1. Introduction
Alkaloids are a large group of important natural compounds, and, thanks to their prophylactic or therapeutic values, they are undoubtedly a stable player in the field of medical treatment for many diseases. Approximately 50–60% of the new drugs approved between 1981 and 2010 either originated from plants or were prepared by variations of the chemical structures of the compounds previously described in plants [
1]. Alkaloids isolated from Amaryllidaceae plants comprise a large group of approximately 600 naturally occurring isoquinoline alkaloids with a vast structural diversity [
2]. Therefore, the large number of Amaryllidaceae alkaloids (AA) have been divided into several structurally divergent skeleton types according to their biosynthetic origin, and ring structure and named after a representative alkaloid from the class [
2,
3].
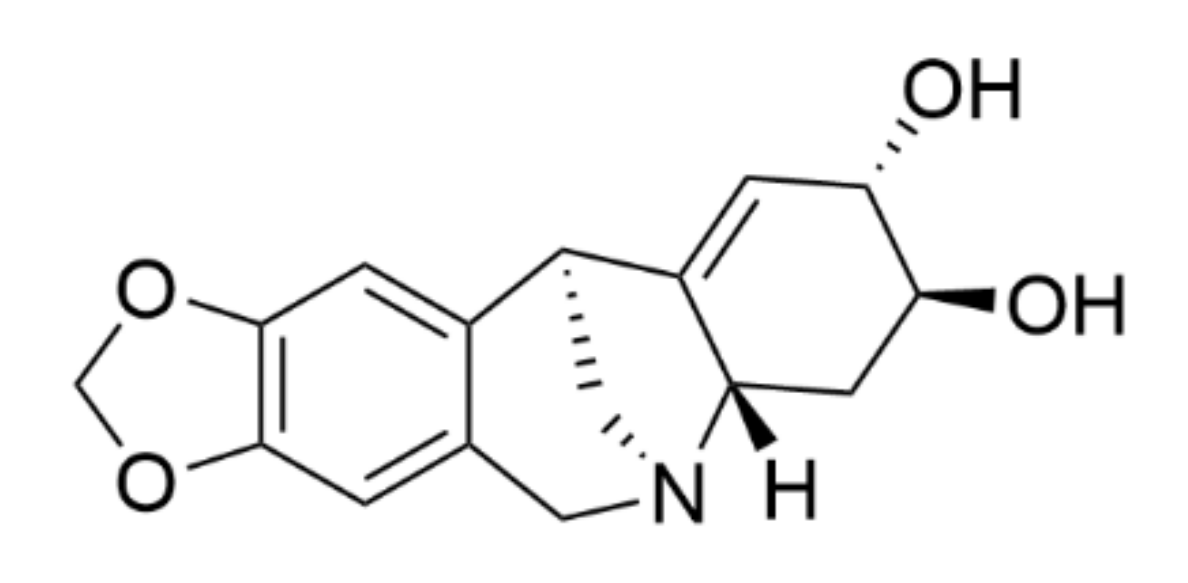
Pancracine (
Figure 1) belongs to the montanine types of AA, which possess a unique pentacyclic 5,11-methanomorphanthridine skeleton ring. At present, fourteen known AA share the 5,11-methanomorphanthridine scaffold, which is believed to be a significant carrier of the biological effects of montanine-type alkaloids [
4,
5]. Pancracine is distinct from other alkaloids of montanine types because of the presence of a double bond between C1 and C11a and two hydroxyls on stereocenters at C2 and C3 in the E ring of the 5,11-methanomorphanthridine framework [
6]. Among the montanine skeleton type, montanine, coccinine, manthidine, and manthine were the first alkaloid members isolated from the
Haemanthus species of the Amaryllidaceae plant family by Wildman et al. [
7]. Thirteen years later, in 1968, Wildman and coworkers described another AA of a montanine type, pancracine, which they isolated from
Pancratium maritimum,
Narcissus poeticus, and
Rhodophiala bifida [
4]. Since then, attention has turned to investigations performed to gain a deeper understanding of montanine-type alkaloids, including either the possibility of isolation from other plants of the Amaryllidaceae family [
6], preparing them by chemical de novo synthesis [
5], or exploring their biological activity [
6,
8,
9,
10].
In our previous work, we screened the cytotoxicity of AA of various structural types isolated from bulbs of
Narcissus cv. Professor Einstein against a panel of eight cancer cell lines [
6]. Among them, pancracine has shown the ability to reduce the proliferation of cancer cells of different histotypes with IC
50 values reaching into the micromolar range after 48 h of treatment. MOLT-4, A549, HT-29, MCF-7, and SAOS-2 are sensitive cell lines with IC
50 values below 3 μM, whereas Jurkat A2780 and HeLa are more resistant, with IC
50 values between 3 and 6 μM [
9]. Consistently, other structure–activity studies investigating the antiproliferative effect of pancracine isolated from
Pancratium canariense described its strong inhibitory effect from 4.3 ± 0.7 µM to 9.1 ± 1.0 µM on the growth of four cancer cell lines derived from ovarian (A2780), lung (SW1573), breast (T47-D), and colon (WiDr) carcinoma [
9]. Moreover, a recent study reported the preparation of semisynthetic montanine-type alkaloid derivatives, which were screened in vitro for their antiproliferative activities against a panel of six cancer cell lines. Although the parent alkaloid of the montanine-type, manthine, was the most effective, the C2-OH and C2-indole-substituted 5,11-methanomorphanthridine derivatives also showed good antiproliferative activity [
5]. In what concerns the other bioactivities, there were two studies reporting the antibiotic, antifungal, and antiparasitic effects of pancracine [
11,
12]. Antimicrobial activity testing using an agar diffusion technique and bacteria species
Staphylococcus aureus,
Escherichia coli, and
Pseudomonas aeruginosa and the yeast
Candida albicans displayed good inhibitory activity of pancracine against
Staphylococcus aureus and
Pseudomonas aeruginosa and moderate activity against
Candida albicans [
11]. The interesting biological activity was complemented by a study describing the antiprotozoal activities of pancracine isolated from
Narcissus angustifolius subsp.
transcarpathicus against
Trypanosoma brucei rhodesiense,
Trypanosoma cruzi, and
Plasmodium falciparum [
12].
The AA and its derivatives exhibited various pharmacological activities. Most notably, the cytotoxic activity of the several AA were noteworthy [
3,
13,
14]. Since recent studies have begun to uncover the mechanisms sustaining the cytotoxicity of AA, deeper investigations on some AA are still fragmentary. The initial pilot studies have suggested that the micromole cytotoxic potency of pancracine towards cancer cell lines deserves further in-depth investigation. Moreover, there is fundamental lack of reports on the mode of action of pancracine in vitro, greatly hindering further investigation on the montanine type of AA. The aim of this study was to reveal the mechanisms underlying the proliferative inhibition after pancracine treatment by evaluating its effect on cell cycle progression, apoptosis, and related signal transduction pathways.
