
Brain Mechanisms of COVID-19-Sleep Disorders

 ,
, 
 and
and
Abstract
1. The ‘Coronasomnia’ Phenomenon
2. Sleep Loss-Associated Neuroinflammation and the Blood–Brain Barrier Disruption

{kind=link}
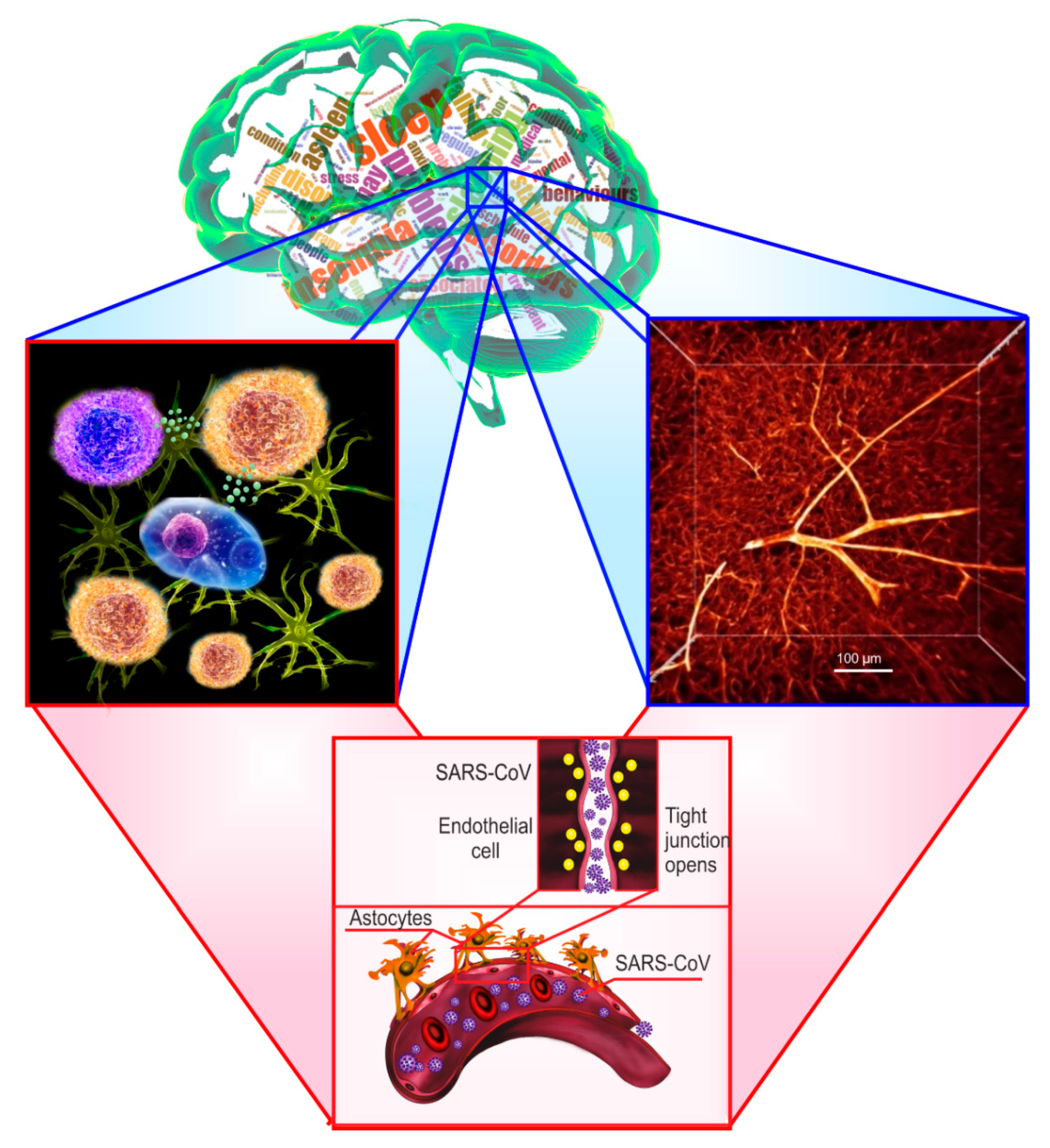{kind=link}
{kind=link}
{kind=link}
| References | Sleep Loss Models | The Effects of Sleep Loss on the BBB Permeability |
|---|---|---|
| Medina-Flores F. et al. (2020) [38] | Wistar rats were sleep-restricted 20 h daily with 4 h sleep recovery for 10 days. | Sleep loss disrupts pericyte-brain endothelial cell interactions. |
| Hurtado-Alvarado G. et al. (2016) [39]. Review | Acute and chronic sleep deprivation, sleep restriction and sleep fragmentation. | Sleep loss induces a low-grade systemic inflammation characterized by the release of several molecules, such as cytokines, chemokines, and acute-phase proteins; all of them may promote changes in cellular components of BBB, particularly on brain endothelial cells. |
| Hurtado-Alvarado G. et al. (2018) [35] | Mice were sleep-restricted during 10 days using the flowerpot technique for 20 h per day with 4 h of daily sleep opportunity. | Cytokines may play a key role in modulating BBB function during sleep restriction via the overexpression of Iba-1, MMP-9 and A2A adenosine receptors. |
| He J. et al. (2014) [50] | Chronic sleep restriction of mice for 6 days in a rotatory bar for 12 h per day. Sleep restriction of this method induced REM sleep loss in the first3 days with partial REM sleep recovery afterward. | Chronic sleep restriction diminished endothelial and inducible nitric oxide synthase, endothelin1, and glucose transporter expression in cerebral microvessels of BBB and decreased 2-deoxy-glucose uptake by the brain. The expression of several tight junction proteins was decreased, whereas the level of cyclooxygenase-2 increased. This coincided with an increase of paracellular permeability of BBB to the small tracers sodium fluorescein and biotin. Chronic sleep restriction for 6 d was sufficient to impair BBB structure and function, although the increase of paracellular permeability returned to baseline after 24 h of recovery sleep. |
| Hurtado-Alvarado G. et al. (2017) [56] | Male Wistar rats were sleep restricted using the modified multiple platform method for 10 days, with a daily schedule of 20-h sleep deprivation plus 4-h sleep recovery at their home-cages. | Chronic sleep restriction disrupts interendothelial tight junctions in the hippocampus and increases BBB permeability to fluorescein-sodium, and decreases interendothelial junction complexity by increasing the frequency of less mature end-to-end and simply overlap junctions, even after sleep recovery, as compared to intact controls. Chronic sleep loss also induces the formation of clefts between narrow zones of adjacent endothelial cell membranes in the hippocampus. |
| Gómez-González B. (2013) [55] | REM sleep restriction was induced by the multiple platform technique; male rats were REM sleep-restricted 20 h daily (with 4 h sleep opportunity) for 10 days; control groups included large platform and intact rats. | REM sleep restriction increased BBB permeability to Evans blue in the whole brain. Brief periods of sleep recovery rapidly and effectively restored the severe alteration of BBB function by reducing BBB transfer of Evans blue. The mechanism of BBB breakdown involved increased caveolae formation at brain endothelial cells. REM sleep regulates the physical barrier properties of BBB. |
| Hurtado-Alvarado G. et al. (2016) [54] | The chronic sleep restriction of male Wistar rats during 10 days. | Sleep restriction increased BBB permeability to FITC-dextrans and Evans blue, and the effect was reverted by the administration of selective A2A adenosine receptor antagonist (SCH58261) in almost all brain regions excluding the cerebellum. Sleep restriction increased the expression of A2A adenosine receptor only in the hippocampus and basal nuclei without changing the expression of adenosine-synthesizing enzyme (CD73) in all brain regions. Sleep restriction reduced the expression of tight junction proteins (claudin-5, occludin, ZO-1) in all brain regions, except in the cerebellum; and SCH58261 restored the levels of tight junction proteins in the cortex, hippocampus and basal nuclei. Sleep restriction-induced neuroinflammatory markers (GFAP and Iba-1) overexpression that was attenuated with the administration of SCH58261. |
| Daulatzai M.A. (2016) [42]. Review | Obstructive sleep apnea | Obstructive sleep apnea is a risk factor triggering neuroinflammation and oxidative-nitrosative stress that in turn decrease nitric oxide and enhance endothelin, amyloid-β deposition, cerebral amyloid angiopathy, and BBB disruption. |
| Lim D.C. and Pack A.I. (2014) [41]. Review | Obstructive sleep apnea | Cyclical intermittent hypoxia is a stressor that disrupts BBB via molecular responses already known to occur in either obstructive sleep apnea patients or animal models of intermittent hypoxia. |
| Voirin A.C. et al. (2020) [43] | Obstructive sleep apnea. The two groups of volunteers were selected, a group of patients suffering newly diagnosed severe obstructive sleep apnea (AHI > 30/h) and a group showing no sleep apnea (AHI <5/h). The human in vitro BBB model of endothelial cells (HBEC-5i) with sera of patients with and without obstructive sleep apnea was studied. | After incubation with sera from patients with obstructive sleep apnea, there was a loss of integrity in the human in vitro BBB model; this was reflected by an increase in permeability (43%; p < 0.001) and correlated with a 50% and 40% decrease in tight junction protein expression of ZO-1 and claudin-5, respectively. There was an upregulation in Pgp protein expression (52%) and functionality and a downregulation in BCRP expression (52%). These results demonstrate that severe BBB disorder after exposure to sera from patients with obstructive sleep apnea was reflected by BBB opening. |
| Benedict C. et al. (2014) [45] | Healthy young men were divided on 2 groups including either 8-h of nocturnal sleep [22:30–06:30] and total sleep deprivation. | Acute sleep deprivation increases serum levels of neuron-specific enolase (NSE) and S100 calcium-binding protein B (S-100B) in healthy young men. |
