
Human Chromosome 18 and Acrocentrics: A Dangerous Liaison

 , , , , , , and
, , , , , , and 
Abstract
1. Introduction
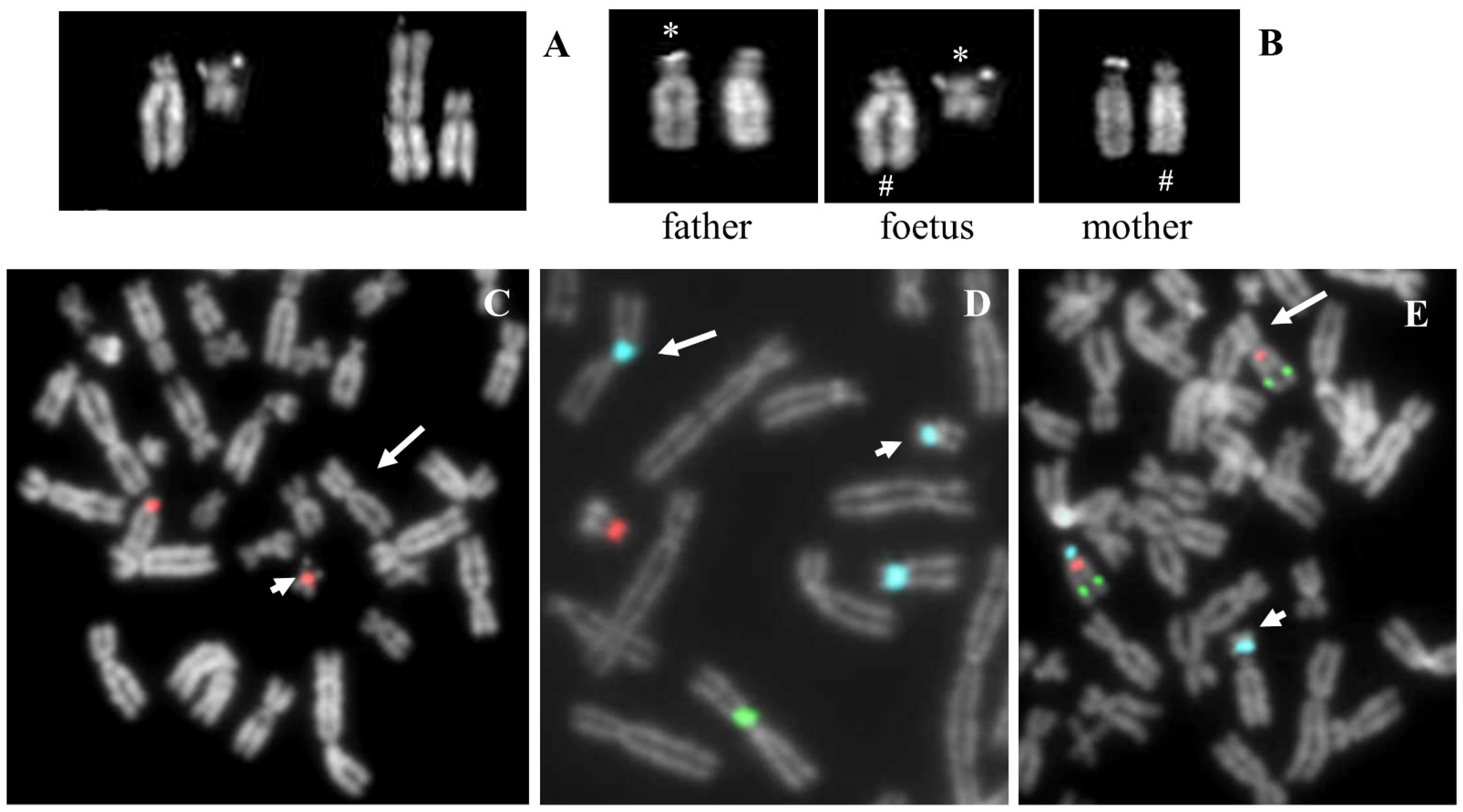
2. Case Reports
2.1. Case 1
2.1.1. Case Description
2.1.2. Results
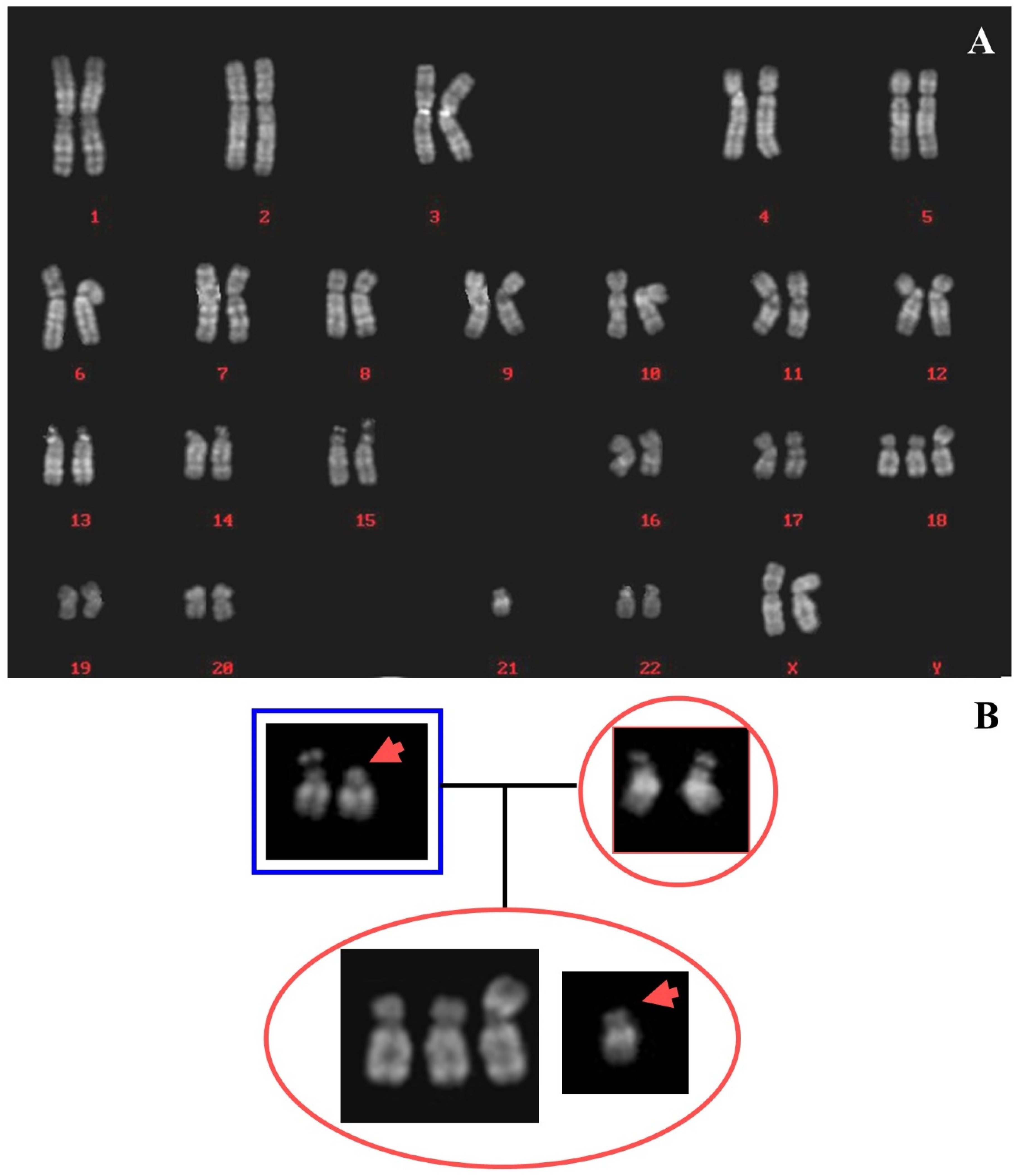
2.2. Case 2
2.2.1. Case Description
2.2.2. Results
2.3. Case 3
2.3.1. Case Description
2.3.2. Results
2.4. Case 4
2.4.1. Case Description
2.4.2. Results
2.5. Case 5
2.5.1. Case Description
2.5.2. Results
2.6. Telomeric Case (Case 6)
2.6.1. Case Description
2.6.2. Results
2.7. Chromosome 18 “Loves” Acrocentrics? What Is the Supporting Evidence?
3. Discussion
4. Materials and Methods
4.1. Chromosome Analysis
4.2. D-FISH Analysis
4.3. Array Comparative Genomic Hybridization (Array-CGH)
4.4. Microsatellite Analysis
4.5. Methylation-Specific PCR
4.6. UCSC Genome Browser Analysis
4.7. Bioinformatics Analysis
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barra, V.; Fachinetti, D. The dark side of centromeres: Types, causes and consequences of structural abnormalities implicating centromeric DNA. Nat. Commun. 2018, 9, 4340. [Google Scholar] [CrossRef]
- Wang, J.C.; Hajianpour, A.; Habibian, R. Centromeric alpha-satellite DNA break in reciprocal translocations. Cytogenet. Genome Res. 2009, 125, 329–333. [Google Scholar] [CrossRef]
- Jarmuz-Szymczak, M.; Janiszewska, J.; Szyfter, K.; Shaffer, L.G. Narrowing the localization of the region breakpoint in most frequent Robertsonian translocations. Chromosome Res. 2014, 22, 517–532. [Google Scholar] [CrossRef]
- Warburton, D. De novo balanced chromosome rearrangements and extra marker chromosomes identified at prenatal diagnosis: Clinical significance and distribution of breakpoints. Am. J. Hum. Genet. 1991, 49, 995–1013. [Google Scholar]
- Hamerton, J.L.; Canning, N.; Ray, M.; Smith, S. A cytogenetic survey of 14,069 newborn infants. I. Incidence of chromosome abnormalities. Clin. Genet. 1975, 8, 223–243. [Google Scholar] [CrossRef]
- Schinzel, A. Catalogue of Unbalanced Chromosome Aberrations in Humans, 2nd ed.; Walter de Gruyter: Berlin, Germany, 2001. [Google Scholar] [CrossRef]