3. Discussion
The unique chemical structure of montanine alkaloids, with their significant biological activity, together with their rarity in nature, has led to a synthetic effort to produce ample amounts for biological studies and subsequent use in clinical therapy [
15,
16,
17,
18,
19,
20,
21]. This also includes the preparation of montanine-type alkaloids by rearrangement of the haemantamine-type ring system [
21,
22]. Unfortunately, without understanding the molecular mechanisms of their actions, it is not possible to take advantage of their promising biological activity over other AAs. If we focus on the anticancer potential of montanine alkaloids, only the IC
50 values have been available so far, with pancracine having values from 2.29 ± 0.43 µM to 5.15 ± 0.34 µM, using tumor cells derived from A549 (lung adenocarcinoma), HT-29 (colon adenocarcinoma), A2780 (ovarian carcinoma), HeLa (cervix carcinoma), MCF-7 (breast carcinoma), SAOS-2 (osteosarcoma), Jurkat (acute T-cell leukemia), and MOLT-4 (acute lymphoblastic leukemia) [
6]. The IC
50 values in the low-micromolar range were found to correspond closely to those found in an earlier study published by Cedrón et al. [
9], which described the cytotoxic effect of pancracine on tumor cells with IC
50 values ranging from 4.3 ± 0.7 µM to 9.1 ± 1.0 µM. Montanine, which is the main representative of this group of substances, seems to be more active than pancracine, because its IC
50 values ranged from 1.04 ± 0.14 µM to 2.30 ± 0.45 µM in the study of Al Shammari et al. [
23]. Since pancracine differs from montanine by only one substituent on the E ring of the 5,11-methanomorphanthridine structure, it can be hypothesized that the molecular mechanism of action of these substances may be similar.
By selecting one apoptosis-resistant cancer cell line and one apoptosis-sensitive leukemia cell line as an experimental model counterpart, the aim of this study was to approximate the effect of pancracine on these cells, including events that lead to either cell cycle perturbation or apoptosis.
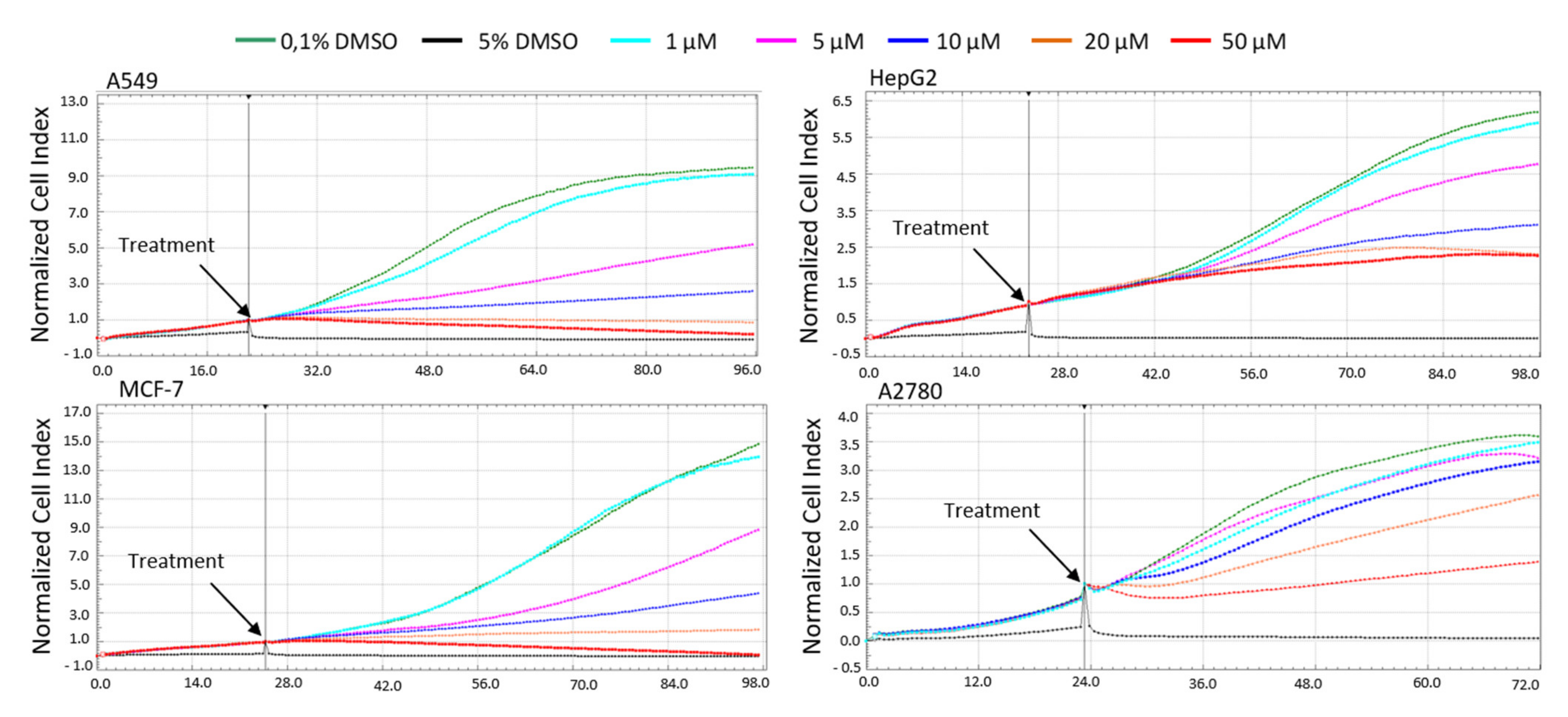
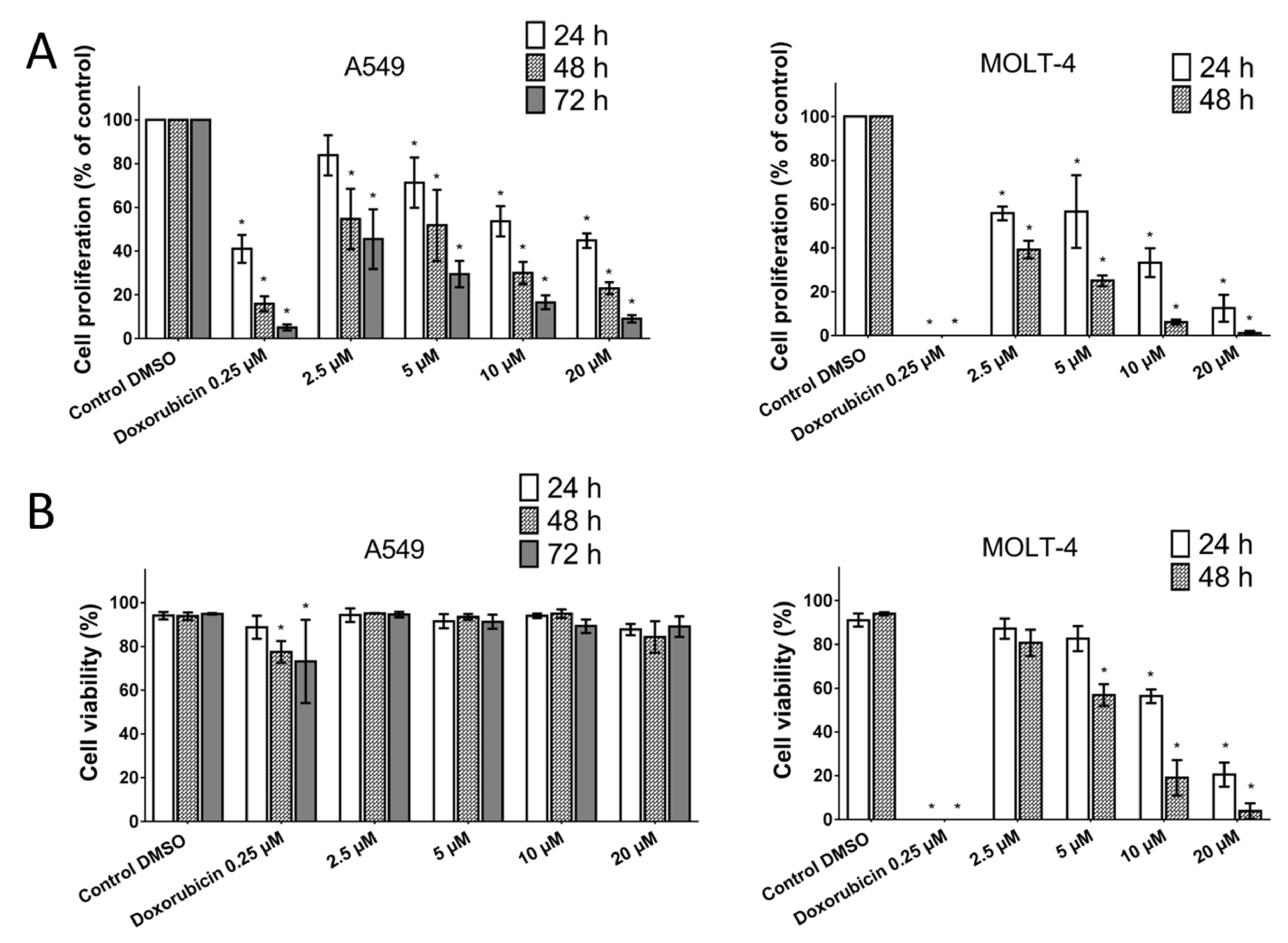
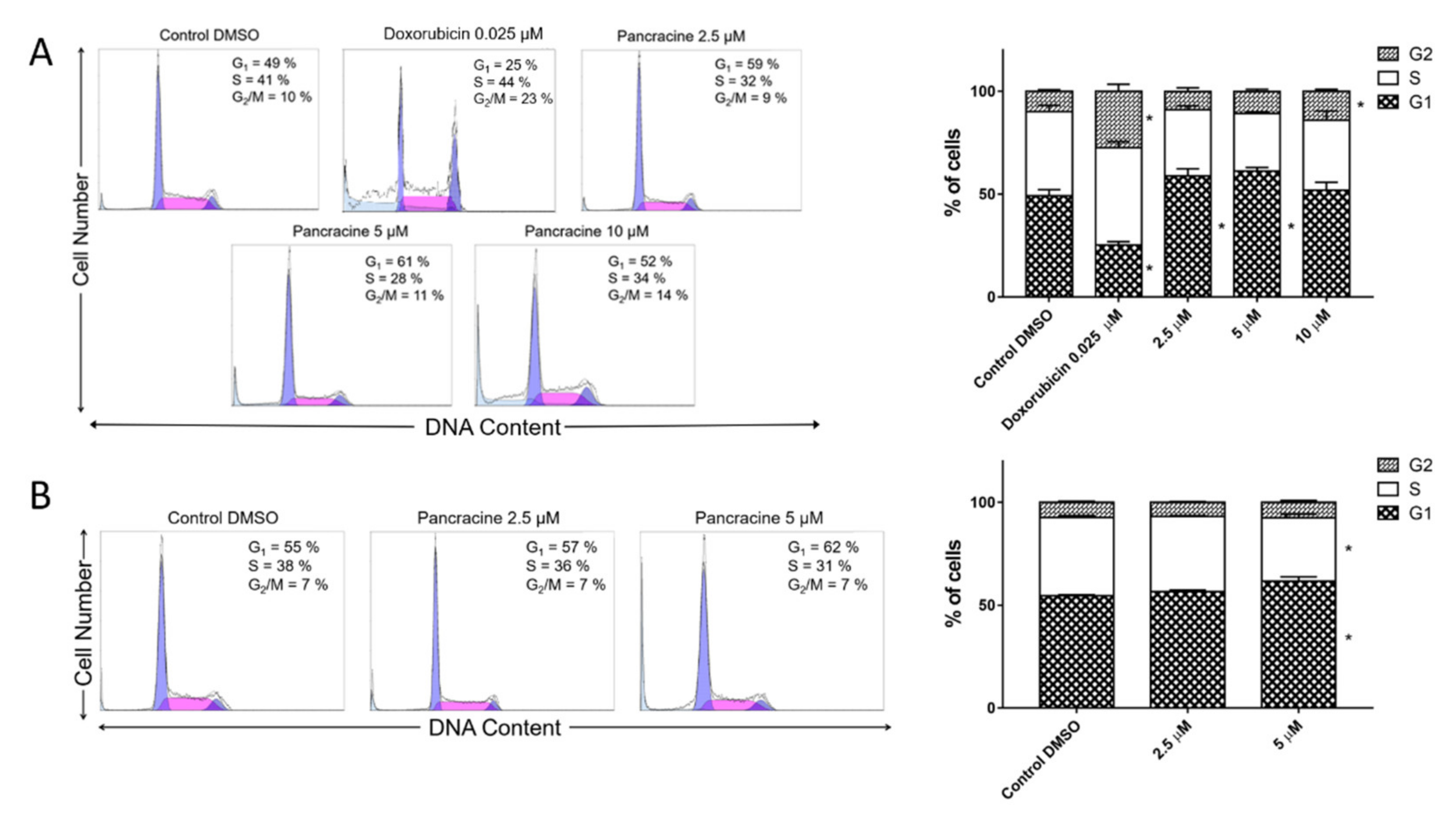
The A549 cancer cell line represents non-small cell lung cancer (NSCLC) with very poor prognosis and with high chemoresistance to the standard cytotoxic drug treatments. Considering the IC
50 values of pancracine for A549 cells (2.29 ± 0.43 µM) [
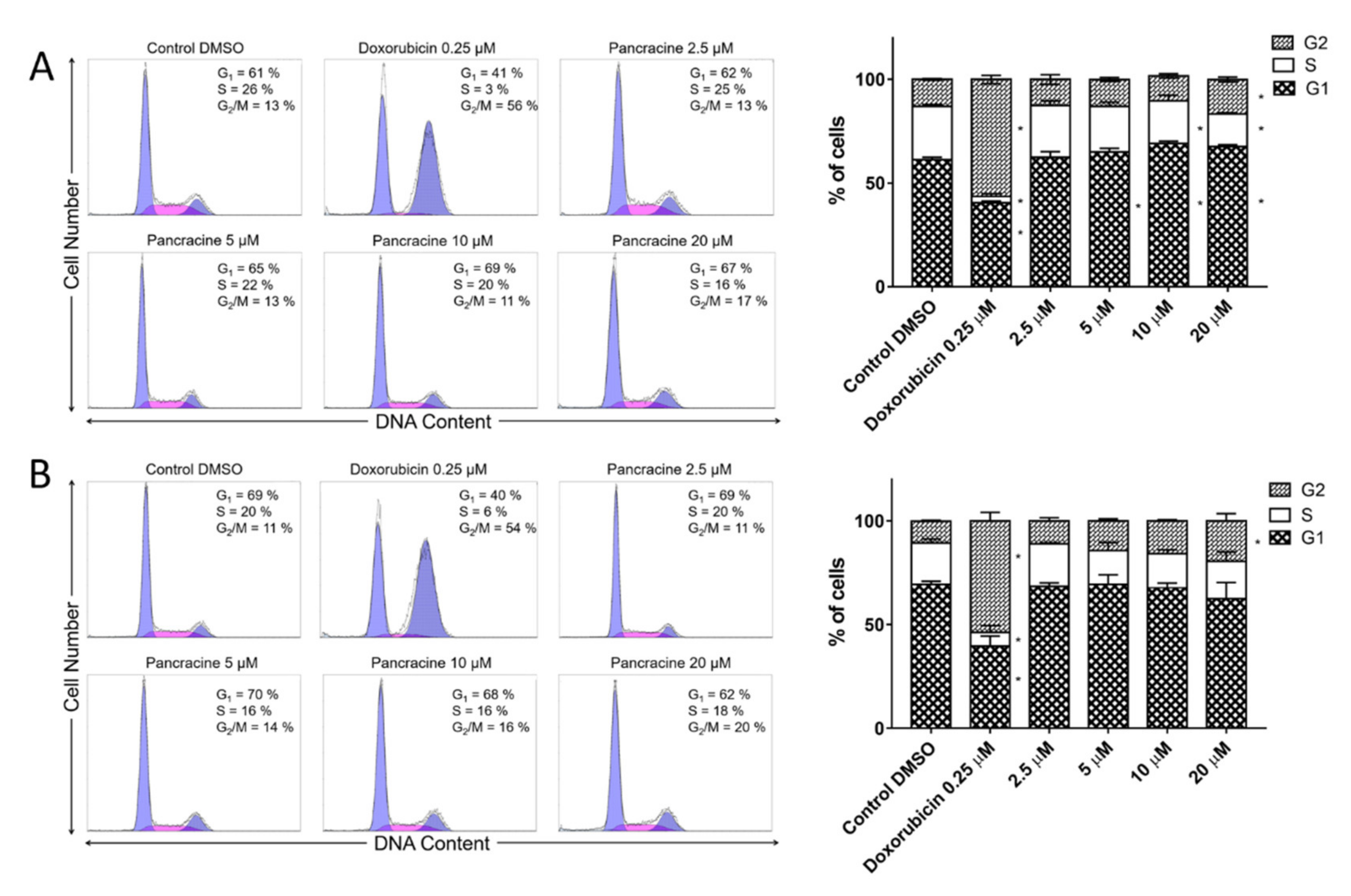
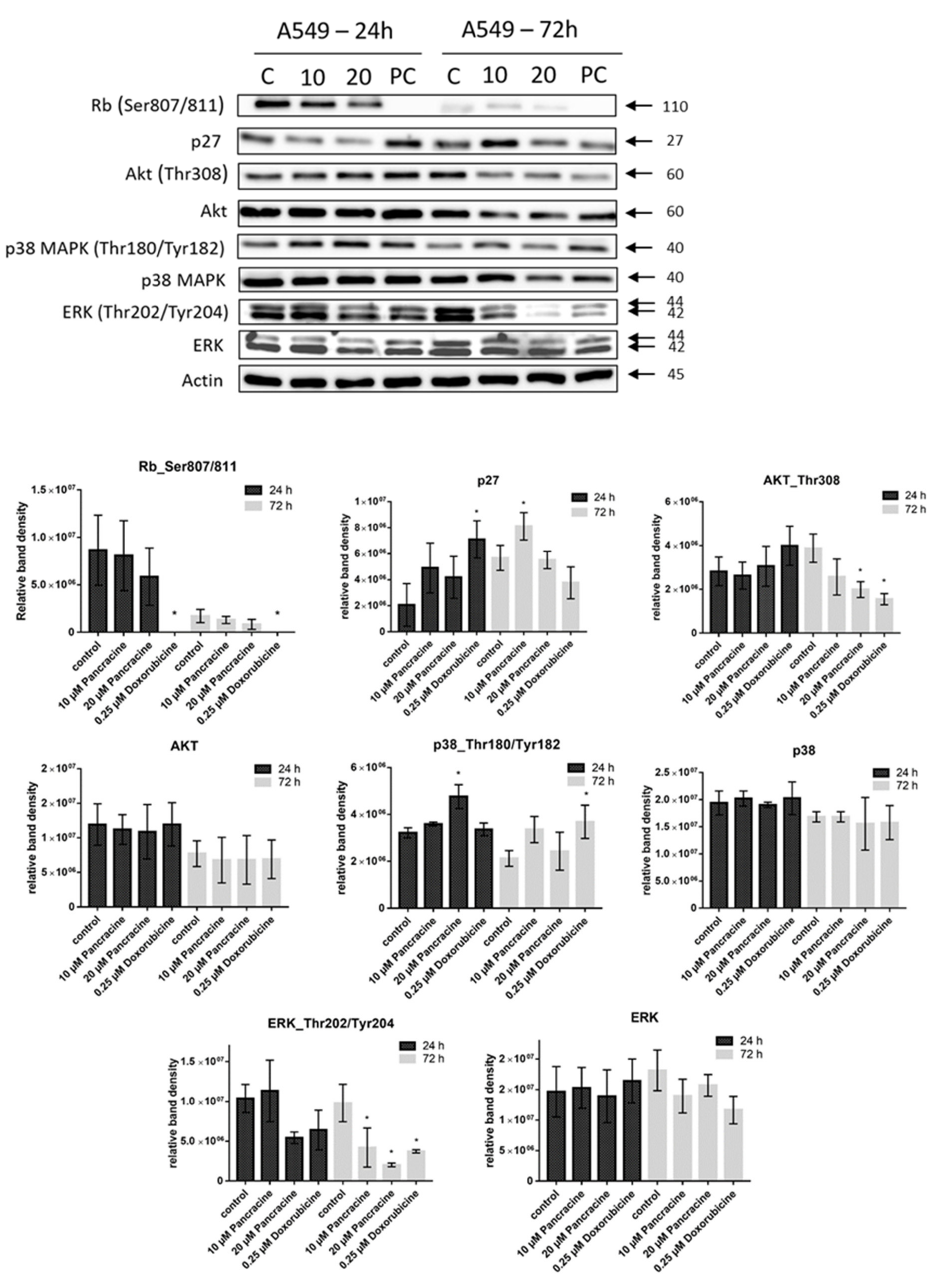
