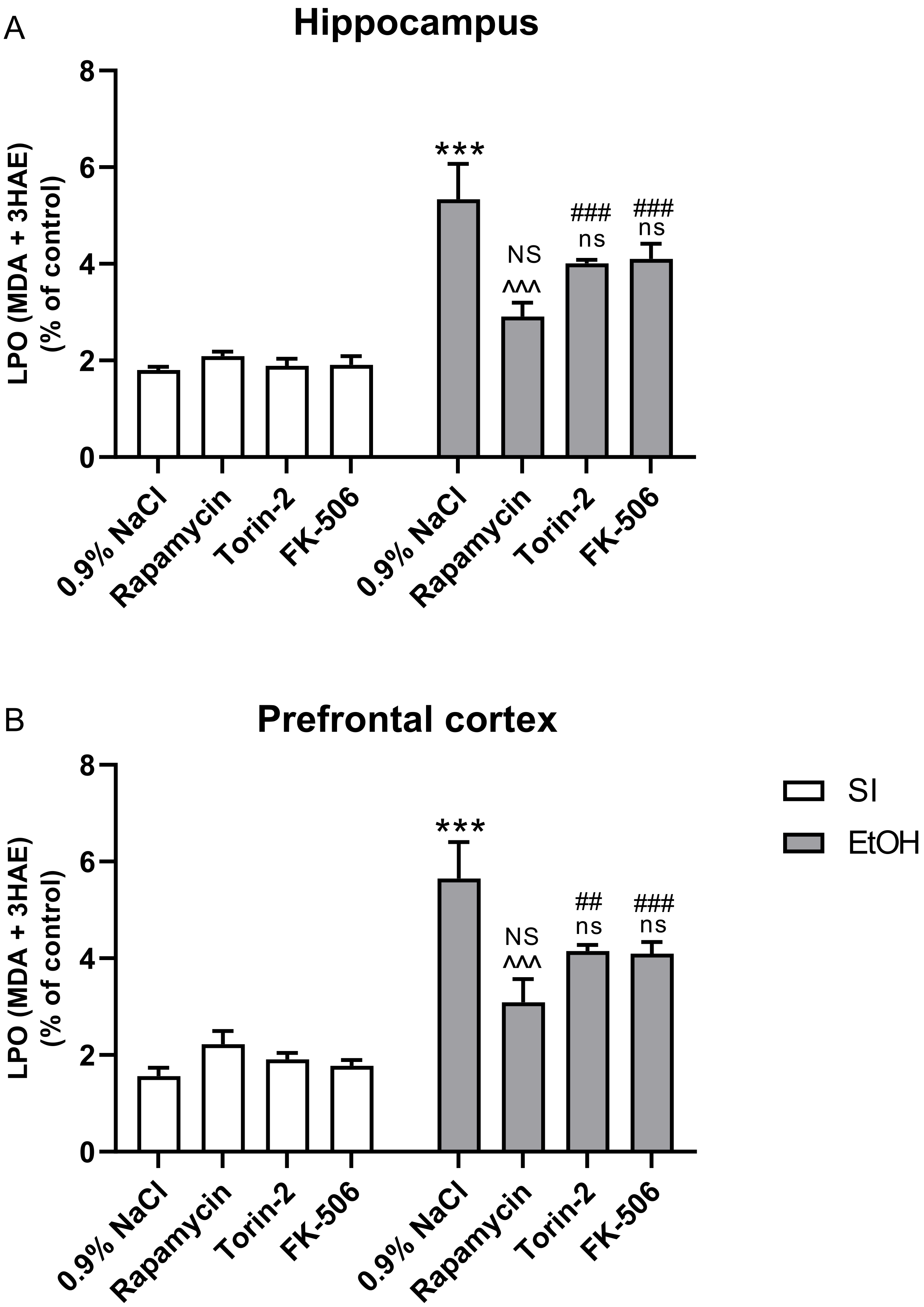
2.4. Influence of Rapamycin, Torin-2, and FK-506 Pre-Treatment before Ethanol Administration (PND4–9) on LPO Concentration in the Hippocampus and Prefrontal Cortex of Adult (PND65) Male Rats
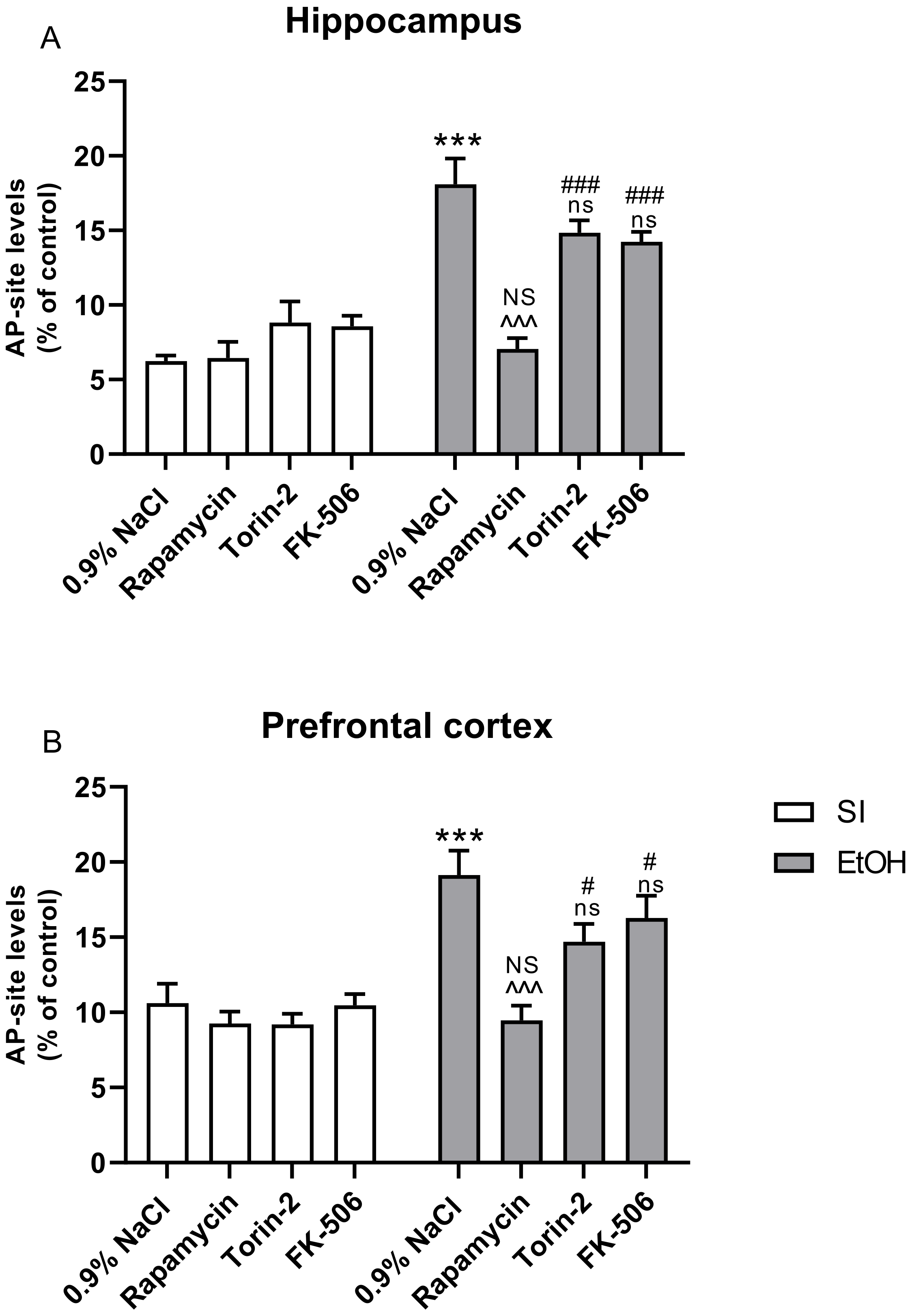
2.5. Influence of Rapamycin, Torin-2, and FK-506 Pre-Treatment before Ethanol Administration (PND4–9) on AP Site Levels in the Hippocampus and Prefrontal Cortex of Adult (PND65) Male Rats
3. Discussion
3.1. Rapamycin Prevents Ethanol-Induced Fear Learning and Memory Impairments
3.2. Rapamycin Prevents Ethanol-Induced Depressive-Like Behavior
3.3. Rapamycin Prevents Ethanol-Induced LPO and Oxidative DNA Damage
3.4. Potential Mechanism
4. Materials and Methods
4.1. Animals
4.2. Drugs and Neonatal Treatment
4.3. Experiments
4.3.1. Passive Avoidance (PA)
4.3.2. Forced Swim Test (FST)
4.3.3. Locomotor Activity (LA)
4.3.4. Determination of Biochemical Parameters
4.3.5. Determination of DNA Oxidative Damage
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilhoit, L.F.; Scott, D.A.; Simecka, B.A. Fetal Alcohol Spectrum Disorders: Characteristics, Complications, and Treatment. Community Ment. Health J. 2017, 53, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Michel, T.; Frangou, S.; Thiemeyer, D.; Camara, S.; Jecel, J.; Nara, K.; Brunklaus, A.; Zoechling, R.; Riederer, P. Evidence for oxidative stress in the frontal cortex in patients with recurrent depressive disorder—A postmortem study. Psychiatry Res. 2007, 151, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, F.; Attilia, M.L.; Spagnolo, P.A.; Rotondo, C.; Mancinelli, R.; Ceccanti, M. National Institute on Alcohol Abuse and Alcoholism and the study of fetal alcohol spectrum disorders. The International Consortium. Ann. dell’Istituto Super. Sanità 2006, 42, 4–7. [Google Scholar]
- Riley, E.P.; Infante, M.A.; Warren, K.R. Fetal Alcohol Spectrum Disorders: An Overview. Neuropsychol. Rev. 2011, 21, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Dobbing, J.; Sands, J. Comparative aspects of the brain growth spurt. Early Hum. Dev. 1979, 3, 79–83. [Google Scholar] [CrossRef]
- Hamilton, G.; Hernandez, I.; Krebs, C.; Bucko, P.; Rhodes, J. Neonatal alcohol exposure reduces number of parvalbumin-positive interneurons in the medial prefrontal cortex and impairs passive avoidance acquisition in mice deficits not rescued from exercise. Neuroscience 2017, 352, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Mohajerani, M.H.; Cherubini, E. Role of giant depolarizing potentials in shaping synaptic currents in the developing hippocampus. Crit. Rev. Neurobiol. 2006, 18, 13–23. [Google Scholar] [CrossRef]
