Characterization of Light-Enhanced Respiration in Cyanobacteria



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
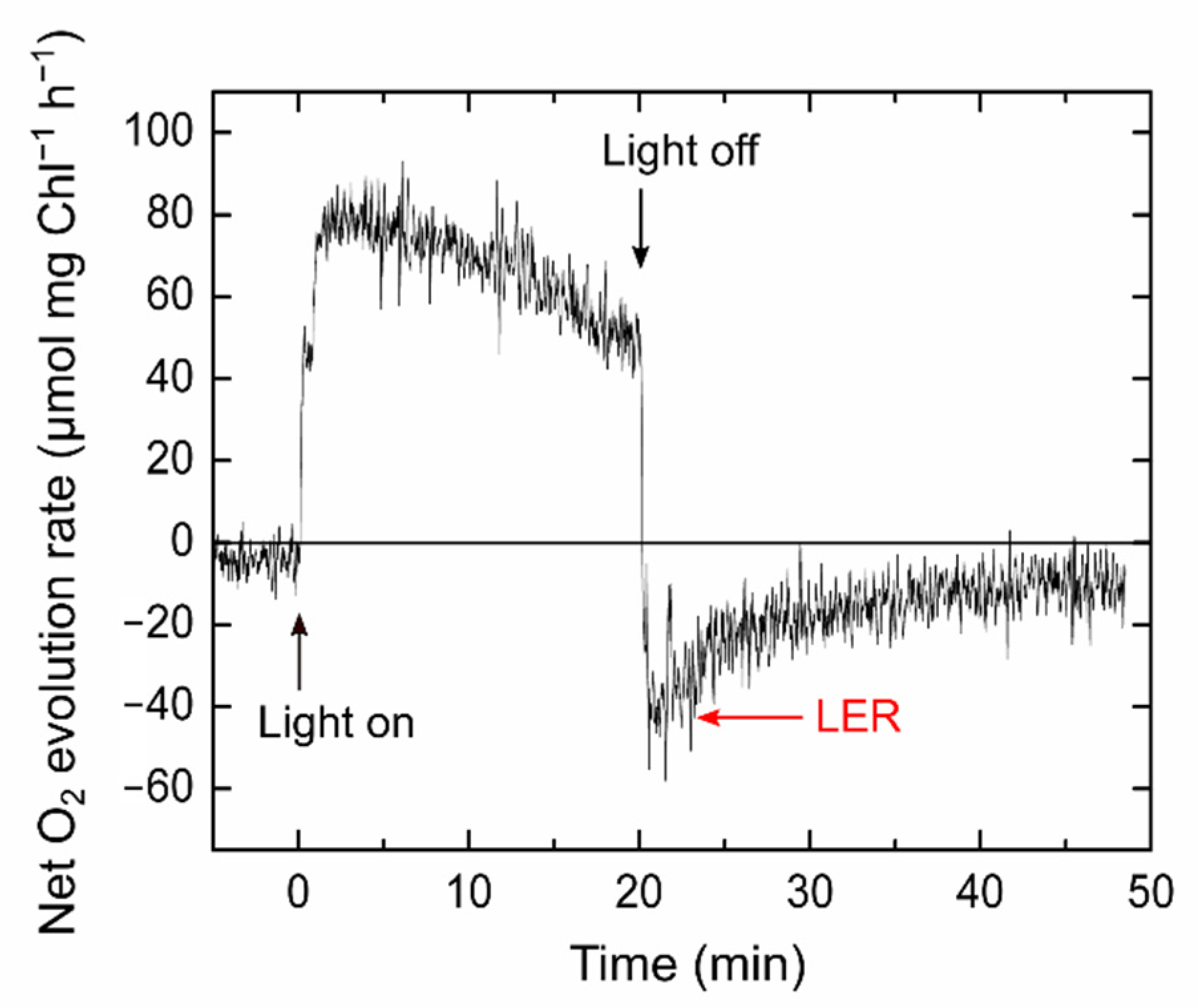
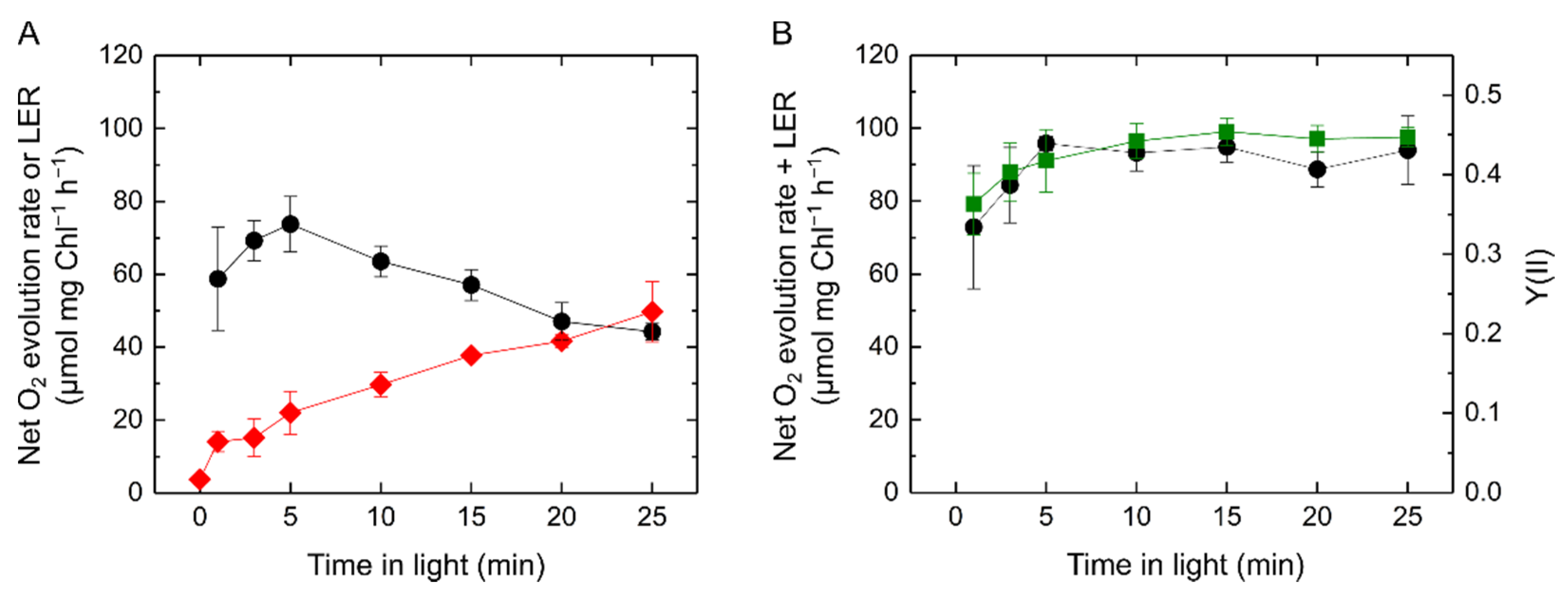
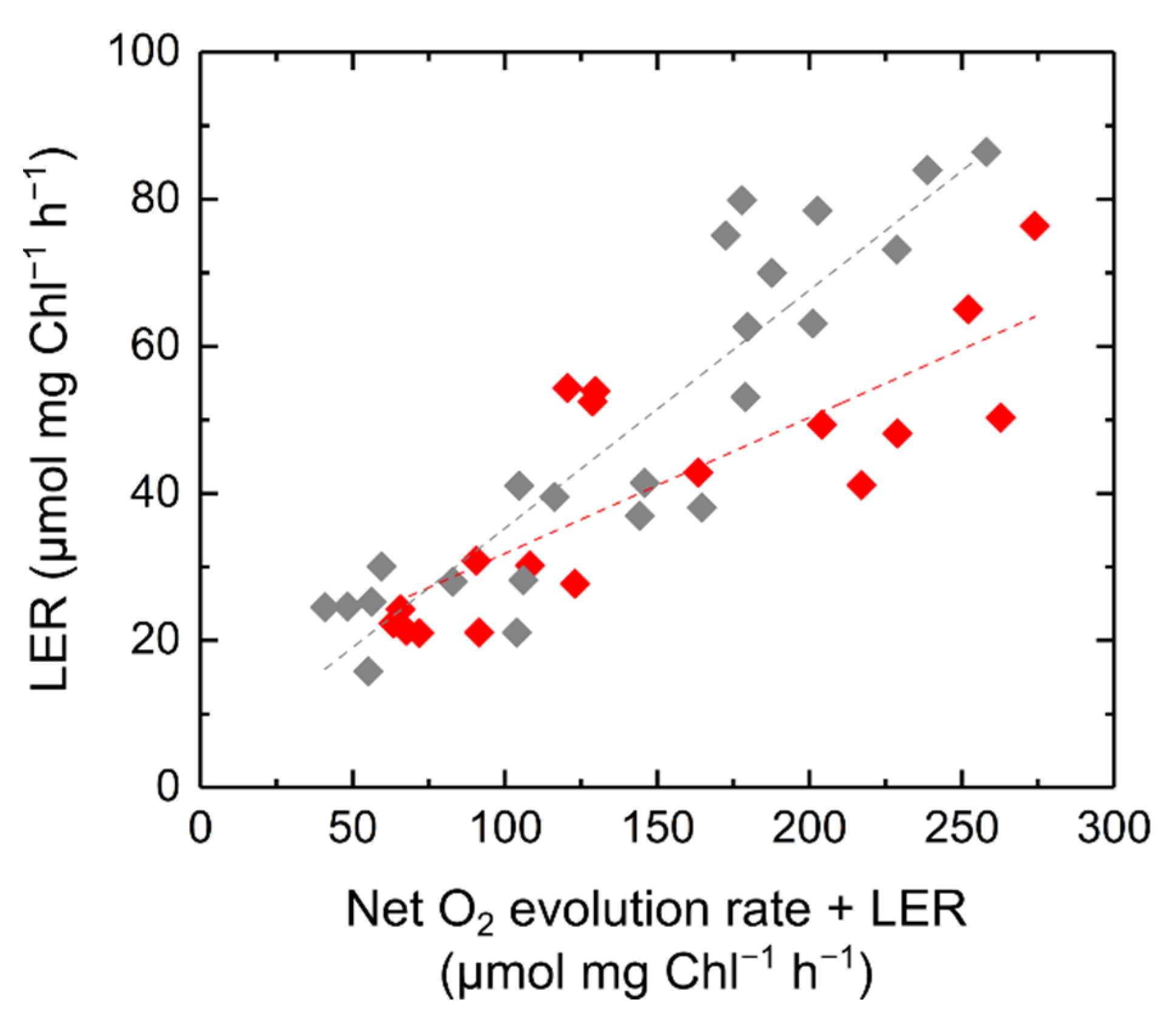
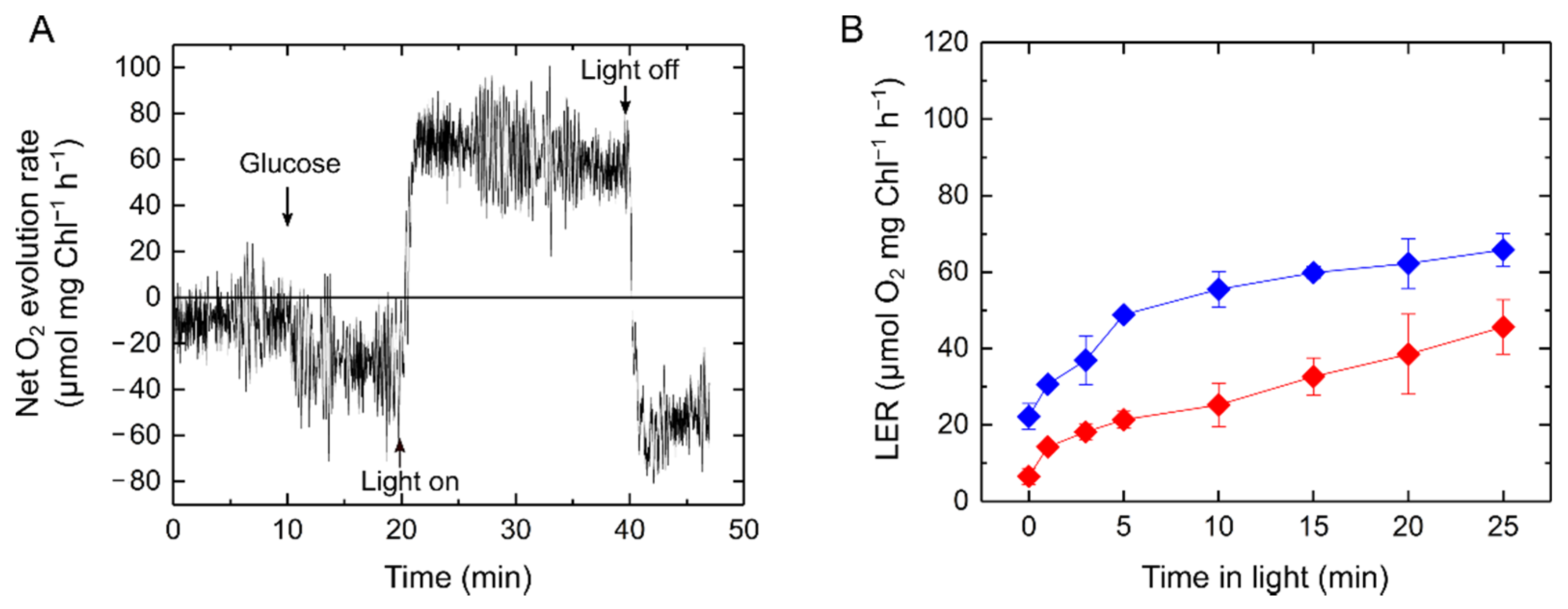
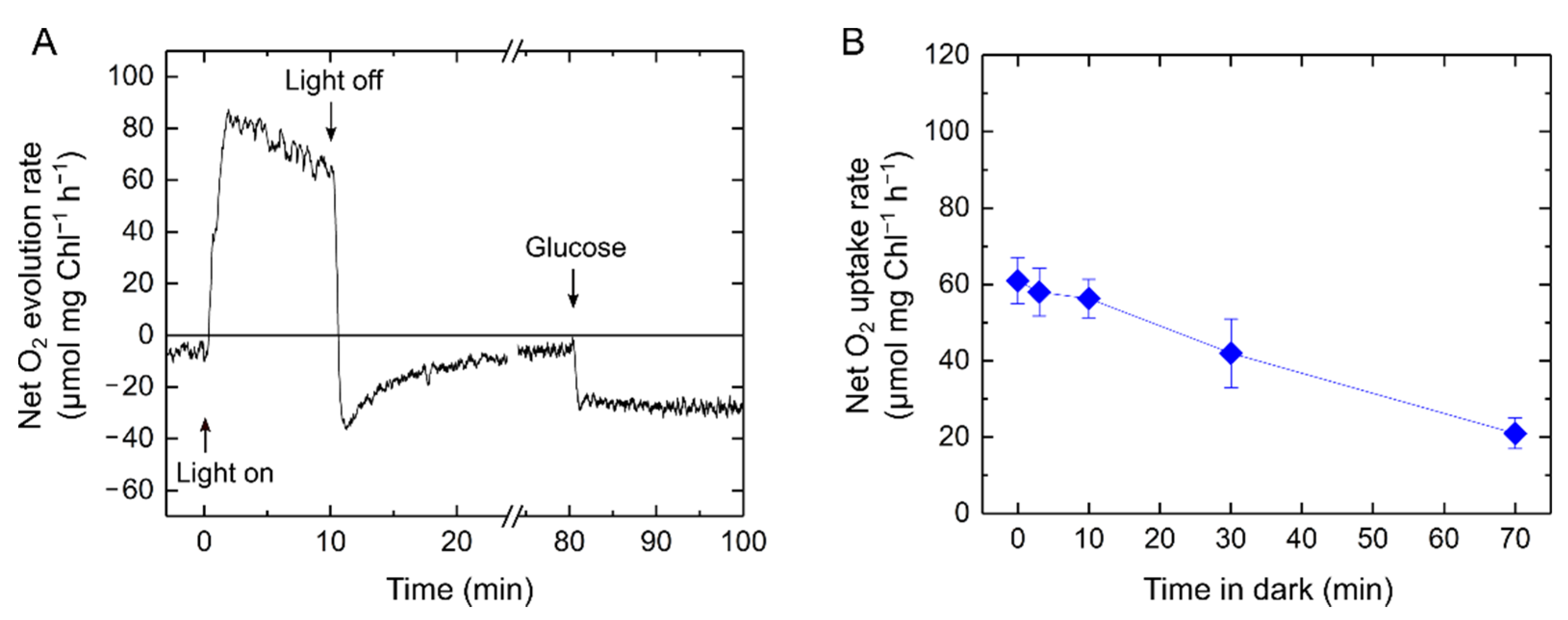
2.1. Relationship of Light-Enhanced Respiration with Photosynthesis in S. 6803
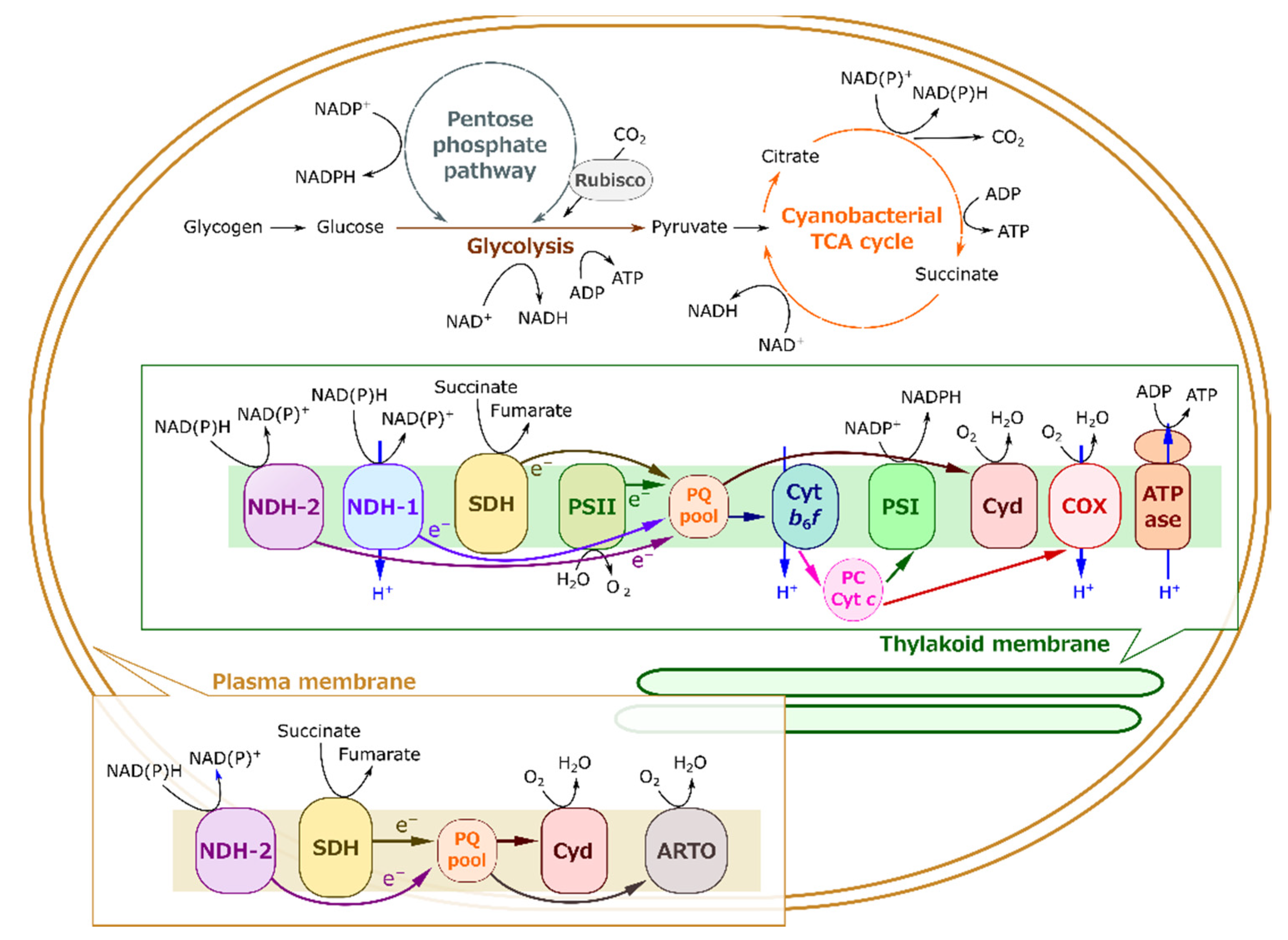
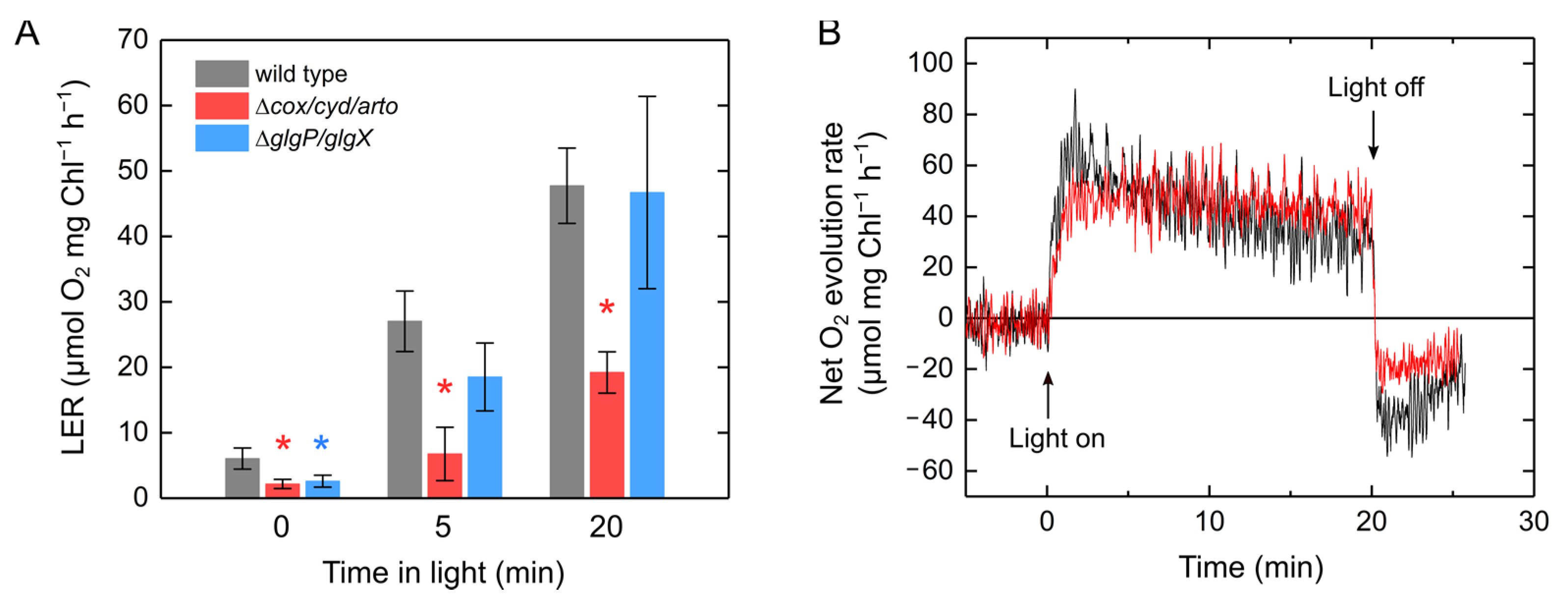
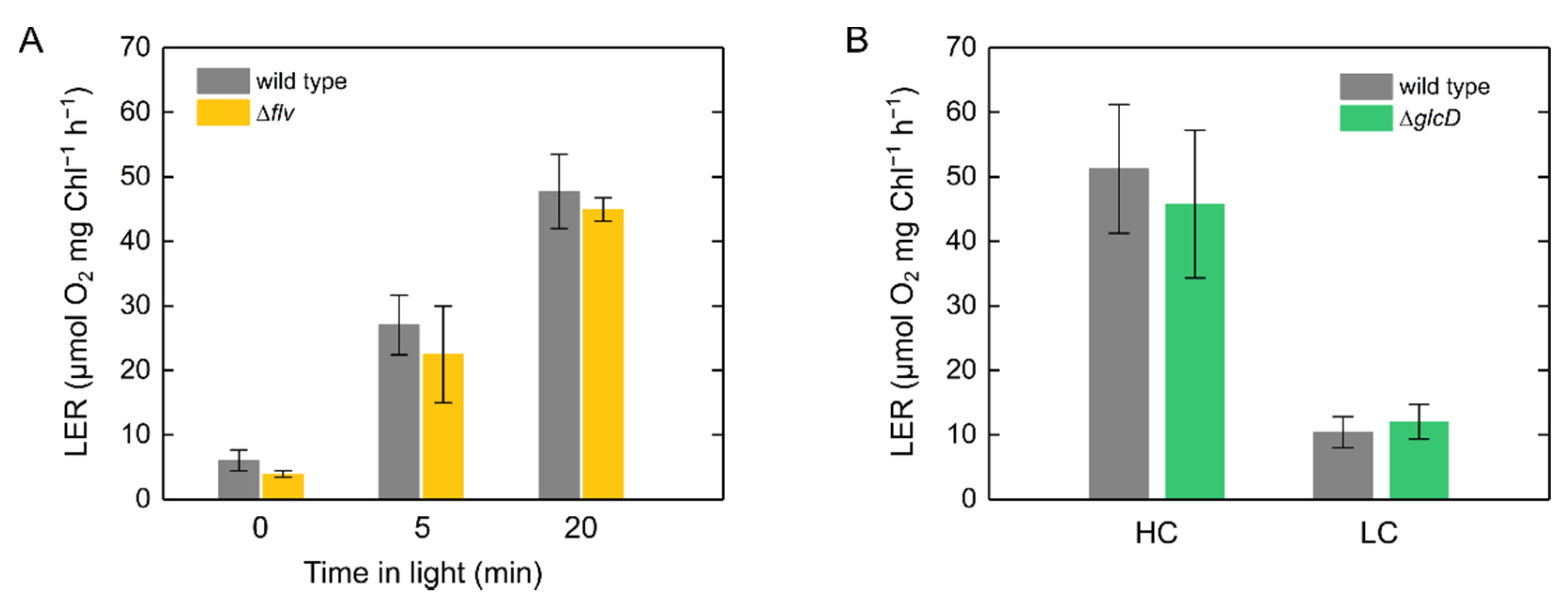
2.2. Molecular Mechanism of Light-Enhanced Respiration in S. 6803
3. Discussion
3.1. Cyanobacterial Light-Enhanced Respiration Is Metabolically Coupled with Photosynthesis
3.2. Cyanobacterial Light-Enhanced Respiration Is Not Restricted Only to Respiratory Substrate Amounts
3.3. Light-Enhanced Respiration Plays a Role in Producing ATP through Consumption of Photosynthates
3.4. Evolutionary Changes of Light-Enhanced Respiration and Photorespiration Capacities from Algae to Plants
4. Materials and Methods
4.1. Growth Conditions and Chl a Determination
4.2. Generation of Mutants
4.3. Measurement of O2 and Chl Fluorescence
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raven, J.A.; Beardall, J. Dark respiration and organic carbon loss. In The Physiology of Microalgae; Borowitzka, M.A., Beardall, J., Raven, J.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 129–140. [Google Scholar] [CrossRef]
- Møller, I.M. Plant mitochondria and oxidative stress: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [PubMed]
- Bailleul, B.; Berne, N.; Murik, O.; Petroutsos, D.; Prihoda, J.; Tanaka, A.; Villanova, V.; Bligny, R.; Flori, S.; Falconet, D.; et al. Energetic coupling between plastids and mitochondria drives CO2 assimilation in diatoms. Nature 2015, 524, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, A.S.; Padmasree, K. Beneficial interactions of mitochondrial metabolism with