
Hsp100 Molecular Chaperone ClpB and Its Role in Virulence of Bacterial Pathogens


Abstract
1. Introduction
2. AAA+ Chaperone ClpB: Its Structure, Function and Mechanism of Action
3. The Role of ClpB in Bacterial Pathogens
4. ClpB as a Druggable Target
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vouga, M.; Greub, G. Emerging bacterial pathogens: The past and beyond. Clin. Microbiol. Infect. 2016, 22, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Mikkelson, H.; Jurgersen, G. Intracellular pathogens: Host immunity and microbial persistence strategies. J. Immunol. Res. 2019, 2019, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Gorbach, S.L. Bacterial infections: Overview. Int. Encycl. Public Health 2008, 273–282. [Google Scholar]
- Cantas, L.; Suer, K. Review: The important bacterial zoonoses in “One Health” concept. Front. Public Health 2014, 2, 144. [Google Scholar] [CrossRef]
- Tomley, F.M.; Shirley, M.W. Livestock infectious diseases and zoonoses. Philos. Trans. R. Soc. B 2009, 364, 2637–2642. [Google Scholar] [CrossRef]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. USA 2013, 10, 8399–8404. [Google Scholar] [CrossRef] [PubMed]
- Albiger, B.; Dahlberg, S.; Henriques-Normark, B.; Normark, S. Role of the innate immune system in host defence against bacterial infections: Focus on the Toll-like receptors. J. Intern. Med. 2007, 261, 511–528. [Google Scholar] [CrossRef]
- Frieri, M.; Kumar, K.; Boutin, A. Antibiotic resistance. J. Infect. Public Health 2017, 10, 369–378. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K.; Chakraborty, R.; Grossart, H.-P.; Reddy, G.S.; Jagannadham, M.V. Antibiotic resistance of bacteria. Biomed. Res. Int. 2015, 2015, 501658. [Google Scholar] [CrossRef]
- Neuwald, A.F.; Aravind, I.; Spouge, J.L.; Koonin, E.V. AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complex. Genome Res. 1999, 9, 27–43. [Google Scholar]
- Hanson, P.I.; Whiteheart, S.W. AAA+ proteins: Have engine, will work. Nat. Rev. Mol. Cell Biol. 2005, 6, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Katikaridis, P.; Meins, L.; Kamal, S.M.; Römling, U.; Mogk, A. ClpG provides increased heat resistance by acting as superior disaggregase. Biomolecules 2019, 9, 815. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Singh, L.K.; Kumari, S.; Hakiem, O.R.; Batra, J.K. ClpB is an essential regulator of Mycobacterium tuberculosis and endows survival advantage to dormant bacilli. Int. J. Med. Microbiol. 2020, 310, 151402. [Google Scholar] [CrossRef] [PubMed]
- Squires, C.L.; Pedersen, S.; Ross, B.M.; Squires, C. ClpB is the Escherichia coli heat shock protein F84.1. J. Bacteriol. 1991, 173, 4254–4262. [Google Scholar] [CrossRef]
- Zolkiewski, M. ClpB cooperates with DnaK, DnaJ, and GrpE in suppressing protein aggregation. J. Biol. Chem. 1999, 274, 28083–28086. [Google Scholar] [CrossRef]
- Goloubinoff, P.; Mogk, A.; Ben-Zvi, A.P.; Tomoyasu, T.; Bukau, B. Sequential mechanism of solubilization and refolding of stable protein aggregates by a bichaperone network. Proc. Natl. Acad. Sci. USA 1999, 96, 13732–13737. [Google Scholar] [CrossRef] [PubMed]