- Lohmann, C.; Kessels, H.W. The developmental stages of synaptic plasticity. J. Physiol. 2013, 592, 13–31. [Google Scholar] [CrossRef]
- Roediger, D.J.; Krueger, A.M.; de Water, E.; Mueller, B.A.; Boys, C.A.; Hendrickson, T.J.; Schumacher, M.J.; Mattson, S.N.; Jones, K.L.; Lim, K.O.; et al. Hippocampal subfield abnormalities and memory functioning in children with fetal alcohol Spectrum disorders. Neurotoxicology Teratol. 2021, 83, 106944. [Google Scholar] [CrossRef]
- Chu, J.; Tong, M.; De La Monte, S.M. Chronic ethanol exposure causes mitochondrial dysfunction and oxidative stress in immature central nervous system neurons. Acta Neuropathol. 2007, 113, 659–673. [Google Scholar] [CrossRef]
- Dembele, K.; Yao, X.-H.; Chen, L.; Nyomba, B.L.G. Intrauterine ethanol exposure results in hypothalamic oxidative stress and neuroendocrine alterations in adult rat offspring. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R796–R802. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- He, J.; Kang, L.; Wu, T.; Zhang, J.; Wang, H.; Gao, H.; Zhang, Y.; Huang, B.; Liu, W.; Kou, Z.; et al. An Elaborate Regulation of Mammalian Target of Rapamycin Activity Is Required for Somatic Cell Reprogramming Induced by Defined Transcription Factors. Stem Cells Dev. 2012, 21, 2630–2641. [Google Scholar] [CrossRef] [PubMed]
- Pla, A.F.; Pascual, M.; Renaupiqueras, J.; Guerri, C. TLR4 mediates the impairment of ubiquitin-proteasome and autophagy-lysosome pathways induced by ethanol treatment in brain. Cell Death Dis. 2014, 5, e1066. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-A. Neuronal Autophagy: A Housekeeper or a Fighter in Neuronal Cell Survival? Exp. Neurobiol. 2012, 21, 1–8. [Google Scholar] [CrossRef]
- Shen, H.-M.; Codogno, P. Autophagy is a survival force via suppression of necrotic cell death. Exp. Cell Res. 2012, 318, 1304–1308. [Google Scholar] [CrossRef] [PubMed]
- Loos, B.; Engelbrecht, A.-M.; Lockshin, R.A.; Klionsky, D.J.; Zakeri, Z. The variability of autophagy and cell death susceptibility. Autophagy 2013, 9, 1270–1285. [Google Scholar] [CrossRef]
- Luo, J. Autophagy and ethanol neurotoxicity. Autophagy 2014, 10, 2099–2108. [Google Scholar] [CrossRef]
- Chen, T.; Wan, H.; Guo, A.; Chen, J.; Long, Y.; Shen, L.; Yu, J.; Zhao, J.; Pei, G. Rapamycin and other longevity-promoting compounds enhance the generation of mouse induced pluripotent stem cells. Aging Cell 2011, 10, 908–911. [Google Scholar] [CrossRef]
