
Effects of Baccharin Isolated from Brazilian Green Propolis on Adipocyte Differentiation and Hyperglycemia in ob/ob Diabetic Mice

 ,
, 
Abstract
:1. Introduction
2. Results
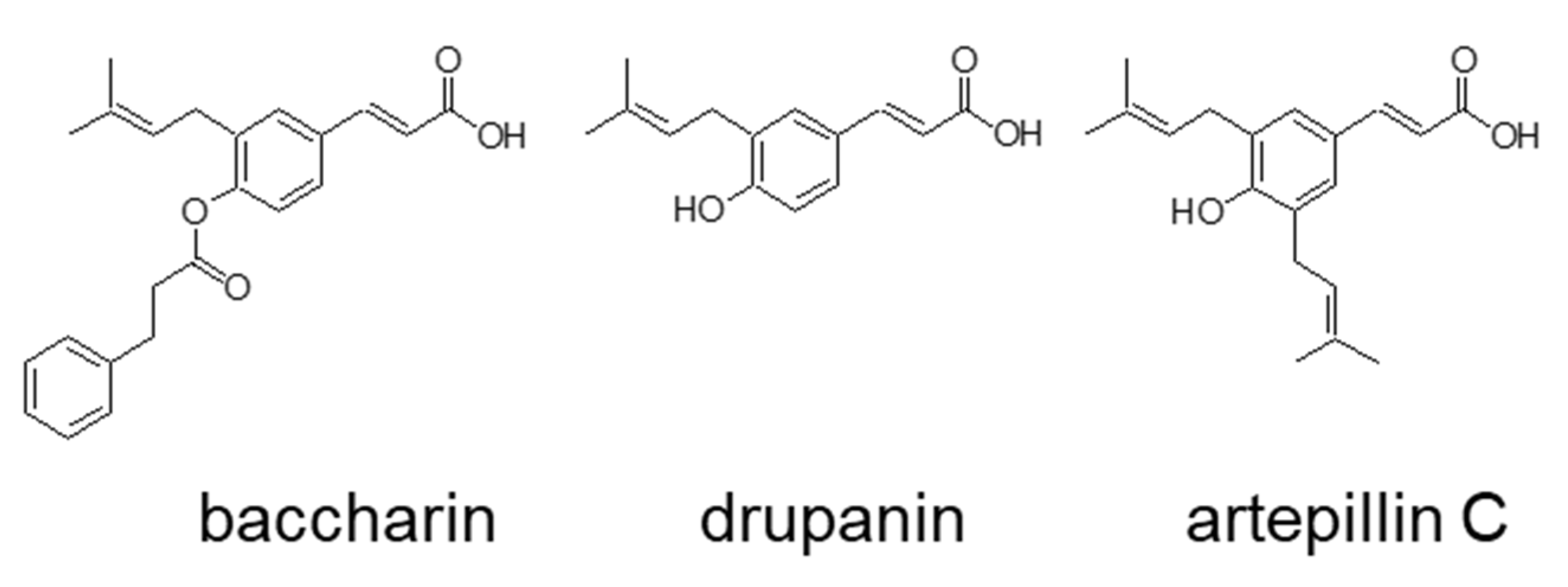
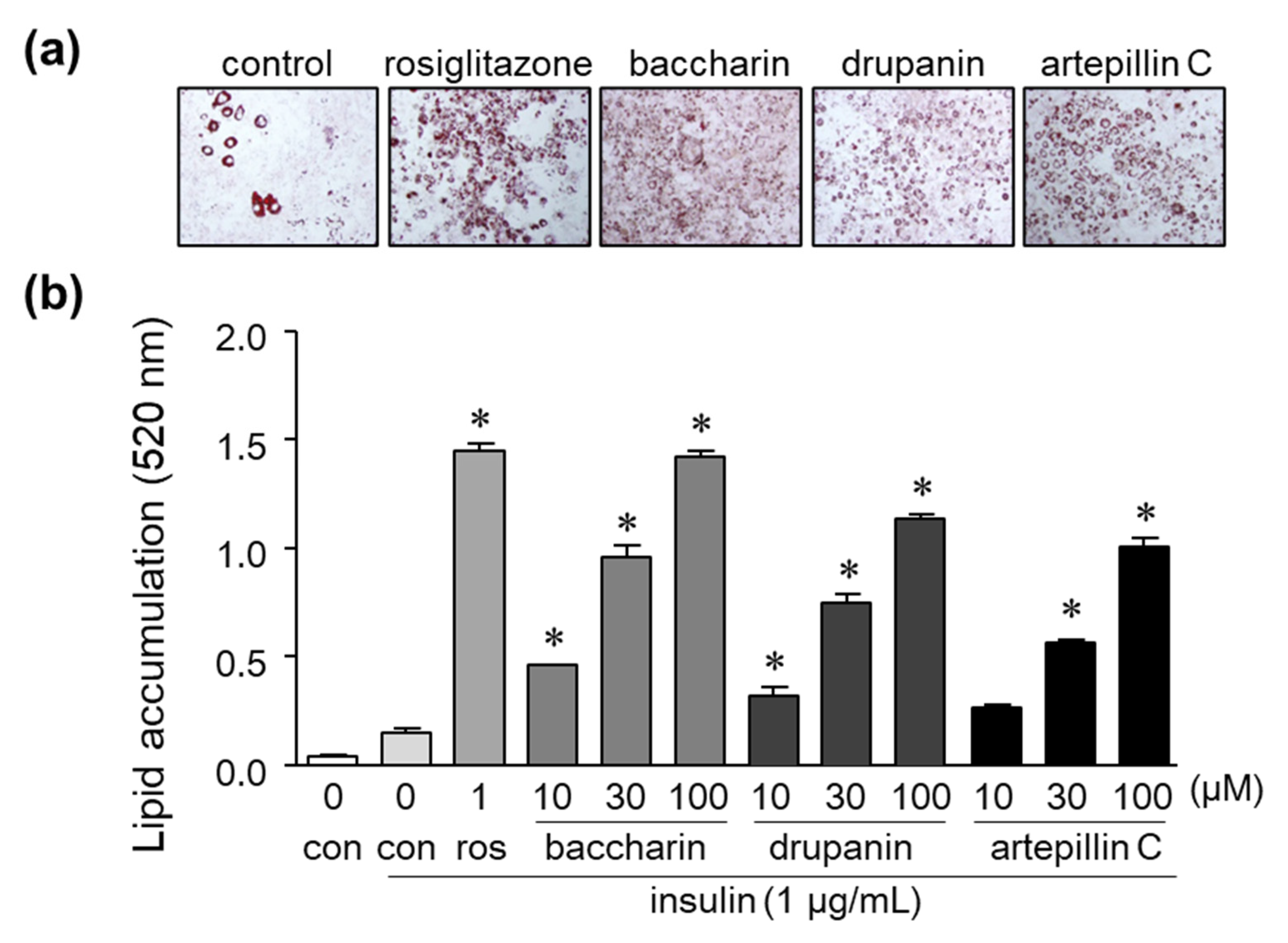
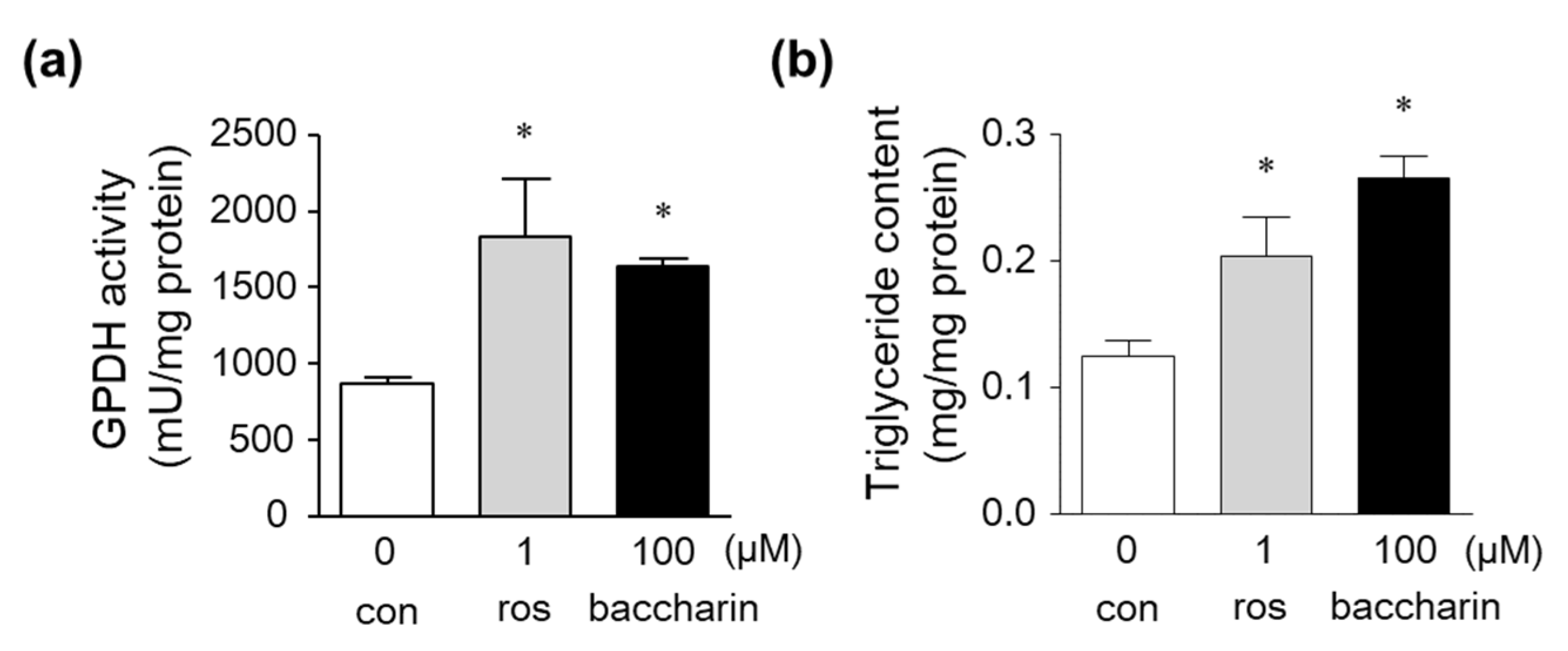
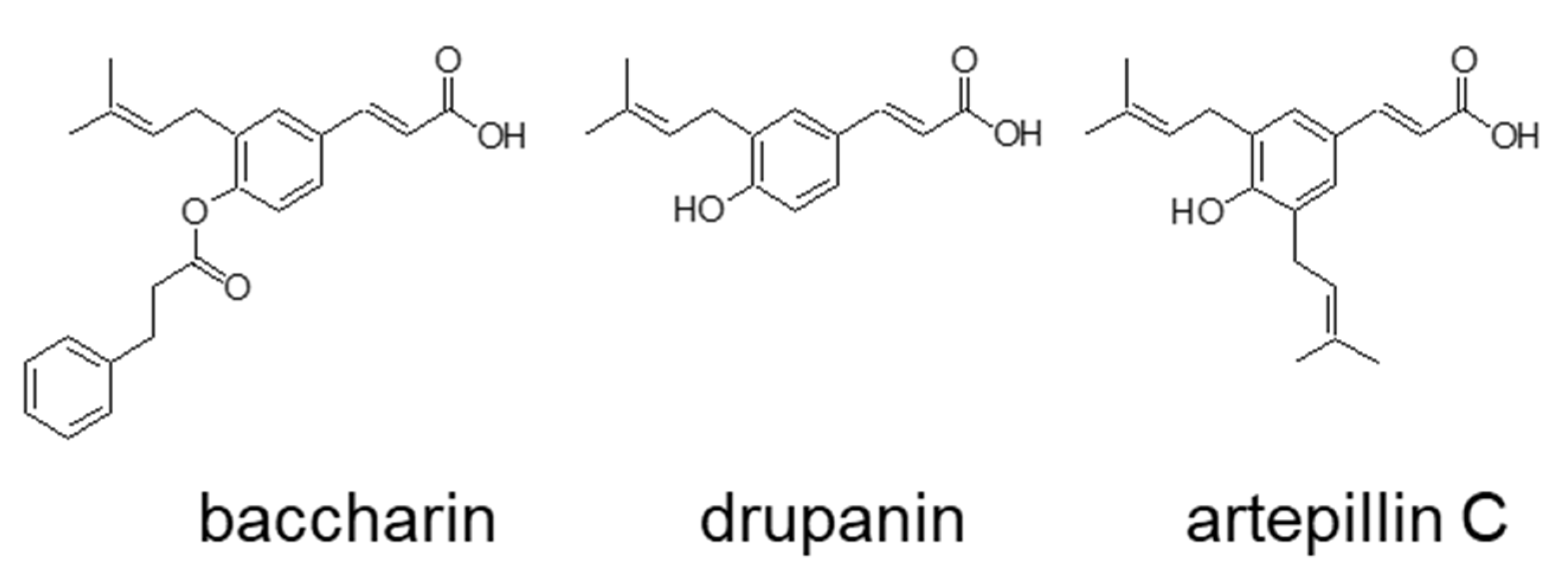
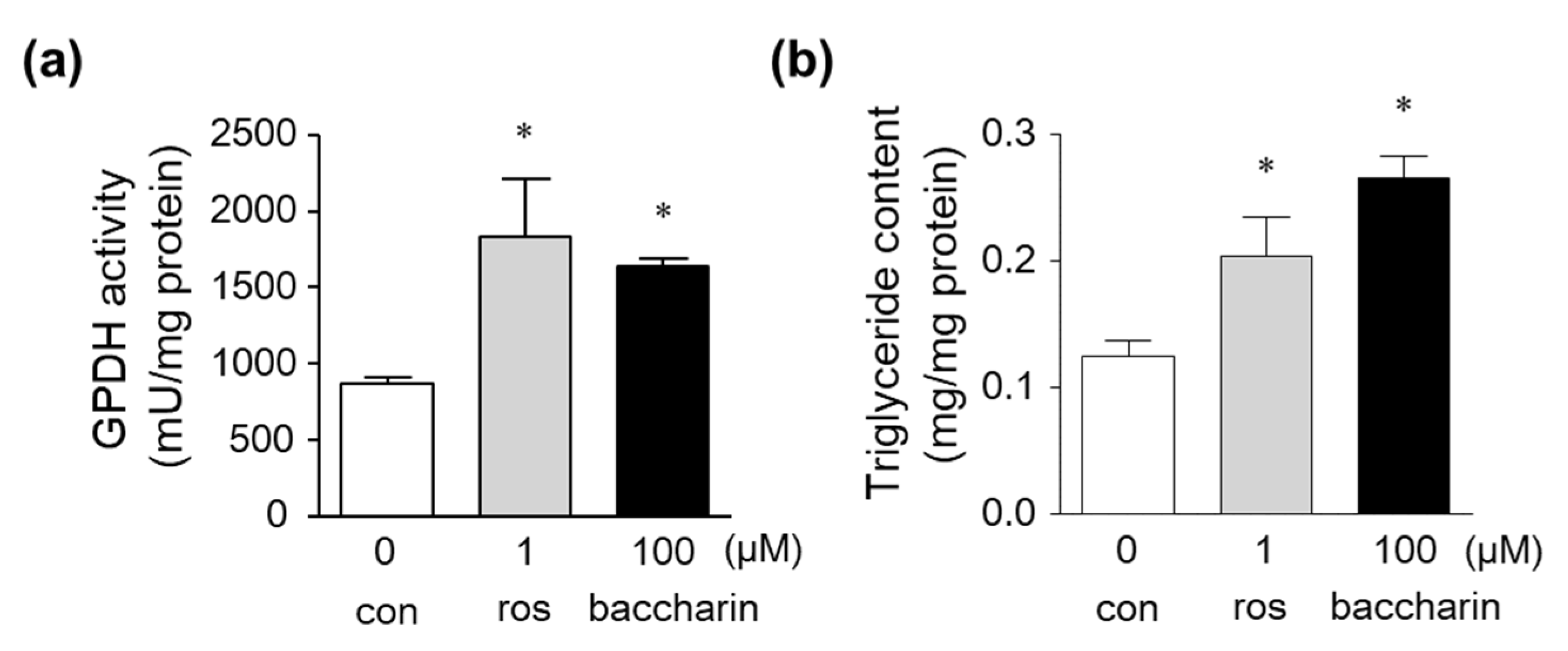
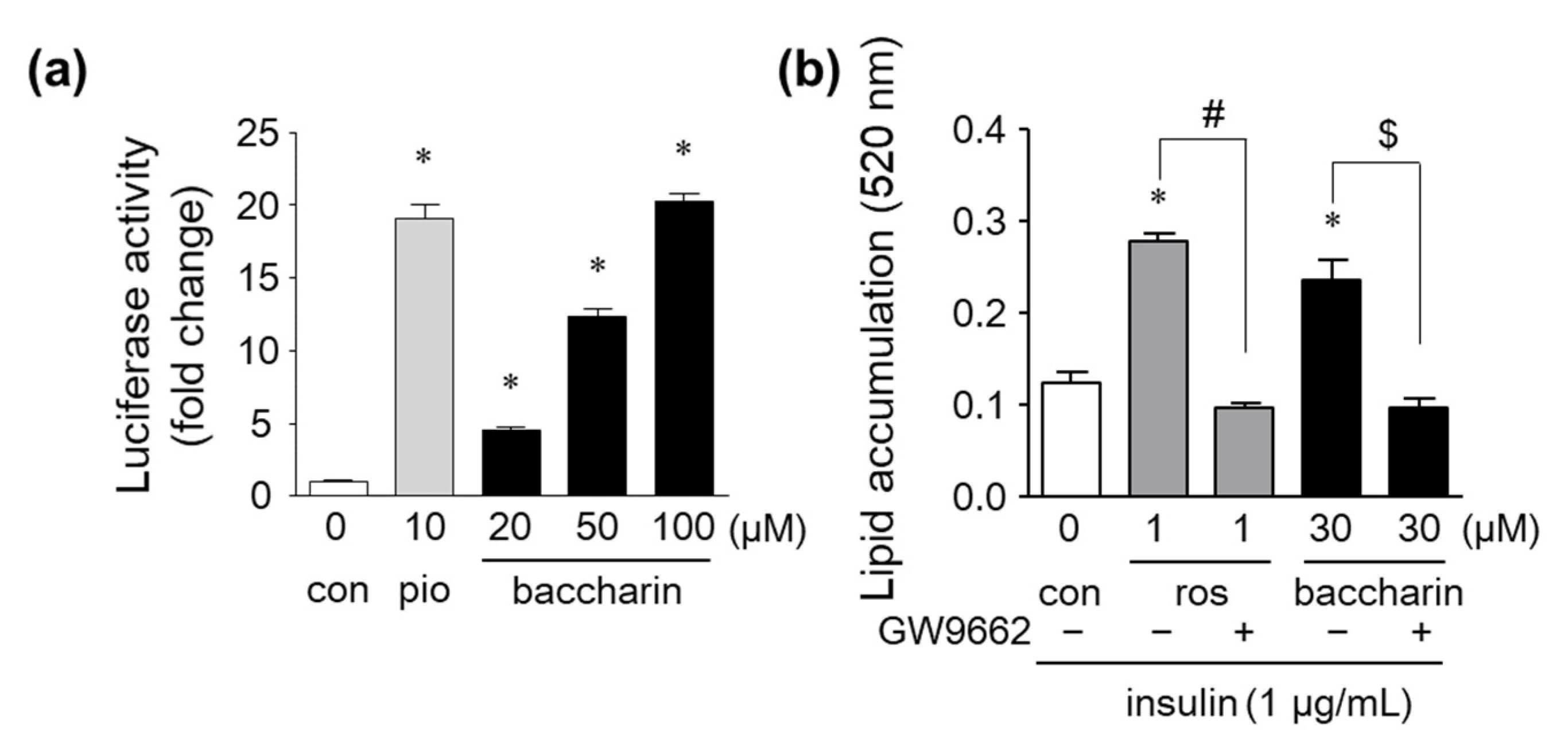
2.1. Effects of Baccharin on 3T3-L1 Adipocyte Differentiation
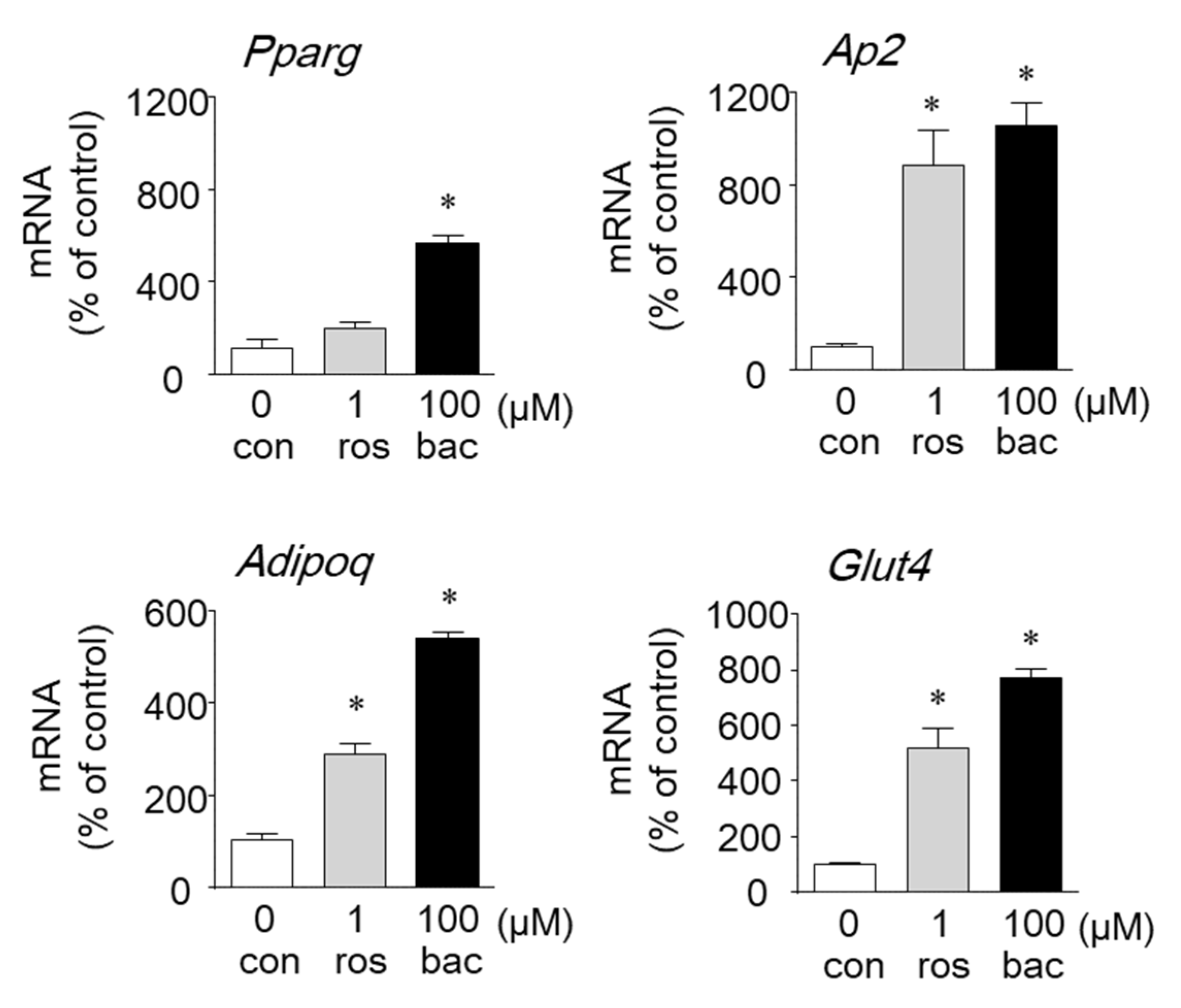
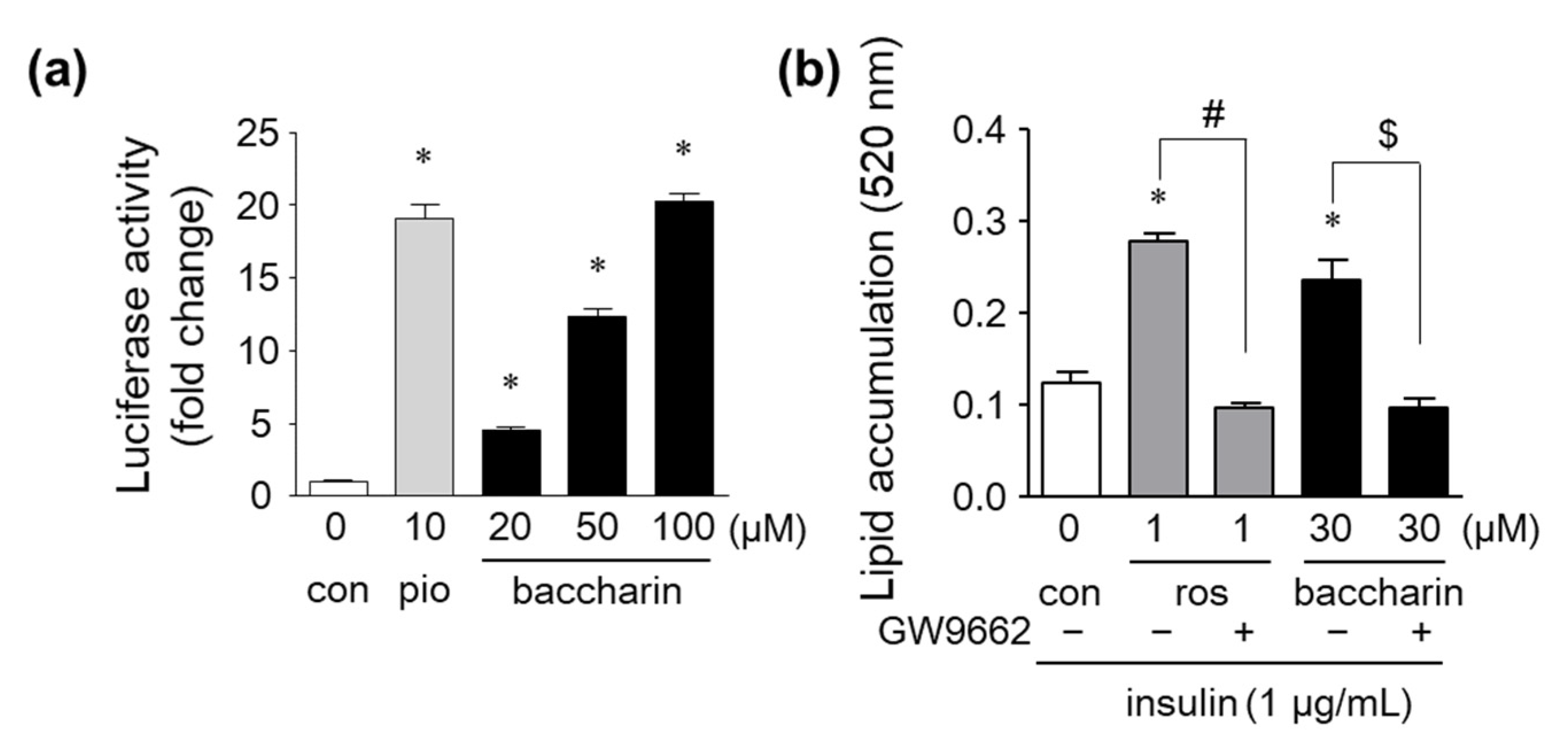
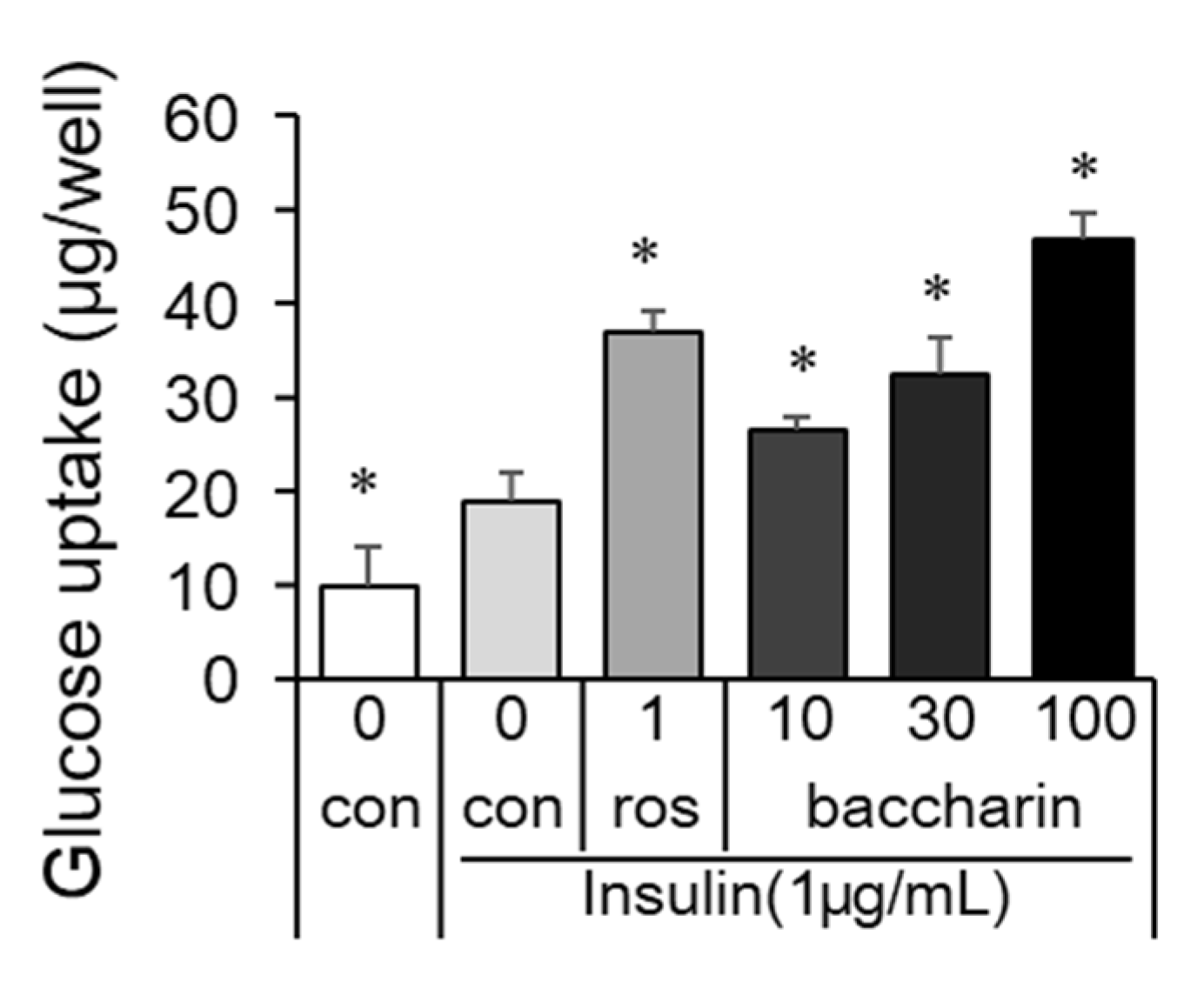
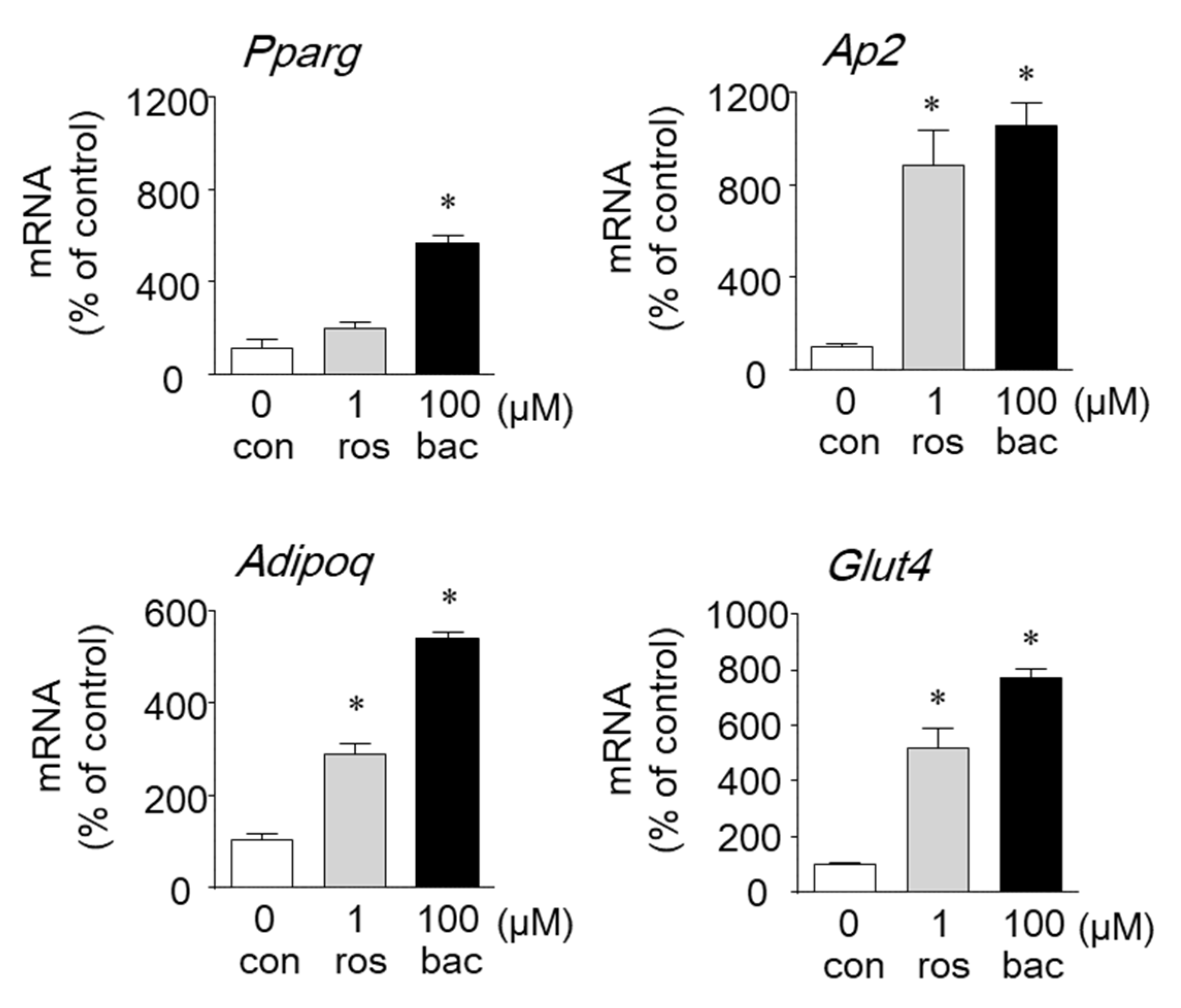
2.2. Effects of Baccharin on the mRNA Expression of Adipocyte-Related Genes and PPARγ-Dependent Transcriptional Activity
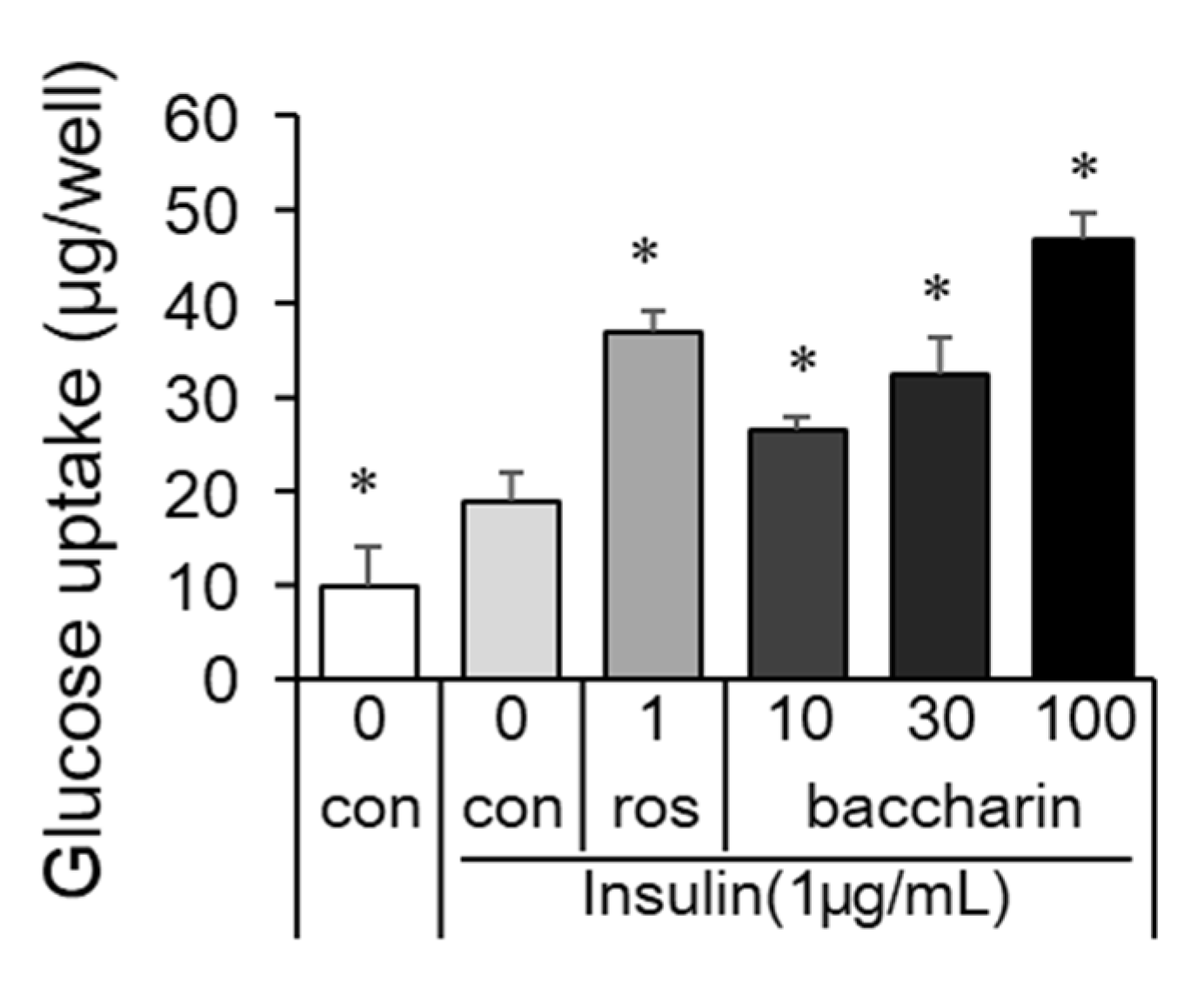
2.3. Effects of Baccharin on Glucose Metabolism in Obese Diabetic ob/ob Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation of Baccharin from Brazilian Green Propolis
4.3. Cell Culture and Adipocyte Differentiation
4.4. Oil Red O Staining
4.5. Triglyceride Levels and GPDH Activity
4.6. Glucose Uptake Assay
4.7. RNA Preparation and qRT-PCR
4.8. Reporter Gene Assay for PPARγ
4.9. Treatment of Obese Type 2 Diabetic ob/ob Mice