photosynthetic carbon assimilation. Trends Plant Sci. 2003, 8, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Sabar, M.; De Paepe, R.; de Kouchkovsky, Y. Complex I impairment, respiratory compensations, and photosynthetic decrease in nuclear and mitochondrial male sterile mutants of Nicotiana sylvestris. Plant Physiol. 2000, 124, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Massoz, S.; Larosa, V.; Horrion, B.; Matagne, R.F.; Remacle, C.; Cardol, P. Isolation of Chlamydomonas reinhardtii mutants with altered mitochondrial respiration by chlorophyll fluorescence measurement. J. Biotechnol. 2015, 215, 27–34. [Google Scholar] [CrossRef]
- Lapaille, M.; Thiry, M.; Perez, E.; González-Halphen, D.; Remacle, C.; Cardol, P. Loss of mitochondrial ATP synthase subunit beta (Atp2) alters mitochondrial and chloroplastic function and morphology in Chlamydomonas. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1533–1539. [Google Scholar] [CrossRef]
- Noguchi, K.; Yoshida, K. Interaction between photosynthesis and respiration in illuminated leaves. Mitochondrion 2008, 8, 87–99. [Google Scholar] [CrossRef]
- Larosa, V.; Meneghesso, A.; La Rocca, N.; Steinbeck, J.; Hippler, M.; Szabò, I.; Morosinotto, T. Mitochondria affect photosynthetic electron transport and photosensitivity in a green alga. Plant Physiol. 2018, 176, 2305–2314. [Google Scholar] [CrossRef]
- Tomaz, T.; Bagard, M.; Pracharoenwattana, I.; Lindén, P.; Lee, C.P.; Carroll, A.J.; Ströher, E.; Smith, S.M.; Gardeström, P.; Millar, A.H. Mitochondrial Malate Dehydrogenase Lowers Leaf Respiration and Alters Photorespiration and Plant Growth in Arabidopsis. Plant Physiol. 2010, 154, 1143–1157. [Google Scholar] [CrossRef]
- Meyer, E.H.; Tomaz, T.; Carroll, A.J.; Estavillo, G.; Delannoy, E.; Tanz, S.K.; Small, I.D.; Pogson, B.J.; Millar, A.H. Remodeled respiration in ndufs4 with low phosphorylation efficiency suppresses Arabidopsis germination and growth and alters control of metabolism at night. Plant Physiol. 2009, 151, 603–619. [Google Scholar] [CrossRef]