| Opp M.R. et al. (2015) [46] | The sleep fragmentation device used in this study. The device consists of a cylindrical Plexiglass® chamber divided into two separate compartments. The floor of the chamber is a disc that is programmed to rotate at specific intervals. In this study, disc rotations were confined to the 12 h light period, and consisted of an 8 s rotation once every 30 s, on average. The direction of the disc rotation was randomized, and the precise placement of the 8 s rotation within the 30 s period was varied to prevent behavioral adaptation by a mouse to disc movements. Sleeping mice wake up when the disc rotates, whereas awake mice continue their normal behavior. During the 12 h dark period, there was no disc rotation, and mice were allowed spontaneous behavior. Mice were housed in the sleep disruption devices, one mouse in each compartment, for 3 days of habituation prior to beginning the sleep fragmentation protocol. | The sleep fragmentation disrupted the BBB and increased tumor necrosis factor-α transport in aged mice but not in young mice. |
| Pan W. and Kastin A.J. (2017) [36]. Review | Acute and chronic sleep deprivation, sleep restriction, and sleep fragmentation. | Authors summarize research on the sleep-BBB interactions in five sections: (1) the structural basis enabling the BBB to serve as a huge regulatory interface; (2) BBB transport and permeation of substances participating in sleep-wake regulation; (3) the circadian rhythm of BBB function; (4) the effect of experimental sleep disruption maneuvers on BBB activities, including regional heterogeneity, possible threshold effect, and reversibility; and (5) implications of sleep disruption-induced BBB dysfunction in neurodegeneration and CNS autoimmune diseases. |
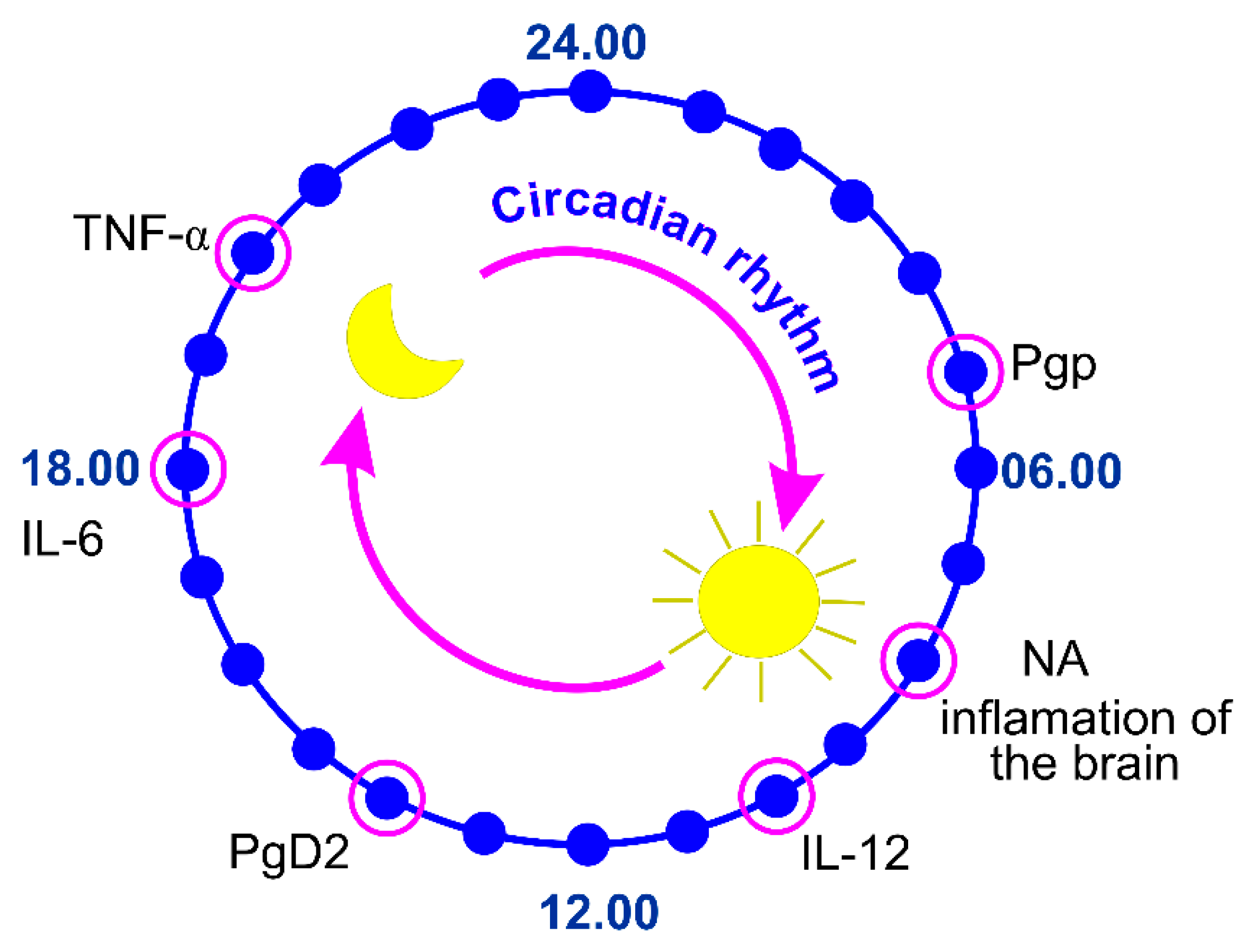
3. Circadian Regulation of the Blood–Brain Barrier Permeability to Cytokines
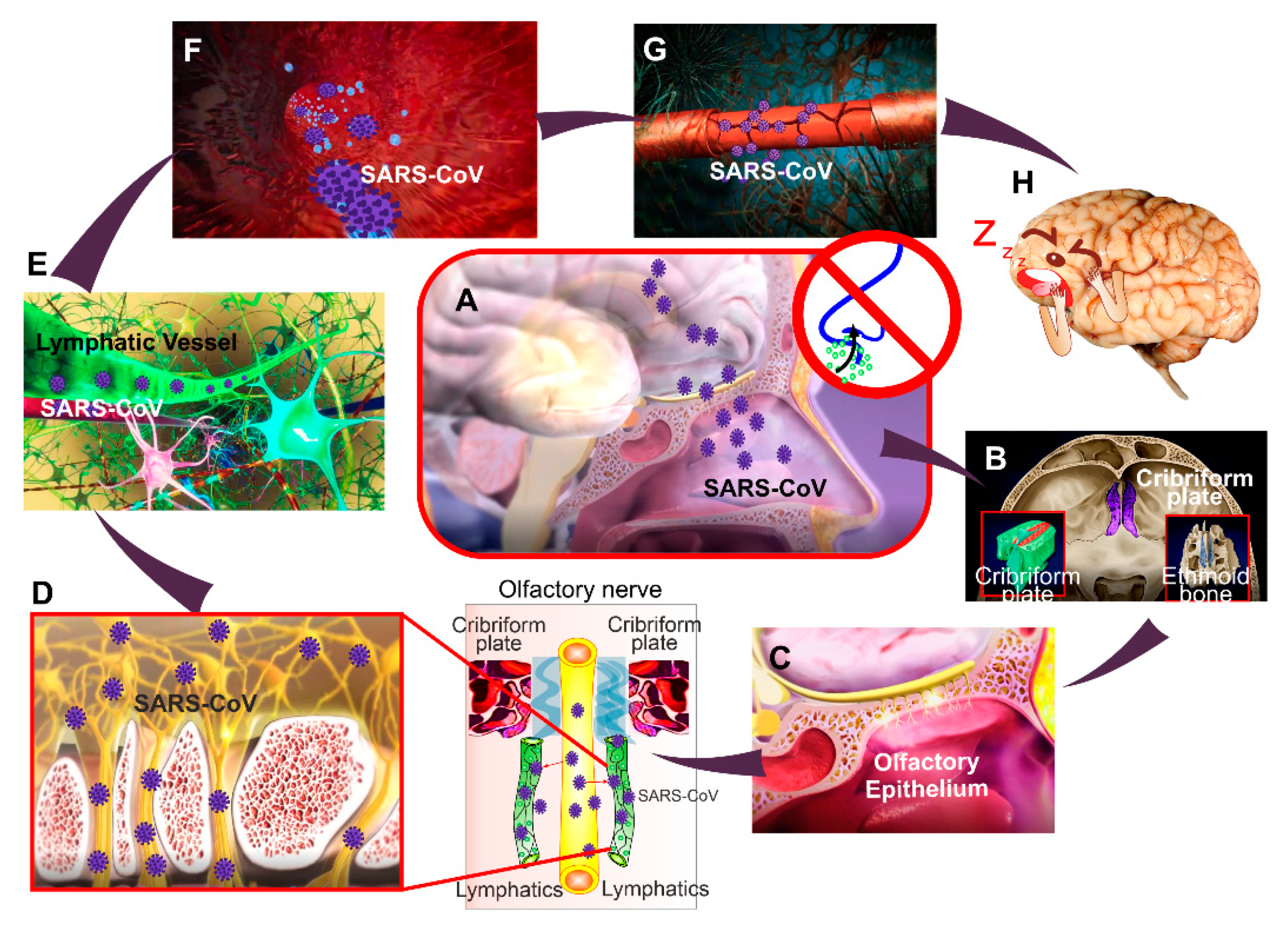
4. The COVID-19 Attack on the Blood–Brain Barrier Integrity
5. The Perspective Strategies for Prevention of COVID-19-Related BBB Disorders
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hui, D.S.; Azhar, E.I.; Madani, T.A.; Ntoumi, F.; Kock, R.; Dar, O.; Ippolito, G.; Mchugh, T.D.; Memish, Z.A.; Drosten, C.; et al. The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health—The latest 2019 novel coronavirus outbreak in Wuhan, China. Int. J. Infec. Dis. 2020, 91, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Gunther, S.; Preiser, W.; Van der Werf, S.; Brodt, H.R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef]
- Ksiazek, T.G.; Erdman, D.; Goldsmith, C.S.; Zaki, S.R.; Peret, T.; Emery, S.; Tong, S.; Urbani, C.; Comer, J.A.; Lim, W.; et al. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1953–1966. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.S.; Lai, S.T.; Poon, L.L.; Guan, Y.; Yam, L.Y.; Lim, W.; Nicholls, J.; Yee, W.K.; Yan, W.W.; Cheung, M.T.; et al. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet 2003, 361, 1319–1325. [Google Scholar] [CrossRef]
- Webster, R.G. Wet markets—A continuing source of severe acute respiratory syndrome and influenza? Lancet 2004, 363, 234–236. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Yuen, K. Infectious diseases emerging from Chinese wet-markets: Zoonotic origins of severe respiratory viral infections. Curr. Opin. Infect. Dis. 2006, 19, 401–407. [Google Scholar] [CrossRef]
- Sharifian-Dorche, M.; Huot, P.; Osherov, M.; Wen, D.; Saveriano, A.; Giacomini, P.; Antela, J.P.; Mowla, A. Neurological complications of coronavirus infection; a comparative review and lessons learned during the COVID-19 pandemic. J. Neurol. Sci. 2020, 417, 117085. [Google Scholar] [CrossRef]
- Varatharaj, A.; Thomas, N.; Ellul, M.A.; Davies, N.W.S.; Pollak, T.A.; Tenorio, E.L.; Sultan, M.; Easton, A.; Breen, G.; Zandi, М.; et al. Neurological and neuropsychiatric complications of COVID-19 in 153 patients: A UK-wide surveillance study. Lancet Psychiatry 2020, 7, 875–882. [Google Scholar] [CrossRef]
- Markuu, P. Sleep research in 2020: COVID-19-related sleep disorders. Lancet Neurol. 2021, 20, 15–17. [Google Scholar]
- Huang, Y.; Zhao, N. Generalized anxiety disorder, depressive symptoms and sleep quality during COVID-19 outbreak in China: A web-based cross-sectional survey. Psychiatry Res. 2020, 288, 112954. [Google Scholar] [CrossRef]
- Mazza, C.; Ricci, E.; Biondi, S.; Colasanti, M.; Ferracuti, S.; Napoli, C.; Roma, P. A nationwide survey of psychological distress among Italian people during the covid-19 pandemic: Immediate psychological responses and associated factors. Int. J. Environ. Res. Public Health 2020, 17, 3165. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, L.; Liu, S.; Ma, S.; Wang, Y.; Cai, Z.; Zhang, B. Survey of insomnia and related social psychological factors among medical staff involved in the 2019 novel Coronavirus disease outbreak. Front. Psychiatry 2020, 11, 306. [Google Scholar] [CrossRef] [PubMed]
- Cellini, N.; Canale, N.; Mioni, G.; Costa, S. Changes in sleep pattern, sense of time and digital media use during COVID-19 lockdown in Italy. J. Sleep Res. 2020, 29, e13074. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.P.; Chesney, E.; Oliver, D.; Pollak, T.A.; McGuire, P.; Fusar-Poli, P.; David, A.S. Psychiatric and neuropsychiatric presentations associated with severe coronavirus infections: A systematic review and meta-analysis with comparison to the COVID-19 pandemic. Lancet Psychiatry 2020, 7, 611–627. [Google Scholar] [CrossRef]
- Partinen, M.; Kronholm, E. Epidemiology: Principles and application in sleep medicine. In Sleep Disorders Medicine: Basic Science, Technical Considerations and Clinical Aspects, 4th ed.; Chokroverty, S., Ed.; Springer: New York, NY, USA, 2017; pp. 485–521. [Google Scholar]
- Belleville, G.; Ouellet, M.C.; Morin, C.M. Post-traumatic stress among evacuees from the 2016 fort mcmurray wildfires: Exploration of psychological and sleep symptoms three months after the evacuation. Int. J. Environ. Res. Public Health 2019, 16, 1604. [Google Scholar] [CrossRef]
- Lavie, P. Sleep disturbances in the wake of traumatic events. N. Engl. J. Med. 2001, 345, 1825–1832. [Google Scholar] [CrossRef]