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- De Oliveira, P.; Lima, I.D.S.; Munari, C.; Bastos, J.; Filho, A.D.S.; Tavares, D. Comparative Evaluation of Antiproliferative Effects of Brazilian Green Propolis, Its Main Source Baccharis dracunculifolia, and Their Major Constituents Artepillin C and Baccharin. Planta Med. 2014, 80, 490–492. [Google Scholar] [CrossRef] [Green Version]
- Berretta, A.A.; Arruda, C.; Miguel, F.G.; Baptista, N.; Nascimento, A.P.; Oliveira, F.M.; Hori, J.I.; Barud, H.D.S.; Damaso, C.R.B.; Ramos, C.; et al. Functional Properties of Brazilian Propolis: From Chemical Composition Until the Market. Superfood Funct. Food Overv. Process. Util. 2017, 55–98. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, R.; Yanagisawa, M.; Takahashi, N.; Kawada, T.; Kumazawa, S.; Yamaotsu, N.; Nakagome, I.; Hirono, S.; Tsuda, T. Brazilian propolis-derived components inhibit TNF-α-mediated downregulation of adiponectin expression via different mechanisms in 3T3-L1 adipocytes. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2011, 1810, 695–703. [Google Scholar] [CrossRef]
- Choi, S.S.; Cha, B.Y.; Iida, K.; Lee, Y.S.; Yonezawa, T.; Teruya, T.; Nagai, T.; Woo, J.T. Artepillin C, as a PPARγ ligand, en-hances adipocyte differentiation and glucose uptake in 3T3-L1 cells. Biochem. Pharmacol. 2011, 81, 925–933. [Google Scholar] [CrossRef]