- Stitt, M.; Zeeman, S.C. Starch turnover: Pathways, regulation and role in growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Mantikci, M.; Hansen, J.L.S.; Markager, S. Photosynthesis enhanced dark respiration in three marine phytoplankton species. J. Exp. Mar. Biol. Ecol. 2017, 497, 188–196. [Google Scholar] [CrossRef]
- Weger, H.G.; Herzig, R.; Falkowski, P.G.; Turpin, D.H. Respiratory losses in the light in a marine diatom: Measurements by short-term mass spectrometry. Limnol. Oceanogr. 1989, 34, 1153–1161. [Google Scholar] [CrossRef]
- Ekelund, N.G.A. Interactions between photosynthesis and ‘light-enhanced dark respiration’ (LEDR) in the flagellate Euglena gracilis after irradiation with ultraviolet radiation. J. Photochem. Photobiol. B Biol. 2000, 55, 63–69. [Google Scholar] [CrossRef]
- Xue, X.; Gauthier, D.A.; Turpin, D.H.; Weger, H.G. Interactions between photosynthesis and respiration in the green alga Chlamydomonas reinhardtii (Characterization of light-enhanced dark respiration). Plant Physiol. 1996, 112, 1005–1014. [Google Scholar] [CrossRef]
- Scherer, S.; Almon, H.; Böger, P. Interaction of photosynthesis, respiration and nitrogen fixation in cyanobacteria. Photosynth. Res. 1988, 15, 95–114. [Google Scholar] [CrossRef]
- Peschek, G.A.; Obinger, C.; Paumann, M. The respiratory chain of blue-green algae (cyanobacteria). Physiol. Plant. 2004, 120, 358–369. [Google Scholar] [CrossRef]
- Binder, A. Respiration and photosynthesis in energy-transducing membranes of cyanobacteria. J. Bioenerg. Biomembr. 1982, 14, 271–286. [Google Scholar] [CrossRef]
- Mi, H.; Klughammer, C.; Schreiber, U. Light-induced dynamic changes of NADPH fluorescence in Synechocystis PCC 6803 and its ndhB-defective mutant M55. Plant Cell Physiol. 2000, 41, 1129–1135. [Google Scholar] [CrossRef]