- Kotlinska, J.H.; Lopatynska-Mazurek, M.; Gawel, K.; Gabka, P.; Jenda-Wojtanowska, M.; Kruk-Slomka, M.; Marszalek-Grabska, M.; Danilczuk, Z.; Kedzierska, E.; Talarek, S.; et al. Impact of the metabotropic glutamate receptor7 (mGlu7) allosteric agonist, AMN082, on fear learning and memory and anxiety-like behavior. Eur. J. Pharmacol. 2019, 858, 172512. [Google Scholar] [CrossRef]
- Gold, P.E. The use of avoidance training in studies of modulation of memory storage. Behav. Neural Biol. 1986, 46, 87–98. [Google Scholar] [CrossRef]
- Roozendaal, B.; McGaugh, J.L. Memory modulation. Behav. Neurosci. 2011, 125, 797–824. [Google Scholar] [CrossRef]
- Poetsch, A.R. The genomics of oxidative DNA damage, repair, and resulting mutagenesis. Comput. Struct. Biotechnol. J. 2020, 18, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Niedernhofer, L.J.; Daniels, J.S.; Rouzer, C.A.; Greene, R.E.; Marnett, L.J. Malondialdehyde, a Product of Lipid Peroxidation, Is Mutagenic in Human Cells. J. Biol. Chem. 2003, 278, 31426–31433. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ke, Z.; Xu, M.; Liao, M.; Wang, X.; Qi, Y.; Zhang, T.; Frank, J.A.; Bower, K.A.; Shi, X.; et al. Autophagy is a protective response to ethanol neurotoxicity. Autophagy 2012, 8, 1577–1589. [Google Scholar] [CrossRef]
- Kane, C.J.; Chang, J.Y.; Roberson, P.K.; Garg, T.K.; Han, L. Ethanol exposure of neonatal rats does not increase biomarkers of oxidative stress in isolated cerebellar granule neurons. Alcohol 2008, 42, 29–36. [Google Scholar] [CrossRef]
- Ramachandran, V.; Perez, A.; Chen, J.; Senthil, D.; Schenker, S.; Henderson, G.I. In utero ethanol exposure causes mitochondrial dysfunction, which can result in apoptotic cell death in fetal brain: A potential role for 4-hydroxynonenal. Alcohol. Clin. Exp. Res. 2001, 25, 862–871. [Google Scholar] [CrossRef]
- Smith, A.M.; Zeve, D.R.; Grisel, J.J.; Chen, W.-J.A. Neonatal alcohol exposure increases malondialdehyde (MDA) and glutathione (GSH) levels in the developing cerebellum. Dev. Brain Res. 2005, 160, 231–238. [Google Scholar] [CrossRef]
- Riley, E.P.; Lochry, E.A.; Shapiro, N.R. Lack of response inhibition in rats prenatally exposed to alcohol. Psychopharmacology 1979, 62, 47–52. [Google Scholar] [CrossRef]
- Rosett, H.L. A Clinical Perspective of the Fetal Alcohol Syndrome. Alcohol. Clin. Exp. Res. 1980, 4, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Teruel, A.; Tobeña, A. Revisiting the role of anxiety in the initial acquisition of two-way active avoidance: Pharmacological, behavioural and neuroanatomical convergence. Neurosci. Biobehav. Rev. 2020, 118, 739–758. [Google Scholar] [CrossRef]