- Mogk, A.; Tomoyasu, T.; Goloubinoff, P.; Rűdiger, S.; Röder, D.; Langen, H.; Bukau, B. Identification of thermolabile Escherichia coli proteins: Prevention and reversion of aggregation by DnaK and ClpB. EMBO J. 1999, 18, 6934–6949. [Google Scholar] [CrossRef]
- Zolkiewski, M.; Zhang, T.; Nagy, M. Aggregate reactivation mediated by the Hsp100 chaperones. Arch. Biochem. Biophys. 2012, 520, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.M.; Hoskins, J.R.; Wicker, S. Collaboration between the ClpB AAA+ remodeling protein and the DnaK chaperone system. Proc. Natl. Acad. Sci. USA 2007, 104, 11138–11144. [Google Scholar] [CrossRef]
- Zhang, T.; Kędzierska-Mieszkowska, S.; Liu, H.; Cheng, C.; Ganta, R.R.; Zolkiewski, M. Aggregate-reactivation activity of the molecular chaperone ClpB from Ehrlichia chaffeensis. PLoS ONE 2013, 8, e62454. [Google Scholar] [CrossRef]
- Krajewska, J.; Modrak-Wójcik, A.; Arent, Z.; Więckowski, D.; Zolkiewski, M.; Bzowska, A.; Kędzierska-Mieszkowska, S. Characterization of the molecular chaperone ClpB from the pathogenic spirochaete Leptospira interrogans. PLoS ONE 2017, 12, e0181118. [Google Scholar] [CrossRef]
- Schlee, S.; Beinker, P.; Akhrymuk, A.; Reinstein, J. A chaperone network for the resolubilization of protein aggregates: Direct interaction of ClpB and DnaK. J. Mol. Biol. 2004, 336, 275–285. [Google Scholar] [CrossRef]
- Miot, M.; Reidy, M.; Doyle, S.M.; Hoskins, J.R.; Johnston, D.M.; Genest, D.; Vitery, M.C.; Massion, D.C.; Wicker, S. Species-specific collaboration of heat shock proteins (Hsp) 70 and 100 in thermotolerance and protein disaggregation. Proc. Natl. Acad. Sci. USA 2011, 108, 6915–6920. [Google Scholar] [CrossRef]
- DeSantis, M.E.; Shorter, J. The elusive middle domain of Hsp104 and ClpB: Location and function. Biochim. Biophys. Acta 2012, 1823, 29–39. [Google Scholar] [CrossRef]
- Lee, S.; Sowa, M.E.; Watanabe, Y.; Sigler, P.B.; Chiu, W.; Yoshida, M.; Tsai, F.T. The structure of ClpB: A molecular chaperone that rescues proteins from an aggregated state. Cell 2003, 115, 229–240. [Google Scholar] [CrossRef]
- Zolkiewski, M.; Kessel, M.; Ginsburg, A.; Maurizi, M.R. Nucleotide-dependent oligomerization of ClpB from Escherichia coli. Protein Sci. 1999, 8, 1899–1903. [Google Scholar] [CrossRef]
- Akoev, V.; Gogol, E.P.; Barnett, M.E.; Zolkiewski, M. Nucleotide-induced switch in oligomerization of the AAA+ ATPase ClpB. Protein Sci. 2004, 13, 567–574. [Google Scholar] [CrossRef]
- Lin, J.; Lucius, A.L. Examination of the dynamic assembly equilibrium for E. coli ClpB. Proteins 2015, 83, 2008–2024. [Google Scholar] [CrossRef]
- Kedzierska, S.; Akoev, V.; Barnett, M.E.; Zolkiewski, M.M. Structure and function of the middle domain of ClpB from Escherichia coli. Biochemistry 2003, 42, 14242–14248. [Google Scholar] [CrossRef]
- Lee, S.; Roh, S.H.; Lee, J.; Sung, N.; Liu, J.; Tsai, F.T.F. Cryo-EM structures of the Hsp104 protein disaggregase captured in the ATP conformation. Cell Rep. 2019, 26, 29–36. [Google Scholar] [CrossRef]
- Weibezahn, J.; Tessarz, P.; Schlieker, C.; Zahn, R.; Maglica, Z.; Lee, S.; Zentgraf, H.; Weber-Ban, E.U.; Dougan, D.A.; Tsai, F.T.; et al. Thermotolerance requires refolding of aggregated proteins by substrate translocation through the central pore of ClpB. Cell 2004, 119, 653–665. [Google Scholar] [CrossRef]