- Shimakawa, G.; Hasunuma, T.; Kondo, A.; Matsuda, M.; Makino, A.; Miyake, C. Respiration accumulates Calvin cycle intermediates for the rapid start of photosynthesis in Synechocystis sp. PCC 6803. Biosci. Biotechnol. Biochem. 2014, 78, 1997–2007. [Google Scholar] [CrossRef]
- Lea-Smith, D.J.; Ross, N.; Zori, M.; Bendall, D.S.; Dennis, J.S.; Scott, S.A.; Smith, A.G.; Howe, C.J. Thylakoid terminal oxidases are essential for the cyanobacterium Synechocystis sp. PCC 6803 to survive rapidly changing light intensities. Plant Physiol. 2013, 162, 484–495. [Google Scholar] [CrossRef]
- Ogawa, T.; Sonoike, K. Dissection of respiration and photosynthesis in the cyanobacterium Synechocystis sp. PCC6803 by the analysis of chlorophyll fluorescence. J. Photochem. Photobiol. B Biol. 2015, 144, 61–67. [Google Scholar] [CrossRef]
- Feilke, K.; Ajlani, G.; Krieger-Liszkay, A. Overexpression of plastid terminal oxidase in Synechocystis sp. PCC 6803 alters cellular redox state. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160379. [Google Scholar] [CrossRef]
- Ermakova, M.; Huokko, T.; Richaud, P.; Bersanini, L.; Howe, C.J.; Lea-Smith, D.J.; Peltier, G.; Allahverdiyeva, Y. Distinguishing the roles of thylakoid respiratory terminal oxidases in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2016, 171, 1307–1319. [Google Scholar] [CrossRef]
- Mills, L.A.; McCormick, A.J.; Lea-Smith, D.J. Current knowledge and recent advances in understanding metabolism of the model cyanobacterium Synechocystis sp. PCC 6803. Biosci. Rep. 2020, 40, BSR20193325. [Google Scholar] [CrossRef]
- Ball, S.G.; Morell, M.K. From bacterial glycogen to starch: Understanding the biogenesis of the plant starch granule. Annu. Rev. Plant Biol. 2003, 54, 207–233. [Google Scholar] [CrossRef]
- Osanai, T.; Azuma, M.; Tanaka, K. Sugar catabolism regulated by light-and nitrogen-status in the cyanobacterium Synechocystis sp. PCC 6803. Photochem. Photobiol. Sci. 2007, 6, 508–514. [Google Scholar] [CrossRef]