- Schaub, R.L.; Reveles, X.T.; Baillargeon, J.; Leach, R.J.; Cody, J.D. Molecular characterization of 18p deletions: Evidence for a breakpoint cluster. Genet. Med. 2002, 4, 15–19. [Google Scholar] [CrossRef]
- Turleau, C. Monosomy 18p. Orphanet J. Rare Dis. 2008, 19, 1–5. [Google Scholar] [CrossRef]
- Hasi-Zogaj, M.; Sebold, C.; Heard, P.; Carter, E.; Soileau, B.; Hill, A.; Rupert, D.; Perry, B.; Atkinson, S.; O’Donnell, L.; et al. A review of 18p deletions. Am. J. Med. Genet. C Semin. Med. Genet. 2015, 169, 251–264. [Google Scholar] [CrossRef]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef] [PubMed]
- Durand, N.C.; Robinson, J.T.; Shamim, M.S.; Machol, I.; Mesirov, J.P.; Lander, E.S.; Aiede, E.L. Juicebox Provides a Visualization System for Hi-C Contact Maps with Unlimited Zoom. Cell Syst. 2016, 3, 99–101. [Google Scholar] [CrossRef]
- Ginzburg, I.A.; Lisichenko, O.V.; Kitaĭnik, G.P. Case of unbalanced translocation (18, 22) in a child with congenital mental retardation. Tsitol. Genet. 1988, 22, 52–53. (In Russian) [Google Scholar] [PubMed]
- Cooper, P.J.; Towe, C.; Crolla, J.A. A balanced whole arm reciprocal translocation resulting in three different adverse pregnancy outcomes. J. Med. Genet. 1993, 30, 417–418. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Nemana, L.; Kou, S.Y.; Habibian, R.; Hajianpour, M.J. Molecular cytogenetic characterization of 18;21 whole arm translocation associated with monosomy 18p. Am. J. Med. Genet. 1997, 71, 463–466. [Google Scholar] [CrossRef]
- McGhee, E.M.; Qu, Y.; Wohlferd, M.M.; Goldberg, J.D.; Norton, M.E.; Cotter, P.D. Prenatal diagnosis and characterization of an unbalanced whole arm translocation resulting in monosomy for 18p. Clin. Genet. 2001, 59, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Sebold, C.; Soileau, B.; Heard, P.; Carter, E.; O’Donnell, L.; Halle, D.E.; Cody, J.D. Whole arm deletions of 18p: Medical and developmental effects. Am. J. Med. Genet. A 2015, 167A, 313–323. [Google Scholar] [CrossRef] [PubMed]
- McNulty, S.M.; Sullivan, B.A. Alpha satellite DNA biology: Finding function in the recesses of the genome. Chromosome Res. 2018, 26, 115–138. [Google Scholar] [CrossRef]
- Sullivan, L.L.; Sullivan, B.A. Genomic and functional variation of human centromeres. Exp. Cell Res. 2020, 389, 111896. [Google Scholar] [CrossRef]
