Cloning, Expression, and Characterization of a Psychrophilic Glucose 6-Phosphate Dehydrogenase from Sphingomonas sp. PAMC 26621


Abstract
1. Introduction
2. Results
2.1. Gene Cloning of SpG6PD1
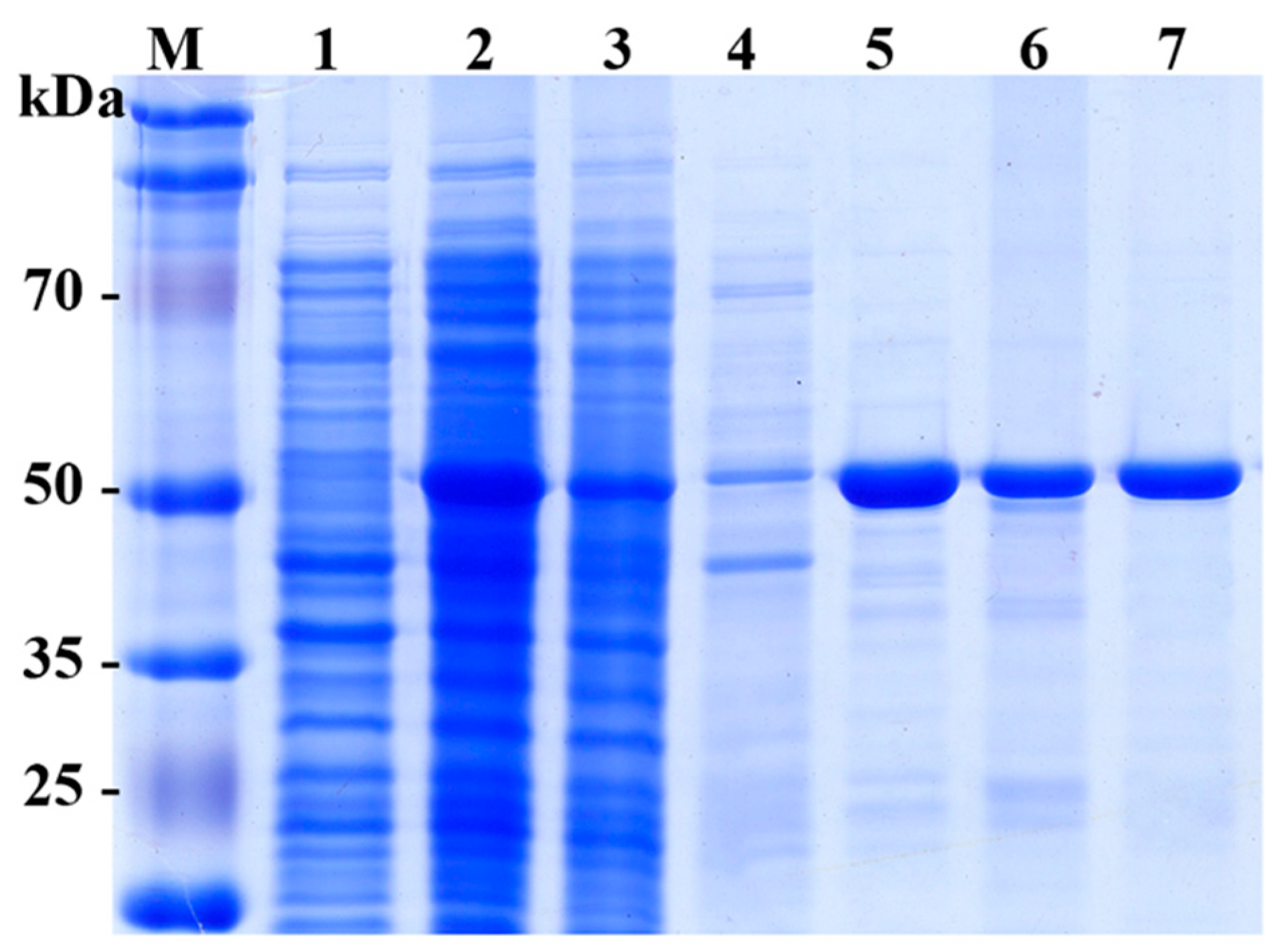
2.2. Expression and Purification of SpG6PD1
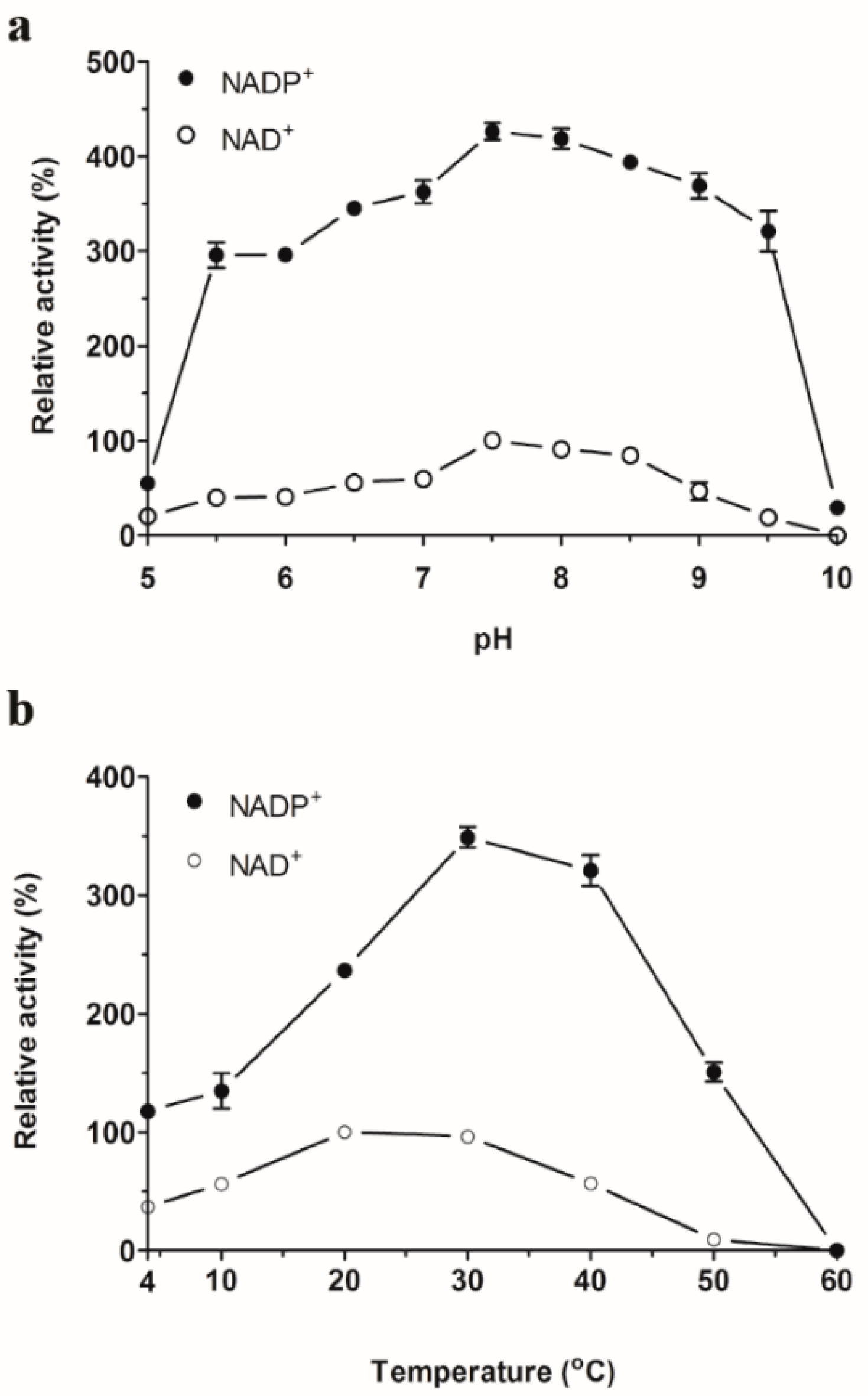
2.3. Optimum pH and Temperature
2.4. Effect of Metal Ions on the SpG6PD1 Activity
2.5. Thermal Stability
2.6. Kinetics and Thermodynamic Analysis
2.7. Spectroscopy Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Gene Cloning of SpG6PD1
4.3. Expression and Purification of SpG6PD1
4.4. Enzyme Assay
4.5. Biochemical Characterization
4.6. Enzyme Kinetics and Thermodynamic Analysis
4.7. Spectroscopy Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kirkman, H.N.; Rolfo, M.; Ferraris, A.M.; Gaetani, G.F. Mechanisms of protection of catalase by NADPH Kinetics and stoichiometry. J. Biol. Chem. 1999, 274, 13908–13914. [Google Scholar] [PubMed]
- Rosemeyer, M. The biochemistry of glucose-6-phosphate dehydrogenase, 6-phosphogluconate dehydrogenase and glutathione reductase. Cell Biochem. Funct. 1987, 5, 79–95. [Google Scholar] [PubMed]
- Mustacich, D.; Powis, G. Thioredoxin reductase. Biochem. J. 2000, 346, 1–8. [Google Scholar] [PubMed]
- Ma, J.-F.; Hager, P.W.; Howell, M.L.; Phibbs, P.V.; Hassett, D.J. Cloning and characterization of the Pseudomonas aeruginosa zwf gene encoding glucose-6-phosphate dehydrogenase, an enzyme important in resistance to methyl viologen (paraquat). J. Bacteriol. 1998, 180, 1741–1749. [Google Scholar] [PubMed]
- Lee, W.T.; Flynn, T.; Lyons, C.; Levy, H. Cloning of the gene and amino acid sequence for glucose 6-phosphate dehydrogenase from Leuconostoc mesenteroides. J. Biol. Chem. 1991, 266, 13028–13034. [Google Scholar] [PubMed]
- Li, Z.; Jiang, N.; Yang, K.; Zheng, J. Cloning, expression, and characterization of a thermostable glucose-6-phosphate dehydrogenase from Thermoanaerobacter tengcongensis. Extremophiles 2016, 20, 149–156. [Google Scholar] [CrossRef]