- Steinhauser, D.; Fernie, A.R.; Araújo, W.L. Unusual cyanobacterial TCA cycles: Not broken just different. Trends Plant Sci. 2012, 17, 503–509. [Google Scholar] [CrossRef]
- Broddrick, J.T.; Rubin, B.E.; Welkie, D.G.; Du, N.; Mih, N.; Diamond, S.; Lee, J.J.; Golden, S.S.; Palsson, B.O. Unique attributes of cyanobacterial metabolism revealed by improved genome-scale metabolic modeling and essential gene analysis. Proc. Natl. Acad. Sci. USA 2016, 113, 8344–8353. [Google Scholar] [CrossRef]
- Welkie, D.G.; Rubin, B.E.; Diamond, S.; Hood, R.D.; Savage, D.F.; Golden, S.S. A hard day’s night: Cyanobacteria in diel cycles. Trends Microbiol. 2019, 27, 231–242. [Google Scholar] [CrossRef]
- Omata, T.; Murata, N. Cytochromes and prenylquinones in preparations of cytoplasmic and thylakoid membranes from the cyanobacterium (blue-green alga) Anacystis nidulans. Biochim. Biophys. Acta Bioenerg. 1984, 766, 395–402. [Google Scholar] [CrossRef]
- Mullineaux, C.W. Co-existence of photosynthetic and respiratory activities in cyanobacterial thylakoid membranes. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.; Schneider, D.; Vermaas, W.F.J.; Rögner, M. Electron transport routes in whole cells of Synechocystis sp. Strain PCC 6803: The role of the cytochrome bd-type oxidase. Biochemistry 2002, 41, 3422–3429. [Google Scholar] [CrossRef] [PubMed]
- Howitt, C.A.; Vermaas, W.F.J. Quinol and cytochrome oxidases in the cyanobacterium Synechocystis sp. PCC 6803. Biochemistry 1998, 37, 17944–17951. [Google Scholar] [CrossRef]
- Shimakawa, G.; Shaku, K.; Nishi, A.; Hayashi, R.; Yamamoto, H.; Sakamoto, K.; Makino, A.; Miyake, C. FLAVODIIRON2 and FLAVODIIRON4 proteins mediate an oxygen-dependent alternative electron flow in Synechocystis sp. PCC 6803 under CO2-limited conditions. Plant Physiol. 2015, 167, 472–480. [Google Scholar] [CrossRef]