- Lopatynska-Mazurek, M.; Pankowska, A.; Gibula-Tarlowska, E.; Pietura, R.; Kotlinska, J. Rapamycin Improves Recognition Memory and Normalizes Amino-Acids and Amines Levels in the Hippocampal Dentate Gyrus in Adult Rats Exposed to Ethanol during the Neonatal Period. Biomolecules 2021, 11, 362. [Google Scholar] [CrossRef]
- Lopatynska-Mazurek, M.; Antolak, A.; Grochecki, P.; Gibula-Tarlowska, E.; Bodzon-Kulakowska, A.; Listos, J.; Kedzierska, E.; Suder, P.; Silberring, J.; Kotlinska, J. Rapamycin Improves Spatial Learning Deficits, Vulnerability to Alcohol Addiction and Altered Expression of the GluN2B Subunit of the NMDA Receptor in Adult Rats Exposed to Ethanol during the Neonatal Period. Biomolecules 2021, 11, 650. [Google Scholar] [CrossRef]
- Gibula-Tarlowska, E.; Korz, V.; Lopatynska-Mazurek, M.; Chlopas-Konowalek, A.; Grochecki, P.; Kalaba, P.; Dragacevic, V.; Kotlinski, R.; Kujawski, R.; Szulc, M.; et al. CE-123, a novel dopamine transporter inhibitor, attenuates locomotor hyperactivity and improves cognitive functions in rat model of fetal alcohol spectrum disorders. Behav. Brain Res. 2021, 410, 113326. [Google Scholar] [CrossRef]
- Sase, A.; Aher, Y.D.; Saroja, S.R.; Ganesan, M.K.; Sase, S.; Holy, M.; Höger, H.; Bakulev, V.; Ecker, G.F.; Langer, T.; et al. A heterocyclic compound CE-103 inhibits dopamine reuptake and modulates dopamine transporter and dopamine D1-D3 containing receptor complexes. Neuropharmacology 2016, 102, 186–196. [Google Scholar] [CrossRef]
- Famy, C.; Streissguth, A.P.; Unis, A.S. Mental Illness in Adults with Fetal Alcohol Syndrome or Fetal Alcohol Effects. Am. J. Psychiatry 1998, 155, 552–554. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, M.J.; Paley, B. The Relationship of Prenatal Alcohol Exposure and the Postnatal Environment to Child Depressive Symptoms. J. Pediatr. Psychol. 2006, 31, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, K.K.; Sheema, S.; Paz, R.D.; Samudio-Ruiz, S.L.; Laughlin, M.H.; Spence, N.E.; Roehlk, M.J.; Alcon, S.N.; Allan, A. Fetal alcohol spectrum disorder-associated depression: Evidence for reductions in the levels of brain-derived neurotrophic factor in a mouse model. Pharmacol. Biochem. Behav. 2008, 90, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Wilcoxon, J.S.; Kuo, A.G.; Disterhoft, J.F.; Redei, E.E. Behavioral deficits associated with fetal alcohol exposure are reversed by prenatal thyroid hormone treatment: A role for maternal thyroid hormone deficiency in FAE. Mol. Psychiatry 2005, 10, 961–971. [Google Scholar] [CrossRef]