- Iyer, R.B.; Wang, J.; Bachas, L.G. Cloning, expression, and characterization of the gsdA gene encoding thermophilic glucose-6-phosphate dehydrogenase from Aquifex aeolicus. Extremophiles 2002, 6, 283–289. [Google Scholar]
- Acero-Navarro, K.E.; Jiménez-Ramírez, M.; Villalobos, M.A.; Vargas-Martínez, R.; Perales-Vela, H.V.; Velasco-García, R. Cloning, overexpression, and purification of glucose-6-phosphate dehydrogenase of Pseudomonas aeruginosa. Protein Expr. Purif. 2018, 142, 53–61. [Google Scholar]
- Ferrara, M.; Guerriero, G.; Cardi, M.; Esposito, S. Purification and biochemical characterisation of a glucose-6-phosphate dehydrogenase from the psychrophilic green alga Koliella antarctica. Extremophiles 2013, 17, 53–62. [Google Scholar]
- Ciardiello, M.A.; Camardella, L.; di Prisco, G. Glucose-6-phosphate dehydrogenase from the blood cells of two Antarctic teleosts: correlation with cold adaptation. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1995, 1250, 76–82. [Google Scholar] [CrossRef]
- Ciardiello, M.A.; Camardella, L.; Carratore, V.; di Prisco, G. Enzymes in Antarctic fish: glucose-6-phosphate dehydrogenase and glutamate dehydrogenase. Comp. Biochem. Physiol., Part A Mol. Integr. Physiol. 1997, 118, 1031–1036. [Google Scholar] [CrossRef]
- Honjoh, K.; Machida, T.; Hagisako, T.; Suga, K.; Yonekura, M.; Shimizu, H.; Ohashi, N.; Miyamoto, T.; Hatano, S.; Iio, M. Molecular cloning and characterization of a cDNA for low-temperature inducible cytosolic glucose 6-phosphate dehydrogenase gene from Chlorella vulgaris and expression of the gene in Saccharomyces cerevisiae. Plant Sci. 2007, 172, 649–658. [Google Scholar] [CrossRef]
- Sadakane, H.; Kabata, K.; Ishibashi, K.; Watanabe, T.; Hatano, S. Studies on frost hardiness in Chlorella ellipsoidea. V. The role of glucose and related compounds. Environ. Exp. Bot. 1980, 20, 297–305. [Google Scholar] [CrossRef]