- Partinen, M.; Bjorvatn, B.; Holzinger, B.; Chung, F.; Penzel, Y.; Espie, C.; Morin, C.; ICOSS-Collaboration Group. Sleep and circadian problems during the coronavirus disease 2019 (COVID-19) pandemic: The International COVID-19 Sleep Study (ICOSS). J. Sleep Res. 2020, 30, e13206. [Google Scholar] [CrossRef] [PubMed]
- Altena, E.; Baglioni, C.; Espie, C.A.; Ellis, J.; Gavriloff, D.; Holzinger, B.; Schlarb, A.; Frase, L.; Jernelov, S.; Riemann, D. Dealing with sleep problems during home confinement due to the COVID-19 outbreak: Practical recommendations from a task force of the European CBT-I Academy. J. Sleep Res. 2020, 29, e13052. [Google Scholar] [CrossRef]
- Casagrande, M.; Favieri, F.; Tambelli, R.; Forte, G. The enemy who sealed the world: Effects quarantine due to the COVID-19 on sleep quality, anxiety, and psychological distress in the Italian population. Sleep Med. 2020, 75, 12–20. [Google Scholar] [CrossRef]
- Sher, L. COVID-19, anxiety, sleep disturbances and suicide. Sleep Med. 2020, 70, 124. [Google Scholar] [CrossRef] [PubMed]
- Esme, S.; Bhvs, O.; Souza, J.C. Sleep and immunity in times of COVID-19. Rev. Assoc. Med. Bras. 2020, 66, 143–147. [Google Scholar]
- Miller, M.A.; Cappuccio, F.P. A systematic review of COVID-19 and obstructive sleep apnoea. Sleep Med. Rev. 2021, 55, 101382. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xie, L.; Xu, Y.; Yu, S.; Yao, B.; Xiang, D. Sleep disturbances among medical workers during the outbreak of COVID-2019. Occup Med. 2020, 70, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Marelli, S.; Castelnuovo, A.; Somma, A.; Castronovo, V.; Mombelli, S.; Bottoni, D.; Leitner, C.; Fossati, A.; Ferini-Strambi, L. Impact of COVID-19 lockdown on sleep quality in university students and administration staff. J. Neurol. 2021, 268, 8–15. [Google Scholar] [CrossRef]
- Deng, J.; Zhou, F.; Hou, W.; Silver, Z.; Wong, C.Y.; Chang, O.; Huang, E.; Zuo, Q.K. The prevalence of depression, anxiety, and sleep disturbances in COVID-19 patients: A meta-analysis. Ann. N. Y. Acad. Sci. 2021, 1486, 90–111. [Google Scholar] [CrossRef]
- Liu, Z.; Tang, H.; Jin, Q.; Wang, G.; Yang, Z.; Chen, H.; Yan, H.; Rao, W.; Owens, J. Sleep of preschoolers during the coronavirus disease 2019 (COVID-19) outbreak. J. Sleep Res. 2021, 30, e13142. [Google Scholar] [CrossRef]
- Martínez-de-Quel, Ó.; Suárez-Iglesias, D.; López-Flores, M.; Pérez, C.A. Physical activity, dietary habits and sleep quality before and during COVID-19 lockdown: A longitudinal study. Appetite 2021, 158, 105019. [Google Scholar] [CrossRef]
- Zhao, X.; Lan, M.; Li, H.; Yang, J. Perceived stress and sleep quality among the non-diseased general public in China during the 2019 coronavirus disease: A moderated mediation model. Sleep Med. 2020, 77, 339–345. [Google Scholar] [CrossRef]
- Lucey, B.P. It’s complicated: The relationship between sleep and Alzheimer’s disease in humans. Neurol. Dis. 2020, 144, 105031. [Google Scholar]
- Shokri-Kojori, E.; Wang, G.J.; Wiers, C.E.; Demiral, S.B.; Guo, M.; Kim, S.W.; Lindgren, E.; Ramirez, V.; Zehra, A.; Freeman, C.; et al. Β-Amyloid accumulation in the human brain after one night of sleep deprivation. Proc. Natl. Acad. Sci. USA 2018, 115, 4483–4488. [Google Scholar] [CrossRef] [PubMed]
- Bishir, M.; Bhat, A.; Essa, M.М.; Ekpo, O.; Ihunwo, A.O.; Veeraraghavan, V.P.; Mohan, S.K.; Mahalakshmi, A.M.; Ray, B.; Tuladhar, S.; et al. Sleep deprivation and neurological disorders. Hindawi BioMed Res. Int. 2020, 2020, 19. [Google Scholar]
- Jahrami, H.; BaHammam, A.S.; Bragazzi, N.L.; Saif, Z.; Faris, M.; Vitiello, M.V. Sleep problems during the COVID-19 pandemic by population: A systematic review and meta-analysis. J. Clin. Sleep Med. 2021, 17, 299–313. [Google Scholar] [CrossRef]
- Tony, A.A.; Tony, E.A.; Ali, S.B.; Ezzeldin, A.M.; Mahmoud, A.A. COVID-19-associated sleep disorders: A case report. Neurobiol. Sleep Circadian Rhythms 2020, 9, 100057. [Google Scholar] [CrossRef]
- Hurtado-Alvarado, G.; Becerril-Villanueva, E.; Contis-Montes, O.A.; Domínguez-Salazar, E.; Salinas-Jazmín, N.; Pérez-Tapia, S.M.; Pavon, L.; Velázquez-Moctezumа, J.; Gómez-González, B. The yin/yang of inflammatory status: Blood-brain barrier regulation during sleep. Brain Behav. Immun. 2018, 69, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Kastin, A.J. The blood-brain barrier: Regulatory roles in wakefulness and sleep. Neuroscientist 2017, 23, 124–136. [Google Scholar] [CrossRef]
- Cuddapah, V.A.; Zhang, S.L.; Sehgal, A. Regulation of the blood-brain barrier by circadian rhythms and sleep. Trends Neurosci. 2019, 42, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Medina-Flores, F.; Hurtado-Alvarado, G.; Contis-Montes de Oca, A.; López-Cervantes, S.P.; Konigsberg, M.; Deli, M.A.; Gómez-González, B. Sleep loss disrupts pericyte-brain endothelial cell interactions impairing blood-brain barrier function. Brain Behav. Immun. 2020, 89, 118–132. [Google Scholar] [CrossRef]
- Hurtado-Alvarado, G.; Domínguez-Salazar, E.; Pavon, L.; Velázquez-Moctezuma, J.; Gómez-González, B. Blood-brain barier disruption induced by chronic sleep loss: Low-grade inflammation may be the link. J. Immunol. Res. 2016, 2016, 4576012. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wu, J.; Hua, F.; Chen, Y.; Zhan, F.; Xu, G. Sleep deprivation induces cognitive impairment by increasing blood-brain barrier permeability via CD44. Front. Neurol. 2020, 11, 563916. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.C.; Pack, A.I. Obstructive sleep apnea and cognitive impairment: Addressing the blood-brain barrier. Sleep Med. Rev. 2014, 18, 35–48. [Google Scholar] [CrossRef]
- Daulatzai, M.A. Cerebral hypoperfusion and glucose hypometabolism: Key pathophysiological modulators promote neurodegeneration, cognitive impairment, and Alzheimer’s disease. J. Neurosci. Res. 2017, 95, 943–972. [Google Scholar] [CrossRef]