- Carneiro, L.M.; Diógenes, J.P.L.; Vasconcelos, S.M.; Aragao, G.F.; Noronha, E.C.; Gomes, P.B.; Viana, G.S. Behavioral and neurochemical effects on rat offspring after prenatal exposure to ethanol. Neurotoxicology Teratol. 2005, 27, 585–592. [Google Scholar] [CrossRef]
- Kessler, R.C. Epidemiology of women and depression. J. Affect. Disord. 2003, 74, 5–13. [Google Scholar] [CrossRef]
- Kessler, R.C.; Akiskal, H.S.; Ames, M.; Birnbaum, H.; Greenberg, P.; Jin, R.; Merikangas, K.R.; Simon, G.E.; Wang, P.S. Prevalence and effects of mood disorders on work performance in a nationally representative sample of U.S. workers. Am. J. Psychiatry. 2006, 163, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Dowling, R.J.; Topisirovic, I.; Fonseca, B.D.; Sonenberg, N. Dissecting the role of mTOR: Lessons from mTOR inhibitors. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2010, 1804, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Codeluppi, S.; Fernández-Zafra, T.; Sándor, K.; Kjell, J.; Liu, Q.; Abrams, M.; Olson, L.; Gray, N.S.; Svensson, C.I.; Uhlén, P. Interleukin-6 Secretion by Astrocytes Is Dynamically Regulated by PI3K-mTOR-Calcium Signaling. PLoS ONE 2014, 9, e92649. [Google Scholar] [CrossRef]
- Ballatori, N.; Krance, S.M.; Notenboom, S.; Shi, S.; Tieu, K.; Hammond, C.L. Glutathione dysregulation and the etiology and progression of human diseases. Biol. Chem. 2009, 390, 191–214. [Google Scholar] [CrossRef]
- Akers, K.; Kushner, S.; Leslie, A.; Clareke, L.; van der Kooy, D.; Lerch, J.P.; Frankland, P.W. Fetal alcohol exposure leads to abnormal olfactory bulb development and impaired odor discrimination in adult mice. Mol. Brain. 2011, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Moreland, N.; La Grange, L.; Montoya, R. Impact of in utero exposure to EtOH on corpus callosum development and paw preference in rats: Protective effects of silymarin. BMC Complement. Altern. Med. 2002, 2, 10. [Google Scholar] [CrossRef]
- Petkov, V.V.; Stoianovski, D.; Petkov, V.D.; Vyglenova, I. Lipid peroxidation changes in the brain in fetal alcohol syndrome. Bull. Exp. Biol. Med. 1992, 113, 500–502. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 1–31. [Google Scholar] [CrossRef]
- Vaváková, M.; Ďuračková, Z.; Trebatická, J. Markers of Oxidative Stress and Neuroprogression in Depression Disorder. Oxidative Med. Cell. Longev. 2015, 2015, 1–12. [Google Scholar] [CrossRef]