- Lee, H.; Shin, S.C.; Lee, J.; Kim, S.J.; Kim, B.-K.; Hong, S.G.; Kim, E.H.; Park, H. Genome sequence of Sphingomonas sp. strain PAMC 26621, an Arctic-lichen-associated bacterium isolated from a Cetraria sp. J. Bacteriol. 2012, 194, 3030. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, M.S.; Gover, S.; Naylor, C.E.; Vandeputte-Rutten, L.; Adams, M.J.; Levy, H.R. An examination of the role of Asp-177 in the His-Asp catalytic dyad of Leuconostoc mesenteroides glucose 6-phosphate dehydrogenase: X-ray structure and pH dependence of kinetic parameters of the D177N mutant enzyme. Biochemistry 2000, 39, 15002–15011. [Google Scholar] [CrossRef] [PubMed]
- Levy, H.R.; Vought, V.E.; Yin, X.; Adams, M.J. Identification of an Arginine residue in the dual coenzyme-specific Glucose-6-phosphate dehydrogenase from Leuconostoc mesenteroides that plays a key role in binding NADP+ but not NAD+. Arch. Biochem. Biophys. 1996, 326, 145–151. [Google Scholar] [CrossRef]
- Rowland, P.; Basak, A.K.; Gover, S.; Levy, H.R.; Adams, M.J. The three–dimensional structure of glucose 6–phosphate dehydrogenase from Leuconostoc mesenteroides refined at 2.0 Å resolution. Structure 1994, 2, 1073–1087. [Google Scholar] [CrossRef]
- Vought, V.; Ciccone, T.; Davino, M.H.; Fairbairn, L.; Lin, Y.; Cosgrove, M.S.; Adams, M.J.; Levy, H.R. Delineation of the roles of amino acids involved in the catalytic functions of Leuconostoc mesenteroides glucose 6-phosphate dehydrogenase. Biochemistry 2000, 39, 15012–15021. [Google Scholar] [CrossRef]
- Cosgrove, M.S.; Naylor, C.; Paludan, S.; Adams, M.J.; Levy, H.R. On the mechanism of the reaction catalyzed by glucose 6-phosphate dehydrogenase. Biochemistry 1998, 37, 2759–2767. [Google Scholar] [CrossRef]