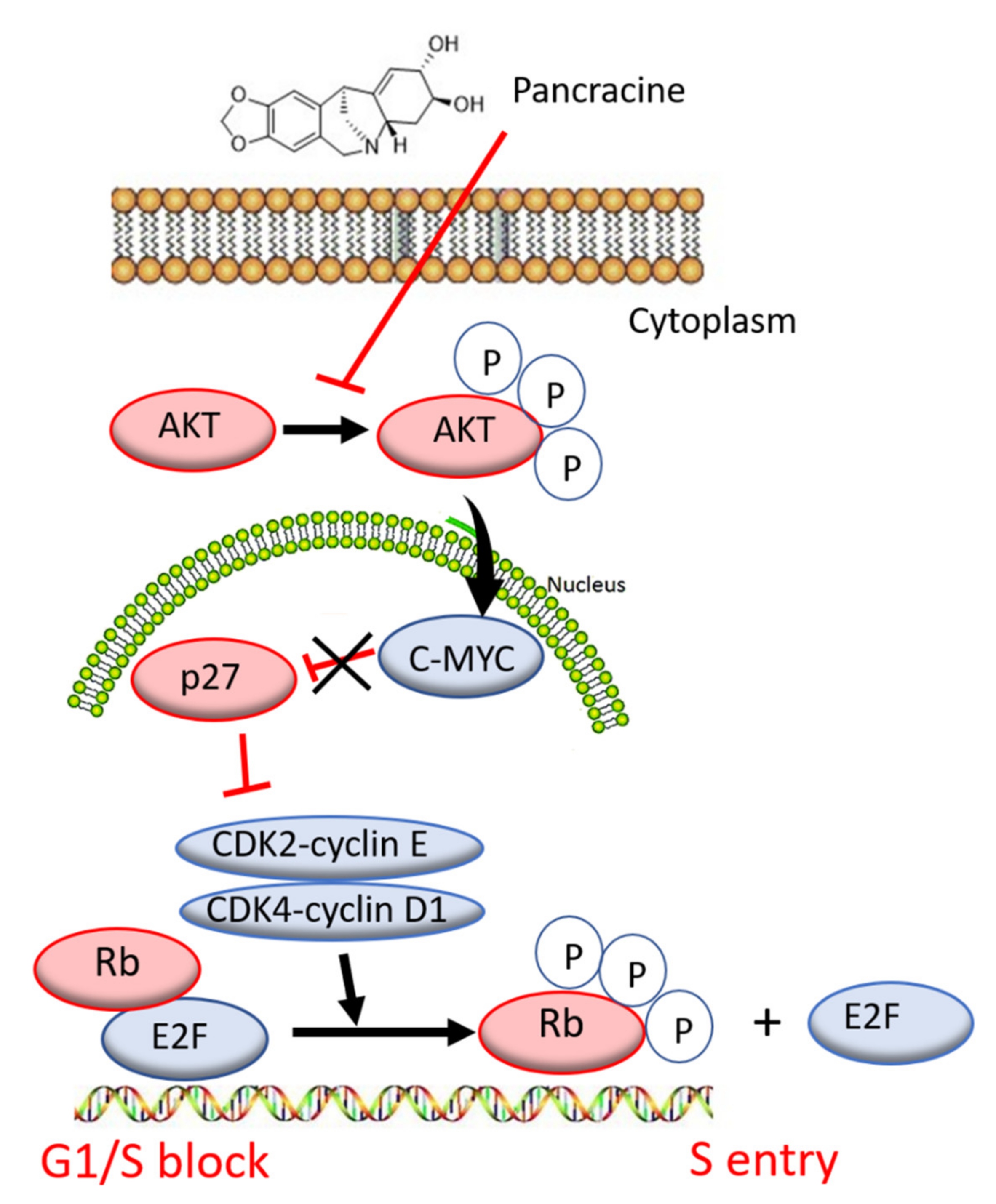
6], the strong antiproliferative effect of this compound on the A549 cancer cell line determined in this study by a real-time cell monitoring method is fully consistent with the previous findings. This growth inhibitory effect persists for 96 h. We extended this real-time cell proliferation measuring with the end-point Trypan blue exclusion test. A statistically significant antiproliferative effect on A549 cells was already demonstrated using a concentration of 2.5 mM at all time intervals (24, 48, and 72 h). The impact on cell viability of the A549 cells was negligible, as concentrations up to 20 μM failed to reduce the A549 viability below 80%. According to the real-time cell monitoring method, a 50-µM concentration of pancracine seemed to be lethal for A549 cancer cells. Cell cycle arrest at the G1/S transition occurs by the activation of several signaling pathways. One of these acts through the activation of MAPK systems by the phosphorylation of p38 MAPK, which phosphorylates several nuclear factors, such as tumor suppressor p53, activating transcription factor ATF-2, myocyte enhancer factor MEF-2, and transcription factor Myc [