- Gates, S.N.; Yokom, A.L.; Lin, J.; Jackrel, M.E.; Rizo, A.N.; Kendsersky, N.M.; Buell, C.E.; Sweeny, E.A.; Mack, K.L.; Chuang, E.; et al. Rachet-like polypeptide translocation mechanism of the AAA+ disaggregase Hsp104. Science 2017, 357, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lupoli, T.J.; Kovach, A.; Meng, X.; Zhao, G.; Nathan, C.F.; Li, H. ATP hydrolysis-coupled peptide translocation mechanism of Mycobacterium tuberculosis ClpB. Proc. Natl. Acad. Sci. USA 2018, 115, E9560–E9569. [Google Scholar] [CrossRef] [PubMed]
- Rizo, A.N.; Lin, J.; Gates, S.N.; Tse, E.; Bart, S.M.; Castellano, L.M.; DiMaio, F.; Shorter, J.; Southworth, D.R. Structural basis for substrate gripping and translocation by the ClpB AAA+ disaggregase. Nat. Commun. 2019, 10, 2393. [Google Scholar] [CrossRef] [PubMed]
- Avellaneda, M.J.; Franke, K.B.; Sunderlikova, V.; Bukau, B.; Mogk, A.; Trans, S.J. Processive extrusion of polypeptide loops by a Hsp100 disaggregase. Nature 2020, 578, 317–320. [Google Scholar] [CrossRef]
- De Los Rios, P.; Ben-Zvi, A.; Slutsky, O.; Azem, A.; Goloubinoff, B. Hsp70 chaperones accelerate protein translocation and the unfolding of stable protein aggregates by entropic pulling. Proc. Natl. Acad. Sci. USA 2006, 103, 6166–6171. [Google Scholar] [CrossRef]
- Wentink, A.S.; Nillegoda, N.B.; Feufel, J.; Ubartaitė, G.; Schneider, C.P.; De Los Rios, P.; Hennig, J.; Barducci, A.; Bukau, B. Molecular dissection of amyloid disaggregation by human Hsp70. Nature 2020, 587, 483–488. [Google Scholar] [CrossRef]
- Erives, A.J.; Fassler, J.S. Metabolic and chaperone gene loss marks the origin of animals: Evidence for hsp104 and Hsp78 chaperones sharing mitochondrial enzymes as clients. PLoS ONE 2015, 10, e0117192. [Google Scholar] [CrossRef]
- Capestany, C.A.; Tribble, G.D.; Maeda, K.; Demuth, D.R.; Lamont, R.J. Role of the ClpB system in stress tolerance, biofilm formation, and intracellular invasion in Porphyromonas gingivalis. J. Bacteriol. 2008, 190, 1436–1446. [Google Scholar] [CrossRef]
- Frees, D.; Chastanet, A.; Qazi, S.; Serensen, K.; Hill, P.; Msadek, T.; Ingmer, H. Clp ATPases are required for stress tolerance, intracellular replication and biofilm formation in Staphylococcus aureus. Mol. Microbiol. 2004, 54, 1445–1462. [Google Scholar] [CrossRef]
- van Steenbergen, T.J.M.; Petit, M.D.A.; Scholte, L.H.M.; van der Velden, U.; de Graff, J. Transmission of Porphyromonas gingivalis between spouses. J. Clin. Periodontol. 1993, 20, 340–345. [Google Scholar] [CrossRef]
- Adhikari, K.; Saimbi, C.S.; Gupta, B.P. Estimation of Porphyromonas gingivalis from mother to child through saliva. J. Nepal. Med. Assoc. 2018, 56, 781–786. [Google Scholar] [CrossRef]
- Kędzierska-Mieszkowska, S.; Arent, Z. AAA+ molecular chaperone ClpB in Leptospira interrogans: Its role and significance in leptospiral virulence and pathogenesis of leptospirosis. Int. J. Mol. Sci. 2020, 21, 6645. [Google Scholar] [CrossRef]