- Kitamura, H.; Naoe, Y.; Kimura, S.; Miyamoto, T.; Okamoto, S.; Toda, C.; Shimamoto, S.; Iwanaga, T.; Miyoshi, I. Beneficial effects of Brazilian propolis on type 2 diabetes in ob/ob mice: Possible involvement of immune cells in mesenteric adipose tis-sue. Adipocyte 2013, 2, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.; Almeida, M.O.; Lemos, M.; Arruda, C.; Casoti, R.; Somensi, L.B.; Boeing, T.; Mariott, M.; da Silva, R.D.C.M.V.D.A.F.; Stein, B.D.P.; et al. Artepillin C, drupanin, aromadendrin-4′-O-methyl-ether and kaempferide from Brazilian green propolis promote gastroprotective action by diversified mode of action. J. Ethnopharmacol. 2018, 226, 82–89. [Google Scholar] [CrossRef]
- Viviane, C.T.; Helia, H.S.; Glaucia, M.P.; Yong, K.P. Recent progress of propolis for its biological and chemical composi-tions and its botanical origin. Evid. Based Complement Alternat. Med. 2013, 697390, 1–13. [Google Scholar]
- Búfalo, M.C.; Candeias, J.M.; Sousa, J.P.; Bastos, J.K.; Sforcin, J.M. In vitro cytotoxic activity of Baccharis dracunculifolia and propolis against HEp-2 cells. Nat. Prod. Res. 2010, 24, 1710–1718. [Google Scholar] [CrossRef]
- Salatino, A.; Teixeira, É.W.; Negri, G.; Message, D. Origin and Chemical Variation of Brazilian Propolis. Evid. -Based Complement. Altern. Med. 2005, 2, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Seidell, J. Obesity, insulin resistance and diabetes—A worldwide epidemic. Br. J. Nutr. 2000, 83, S5–S8. [Google Scholar] [CrossRef] [Green Version]
- Lam, D.W.; Le Roith, D. The worldwide diabetes epidemic. Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 93–96. [Google Scholar] [CrossRef]