- Voirin, A.C.; Celle, S.; Perek, N.; Roche, F. Sera of elderly obstructive sleep apnea patients alter blood-brain barrier integrity in vitro: A pilot study. Sci. Rep. 2020, 10, 11309. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Alvarado, G.; Pavón, L.; Castillo-García, S.A.; Hernández, M.E.; Domínguez-Salazar, E.; Velázquez-Moctezuma, J.; Gómez-González, B. Sleep loss as a factor to induce cellular and molecular inflammatory variations. Clin. Dev. Immunol. 2013, 2013, 801341. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Cedernaes, J.; Giedraitis, V.; Nilsson, E.K.; Hogenkamp, P.S.; Vågesjö, E.; Massena, S.; Pettersson, U.; Christoffersson, G.; Phillipson, M.; et al. Acute sleep deprivation increases serum levels of neuron-specific enolase (NSE) and S100 calcium binding protein B (S-100B) in healthy young men. Sleep 2014, 37, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Opp, M.R.; George, A.; Ringgold, K.M.; Hansen, K.M.; Bullock, K.M.; Banks, W.A. Sleep fragmentation and sepsis difeetially impact blood-brain barrier integrity and transport of tumor necrosis factorin aging. Brain Behav. Immun. 2015, 50, 259–265. [Google Scholar] [CrossRef]
- Krueger, J.M.; Majde, J.A. Humoral links between sleep and the immune system: Research issues. Ann. N. Y. Acad. Sci. 2003, 992, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.; Männel, D. How the immune system puts the brain to sleep. Nat. Med. 1999, 5, 877–879. [Google Scholar] [CrossRef]
- Hirotsu, C.; Rydlewski, M.; Araújo, M.S.; Tufik, S.; Andersen, M.L. Sleep loss and cytokines levels in an experimental model of psoriasis. PLoS ONE 2012, 7, e51183. [Google Scholar] [CrossRef]
- He, J.; Hsuchou, H.; He, Y.; Kastin, A.J.; Wang, Y.; Pan, W. Sleep restriction impairs blood-brain barrier function. J. Neurosci. 2014, 34, 14697–14706. [Google Scholar] [CrossRef]
- Ibrahim, L.; Duncan, W.; Luckenbaugh, D.A.; Yuan, P.; Machado-Vieira, R.; Zarate, C.A., Jr. Rapid antidepressant changes with sleep deprivation in major depressive disorder are associated with changes in vascular endothelial growth factor (VEGF): A pilot study. Brain Res. Bull. 2011, 86, 129–133. [Google Scholar] [CrossRef][Green Version]
- Li, K.; Wei, P.; Qin, Y.; Wei, Y. Is C-reactive protein a marker of obstructive sleep apnea? Medicine 2017, 96, e6850. [Google Scholar] [CrossRef] [PubMed]
- Gabryelska, A.; Szmyd, B.; Panek, M.; Szemraj, J.; Kuna, P.; Białasiewicz, P. Serum hypoxia-inducible factor-1α protein level as a diagnostic marker of obstructive sleep apnea. Pol. Arch. Intern. Med. 2020, 130, 158–160. [Google Scholar] [CrossRef]
- Hurtado-Alvarado, G.; Domínguez-Salazar, E.; Velázquez-Moctezuma, J.; Gómez-González, В. A2A adenosine receptor antagonism reverts the blood-brain barrier dysfunction induced by sleep restriction. PLoS ONE 2016, 11, e0167236. [Google Scholar] [CrossRef]
- Gomez-Gonzalez, B.; Hurtado-Alvarado, G.; Esqueda-Leon, E.; Santana-Miranda, R.; Rojas-Zamorano, J.А.; Velazquez-Moctezuma, J. REM Sleep Loss and Recovery Regulates Blood-Brain Barrier Function. Curr. Neurovasc. Res. 2013, 10, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Alvarado, G.; Velázquez-Moctezuma, J.; Gómez-González, B. Chronic sleep restriction disrupts interendothelial tight junctions in the hippocampus and increases blood-brain barrier permeability. J. Microsc. 2017, 268, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Saunders, N.R.; Dziegielewska, K.M.; Mollgard, K.; Habgood, M.D. Markers for blood-brain barrier integrity: How apprpriate is Evans blue in the twenty-first century and what are the alternatives. Front. Neurosci. 2015, 9, 385. [Google Scholar] [CrossRef]
- Chennaoui, M.; Sauvet, F.; Drogou, C.; Van Beers, P.; Langrume, C.; Guillard, M.; Gourby, B.; Bourrilhon, C.; Florence, G.; Gomez-Merino, D. Effect of one night of sleep loss on changes in tumor necrosis factor alpha (TNF-α) levels in healthy men. Cytokine 2011, 56, 318–324. [Google Scholar] [CrossRef]
- Zielinski, M.R.; Kim, Y.; Karpova, S.A.; McCarley, R.W.; Strecker, R.E.; Gerashchenko, D. Chronic sleep restriction elevates brain interleukin-1 beta and tumor necrosis factor-alpha and attenuates brain-derived neurotrophic factor expression. Neurosci. Lett. 2014, 580, 27–31. [Google Scholar] [CrossRef]
- Vgontzas, A.N.; Zoumakis, M.; Papanicolaou, D.A.; Bixler, E.; Prolo, P.; Lin, H.-M.; Vela-Bueno, A.; Kales, A.; Chrousos, G. Chronic insomnia is associated with a shift of interleukin-6 and tumor necrosis factor secretion from nighttime to daytime. Metabolism 2002, 51, 887–892. [Google Scholar] [CrossRef]
- Van Leeuwen, W.M.A.M.; Karisola, L.P.; Karisola, P.; Lindholm, H.; Luukkonen, R.; Sallinen, M.; Alenius, H. Sleep restriction increases the risk of developing cardiovascular diseases by augmenting proinflammatory responses through IL-17 and CRP. PLoS ONE 2009, 4, e4589. [Google Scholar] [CrossRef]
- Okun, M.L.; Coussons-Read, M.; Hall, M. Disturbed sleep is associated with increased C-reactive protein in young women. Brain Behav. Immun. 2009, 23, 351–354. [Google Scholar] [CrossRef]
- Carver, K.A.; Lourim, D.; Tryba, A.K.; Harder, D.R. Rhythmic expression of cytochrome P450 epoxygenases CYP4×1 and CYP2c11 in the rat brain and vasculature. Am. J. Physiol. Cell Physiol. 2014, 307, 989. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Meng, D.; Sun, N.; Zhu, Z.; Zhao, R.; Lu, C.; Chen, S.; Hua, L.; Qian, R. Clock upregulates intercellular adhesion molecule-1 expression and promotes mononuclear cells adhesion to endothelial cells. Biochem. Biophys. Res. Commun. 2014, 443, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Mohawk, J.A.; Green, C.; Takahashi, J. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef]
- Davidson, A.J.; London, B.; Block, G.; Menaker, M. Cardiovascular tissues contain independent circadian clocks. Clin. Exp. Hypertens. 2005, 27, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Durgan, D.J.; Grossland, R.; Bryan, J. The rat cerebral vasculature exhibits time-of-day-dependent oscillations in circadian clock genes and vascular function that are attenuated following obstructive sleep apnea. J. Cereb. Blood Flow Metab. 2017, 37, 2806–2819. [Google Scholar] [CrossRef]
- Shintaro, H.; Mona, U.; Kumiko, T.; Rie, H.; Yuki, T.; Masaaki, M.; Toshio, H. How COVID-19 induces cytokine storm with high mortality. Inflamm. Regen. 2020, 40, 37. [Google Scholar]
- Tay, M.Z.; Poh, C.M.; Renia, L.; MacAry, P.A.; Ng, L.F.P. The trinity of COVID-19: Immunity, inflammation and intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qui, P.; Wang, J.; Liu, J.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Coronaviridae Study Group of the International Committee on Taxonomy of V. The species severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, J.; Song, B.; Gi, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Liu, B.; Li, M.; Zhou, Z.; Guan, X.; Xiang, Y. Can we use interleukin-6 (IL-6) blockade for coronavirus disease 2019 (COVID-19)-induced cytokine release syndrome (CRS)? J. Autoimmun. 2020, 111, 102452. [Google Scholar] [CrossRef]