- Chastanet, A.; Derre, I.; Nair, S.; Msadek, T. clpB, a novel number of the Listeria monocytogenes CtsR regulon, is involved in virulence but not in general stress tolerance. J. Bacteriol. 2004, 186, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Higuita, N.I.A.; Huycke, M.M. Enterococcal disease, epidemiology, and implications for treatment. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shanker, N., Eds.; Massachusetss Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Nigrovic, L.E.; Wingerter, S.L. Tularemia. Infect. Dis. Clin. N. Am. 2008, 22, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, S.W.M.; Orsel, K.; Wagenaar, J.A.; Miko, A.; van Duijkeren, E. Animal-to human transmission of Salmonella typhimurium DT104A variant. Emerg. Infect. Dis. 2004, 10, 2225–2227. [Google Scholar] [CrossRef]
- Meibom, K.L.; Dubail, I.; Dupuis, M.; Barel, M.; Lenco, J.; Stulik, J.; Golovliov, I.; Sjöstedt, A.; Charbit, A. The heat shock protein ClpB of Francisella tularensis is involved in stress tolerance and is required for multiplication in target organs of infected mice. Mol. Microbiol. 2008, 67, 1384–1401. [Google Scholar] [CrossRef]
- Alam, A.; Golovliov, I.; Javed, E.; Sjöstedt, A. ClpB mutants of Francisella tularensis subspecies holartica and tularensis are defective for type VI secretion and intracellular replication. Sci. Rep. 2018, 8, 11324. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Rodriques, P.H.; Belanger, M.; Dunn, W., Jr.; Progulske-Fox, A. The Porphyromonas gingivalis clpB gene is involved in cellular invasion In Vitro and virulence In Vivo. FEMS Immunol. Med. Microbiol. 2007, 51, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.K.; Lovell, M.A.; Hulme, S.D.; Zhang-Barber, L.; Barrow, P. Identification of Salmonella typhimurium genes required for colonization of the chickem alimentary tract and for virulence in newly hatched chicks. Intect. Immun. 1998, 66, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Vaubourgeix, J.; Lin, G.; Dhar, N.; Chenouard, N.; Jiang, X.; Botella, H.; Lupoli, T.; Mariani, O.; Yang, G.; Ouerfelli, O.; et al. Stressed mycobacteria use the chaperone ClpB to sequester irreversibly oxidized proteins asymmetrically within and between cells. Cell Host Microbe 2015, 17, 178–190. [Google Scholar] [CrossRef]
- de Oliveira, N.E.M.; Abranches, J.; Gaca, A.O.; Laport, M.S.; Damaso, C.R.; Bastos, M.C.; Lemos, J.A.; Giambiagi-deMarval, M. clpB, a class III heat-shock gene regulated by CtsR, is involved in thermotolerane and virulence of Enterococcus faecalis. Microbiology 2011, 157, 656–665. [Google Scholar] [CrossRef]
- Lourdault, K.; Cerqueira, G.M.; Wunder, E.A., Jr.; Picardeau, M. Inactivation of clpB in the pathogen Leptospira interrogans reduces virulence and resistance to stress conditions. Infect. Immun. 2011, 79, 3711–3717. [Google Scholar] [CrossRef]