- Storey, K.B. Oxidative stress: Animal adaptations in nature. Braz. J. Med. Biol. Res. 1996, 29, 1715–1733. [Google Scholar]
- Şimsek, Ş.; Yüksel, T.; Kaplan, I.; Uysal, C.; Aktaş, H. The Levels of Cortisol and Oxidative Stress and DNA Damage in Child and Adolescent Victims of Sexual Abuse with or without Post-Traumatic Stress Disorder. Psychiatry Investig. 2016, 13, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Yakes, F.M.; Van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc. Natl. Acad. Sci. USA 1997, 94, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Englander, E.W. Brain capacity for repair of oxidatively damaged DNA and preservation of neuronal function. Mech. Ageing Dev. 2008, 129, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Shenoy, A.; Cui, J.; Huang, W.; Liu, P.K. In situ detection of AP sites and DNA strand breaks bearing 3’-phosphate termini in ischemic mouse brain. FASEB J. 2000, 14, 407–417. [Google Scholar] [CrossRef]
- Filler, K.; Lyon, D.; Bennett, J.; McCain, N.; Elswick, R.; Lukkahatai, N.; Saligan, L.N. Association of mitochondrial dysfunction and fatigue: A review of the literature. BBA Clin. 2014, 1, 12–23. [Google Scholar] [CrossRef]
- Brocardo, P.S.; Gil-Mohapel, J.; Christie, B.R. The role of oxidative stress in fetal alcohol spectrum disorders. Brain Res. Rev. 2011, 67, 209–225. [Google Scholar] [CrossRef]
- Bhatia, S.; Drake, D.M.; Miller, L.; Wells, P.G. Oxidative stress and DNA damage in the mechanism of fetal alcohol spectrum disorders. Birth Defects Res. 2019, 111, 714–748. [Google Scholar] [CrossRef]
- De La Monte, S.; Wands, J. Chronic gestational exposure to ethanol impairs insulin-stimulated survival and mitochondrial function in cerebellar neurons. Cell. Mol. Life Sci. 2002, 59, 882–893. [Google Scholar] [CrossRef]
- Bukiya, A.N. Fetal Cerebral Artery Mitochondrion as Target of Prenatal Alcohol Exposure. Int J Environ Res Public Health 2019, 16, 1586. [Google Scholar] [CrossRef]
- Soscia, S.J.; Tong, M.; Xu, X.J.; Cohen, A.C.; Chu, J.; Wands, J.R.; De La Monte, S.M. Chronic gestational exposure to ethanol causes insulin and IGF resistance and impairs acetylcholine homeostasis in the brain. Cell. Mol. Life Sci. 2006, 63, 2039–2056. [Google Scholar] [CrossRef]
- Xu, J.; Yeon, J.E.; Chang, H.; Tison, G.; Chen, G.J.; Wands, J.; de la Monte, S. Ethanol Impairs Insulin-stimulated Neuronal Survival in the Developing Brain. J. Biol. Chem. 2003, 278, 26929–26937. [Google Scholar] [CrossRef]