- Shimakawa, G.; Shaku, K.; Miyake, C. Oxidation of P700 in photosystem I is essential for the growth of cyanobacteria. Plant Physiol. 2016, 172, 1443–1450. [Google Scholar] [CrossRef]
- Helman, Y.; Tchernov, D.; Reinhold, L.; Shibata, M.; Ogawa, T.; Schwarz, R.; Ohad, I.; Kaplan, A. Genes encoding A-type flavoproteins are essential for photoreduction of O2 in cyanobacteria. Curr. Biol. 2003, 13, 230–235. [Google Scholar] [CrossRef]
- Sétif, P.; Shimakawa, G.; Krieger-Liszkay, A.; Miyake, C. Identification of the electron donor to flavodiiron proteins in Synechocystis sp. PCC 6803 by in vivo spectroscopy. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148256. [Google Scholar] [CrossRef]
- Hanawa, H.; Ishizaki, K.; Nohira, K.; Takagi, D.; Shimakawa, G.; Sejima, T.; Shaku, K.; Makino, A.; Miyake, C. Land plants drive photorespiration as higher electron-sink: Comparative study of post-illumination transient O2-uptake rates from liverworts to angiosperms through ferns and gymnosperms. Physiol. Plant. 2017, 161, 138–149. [Google Scholar] [CrossRef]
- Sejima, T.; Hanawa, H.; Shimakawa, G.; Takagi, D.; Suzuki, Y.; Fukayama, H.; Makino, A.; Miyake, C. Post-illumination transient O2-uptake is driven by photorespiration in tobacco leaves. Physiol. Plant. 2016, 156, 227–238. [Google Scholar] [CrossRef]
- Shimakawa, G.; Miyake, C. Oxidation of P700 ensures robust photosynthesis. Front. Plant Sci. 2018, 9, 1617. [Google Scholar] [CrossRef] [PubMed]
- Shimakawa, G.; Miyake, C. Respiratory terminal oxidases alleviate photo-oxidative damage in photosystem I during repetitive short-pulse illumination in Synechocystis sp. PCC 6803. Photosynth. Res. 2018, 137, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Cooley, J.W.; Vermaas, W.F.J. Succinate dehydrogenase and other respiratory pathways in thylakoid membranes of Synechocystis sp. strain PCC 6803: Capacity comparisons and physiological function. J. Bacteriol. 2001, 183, 4251–4258. [Google Scholar] [CrossRef] [PubMed]