- Krajewska, J.; Arent, Z.; Zolkiewski, M.; Kędzierska-Mieszkowska, S. Isolation and identification of putative protein substrates of the AAA+ molecular chaperone ClpB from the pathogenic spirochaete Leptospira interrogans. Int. J. Mol. Sci. 2018, 19, 1234. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Lata, K.S.; Sharma, P.; Bhairappanavar, S.B.; Soni, S.; Das, J. Inferring pathogen-host interactions between Leptospira interrogans and Homo sapiens using network theory. Sci. Rep. 2019, 9, 1434. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.S.; Breiman, A.; Allain, S.; Deknuydt, F.; Altare, F. The tuberculosis granuloma: A unsuccessful host defence mechanism providing a safety shelter for the bacteria? J. Immunol. Res. 2012, 2012, 139127. [Google Scholar] [CrossRef]
- Pu, Y.; Li, Y.; Jin, X.; Leake, M.C.; Lo, C.-J.; Bai, F. ATP-dependent dynamic protein aggregation regulates bacterial dormancy depth critical for antibiotic tolerance. Mol. Cell 2019, 73, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Kannan, T.R.; Musatovova, O.; Gowda, P.; Baseman, J.B. Characterization of a unique ClpB protein of Mycoplasma pneumoniae and its impact on growth. Infect. Immun. 2008, 76, 5082–5092. [Google Scholar] [CrossRef]
- Krajewska, J.; Arent, Z.; Więckowski, D.; Zolkiewski, M.; Kędzierska-Mieszkowska, S. Immunoreactivity of the AAA+ chaperone ClpB from Leptospira interrogans with sera from Leptospira-infected animals. BMC Microbiol. 2016, 16, 151–158. [Google Scholar] [CrossRef]
- Barrigan, L.M.; Tuladhar, S.; Brunton, J.C.; Woolard, M.D.; Chen, C.; Saini, D.; Frothingham, R.; Sempowski, G.D.; Kawula, T.H.; Frelinger, J.A. Infection with Francisella tularensis LVS clpB leads to an altered yet protective immune response. Infect. Immun. 2013, 81, 2028–2042. [Google Scholar] [CrossRef] [PubMed]
- Kuczyńska-Wisnik, D.; Cheng, C.; Ganta, R.R.; Zolkiewski, M. Protein aggregation in Ehrlichia chaffeensis during infection of mammalian Wells. FEMS Microbiol. Lett. 2017, 364, 1–5. [Google Scholar] [CrossRef]
- Dong, S.; Hu, W.; Me, Y.; Ojcius, D.M.; Lin, X.; Yan, J. A leptospiral AAA+ chaperone-Ntn peptidase complex, HslUV, contributes to the intracellular survival of Leptospira interrogans in hosts and the transmission of leptospirosis. Emerg. Microbes Infect. 2017, 6, e105. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.; Masison, D.C. Guanidine hydrochloride inhibits Hsp104 activity In Vivo: A possible explanation for its effect in curing yeast prions. Curr. Microbiol. 2001, 43, 7–10. [Google Scholar] [CrossRef]
- Grimminger, V.; Richter, K.; Imhof, A.; Buchner, J.; Walter, S. The prion curing agent guanidinum chloride specifically inhibits ATP hydrolysis by Hsp104. J. Biol. Chem. 2004, 279, 7378–7383. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, Ł.; Leźnicki, P.; Morawiec, E.; Litwińczuk, N.; Liberek, K. Role of a conserved aspartic acid in nucleotide binding domain 1 (NBD1) of Hsp100 chaperones in their activities. Cell Stress Chaperones 2012, 17, 361–373. [Google Scholar] [CrossRef]
- Zeymer, C.; Werbeck, N.D.; Schlichting, I.; Reinstein, J. The molecular mechanism of Hsp100 chaperone inhibition by the prion curing agent guanidinum chloride. J. Biol. Chem. 2013, 288, 7065–7076. [Google Scholar] [CrossRef]
- Martin, I.; Underhaug, J.; Celaya, G.; Moro, F.; Teigen, K.; Martinez, A.; Muga, A. Screening and evaluation of small organic molecules as ClpB inhibitors and potential antimicrobials. J. Med. Chem. 2012, 56, 7177–7189. [Google Scholar] [CrossRef]