24]. Another possibility is signaling through the Akt kinase. The phosphorylated pAkt kinase inhibits the p27 protein [
25]. p27 is a tumor suppressor that inhibits the phosphorylation of Rb by inhibition of the cyclin-dependent kinase (CDK) complex and, as a result, prevents the separation of transcription factor E2F from Rb, which, in total, prevents the transcription of genes required for G1/S transition [
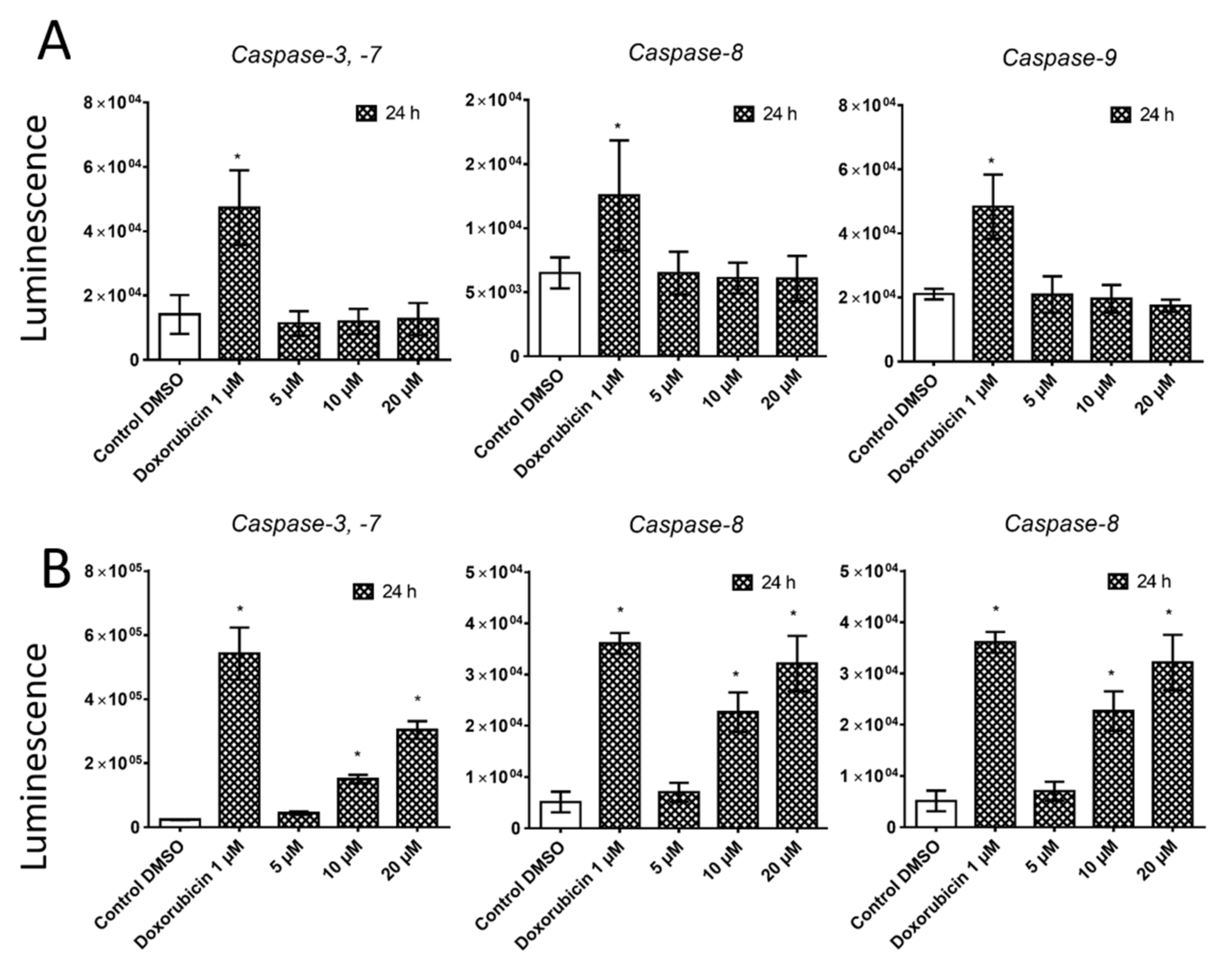
25]. Our results show that the proliferation of A549 cells is inhibited by cell cycle arrest in the G1 phase through the upregulation of phosphorylated p38 MAPK after 24 h and either a 10- or 20-µM pancracine treatment. Cell cycle arrest initiated through the downregulation of phosphorylated pRb protein, upregulation of p27 concomitantly with the downregulation of Akt kinase phosphorylated on Thr308, and downregulation of ERK was observed after a longer time interval (72 h) using the same experimental conditions. Besides, apoptosis is not induced at the cellular level, as proved by the activity of caspase-3/7, -8, and -9 in A549 cells. For a better overview,
Figure 10 shows the schematic representation of proposed AKT/p27/pRb upstream signaling events involved in the antiproliferative activity of pancracine against A549 lung adenocarcinoma cells.