- Reaven, G.M. Role of insulin resistance in human disease. Diabetes 1988, 37, 1595–1607. [Google Scholar] [CrossRef]
- Kahn, B.B. Type 2 Diabetes: When Insulin Secretion Fails to Compensate for Insulin Resistance. Cell 1998, 92, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Daniele, G.; Mendoza, R.G.; Winnier, D.; Fiorentino, T.V.; Pengou, Z.; Cornell, J.; Andreozzi, F.; Jenkinson, C.; Cersosimo, E.; Federici, M.; et al. The inflammatory status score including IL-6, TNF-α, osteopontin, fractalkine, MCP-1 and adiponectin underlies whole-body insulin resistance and hyperglycemia in type 2 diabetes mellitus. Acta Diabetol. 2014, 51, 123–131. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K.; Tobe, K. Adiponectin and adiponectin receptors in insulin re-sistance, diabetes, and the metabolic syndrome. J. Clin. Investig. 2006, 116, 1784–1792. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.G.; Javorschi, S.; Hevener, A.L.; Kruszynska, Y.T.; Norman, R.A.; Sinha, M.; Olefsky, J.M. The effect of thiazolidinedi-ones on plasma adiponectin levels in normal, obese, and type diabetic subjects. Diabetes 2002, 51, 2968–2974. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, V. Effect of thiazolidinediones on body weight in patients with diabetes mellitus. Am. J. Med. 2003, 115, 42–48. [Google Scholar] [CrossRef]