- Kim, I.; Lemasters, J.J. Mitochondrial degradation by autophagy (mitophagy) in GFP-LC3 transgenic hepatocytes during nutrient deprivation. Am. J. Physiol. Physiol. 2011, 300, C308–C317. [Google Scholar] [CrossRef]
- Kim, I.; Rodriguez-Enriquez, S.; Lemasters, J.J. Selective degradation of mitochondria by mitophagy. Arch. Biochem. Biophys. 2007, 462, 245–253. [Google Scholar] [CrossRef]
- Davis-Anderson, K.L.; Wesseling, H.; Siebert, L.M.; Lunde-Young, E.R.; Naik, V.D.; Steen, H.; Ramadoss, J. Fetal regional brain protein signature in FASD rat model. Reprod. Toxicol. 2018, 76, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One Drug, Many Effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef]
- Goodlett, C.R.; Peterson, S.D. Sex Differences in Vulnerability to Developmental Spatial Learning Deficits Induced by Limited Binge Alcohol Exposure in Neonatal Rats. Neurobiol. Learn. Mem. 1995, 64, 265–275. [Google Scholar] [CrossRef]
- Johnson, T.B.; Goodlett, C.R. Selective and enduring deficits in spatial learning after limited neonatal binge alcohol exposure in male rats. Alcohol. Clin. Exp. Res. 2002, 26, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Dursun, I.; Jakubowska-Dogru, E.; Elibol-Can, B.; Van Der List, D.; Chapman, B.; Qi, L.; Berman, R.F. Effects of early postnatal alcohol exposure on the developing retinogeniculate projections in C57BL/6 mice. Alcohol 2013, 47, 173–179. [Google Scholar] [CrossRef]
- Lin, J.; Liu, L.; Wen, Q.; Zheng, C.; Gao, Y.; Peng, S.; Tan, Y.; Li, Y. Rapamycin prevents drug seeking via disrupting reconsolidation of reward memory in rats. Int. J. Neuropsychopharmacol. 2013, 17, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Vogel, K.R.; Ainslie, G.R.; Gibson, K.M. mTOR inhibitors rescue premature lethality and attenuate dysregulation of GABAergic/glutamatergic transcription in murine succinate semialdehyde dehydrogenase deficiency (SSADHD), a disorder of GABA metabolism. J. Inherit. Metab. Dis. 2016, 39, 877–886. [Google Scholar] [CrossRef] [PubMed]
- McAllister-Williams, R.H.; Bertrand, D.; Rollema, H.; Hurst, R.S.; Spear, L.P.; Kirkham, T.C.; Steckler, T.; Capdevielle, D.; Boulenger, J.-P.; Höschl, C.; et al. Passive Avoidance. In Encyclopedia of Psychopharmacology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2010; pp. 960–967. [Google Scholar]