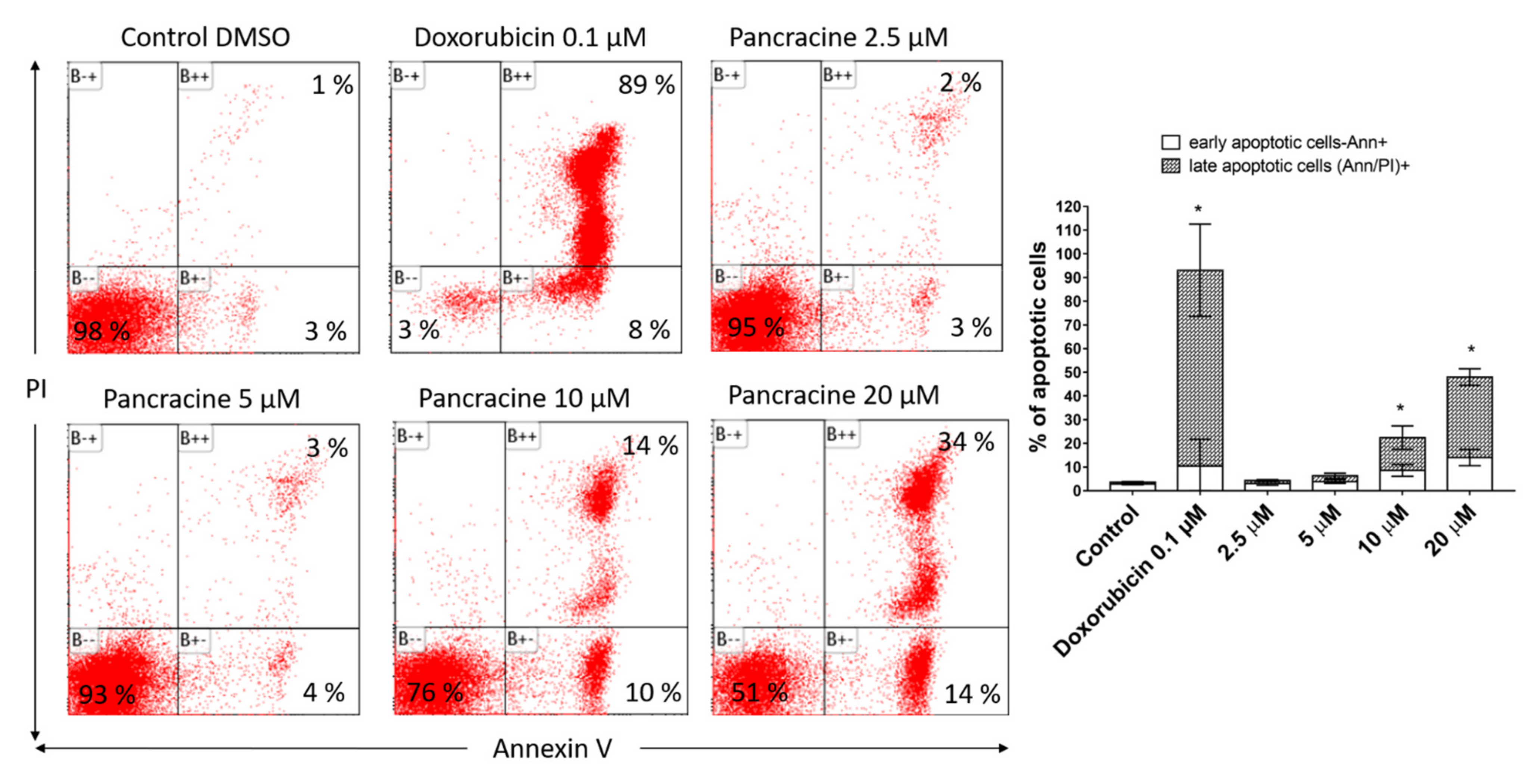
MOLT-4 is a T-lymphoblastic cell line originally derived from the peripheral blood of a 19-year-old patient with acute lymphoblastic leukemia in relapse. These cells responded to pancracine treatment with a half-maximal inhibitory concentration (IC
50) of 2.71 ± 0.25 µM [
6]. A similar strong antiproliferative effect as that seen on A549 cells was observed after the pancracine treatment of MOLT-4 cells. However, unlike in the A549 cell line, a significant strong activity in decreasing the viability of leukemic cells was observed. The viability of MOLT-4 cells decreased, even after treatment with 2.5-μM pancracine, and treatment with the higher dose of 20 μM led to a decrease of viable cells down to 20%. Due to the greater potency of pancracine with decreasing viability rates at higher concentrations, only the lower concentrations of 2.5, 5, and 10 μM after 24 h and 2.5 and 5 μM after 48 h of treatment could be used to detect the percentage of leukemic MOLT-4 cells in the cell cycle. Several publications showed that leukemic cells undergo apoptotic cell death [
26,
27,
28]. MOLT-4 cells die due to ionizing radiation and other DNA damage-inducing agents by the apoptotic process of the activation of caspase-8 and -9 and release of cytochrome c [
27]. In this study, the leukemic MOLT-4 cells died due to apoptosis activation at 5 μM, increasing the activity of effector caspase-3/7 and the activation of caspase -9 responsible for the mitochondrial apoptosis pathway and caspase-8 responsible for the pathway through death receptors. The apoptosis of MOLT-4 cells significantly increased 24 h after the exposure to a 10-μM dose of pancracine, as detected using Annexin V and PI staining. Using a concentration of 20 μM, 50% of the cells were either in the early or late phase of apoptosis. Activation of the ATM/Chk2/p53 signaling pathway was described in MOLT-4 cells after ionizing radiation exposure [
29]. Additionally, the cytotoxic effect of valproic acid is accompanied by the activation of p21 and upregulation of p53 phosphorylated on Ser392 [
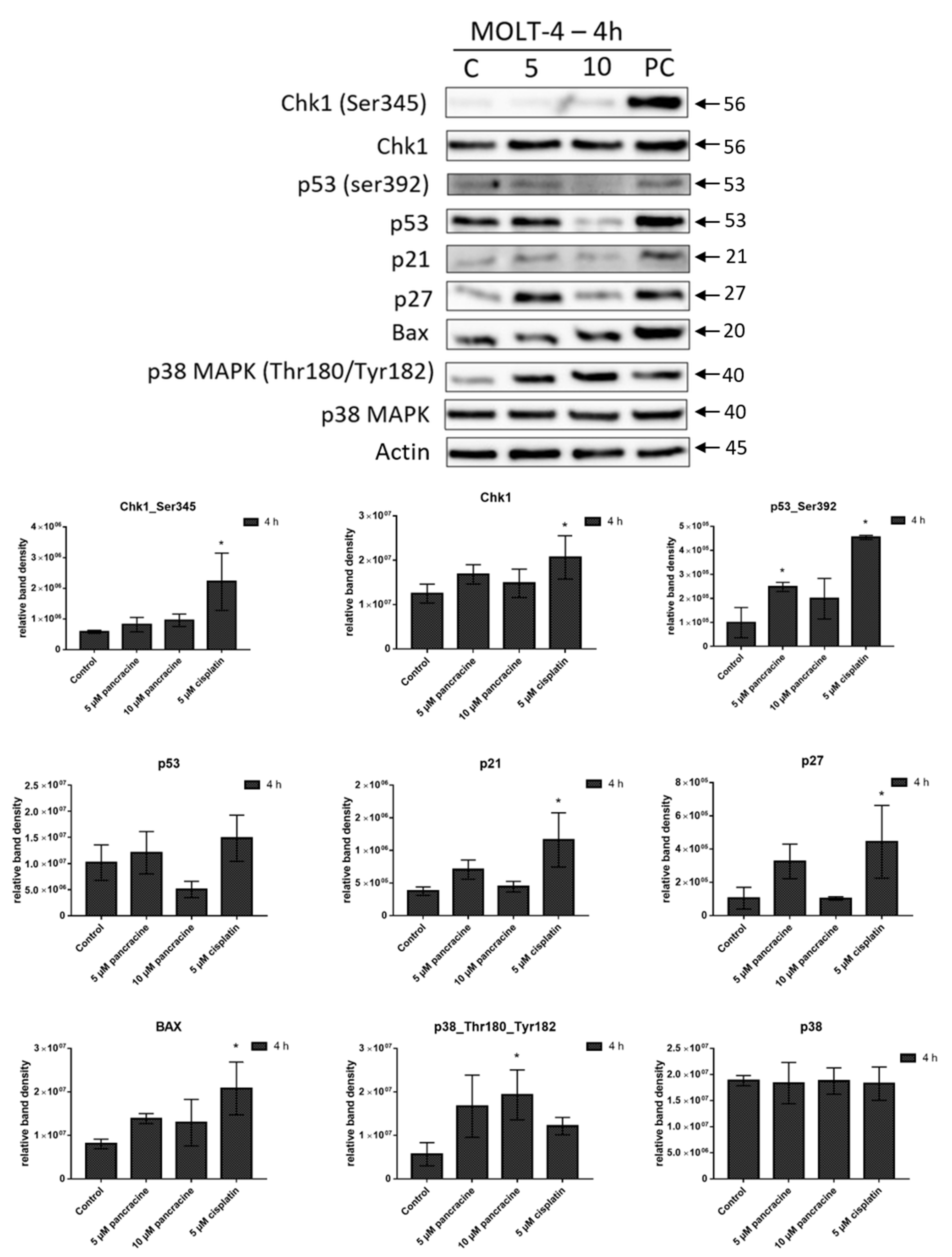
28]. In our study, the significant apoptosis-inducing effect of pancracine was accompanied by the signaling pathway, including an increase of p27 protein, Bax, and activation of p38 MAPK through phosphorylation at Thr180 and Tyr182, and seemed to be activated by the upregulation of p53 phosphorylated at Ser392.
As far as we know, a study of the molecular mechanism of action of montanine-type AA has not yet been published, but when considering the biosynthesis of this group of alkaloids, one can consider other AA with similar structures. Haemantamine, with its α-crinane structure, also belongs to the isoquinoline group of Amaryllidaceae alkaloids, and its activity has been recently reviewed [
3]. Haemantamine, even at a dose of 5 µM, also induced apoptosis in p53-deficient acute T-cell leukemia Jurkat cells [
14]. In addition, alkaloids based on the montanine skeleton can be prepared by a rearrangement of the haemantamine (α-crinane)-type ring system [
21,
22]. The mechanism of action of haemantamine lies in the targeting of the A-site cleft on the large ribosomal subunit and rearranging rRNA to halt the elongation phase of translation, which leads to repressing cancer cell growth [
30].