- Lago, R.M.; Singh, P.P.; Nesto, R.W. Congestive heart failure and cardiovascular death in patients with prediabetes and type 2 diabetes given thiazolidinediones: A meta-analysis of randomized clinic trials. Lancet 2007, 370, 1129–1136. [Google Scholar] [CrossRef]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.; et al. Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARγ): A review. Biochem. Pharmacol. 2014, 92, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, K.; Murakami, T.; Tanabe, H.; Inoue, M. Identification of a naturally occurring retinoid X receptor agonist from Brazilian green propolis. Biochim. Biophys. Acta 2014, 1840, 3034–3041. [Google Scholar] [CrossRef]
- Wise, L.; Green, H. Participation of one isozyme of cytosolic glycerophosphate dehydrogenase in the adipose conversion of 3T3 cells. J. Biol. Chem. 1979, 254, 273–275. [Google Scholar] [CrossRef]
- Bankova, V.; Marcucci, M.C.; Simova, S.; Nikolova, N.; Kujumgiev, A.; Popov, S. Anti-bacterial diterpenic acids from Bra-zilian propolis. Z. Nat. C 1996, 51, 277–280. [Google Scholar]
- Sforcin, J.M.; Bankova, V. Propolis: Is there a potential for the development of new drugs? J. Ethnopharmacol. 2011, 133, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, P.K.; Doble, M. Interaction of Cinnamic Acid Derivatives with Commercial Hypoglycemic Drugs on 2-Deoxyglucose Uptake in 3T3-L1 Adipocytes. J. Agric. Food Chem. 2011, 59, 9835–9844. [Google Scholar] [CrossRef]