- Hirano, T.; Murakami, M. COVID-19: A new virus, but a familiar receptor and cytokine release syndrome. Immunity 2020, 52, 731–733. [Google Scholar] [CrossRef]
- Mahmudpour, M.; Roozbeh, J.; Keshavarz, M.; Farrokhi, S.; Nabipour, I. COVID-19 cytokine storm: The anger of inflammation. Cytokine 2020, 133, 155151. [Google Scholar] [CrossRef] [PubMed]
- McGonagle, D.; Sharif, K.; O’Regan, A.; Bridgewood, C. The role of cytokines including interleukin-6 in COVID-19 induced pneumonia and macrophage activation syndrome-like disease. Autoimmun. Rev. 2020, 9, 102537. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Cornélissen, G.; Halberg, F.; Kastin, A.J. Selected contribution: Circadian rhythm of tumor necrosis factor-alpha uptake into mouse spinal cord. J. Appl. Physiol. 2002, 92, 62. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Kastin, А.J. TNFalpha transport across the blood-brain barrier is abolished in receptor knockout mice. Exp. Neurol. 2002, 174, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Agorastos, A.; Hauger, R.L.; Barkauskas, D.A.; Moeller-Bertram, T.; Clopton, P.L.; Haji, U.; Lohr, J.B.; Geracioti, T.D., Jr.; Patel, P.M.; Chrousos, G.P.; et al. Circadian rhythmicity, variability and correlation of interleukin-6 levels in plasma and cerebrospinal fluid of healthy men. Psychoneuroendocrinology 2014, 44, 71–82. [Google Scholar] [CrossRef]
- Banks, W.A.; Kastin, A.J.; Gutierrez, E.G. Penetration of interleukin-6 across the murine blood-brain barrier. Neurosci. Lett. 1994, 179, 53–56. [Google Scholar] [CrossRef]
- Banks, W.A. Diurnal uptake of circulating interleukin-1alpha by brain, spinal cord, testis and muscle. Neuroimmunomodulation 1998, 5, 36–41. [Google Scholar] [CrossRef]
- Choy, E.H.; De Benedetti, F.; Takeuchi, T.; Hashizume, M.; John, M.R.; Kishimoto, T. Translating IL-6 biology into effective treatments. Nat. Rev. Rheumatol. 2020, 16, 335–345. [Google Scholar] [CrossRef]
- Maric, J.; Ravindran, A.; Mazzurana, L.; Van Acker, A.; Rao, A.; Kokkinou, E.; Ekoff, M.; Thomas, D.; Fauland, A.; Nilsson, G.; et al. Cytokine-induced endogenous production of prostaglandin D2 is essential for human group 2 innate lymphoid cell activation. J. Allergy Clin. Immunol. 2019, 143, 2202–2214. [Google Scholar] [CrossRef]
- Oyesola, O.O.; Duque, C.; Huang, L.C.; Larson, E.M.; Früh, S.P.; Webb, L.M.; Peng, S.A.; Tait Wojno, E.D. The Prostaglandin D2 Receptor CRTH2 Promotes IL-33–Induced ILC2 Accumulation in the Lung. J. Immunol. 2020, 204, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Pandey, H.P.; Ram, A.; Matsumura, H.; Hayaishi, O. Concentration of prostaglandin D2 in cerebrospinal fluid exhibits a circadian alteration in conscious rats. Biochem. Mol. Biol. Int. 1997, 37, 431–437. [Google Scholar]
- Ram, A.; Pandey, H.P.; Matsumura, H.; Kasahara-Orita, K.; Nakajima, T.; Takahata, R.; Satoh, S.; Terao, A.; Hayaishi, O. CSF levels of prostaglandins, especially the level of prostaglandin D2, are correlated with increasing propensity towards sleep in rats. Brain Res. 1997, 751, 81–89. [Google Scholar] [CrossRef]
- McNamee, E.N.; Ryan, K.M.; Griffin, E.W.; González-Reyes, R.E.; Ryan, K.J.; Harkin, A.; Connor, T.J. Noradrenaline acting at central beta-adrenoceptors induces interleukin-10 and suppressor of cytokine signaling-3 expression in rat brain: Implications for neurodegeneration. Brain Behav. Immun. 2010, 24, 660–671. [Google Scholar] [CrossRef]
- O’Neill, E.; Harkin, A. Targeting the noradrenergic system for anti-inflammatory and neuroprotective effects: Implications for Parkinson’s disease. Neural Regen. Res. 2018, 13, 1332–1337. [Google Scholar] [PubMed]
- Staedtke, V.; Bai, R.Y.; Kim, K.; Darvas, M.; Davilo, M.L.; Riggins, G.J.; Rothman, P.B.; Papadopoulos, N.; Kinzler, K.W.; Vogelstein, B.; et al. Disruption of a self-amplifying catecholamine loop reduces cytokine release syndrome. Nature 2018, 564, 273–277. [Google Scholar] [CrossRef]
- Pan, L.; Dong, L. Epinephrine use in COVID-19: Friend or foe? Eur. J. Hosp. Pharm. 2021, 28, e1. [Google Scholar]
- Wood, J.H.; Ziegler, M.C.; Lake, C.R.; Ebert, M.H. Circadian rhythm in cerebrospinal fluid noradrenaline of man and monkey. Nature 1976, 264, 656–658. [Google Scholar] [CrossRef] [PubMed]
- Manshardt, J.; Wurtman, R. Daily rhythm in the noradrenaline content of rat hypothalamus. Nature 1968, 217, 574–575. [Google Scholar] [CrossRef] [PubMed]
- Reis, D.J.; Wurtman, R.J. Diurnal changes in brain noradrenalin. Life Sci. 1968, 7, 91–98. [Google Scholar] [CrossRef]
- Reis, D.J.; Weinbren, M.; Corvelli, A. A circadian rhythm of norepinephrine regionally in cat brain: Its relationship to environmental lighting and to regional diurnal variations in brain serotonin. J. Pharmacol. Exp. Ther. 1968, 164, 135–145. [Google Scholar]
- Freidman, A.H.; Walker, C.A. Circadian rhythms in rat mid-brain and caudate nucleus biogenic amine levels. J. Physiol. 1968, 197, 77–85. [Google Scholar] [CrossRef]
- Lew, G.M.; Quay, W.B. The mechanism of circadian rhythms in brain and organ contents of norepinephrine: Circadian changes in the effects of methyltyrosine and 6-hydroxydopamine. Comp. Gen. Pharmacol. 1973, 4, 375–381. [Google Scholar] [CrossRef]
- Eleftheriou, B.S. Circadian rhythm in blood and brain biogenic amines and other biochemical changes in rabbits. Brain Res. 1974, 75, 145–152. [Google Scholar] [CrossRef]
- Mitchell, H.A.; Weinshenker, D. Good night and good luck: Norepinephrine in sleep pharmacology. Biochem. Pharm. 2010, 79, 801–809. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Bloom, F.E. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J. Neurosci. 1981, 1, 876–886. [Google Scholar] [CrossRef]
- Shimba, A.; Ikuta, K. Glucocorticoids regulate circadian rhythm of innate and adaptive immunity. Front. Immunol. 2020, 11, 2143. [Google Scholar] [CrossRef]
- Szmyd, B.; Rogut, M.; Białasiewicz, P.; Gabryelska, A. The impact of glucocorticoids and statins on sleep quality. Sleep Med. Rev. 2021, 55, 101380. [Google Scholar] [CrossRef]