- Schuurmans, R.M.; van Alphen, P.; Schuurmans, J.M.; Matthijs, H.C.P.; Hellingwerf, K.J. Comparison of the photosynthetic yield of cyanobacteria and green algae: Different methods give different answers. PLoS ONE 2015, 10, e0139061. [Google Scholar] [CrossRef]
- Viola, S.; Bailleul, B.; Yu, J.; Nixon, P.; Sellés, J.; Joliot, P.; Wollman, F.-A. Probing the electric field across thylakoid membranes in cyanobacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 21900–21906. [Google Scholar] [CrossRef]
- Scherer, S.; Böger, P. Respiration of blue-green algae in the light. Arch. Microbiol. 1982, 132, 329–332. [Google Scholar] [CrossRef]
- Ito, S.; Osanai, T. Unconventional biochemical regulation of the oxidative pentose phosphate pathway in the model cyanobacterium Synechocystis sp. PCC 6803. Biochem. J. 2020, 477, 1309–1321. [Google Scholar] [CrossRef]
- Benson, P.J.; Purcell-Meyerink, D.; Hocart, C.H.; Truong, T.T.; James, G.O.; Rourke, L.; Djordjevic, M.A.; Blackburn, S.I.; Price, G.D. Factors altering pyruvate excretion in a glycogen storage mutant of the cyanobacterium, Synechococcus PCC7942. Front. Microbiol. 2016, 7, 475. [Google Scholar] [CrossRef]
- Braakman, R.; Follows, M.J.; Chisholm, S.W. Metabolic evolution and the self-organization of ecosystems. Proc. Natl. Acad. Sci. USA 2017, 114, E3091–E3100. [Google Scholar] [CrossRef]
- Eisenhut, M.; Ruth, W.; Haimovich, M.; Bauwe, H.; Kaplan, A.; Hagemann, M. The photorespiratory glycolate metabolism is essential for cyanobacteria and might have been conveyed endosymbiontically to plants. Proc. Natl. Acad. Sci. USA 2008, 105, 17199–17204. [Google Scholar] [CrossRef]
- Shimakawa, G.; Kohara, A.; Miyake, C. Medium-chain dehydrogenase/reductase and aldo-keto reductase scavenge reactive carbonyls in Synechocystis sp. PCC 6803. FEBS Lett. 2018, 592, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Grimme, L.H.; Boardman, N.K. Photochemical activities of a particle fraction P1 obtained from the green alga Chlorella fusca. Biochem. Biophys. Res. Commun. 1972, 49, 1617–1623. [Google Scholar] [CrossRef]