- Koya-Miyata, S.; Koya-Miyata, S.; Arai, N.; Mizote, A.; Taniguchi, Y.; Ushio, S.; Iwaki, K.; Fukuda, S. Propolis prevents di-et-induced hyperlipidemia and mitigates weight gain in diet-induced obesity in mice. Biol. Pharm. Bull. 2009, 12, 2022–2028. [Google Scholar] [CrossRef] [Green Version]
- Sakai, T.; Ohhata, M.; Fujii, M.; Oda, S.; Kusaka, Y.; Matsumoto, M.; Nakamoto, A.; Taki, T.; Nakamoto, M.; Shuto, E. Brazilian Green Propolis Promotes Weight Loss and Reduces Fat Accumulation in C57BL/6 Mice Fed A High-Fat Diet. Biol. Pharm. Bull. 2017, 40, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakuta, H.; Kurosaki, E.; Niimi, T.; Gato, K.; Kawasaki, Y.; Suwa, A.; Honbou, K.; Yamaguchi, T.; Okumura, H.; Sanagi, M.; et al. Distinct Properties of Telmisartan on Agonistic Activities for Peroxisome Proliferator-Activated Receptor γ among Clinically Used Angiotensin II Receptor Blockers: Drug-Target Interaction Analyses. J. Pharmacol. Exp. Ther. 2014, 349, 10–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schupp, M.; Clemenz, M.; Gineste, R.; Witt, H.; Janke, J.; Helleboid, S.; Hennuyer, N.; Ruiz, P.; Unger, T.; Staels, B.; et al. Molecular characterization of new selective peroxisome proliferator-activated receptor gamma modula-tors with angiotensin receptor blocking activity. Diabetes 2005, 54, 3442–3452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aga, H.; Shibuya, T.; Sugimoto, T.; Kurimoto, M.; Nakajima, S. Isolation and Identification of Antimicrobial Compounds in Brazilian Propolis. Biosci. Biotechnol. Biochem. 