- Carvalho, C.M.; Lupski, J.R. Mechanisms underlying structural variant formation in genomic disorders. Nat. Rev. Genet. 2016, 17, 224–238. [Google Scholar] [CrossRef]
- Alsop, A.E.; Teschendorff, A.E.; Edwards, P.A. Distribution of breakpoints on chromosome 18 in breast, colorectal, and pancreatic carcinoma cell lines. Cancer Genet. Cytogen. 2006, 164, 97–109. [Google Scholar] [CrossRef]
- Golfier, G.; Chibon, F.; Aurias, A.; Chen, X.N.; Korenberg, J.; Rossier, J.; Potier, M.C. The 200-kb segmental duplication on human chromosome 21 originates from a pericentromeric dissemination involving human chromosomes 2, 18 and 13. Gene 2003, 312, 51–59. [Google Scholar] [CrossRef]
- Kempfer, R.; Pombo, A. Methods for mapping 3D chromosome architecture. Nat. Rev. Genet. 2020, 21, 207–226. [Google Scholar] [CrossRef] [PubMed]
- Dillinger, S.; Straub, T.; Németh, A. Nucleolus association of chromosomal domains is largely maintained in cellular senescence despite massive nuclear reorganisation. PLoS ONE 2017, 12, e0178821. [Google Scholar] [CrossRef] [PubMed]
- Cremer, M.; von Hase, J.; Volm, T.; Brero, A.; Kreth, G.; Walter, J.; Fischer, C.; Solovei, I.; Cremer, C.; Cremer, T. Non-random radial higher-order chromatin arrangements in nuclei of diploid human cells. Chromosome Res. 2001, 9, 541–567. [Google Scholar] [CrossRef] [PubMed]
- Kind, J.; Pagie, L.; de Vries, S.S.; Nahidiazar, L.; Dey, S.S.; Bienko, M.; Zhan, Y.; Lajoie, B.; de Graaf, C.A.; Amendola, M.; et al. Genome-wide maps of nuclear lamina interactions in single human cells. Cell 2015, 163, 134–147. [Google Scholar] [CrossRef]
- Associazione Italiana di Citogenetica Medica. Diagnostica Citogenetica, Consensus. Analysis 1995, 8, 12–42. [Google Scholar]
- McGowan-Jordan, J.; Hastings, R.J.; Moore, S. (Eds.) International Standing Committee on Human Cytogenomic Nomenclature. In ISCN: An International System for Human Cytogenomic Nomenclature; Karger: Basel, Switzerland, 2020. [Google Scholar]
- Lissoni, S.; Baronchelli, S.; Villa, N.; Lucchini, V.; Betri, E.; Cavalli, P.; Dalpra, L. Chromosome territories, X; Y translocation and Premature Ovarian Failure: Is there a relationship? Mol. Cytogen. 2009, 2, 19. [Google Scholar] [CrossRef]