- Elhai, J.; Wolk, C.P. A versatile class of positive-selection vectors based on the nonviability of palindrome-containing plasmids that allows cloning into long polylinkers. Gene 1988, 68, 119–138. [Google Scholar] [CrossRef]
- Hayashi, R.; Shimakawa, G.; Shaku, K.; Shimizu, S.; Akimoto, S.; Yamamoto, H.; Amako, K.; Sugimoto, T.; Tamoi, M.; Makino, A.; et al. O2-dependent large electron flow functioned as an electron sink, replacing the steady-state electron flux in photosynthesis in the cyanobacterium Synechocystis sp. PCC 6803, but not in the cyanobacterium Synechococcus sp. PCC 7942. Biosci. Biotechnol. Biochem. 2014, 78, 384–393. [Google Scholar] [CrossRef]
- Sakurai, I.; Mizusawa, N.; Wada, H.; Sato, N. Digalactosyldiacylglycerol is required for stabilization of the oxygen-evolving complex in photosystem II. Plant. Physiol. 2007, 145, 1361–1370. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimakawa, G.; Kohara, A.; Miyake, C. Characterization of Light-Enhanced Respiration in Cyanobacteria. Int. J. Mol. Sci. 2021, 22, 342. https://doi.org/10.3390/ijms22010342
Shimakawa G, Kohara A, Miyake C. Characterization of Light-Enhanced Respiration in Cyanobacteria. International Journal of Molecular Sciences. 2021; 22(1):342. https://doi.org/10.3390/ijms22010342
Chicago/Turabian StyleShimakawa, Ginga, Ayaka Kohara, and Chikahiro Miyake. 2021. "Characterization of Light-Enhanced Respiration in Cyanobacteria" International Journal of Molecular Sciences 22, no. 1: 342. https://doi.org/10.3390/ijms22010342
APA StyleShimakawa, G., Kohara, A., & Miyake, C. (2021). Characterization of Light-Enhanced Respiration in Cyanobacteria. International Journal of Molecular Sciences, 22(1), 342. https://doi.org/10.3390/ijms22010342