In conclusion, the later stages of preclinical testing of new chemotherapeutics have always preceded a deeper knowledge of the molecular mechanisms of their cytostatic and cytotoxic activity. Pancracine, with its strong inhibitory effect on tumor cell proliferation and viability, may be one of the potential pharmacophore scaffolds in cancer therapy. Additionally, this initial pilot study presented here attempts to open a more detailed study of the mechanisms of action of montanine-type AA.
4. Materials and Methods
4.1. Cell Culture and Culture Conditions
Experiments were performed with selected human tumor cell lines MOLT-4 (acute lymphoblastic leukemia), A549 (lung adenocarcinoma), A2780 (ovarian carcinoma), MCF-7 (breast adenocarcinoma), and HepG2 (hepatocellular carcinoma), which were purchased from the European Collection of Cell Cultures (ECACC, Salisbury, UK) and cultured in accordance with the provider’s culture method guidelines. All cell lines were maintained under standard cell culture conditions at 37 °C in a humidified incubator in an atmosphere of 5% CO2 and 95% air. Cells were passaged every 2 to 3 days to obtain exponential growth. Cells in the maximum range of 20 passages were used for this study.
4.2. Cell Treatment
Pancracine (purity > 95%) was provided by the ADINACO Research Group from the Department of Pharmaceutical Botany, Faculty of Pharmacy in Hradec Kralove, Charles University, Hradec Kralove, Czech Republic. Results of the
1H-NMR, HPLC/UV, and GC/MS analyses are shown in
Supplementary Figures S2–S4. The compound was isolated from fresh bulbs of
Narcissus L. cv. Professor Einstein (Amaryllidaceae) within a detailed phytochemical study [
5]. Fresh stock solutions of pancracine in concentrations of 50 mM were dissolved in dimethyl sulfoxide (DMSO) (Sigma-Aldrich, St. Louis, MO, USA). Stock solutions were freshly prepared before use. For the experiments, the stock solutions were diluted with the complete culture medium to create final concentrations of 1–50 μM (50 μM is the highest concentration used in xCELLigence measurements), making sure that the concentration of DMSO was < 0.1% to avoid toxic effects on the cells. Negative control cells were sham-treated with a DMSO vehicle only (0.1%; control). Cells treated with 5% DMSO; cisplatin (Sigma-Aldrich, St. Louis, MO, USA) at 5 µM; or doxorubicin (Sigma-Aldrich, St. Louis, MO, USA) at 0.25 nM, 0.25 µM, and 1 µM were used as a positive control.
4.3. Screening for Antiproliferative Activity Using the xCELLigence System
The xCELLigence system (Roche, Basel, Switzerland and ACEA Biosciences, San Diego, CA, USA) was used to monitor cell adhesion, proliferation, and cytotoxicity. It was connected and tested by s resistor plate before the RTCA single-plate station was placed inside the incubator at 37 °C with an atmosphere containing 5% CO2. First, the seeding concentration for the experiments was optimized for each cell line. After seeding, the respective number of cells in 190 µL of medium per well of the E-plate 96 and the proliferation, attachment, and spreading of the cells were monitored every 30 min by the xCELLigence system. Approximately 24 h after seeding, when the cells were in the log growth phase, they were exposed in triplicates to 10 µL of sterile deionized water containing pancracine to obtain final concentrations of 1–50 μM. Controls received sterile deionized water + DMSO with a final concentration of 0.1%. Cells treated with 5% DMSO were used as a positive control. Growth curves were normalized to the time point of treatment. Evaluations were performed using xCELLigence 1.2.1 software (Roche, Basel, Switzerland and ACEA Biosciences, San Diego, CA, USA).