- Daneman, R.; Prat, A. The blood-brain barrier. Cold Spring Harb Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef]
- Abbott, J.; Patabendige, A.; Dolman, D.; Yusof, S.; Begley, D. Structure and function of the blood–brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- King, M.; Su, W.; Chang, A.; Zuckerman, A.; Pasternak, G.W. Transport of opioids from the brain to the periphery by P-glycoprotein: Peripheral actions of central drugs. Nat. Neurosci. 2001, 4, 268–274. [Google Scholar] [CrossRef]
- Loscher, W.; Potschka, H. Blood-brain barrier active efflux transporters: ATP-binding cassette gene family. NeuroRx 2005, 2, 86–98. [Google Scholar] [CrossRef]
- Kervezee, L.; Hartman, R.; van den Berg, D.J.; Shimizu, S.; Emoto-Yamamoto, Y.; Meijer, J.H.; de Lange, E.C. Diurnal variation in P-glycoprotein-mediated transport and cerebrospinal fluid turnover in the brain. AAPS J. 2014, 16, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.L.; Lahens, N.F.; Yue, Z.; Arnold, D.M.; Pakstis, P.P.; Schwarz, J.E.; Sehgal, A. A circadian clock in the blood-brain barrier regulates xenobiotic efflux. Cell 2018, 173, 130–139.e10. [Google Scholar] [CrossRef]
- Hindle, S.J.; Munji, R.N.; Dolghih, E.; Gaskins, G.; Orng, S.; Ishimoto, H.; Soung, A.; DeSalvo, M.; Kitamoto, T.; Keiser, M.J.; et al. Evolutionarily Conserved roles for blood-brain barrier xenobiotic transporters in endogenous steroid partitioning and behavior. Cell Rep. 2017, 21, 1304–1316. [Google Scholar] [CrossRef]
- Baig, A.M. Neurological manifestations in COVID-19 caused by SARS-CoV-2. CNS Neurosci. 2020, 26, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A multibasic cleavage site in the Spike protein of SARS-CoV-2 is essential for infection of human lung cells. Mol. Cell 2020, 78, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mayi, B.S.; Leibowitz, J.A.; Woods, A.T.; Ammon, K.A.; Liu, A.E.; Raja, A. The role of Neuropilin-1 in COVID-19. PLoS Pathog. 2021, 17, e1009153. [Google Scholar] [CrossRef] [PubMed]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Hamming, W.T.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, Y.; Mangalam, A.K.; Guo, Y.; LaFrance-Corey, R.G.; Gamez, J.D.; Atanga, P.A.; Clarkson, B.D.; Zhang, Y.; Wang, E.; et al. Neuropilin-1 modulates interferon-gamma-stimulated signaling in brain microvascular endothelial cells. J. Cell Sci. 2016, 129, 3911–3921. [Google Scholar]
- Paniz-Mondolfi, A.; Bryce, C.; Grimes, Z.; Gordon, R.E.; Reidy, J.; Lednicky, J.; Sordillo, E.M.; Fowkes, M. Central nervous system involvement by severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). J. Med. Virol. 2020, 92, 699–702. [Google Scholar] [CrossRef] [PubMed]
- Burks, S.M.; Rosas-Hernandez, H.; Alenjandro Ramirez-Lee, M.; Cuevas, E.; Talpos, J.C. Can SARS-CoV-2 infect the central nervous system via the olfactory bulb or the blood-brain barrier? Brain Behav. Immun. 2021, 95, 7–14. [Google Scholar] [CrossRef]
- Bleau, C.; Filliol, A.; Samson, M.; Lamontagne, L.; Perlman, S. Brain invasion by mouse hepatitis virus depends on impairment of tight junctions and beta interferon production in brain microvascular endothelial cells. J. Virol. 2015, 89, 9896–9908. [Google Scholar] [CrossRef]
- Cabirac, G.F.; Soike, K.F.; Butunoi, C.; Hoel, K.; Johnson, S.; Cai, G.Y.; Murray, R.S. Coronavirus JHM OMP1 pathogenesis in owl monkey CNS and coronavirus infection of owl monkey CNS via peripheral routes. Adv. Exp. Med. Biol. 1993, 342, 347–352. [Google Scholar]
- Cabirac, G.F.; Soike, K.F.; Zhang, J.Y.; Hoel, K.; Butunoi, C.; Cai, G.Y.; Johnson, S.; Murray, R.S. Entry of coronavirus into primate CNS following peripheral infection. Microb. Pathog. 1994, 16, 349–357. [Google Scholar] [CrossRef]
- Cabirac, G.F.; Murray, R.S.; McLaughlin, L.B.; Skolnick, D.M.; Hogue, B.; Dorovini-Zis, K.; Didier, P.J. In vitro interaction of coronaviruses with primate and human brain microvascular endothelial cells. Adv. Exp. Med. Biol. 1995, 380, 79–88. [Google Scholar]
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood-brain barrier. Neurobiol. Dis. 2020, 146, 105131. [Google Scholar] [CrossRef]
- Bellon, M.; Schweblin, C.; Lambeng, N.; Cherpillod, P.; Vazquez, J.; Lalive, P.H.; Schibler, M.; Deffert, C. Cerebrospinal fluid features in SARS-CoV-2 RT-PCR positive patients. Clin. Infect. Dis. 2020, ciaa1165. [Google Scholar] [CrossRef]
- Alam, S.B.; Willows, S.; Kulka, M.; Sandhu, J.K. Severe acute respiratory syndrome coronavirus 2 may be an underappreciated pathogen of the central nervous system. Eur. J. Neurol. 2020, 27, 2348–2360. [Google Scholar] [CrossRef]
- Robinson, C.P.; Busl, K.M. Neurologic manifestations of severe respiratory viral contagions. Crit. Care Explor. 2020, 2, e0107. [Google Scholar] [CrossRef]
- Achar, A.; Ghosh, C. COVID-19-associated neurological disorders: The potential route of CNS invasion and blood-brain barrier relevance. Cells 2020, 9, 2360. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Gong, E.; Zhang, B.; Zheng, J.; Gao, Z.; Zhong, Y.; Zou, W.; Zhan, J.; Wang, S.; Xie, Z.; et al. Multiple organ infection and the pathogenesis of SARS. J. Exp. Med. 2005, 202, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, J.M.; Butany, J.; Poon, L.L.M.; Chan, K.H.; Beh, S.L.; Poutanen, S.; Peiris, J.S.M.; Wong, M. Time course and cellular localization of SARS-CoV nucleoprotein and RNA in lungs from fatal cases of SARS. PLoS Med. 2006, 3, e27. [Google Scholar] [CrossRef]
- Spiegel, M.; Schneider, K.; Weber, F.; Weidmann, M.; Hufert, F.T. Interaction of severe acute respiratory syndrome-associated coronavirus with dendritic cells. J. Gen. Virol. 2006, 87, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Trojanowicz, B.; Ulrich, C.; Kohler, F.; Bode, V.; Seibert, E.; Fiedler, R.; Girndt, M. Monocytic angiotensin-converting enzyme 2 relates to atherosclerosis in patients with chronic kidney disease. Nephrol. Dial. Transpl. 2017, 32, 287–298. [Google Scholar] [CrossRef][Green Version]