1994, 58, 945–946. [Google Scholar] [CrossRef]
- Hattori, H.; Okuda, K.; Murase, T.; Shigetsura, Y.; Narise, K.; Semenza, G.L.; Nagasawa, H. Isolation, identification, and biological evaluation of HIF-1-modulating compounds from Brazilian green propolis. Bioorg. Med. Chem. 2011, 19, 5392–5401. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Control | ob/ob Control | ob/ob Baccharin 10 mg/kg | ob/ob Baccharin 50 mg/kg | |
|---|---|---|---|---|
| Body weight (g) | 22.1 ± 0.4 * | 39.7 ± 1.2 | 39.4 ± 1.8 | 37.7 ± 1.1 |
| Liver weight (g/100 g body weight) | 4.39 ± 0.09 * | 6.30 ± 0.36 | 6.21 ± 0.34 | 5.64 ± 0.23 |
| White adipose tissue weight (g/100 g body weight) | 2.52 ± 0.17 * | 18.95 ± 0.45 | 17.65 ± 0.65 | 17.13 ± 0.47 |
| Fasting blood glucose (mg/dL) | 92.8 ± 3.4 * | 308 ± 40 | 340 ± 33 | 180 ± 35 * |
| Non-fasting blood glucose (mg/dL) | 163.4 ± 7.3 * | 362.0 ± 29.4 | 364.4 ± 32.8 | 239.3 ± 43.3 * |
| Plasma insulin (ng/mL) | N.D. | 4.04 ± 0.64 | 4.94 ± 0.83 | 1.98 ± 0.72 |
| Plasma adiponectin (µg/mL) | 13.6 ± 0.9 | 20.3 ± 3.6 | 20.8 ± 3.1 | 17.9 ± 2.6 |