4.4. Proliferation and Viability Measurement Using Trypan Blue Exclusion Test
Cell proliferation and viability of MOLT-4 and A549 cells were monitored 24, 48, and 72 h after treatment with 2.5, 5, 10, and 20 μM of pancracine in the case of A549 cells (1.5 × 105 cell in 5 mL placed in 25-cm2 Falcon flasks) and 24 and 48 h in the case of MOLT-4 cells (1 × 106 cell in 5 mL placed in 25-cm2 Falcon flasks) with the same concentration range of pancracine. Cells treated with 0.25-µM doxorubicin were used as a positive control. Cell membrane integrity was determined using the Trypan blue exclusion technique—mixing 10 μL of 0.4% Trypan blue and 10 μL of cell suspension. The Trypan blue dye exclusion test is used to determine the percentage of viable cells and number of viable cells present in a cell suspension. It is based on the principle that live cells possess intact cell membranes that exclude certain dyes, such as Trypan Blue, whereas dead cells do not. The assay is easy to perform; a cell suspension is mixed with dye and then visually examined by microscope to determine whether cells take up or exclude the dye. Then, the individual cells are observed using a bright-field Nikon Eclipse E200 light microscope (Nikon, Tokyo, Japan); a viable cell will have a clear cytoplasm, whereas a nonviable cell will have a blue cytoplasm. In this assay, a hemocytometer (Bürker counting chamber) was used to determine exactly the number of cells in a suspension. We counted 25 squares (=1 horizontal line + 1 vertical line + 1 square). We counted only cells that were within the 0.2 mm × 0.2 mm squares. We did not count cells that overlapped over the border. The surface of each square was 0.2 mm × 0.2 mm = 0.04 mm2. With a height of 0.1 mm, the volume was 0.004 mm3. Therefore, 25 squares corresponded to a volume of 0.1 mm3 (=0.1 μL). Multiplication by 10,000 gave 1 mL. The calculation of proliferation in % followed this formula: (number of viable cells after treatment/number of viable cells in control DMSO) multiplied by 100. The calculation of viability in % followed this formula: (number of viable cells after treatment/number of total cells) multiplied by 100.
4.5. Cell Cycle Distribution and Internucleosomal DNA Fragmentation Analysis
The cells were washed with ice-cold PBS and fixed with 70% ethanol. In order to detect low molecular weight fragments of DNA, the cells were incubated for 5 min at room temperature in a buffer (192 mL of 0.2-M Na2HPO4 + 8 mL of 0.1-M citric acid, pH 7.8) and then labeled with propidium iodide in Vindelov’s solution for 1 h at 37 °C. The DNA content was determined using a CyAn flow cytometer (Beckman Coulter, Miami, FL, USA) with an excitation wavelength of 488 nm. The data were analyzed using Multicycle AV software (Phoenix Flow Systems, San Diego, CA, USA).
4.6. Activity of Caspases
The induction of programmed cell death was determined by monitoring the activities of caspases-3/7, caspase-8, and caspase-9 by Caspase-Glo Assays (Promega, Madison, WI, USA) 24 h after treatment with 5, 10, and 20 μM of pancracine. Cells treated with 1 µM of doxorubicin were used as a positive control. The assay provides a proluminogenic substrate in an optimized buffer system. The addition of a Caspase-Glo Reagent results in cell lysis, followed by caspase cleavage of the substrate and the generation of a luminescent signal. A total of 1 × 104 cells were seeded per well using a 96-well plate format (Sigma-Aldrich, St. Louis, MO, USA). After treatment, the Caspase-Glo Assay Reagent was added to each well (50 μL/well) and incubated for 30 min before luminescence was measured using a Tecan Infinite M200 microplate reader (Tecan Group, Männedorf, Switzerland).
4.7. Analysis of Apoptosis
Apoptosis was determined by flow cytometry using an Alexa Fluor®488 Annexin V/Dead Cell Apoptosis kit (Life Technologies, Grand Island, NY, USA) in accordance with the manufacturer’s instructions. The Alexa Fluor®488 Annexin V/Dead Cell Apoptosis kit employs the property of Alexa Fluor®488 conjugated to Annexin V to bind to phosphatidylserine in the presence of Ca2+, and the ability of propidium iodide (PI) to enter cells with damaged cell membranes and bind to DNA. Measurements were performed immediately using a CytoFLEX LX flow cytometer (Beckman Coulter, Miami, FL, USA). List mode data were analyzed using Kaluza Analysis 2.1 software (Beckman Coulter, Miami, FL, USA).
4.8. Western Blot Analysis
Whole-cell lysates (Cell Lysis Buffer; Cell Signaling Technology, Danvers, MA, USA) were prepared 24 and 72 h following the treatment of A549 with 10 μM and 20 μM of pancracine and 4 h following the treatment of MOLT-4 cells with 5 μM and 10 μM of pancracine. Cells treated with 0.1% DMSO were used as a negative control. Cells treated with either 5 μM of cisplatin or 0.25 µM of doxorubicin were used as a positive control. Quantification of the protein content was performed using the BCA assay (Sigma-Aldrich, St. Louis, MO, USA). The lysates (20 µg of purified protein) were loaded into lanes of polyacrylamide gel. After electrophoresis separation, the proteins were transferred to a PVDF membrane (Bio-Rad, Hercules, CA, USA). Any nonspecific bindings of the membranes were blocked for 1 h in a Tris-buffered saline (TBS) containing 0.05% Tween 20 and 5%, w/v, nonfat dry milk. The membranes were washed in TBS. Incubation with a primary antibody against specific antigens and at the appropriate dilutions (1:1000 Chk1, 1:1000 Chk1_serine 345—Cell Signalling, Danvers, MA, USA; 1:1000 Rb, 1:1000 Rb_serine 807/811—Cell Signalling, Danvers, MA, USA; 1:10,000 β-actin—Sigma-Aldrich, St. Louis, MO, USA; 1:1000 ERK, 1:1000 ERK ½_threonine 202 and tyrosine 204—Cell Signalling, Danvers, MA, USA; 1:1000 p53 and 1:1000 p53_serine 392—Exbio, Prague, Czech Republic; 1:1000 p53_serine 15—Cell Signalling, Danvers, MA, USA; 1:1000 p21—Cell Signalling, Danvers, MA, USA; 1:1000 p27—Cell Signalling, Danvers, MA, USA; 1:1000 Akt and 1:1000 Akt_threonine 308—Cell Signalling, Danvers, MA, USA; 1:1000 p38, 1:1000 p38_ threonine 180, and tyrosine 182—Cell Signalling, Danvers, MA, USA; and 1:1000 Bax—Cell Signalling, Danvers, MA, USA) was performed at 4 °C overnight. The following day, the membranes were washed 5 times with TBS, each time for 5 min, and once with TBS for 10 min and then incubated with an appropriate secondary antibody (DakoCytomation, Glostrup, Denmark, or Cell Signalling, Danvers, MA, USA) in a dilution of 1:1000 for 1 h at room temperature. Band detection was performed using a chemiluminescence detection kit (Roche, Basel, Switzerland). To ensure equal protein loading, each membrane was reprobed, and β-actin was detected. The densities of the proteins of interest were analyzed using the GeneTools image analysis system (Syngene, Cambridge, UK).
4.9. Statistical Analysis
The descriptive statistics of the results were calculated and the charts made using either Microsoft Office 365 Excel (Microsoft, Redmond, WA, USA) or GraphPad Prism 7 biostatistics (GraphPad Software, La Jolla, CA, USA) software. In this study, all the values were expressed as arithmetic means with SD of triplicates unless otherwise noted. For quantitative data, normality testing was performed to assess whether parametric or nonparametric tests should be used. For experiments with parametric variables, one-way analysis of variance (ANOVA) followed by post-hoc Dunnett’s test was used to compare the mean values among different groups, with a p-value less than 0.05 considered as statistically significant.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}