- Sankowski, R.; Mader, S.; Valdés-Ferrer, S.I. Systemic Inflammation and the Brain: Novel Roles of Genetic, Molecular, and Environmental Cues as Drivers of Neurodegeneration. Front. Cell. Neurosci. 2015, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, M.; Lanza, G.; Falzone, L.; Fisicaro, F.; Ferri, R.; Bella, R. SARS-CoV-2 and the nervous system: From clinical features to molecular mechanisms. Int. J. Mol. Sci. 2020, 21, 5475. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J.; HLH Across Speciality Collaboration, UK. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Steardo, L.; Steardo, L., Jr.; Zorec, R.; Verkhratsky, A. Neuroinfection may contribute to pathophysiology and clinical manifestations of COVID-19. Acta Physiol. 2020, 229, e13473. [Google Scholar] [CrossRef]
- Varatharaj, A.; Galea, I. The blood-brain barrier in systemic inflammation. Brain Behav. Immun. 2017, 60, 1–12. [Google Scholar] [CrossRef]
- Koh, L.; Zakharov, A.; Johnston, M. Integration of the subarachnoid space and lymphatics: Is it time to embrace a new concept of cerebrospinal fluid absorption? Cereb. Fluid Res. 2005, 2, 6. [Google Scholar] [CrossRef]
- Cserr, H.F.; Knopf, P.M. Cervical lymphatics, the blood-brain barrier and the immunoreactivity of the brain: A new view. Immunol. Today 1992, 13, 507–512. [Google Scholar] [CrossRef]
- Bradbury, M. Lymphatics and the central nervous system. Trends Neurosci. 1981, 4, 100–101. [Google Scholar] [CrossRef]
- Ma, Q.; Ries, M.; Decker, Y.; Muller, A.; Riner, C.; Bucker, A.B.; Fassbender, K.; Detmar, M.; Proulx, S.T. Rapid lymphatic efflux limits cerebrospinal fluid flow to the brain. Acta Neuropathol. 2019, 137, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Weller, R.O.; Kida, S.; Zhang, E.T. Pathways of fluid drainage from the brain-morphological aspects and immunological significance in rat and man. Brain Pathol. 1992, 2, 277–284. [Google Scholar] [CrossRef]
- Kida, S.; Pantazis, A.; Weller, R. CSF drains directly from the subarachnoid space into nasal lymphatics in the rat. anatomy, histology and immunological significance. Neuropathol. Appl. Neurobiol. 1993, 19, 480–488. [Google Scholar] [CrossRef]
- Johnston, M.; Zakharov, A.; Papaiconomou, C.; Salmasi, G.; Armstrong, D. Evidence of connections between cerebrospinal fluid and nasal lymphatic vessels in humans, non-human primates and other mammalian species. Cereb. Fluid Res. 2004, 1, 2. [Google Scholar] [CrossRef]
- De Leon, M.J.; Li, Y.; Okamura, N.; Tsui, W.H.; Saint-Louis, L.A.; Glodzik, L.; Osorio, R.S.; Fortea, J.; Butler, T.; Pirraglia, E.; et al. Cerebrospinal fluid clearance in Alzheimer’s disease measured with dynamic pet. J. Nucl. Med. 2017, 58, 1471–1476. [Google Scholar] [CrossRef]
- Song, J.; Lee, S.-S.; Lim, S.; Yeo, S. Mechanism of the neuroprotective effect of injecting brain cells on st36 in an animal model of Parkinson’s disease. Neurosci. Lett. 2020, 717, 34698. [Google Scholar] [CrossRef]
- Zhao, P.; Le, Z.; Liu, L.; Chen, Y. Therapeutic delivery to the brain via the lymphatic vasculature. Nano Lett. 2020, 20, 5415–5420. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O. Biophotonic strategies of measurement and stimulation of the cranial and the extracranial lymphatic drainage function. IEEE J. Sel. Top. Quantum Electron. 2021, 27, 1–13. [Google Scholar] [CrossRef]
- Li, J.; Gao, J.; Xu, Y.P.; Zhou, T.L.; Jin, Y.Y.; Lou, J.N. Expression of severe acute respiratory syndrome coronavirus receptors, ACE2 and CD209L in different organ derived microvascular endothelial cells. Zhonghua Yi Xue Za Zhi 2007, 87, 833–837. (In Chinese) [Google Scholar] [PubMed]
- Butowt, R.; Christopher, S.B. Anosmia in COVID-19: Underlying mechanisms and assessment of an olfactory route to brain infection. Neuroscientist 2020, 11, 1073858420956905. [Google Scholar] [CrossRef]
- Aragão, M.D.F.V.V.; Leal, M.C.; Cartaxo Filho, O.Q.; Fonseca, T.M.; Valença, M.M. Anosmia in covid-19 associated with injury to the olfactory bulbs evident on mri. Am. J. Neuroradiol. 2020, 41, 1703–1706. [Google Scholar] [CrossRef]
- Baig, A.M.; Khaleeq, A.; Ali, U.; Syeda, H. Evidence of the covid-19 virus targeting the CNS: Tissue distribution, host–virus interaction, and proposed neurotropic mechanisms. ACS Chem. Neurosci. 2020, 11, 995–998. [Google Scholar] [CrossRef]
- Briguglio, M.; Bona, A.; Porta, M.; Dell’Osso, B.; Pregliasco, F.E.; Banfi, G. Disentangling the hypothesis of host dysosmia and sars-cov 2: The bait symptom that hides neglected neurophysiological routes. Front. Physiol. 2020, 11, 671. [Google Scholar] [CrossRef] [PubMed]
- Gilani, S.; Roditi, R.; Naraghi, M. Covid-19 and anosmia in tehran, iran. Med. Hypotheses 2020, 141, 109757. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Galougahi, M.; Yousefi-Koma, A.; Bakhshayeshkaram, M.; Raad, N.; Haseli, S. 18fdg pet/ct scan reveals hypoactive or bitofrontal cortex in anosmia of covid-19. Acad. Radiol. 2020, 27, 1042–1043. [Google Scholar] [CrossRef] [PubMed]
- Beeraka, N.M.; Sadhu, S.P.; Madhunapantula, S.V.; Pragada, R.R.; Svistunov, A.A.; Nikolenko, V.N.; Mikhaleva, L.M.; Aliev, G. Strategies for targeting sars cov-2: Small molecule inhibitors—The current status. Front. Immunol. 2020, 11, 552925. [Google Scholar] [CrossRef] [PubMed]
- Chassidim, Y.; Veksler, R.; Lublinsky, S.; Pell, G.; Friedman, A.; Shelef, I. Quantitative imaging assessment of blood-brain barrier permeability in humans. Fluids Barriers CNS 2013, 10, 9. [Google Scholar] [CrossRef]
- Heye, A.K.; Culling, R.D.; Valdés Hernández, M.C.; Thrippleton, M.J.; Wardlaw, J.M. Assessment of blood–brain barrier disruption using dynamic contrast-enhanced MRI. A systematic review. NeuroImage Clin. 2014, 6, 262–274. [Google Scholar] [CrossRef]
- Neuwelt, E.A.; Maravilla, K.; Frenkel, E.; Ripaport, S.; HilL, S.; Barnett, P. Osmotic blood-brain barrier disruption. Computerized tomographic monitoring of chemotherapeutic agent delivery. J. Clin. Investig. 1979, 64, 684–688. [Google Scholar] [CrossRef]
- Roman-Goldstein, S.; Clunie, D.; Stevens, D.; Hogan, R.; Monard, J.; Ramsay, F.; Neuwelt, E. Osmotic blood-brain barrier disruption: CT and radionuclide imaging. AJNR Am. J. Neuroradiol. 1994, 15, 581–590. [Google Scholar]
- Rogosnitzky, M.; Branch, S. Gadolinium-based contrast agent toxicity: A review of known and proposed mechanisms. Biometals 2016, 29, 365–376. [Google Scholar] [CrossRef]
- Kaller, M.O.; An, J.; Contrast Agent Toxicity. StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK537159/ (accessed on 10 May 2021).