- Paydar, M.J.; Zakeri, M.; Norouzi, A.; Rastegar, H.; Mirazi, N.; Dehpour, A.R. Involvement of nitric oxide in granisetron improving effect on scopolamine-induced memory impairment in mice. Brain Res. 2012, 1429, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Anton, G.; Blavet, N.; Jalfre, M. Behavioural despair in rats: A new model sensitive to antidepressant treatments. Eur. J. Pharmacol. 1978, 47, 379–391. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Distance Traveled (m) ± SEM |
|---|---|---|
| 1. 0.9% NaCl + SI | 8 | 52.87 ± 2.631 (NS) |
| 2. Rapamycin + SI | 8 | 48.39 ± 4.486 (NS) |
| 3. Torin-2 + SI | 8 | 50.89 ± 1.982 (NS) |
| 4. FK-506 + SI | 8 | 50.84 ± 2.036 (NS) |
| 5. 0.9% NaCl + EtOH | 8 | 50.15 ± 2.72 1(NS) |
| 6. Rapamycin + EtOH | 8 | 53.52 ± 0.650 (NS) |
| 7. Torin-2 + EtOH | 8 | 53.80 ± 0.927 (NS) |
| 8. FK-506 + EtOH | 8 | 55.93 ± 3.058 (NS) |
| Number of Groups | Intraperitoneally (i.p.) Injection | Intragastric (i.g.) Intubation | N |
|---|---|---|---|
| 1. | 0.9%NaCl | SI | 8 |
| 2. | Rapamycin | SI | 8 |
| 3. | Torin-2 | SI | 8 |
| 4. | FK-506 | SI | 8 |
| 5. | 0.9%NaCl | EtOH | 8 |
| 6. | Rapamycin | EtOH | 8 |
| 7. | Torin-2 | EtOH | 8 |
| 8. | FK-506 | EtOH | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopatynska-Mazurek, M.; Komsta, L.; Gibula-Tarlowska, E.; Kotlinska, J.H. Aversive Learning Deficits and Depressive-Like Behaviors Are Accompanied by an Increase in Oxidative Stress in a Rat Model of Fetal Alcohol Spectrum Disorders: The Protective Effect of Rapamycin. Int. J. Mol. Sci. 2021, 22, 7083. https://doi.org/10.3390/ijms22137083
Lopatynska-Mazurek M, Komsta L, Gibula-Tarlowska E, Kotlinska JH. Aversive Learning Deficits and Depressive-Like Behaviors Are Accompanied by an Increase in Oxidative Stress in a Rat Model of Fetal Alcohol Spectrum Disorders: The Protective Effect of Rapamycin. International Journal of Molecular Sciences. 2021; 22(13):7083. https://doi.org/10.3390/ijms22137083
Chicago/Turabian StyleLopatynska-Mazurek, Malgorzata, Lukasz Komsta, Ewa Gibula-Tarlowska, and Jolanta H. Kotlinska. 2021. "Aversive Learning Deficits and Depressive-Like Behaviors Are Accompanied by an Increase in Oxidative Stress in a Rat Model of Fetal Alcohol Spectrum Disorders: The Protective Effect of Rapamycin" International Journal of Molecular Sciences 22, no. 13: 7083. https://doi.org/10.3390/ijms22137083
APA StyleLopatynska-Mazurek, M., Komsta, L., Gibula-Tarlowska, E., & Kotlinska, J. H. (2021). Aversive Learning Deficits and Depressive-Like Behaviors Are Accompanied by an Increase in Oxidative Stress in a Rat Model of Fetal Alcohol Spectrum Disorders: The Protective Effect of Rapamycin. International Journal of Molecular Sciences, 22(13), 7083. https://doi.org/10.3390/ijms22137083