- Redaelli, S.; Maitz, S.; Crosti, F.; Sala, E.; Villa, N.; Spaccini, L.; Selicorni, A.; Rigoldi, M.; Conconi, D.; Dalpra, L.; et al. Refining the Phenotype of Recurrent Rearrangements of Chromosome 16. Int. J. Mol. Sci 2019, 20, 1095. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reciprocal Translocations Involving 18 | Chromosome Translocations between 18 and One Acrocentric | Chr. Translocations between 18 and Non-Acrocentric | ||
|---|---|---|---|---|
| Break at the Centromere of 18 | Break Not at the Centromere of 18 | |||
| Laboratory 1 | 11 | 2 (Chr 22) 1 (Chr 15) | 1 (Chr 13) | 5 |
| 1 (Chr 14) | ||||
| 1 (Chr 22) | ||||
| Laboratory 2 | 7 | 1 (Chr 13) | 1 (Chr 13) | 4 |
| 1 (Chr 14) | ||||
| Laboratory 3 | 13 | 1 (Chr 21) | 2 (Chr 13) | 6 |
| 3 (Chr 14) | ||||
| 1 (Chr 22) | ||||
| Total | 31 | 5 (16.1%) | 11 (35.5%) | 15 (48.4%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villa, N.; Redaelli, S.; Sala, E.; Conconi, D.; Romitti, L.; Manfredini, E.; Crosti, F.; Roversi, G.; Lavitrano, M.; Rodeschini, O.; et al. Human Chromosome 18 and Acrocentrics: A Dangerous Liaison. Int. J. Mol. Sci. 2021, 22, 5637. https://doi.org/10.3390/ijms22115637
Villa N, Redaelli S, Sala E, Conconi D, Romitti L, Manfredini E, Crosti F, Roversi G, Lavitrano M, Rodeschini O, et al. Human Chromosome 18 and Acrocentrics: A Dangerous Liaison. International Journal of Molecular Sciences. 2021; 22(11):5637. https://doi.org/10.3390/ijms22115637
Chicago/Turabian StyleVilla, Nicoletta, Serena Redaelli, Elena Sala, Donatella Conconi, Lorenza Romitti, Emanuela Manfredini, Francesca Crosti, Gaia Roversi, Marialuisa Lavitrano, Ornella Rodeschini, and et al. 2021. "Human Chromosome 18 and Acrocentrics: A Dangerous Liaison" International Journal of Molecular Sciences 22, no. 11: 5637. https://doi.org/10.3390/ijms22115637
APA StyleVilla, N., Redaelli, S., Sala, E., Conconi, D., Romitti, L., Manfredini, E., Crosti, F., Roversi, G., Lavitrano, M., Rodeschini, O., Recalcati, M. P., Piazza, R., Dalprà, L., Riva, P., & Bentivegna, A. (2021). Human Chromosome 18 and Acrocentrics: A Dangerous Liaison. International Journal of Molecular Sciences, 22(11), 5637. https://doi.org/10.3390/ijms22115637