- Elbeshlawi, I.; AbdelBaki, M.S. Safety of gadolinium administration in children. Pediatr. Neurol. 2018, 86, 27–32. [Google Scholar] [CrossRef]
- Perazella, M.A. Gadolinium-contrast toxicity in patients with kidney disease: Nephrotoxicity and nephrogenic systemic fibrosis. Curr. Drug. Saf. 2008, 3, 67–75. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Esmat, A.; Bragin, D.; Bragina, O.; Shirokov, A.; Navolokin, N.; Yang, Y.; Abdurashitov, A.; Khorovodov, A.; Terskov, A. Phenomenon of music-induced opening of the blood-brain barrier in healthy mice. Proc. R. Soc. B 2020, 287, 20202337. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Chehonin, V.; Borisova, E.; Fedosov, I.; Namykin, A.; Abdurashitov, A.; Shirokov, A.; Khlebtsov, B.; Lyubun, Y.; Navolokin, N.; et al. Photodynamic opening of the blood-brain barrier and pathways of brain clearing. J. Biophotonics 2018, 11, e201700287. [Google Scholar] [CrossRef]
- Lipsman, N.; Meng, Y.; Bethune, A.J.; Huang, Y.; Lam, B.; Masellis, M.; Herrmann, N.; Heyn, C.; Aubert, I.; Boutet, A.; et al. Blood–brain barrier opening in Alzheimer’s disease using MR-guided focused ultrasound. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Jordão, J.F.; Thévenot, E.; Markham-Coultes, K.; Scarcelli, T.; Weng, Y.Q.; Xhima, K.; O’Reilly, M.; Huang, Y.; McLaurin, J.; Hynynen, K.; et al. Amyloid-β plaque reduction, endogenous antibody delivery and glial activation by brain-targeted, transcranial focused ultrasound. Exp. Neurol. 2013, 248, 16–29. [Google Scholar] [CrossRef]
- Leinenga, G.; Götz, J. Scanning ultrasound removes amyloid-β and restores memory in an Alzheimer’s disease mouse model. Sci. Transl. Med. 2015, 7, 278ra33. [Google Scholar] [CrossRef]
- Burgess, A.; Dubey, S.; Yeung, S.; Hough, O.; Eterman, N.; Aubert, I.; Hynynen, K. Alzheimer disease in a mouse model: MR imaging–guided focused ultrasound targeted to the hippocampus opens the blood-brain barrier and improves pathologic abnormalities and behavior. Radiology 2014, 273, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Fultz, N.E.; Bonmassar, G.; Setsompop, K.; Stickgold, R.A.; Rosen, B.R.; Polimeni, J.R.; Lewis, L.D. Coupled electrophysiological, hemodynamic, and cerebrospinal fluid oscillations in human sleep. Science 2019, 366, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, A.; Dubrovsky, A.; Koronovskii, A., Jr.; Pavlova, O.; Semyachkina-Glushkovskaya, O.; Kurths, J. Extended detrended fluctuation analysis of sound-induced changes in brain electrical activity. Chaos Solitons Fractals 2020, 139, 109989. [Google Scholar] [CrossRef]
- Pavlov, A.; Dubrovsky, A.; Koronovskii, A., Jr.; Pavlova, O.; Semyachkina-Glushkovskaya, O.; Kurths, J. Extended detrended fluctuation analysis of electroencephalograms signals during sleep and the opening of the blood–brain barrier. Chaos Interdiscip. J. Nonlinear Sci. 2020, 30, 073138. [Google Scholar] [CrossRef] [PubMed]
- Hablitz, L.M.; Vinitsky, H.S.; Sun, Q.; Stæger, F.F.; Sigurdsson, B.; Mortensen, K.N.; Lilius, T.O.; Nedergaard, M. Increased glymphatic influx is correlated with high EEG delta power and low heart rate in mice under anesthesia. Sci. Adv. 2019, 5, eaav5447. [Google Scholar] [CrossRef] [PubMed]
- Kiviniemi, V.; Korhonen, V.; Kortelainen, J.; Rytky, S.; Keinänen, T.; Tuovinen, T.; Isokangas, M.; Sonkajärvi, E.; Siniluoto, T.; Nikkinen, J.; et al. Real-time monitoring of human blood-brain barrier disruption. PLoS ONE 2017, 12, e0174072. [Google Scholar] [CrossRef]
- Shuvaev, A.; Kuvacheva, N.; Morgun, A.; Khilazheva, E.; Salmina, A. The role of ion channels expressed in cerebral endothelial cells in the functional integrity of the blood-brain barrier (Review). Sovrem. Tehnol. V Med. 2016, 8, 241–250. [Google Scholar] [CrossRef]
- Callies, C.; Fels, J.; Liashkovich, I.; Kliche, K.; Jeggle, P.; Kusche-Vihrog, K.; Oberleithner, H. Membrane potential depolarization decreases the stiffness of vascular endothelial cells. J. Cell Sci. 2011, 124, 1936–1942. [Google Scholar] [CrossRef] [PubMed]
- Vanhatalo, S.; Voipio, J.; Kaila, K. Infraslow. EEG activity. In Niedermeyer’s Electroencephalography: Basic Principles, Clinical Applications, and Related Fields; Wolters Kluwer/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2011; pp. 741–747. [Google Scholar]
- Woody, C.; Marshall, W.; Besson, J.; Thompson, H.; Aleonard, P.; Albe-Fessard, D. Brain potential shift with respiratory acidosis in the cat and monkey. Am. J. Physiol. Leg. Content 1970, 218, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Revest, P.A.; Jones, H.C.; Abbott, N.J. The transendothelial DC potential of rat blood-brain barrier vessels in situ. In Frontiers in Cerebral Vascular Biology; Springer: Berlin/Heidelberg, Germany, 1993; pp. 71–74. [Google Scholar]
- Revest, P.A.; Jones, H.C.; Abbott, N.J. Transendothelial electrical potential across pial vessels in anaesthetised rats: A study of ion permeability and transport at the blood-brain barrier. Brain Res. 1994, 652, 76–82. [Google Scholar] [CrossRef]
- Monto, S.; Palva, S.; Voipio, J.; Palva, J.M. Very slow EEG fluctuations predict the dynamics of stimulus detection and oscillation amplitudes in humans. J. Neurosci. 2008, 28, 8268–8272. [Google Scholar] [CrossRef]
- Hiltunen, T.; Kantola, J.; Abou Elseoud, A.; Lepola, P.; Suominen, K.; Starck, T.; Nikkinen, J.; Remes, J.; Tervonen, O.; Palva, S.; et al. Infra-slow EEG fluctuations are correlated with resting-state network dynamics in fMRI. J. Neurosci. 2014, 34, 356–362. [Google Scholar] [CrossRef]
- Brockett, A.T.; Kane, G.A.; Monari, P.K.; Briones, B.A.; Vigneron, P.A.; Barber, G.A.; Bermudez, A.; Dieffenbach, U.; Kloth, A.D.; Buschman, T.J.; et al. Evidence supporting a role for astrocytes in the regulation of cognitive flexibility and neuronal oscillations through the Ca2+ binding protein S100β. PLoS ONE 2018, 13, e0195726. [Google Scholar] [CrossRef] [PubMed]
- Bellot-Saez, A.; Cohen, G.; van Schaik, A.; Ooi, L.; Morley, J.W.; Buskila, Y. Astrocytic modulation of cortical oscillations. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Henneberger, C.; Papouin, T.; Oliet, S.H.; Rusakov, D.A. Long-term potentiation depends on release of D-serine from astrocytes. Nature 2010, 463, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Takata, N.; Mishima, T.; Hisatsune, C.; Nagai, T.; Ebisui, E.; Mikoshiba, K.; Hirase, H. Astrocyte calcium signaling transforms cholinergic modulation to cortical plasticity in vivo. J. Neurosci. 2011, 31, 18155–18165. [Google Scholar] [CrossRef]
- Bellesi, M.; de Vivo, L.; Tononi, G.; Cirelli, C. Effects of sleep and wake on astrocytes: Clues from molecular and ultrastructural studies. BMC Biol. 2015, 13, 66. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Postnov, D.; Penzel, T.; Kurths, J. Sleep as a novel biomarker and a promising therapeutic target for cerebral small vessel disease: A review focusing on Alzheimer’s disease and the blood-brain barrier. Int. J. Mol. Sci. 2020, 21, 6293. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semyachkina-Glushkovskaya, O.; Mamedova, A.; Vinnik, V.; Klimova, M.; Saranceva, E.; Ageev, V.; Yu, T.; Zhu, D.; Penzel, T.; Kurths, J. Brain Mechanisms of COVID-19-Sleep Disorders. Int. J. Mol. Sci. 2021, 22, 6917. https://doi.org/10.3390/ijms22136917
Semyachkina-Glushkovskaya O, Mamedova A, Vinnik V, Klimova M, Saranceva E, Ageev V, Yu T, Zhu D, Penzel T, Kurths J. Brain Mechanisms of COVID-19-Sleep Disorders. International Journal of Molecular Sciences. 2021; 22(13):6917. https://doi.org/10.3390/ijms22136917
Chicago/Turabian StyleSemyachkina-Glushkovskaya, Oxana, Aysel Mamedova, Valeria Vinnik, Maria Klimova, Elena Saranceva, Vasily Ageev, Tingting Yu, Dan Zhu, Thomas Penzel, and Jürgen Kurths. 2021. "Brain Mechanisms of COVID-19-Sleep Disorders" International Journal of Molecular Sciences 22, no. 13: 6917. https://doi.org/10.3390/ijms22136917
APA StyleSemyachkina-Glushkovskaya, O., Mamedova, A., Vinnik, V., Klimova, M., Saranceva, E., Ageev, V., Yu, T., Zhu, D., Penzel, T., & Kurths, J. (2021). Brain Mechanisms of COVID-19-Sleep Disorders. International Journal of Molecular Sciences, 22(13), 6917. https://doi.org/10.3390/ijms22136917