| HOMA-IR | N.D. | 75.4 ± 10.1 | 107.0 ± 20.5 | 20.0 ± 6.2 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, A.; de Almeida, M.O.; Deguchi, Y.; Kozuka, R.; Arruda, C.; Berreta, A.A.; Bastos, J.K.; Woo, J.-T.; Yonezawa, T. Effects of Baccharin Isolated from Brazilian Green Propolis on Adipocyte Differentiation and Hyperglycemia in ob/ob Diabetic Mice. Int. J. Mol. Sci. 2021, 22, 6954. https://doi.org/10.3390/ijms22136954
Watanabe A, de Almeida MO, Deguchi Y, Kozuka R, Arruda C, Berreta AA, Bastos JK, Woo J-T, Yonezawa T. Effects of Baccharin Isolated from Brazilian Green Propolis on Adipocyte Differentiation and Hyperglycemia in ob/ob Diabetic Mice. International Journal of Molecular Sciences. 2021; 22(13):6954. https://doi.org/10.3390/ijms22136954
Chicago/Turabian StyleWatanabe, Akio, Marília Oliveira de Almeida, Yusuke Deguchi, Ryuzo Kozuka, Caroline Arruda, Andresa Aparecida Berreta, Jairo Kenupp Bastos, Je-Tae Woo, and Takayuki Yonezawa. 2021. "Effects of Baccharin Isolated from Brazilian Green Propolis on Adipocyte Differentiation and Hyperglycemia in ob/ob Diabetic Mice" International Journal of Molecular Sciences 22, no. 13: 6954. https://doi.org/10.3390/ijms22136954
APA StyleWatanabe, A., de Almeida, M. O., Deguchi, Y., Kozuka, R., Arruda, C., Berreta, A. A., Bastos, J. K., Woo, J.-T., & Yonezawa, T. (2021). Effects of Baccharin Isolated from Brazilian Green Propolis on Adipocyte Differentiation and Hyperglycemia in ob/ob Diabetic Mice. International Journal of Molecular Sciences, 22(13), 6954. https://doi.org/10.3390/ijms22136954