- Baell, J.; Walters, M.A. Chemistry: Chemical con artists foil drug discovery. Nature 2014, 25, 481–483. [Google Scholar] [CrossRef]
- Torrente, M.P.; Castellano, L.M.; Shorter, J. Suramin inhibits Hsp104 ATPase and disaggregase activity. PLoS ONE 2014, 9, e110115. [Google Scholar] [CrossRef]
- Stein, A. Suramin: A novel antineoplastic agent with multiple potential mechanisms of action. Cancer Res. 1993, 53, 2239–2248. [Google Scholar]
- Morgan, H.P.; McNae, I.W.; Nowicki, M.W.; Zhong, W.; Michels, P.A.; Auld, D.S.; Fothergill-Gilmore, L.A.; Walkinshaw, M.D. The trypanocidal drug suramin and other trypan blue mimetics are inhibitors of pyruvate kinases and bind to the adenosine site. J. Biol. Chem. 2011, 286, 31232–31240. [Google Scholar] [CrossRef] [PubMed]
- Glaza, P.; Ranaweera, C.B.; Shiva, S.; Roy, A.; Geisbrecht, B.V.; Schoenen, F.J.; Zolkiewski, M. Repurposing p97 inhibitors for chemical modulation of the bacterial ClpB-DnaK bichaperone system. J. Biol. Chem. 2020, 296, 100079. [Google Scholar] [CrossRef] [PubMed]
- DeGraw, J.I.; Brown, V.H.; Colwell, W.T. Potential antileprotic agents. 3. inhibition of mycobacterial dihydrofolic reductase by 2,4-diamino-5-methyl-6-alkylquinazolines. J. Med. Chem. 1974, 17, 762–764. [Google Scholar] [CrossRef]
- Van Horn, K.S.; Burda, W.N.; Fleeman, R.; Shaw, L.N.; Manetsch, R. Antibacterial activity of a series of N2,N4-disubstituted quinazoline-2,4-diamines. J. Med. Chem. 2014, 57, 3075–3093. [Google Scholar] [CrossRef] [PubMed]
- Oguchi, Y.; Kummer, E.; Seyffer, F.; Berynskyy, M.; Anstett, B.; Zahn, R.; Wade, R.C.; Mogk, A.; Bukau, B. A tightly regulated molecular toggle controls AAA+ disaggregase. Nat. Struct. Mol. Biol. 2012, 19, 1338–1346. [Google Scholar] [CrossRef]
- Lipińska, N.; Ziętkiewicz, S.; Sobczak, A.; Jurczyk, A.; Potocki, W.; Morawiec, E.; Wawrzycka, A.; Gumowski, K.; Ślusarz, M.; Rodziewicz-Motowidło, S.; et al. Disruption of ionic interactions between the nucleotide binding domain 1 (NBD1) and middle (M) domain in Hsp00 disaggregase unleashes toxic hyperactivity and partial independence from Hsp70. J. Biol. Chem. 2013, 288, 2857–2869. [Google Scholar] [CrossRef]
- Chamera, T.; Kłosowska, A.; Janta, A.; Wyszkowski, H.; Obuchowski, I.; Gumowski, K.; Liberek, K. Selective Hsp70-dependent docking of Hsp104 to protein aggregates protects the cell from the toxicity of the disaggregase. J. Mol. Biol. 2019, 431, 2180–2196. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
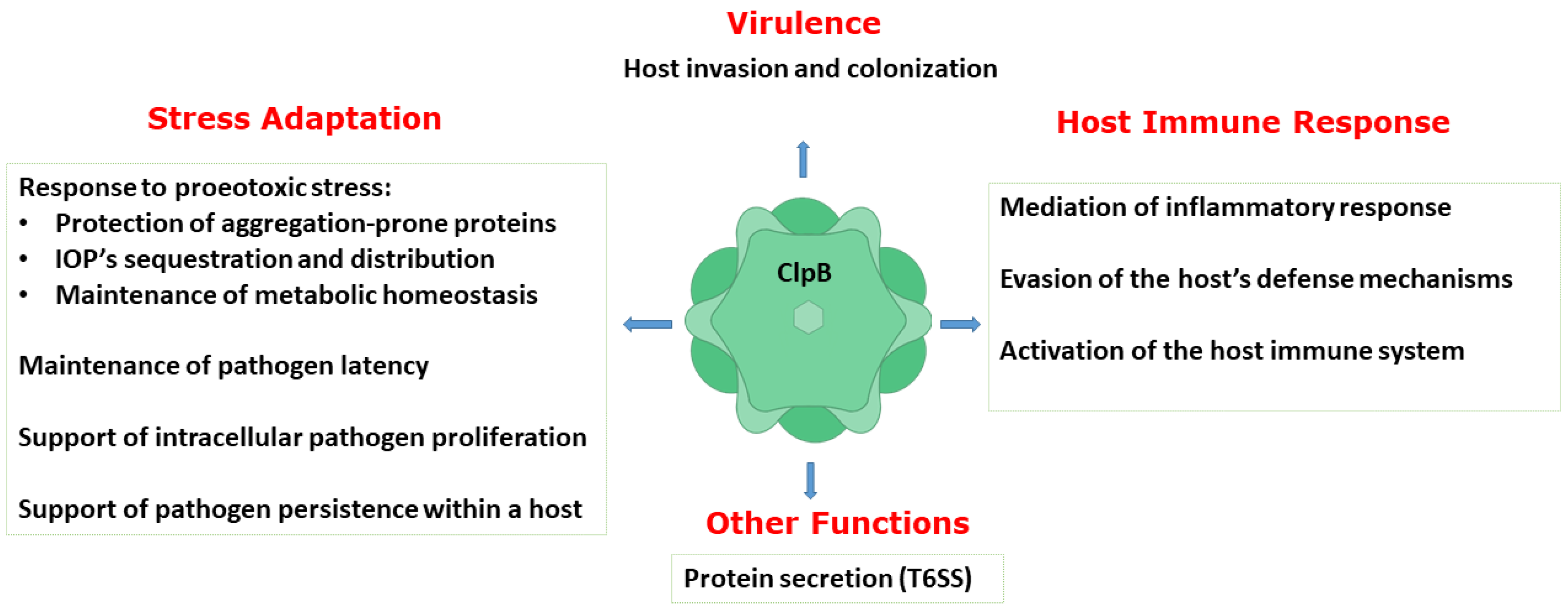{kind=link}
| Bacterial Species | Disease | Transmission |
|---|---|---|
| Ehrlichia chaffeensis * (Gram-negative Rickettsia bacterium) | human monocytic ehrlichiosis (HME) | zoonosis transmitted through an infected tick |
| Enterococcus faecalis (Gram-positive cocci) | bacteremia, endocarditis, intra-abdominal, pelvic and soft tissue infections, and urinary tract infections | transmission via a physical contact (person−person) or a contact with contaminated surfaces |
| Francisella tularensis (Gram-negative coccobacillus) | tularemia (also known as a rabbit fever) | zoonosis transmitted to humans in numerous ways, including ticks, deerfly bites, direct handling of infected tissues, ingestion of contaminated water or tissues, or inhalation of infective materials |
| Leptospira interrogans (Gram-negative spirochaete) | leptospirosis in mammals, including humans | zoonosis (wild and domestic animals are a main source of this pathogen) transmitted mainly through urine of infected animals, contact with a urine-contaminated environment, i.e., water or moist soil |
| Listeria monocytogenes (Gram-positive rod) | food-borne infections; listeriosis of pregnancy; neonatal listeriosis; clinical syndromes associated with listeriosis: meningoencephalitis, meningitis, septicemia, spontaneous abortions, stillbirth, premature labor, and neonatal disease | infection occurs through consumption of contaminated food (unpasteurized milk, soft cheeses, vegetables and some meat products), transmission is possible from mother to fetus and from mother to child during birth |
| Mycobacterium tuberculosis (neither Gram-positive nor Gram-negative bacterium) | tuberculosis | transmission through airborne particles called droplet nuclei generated by a sick person coughing, sneezing, shouting, or singing |
| Mycoplasma pneumoniae * (Gram-negative) | acute and chronic respiratory diseases; pneumonia | transmission through droplets generated by an infected person coughing or sneezing |
| Porphyromonas gingivalis (Gram-negative rod) | periodontal diseases, chronic periodontitis, systemic diseases, including heart disease, stroke, and diabetes mellitus as well as preterm delivery of low birth-weight infants | colonization of the mouth; transmission through saliva |
| Staphylococcus aureus (Gram-positive) | wide spectrum of infections from superficial wound infections, Staph food poisoning to life threatening septicemia and toxic shock syndrome; | skin and mucous membrane colonization; transmission through direct contact (person−person) |
| Salmonella enterica serovar typhimurium (Gram-negative) | food-borne illness; enteric (typhoid) fever, food poisoning in humans | zoonosis transmitted through contact with infected animals, contaminated water, and the environment; food-borne infections |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kędzierska-Mieszkowska, S.; Zolkiewski, M. Hsp100 Molecular Chaperone ClpB and Its Role in Virulence of Bacterial Pathogens. Int. J. Mol. Sci. 2021, 22, 5319. https://doi.org/10.3390/ijms22105319
Kędzierska-Mieszkowska S, Zolkiewski M. Hsp100 Molecular Chaperone ClpB and Its Role in Virulence of Bacterial Pathogens. International Journal of Molecular Sciences. 2021; 22(10):5319. https://doi.org/10.3390/ijms22105319
Chicago/Turabian StyleKędzierska-Mieszkowska, Sabina, and Michal Zolkiewski. 2021. "Hsp100 Molecular Chaperone ClpB and Its Role in Virulence of Bacterial Pathogens" International Journal of Molecular Sciences 22, no. 10: 5319. https://doi.org/10.3390/ijms22105319
APA StyleKędzierska-Mieszkowska, S., & Zolkiewski, M. (2021). Hsp100 Molecular Chaperone ClpB and Its Role in Virulence of Bacterial Pathogens. International Journal of Molecular Sciences, 22(10), 5319. https://doi.org/10.3390/ijms22105319