- Vartak, N.B.; Lin, C.C.; Cleary, J.M.; Fagan, M.J.; Saier Jr, M.H. Glucose metabolism in ‘Sphingomonas elodea’: Pathway engineering via construction of a glucose-6-phosphate dehydrogenase insertion mutant. Microbiology 1995, 141, 2339–2350. [Google Scholar] [CrossRef]
- Maleki, S.; Mærk, M.; Valla, S.; Ertesvåg, H. Mutational analyses of glucose dehydrogenase and glucose-6-phosphate dehydrogenase genes in Pseudomonas fluorescens reveal their effects on growth and alginate production. Appl. Environ. Microbiol. 2015, 81, 3349–3356. [Google Scholar] [CrossRef] [PubMed]
- Au, S.W.; Gover, S.; Lam, V.M.; Adams, M.J. Human glucose-6-phosphate dehydrogenase: the crystal structure reveals a structural NADP+ molecule and provides insights into enzyme deficiency. Structure 2000, 8, 293–303. [Google Scholar] [CrossRef]
- Fuentealba, M.; Muñoz, R.; Maturana, P.; Krapp, A.; Cabrera, R. Determinants of cofactor specificity for the glucose-6-phosphate dehydrogenase from Escherichia coli: simulation, kinetics and evolutionary studies. PloS one 2016, 11, e0152403. [Google Scholar] [CrossRef] [PubMed]
- Saeed, H.; Ismaeil, M.; Embaby, A.; Ataya, F.; Malik, A.; Shalaby, M.; El-Banna, S.; Ali, A.A.M.; Bassiouny, K. Overexpression, purification and enzymatic characterization of a recombinant Arabian camel Camelus dromedarius glucose-6-phosphate dehydrogenase. Protein Expr. Purif. 2018, 142, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Cardi, M.; Chibani, K.; Castiglia, D.; Cafasso, D.; Pizzo, E.; Rouhier, N.; Jacquot, J.-P.; Esposito, S. Overexpression, purification and enzymatic characterization of a recombinant plastidial glucose-6-phosphate dehydrogenase from barley (Hordeum vulgare cv. Nure) roots. Plant Physiol. Biochem. 2013, 73, 266–273. [Google Scholar] [CrossRef]
- Hansen, T.; Schlichting, B.; Schönheit, P. Glucose-6-phosphate dehydrogenase from the hyperthermophilic bacterium Thermotoga maritima: expression of the g6pd gene and characterization of an extremely thermophilic enzyme. FEMS Microbiol Lett 2002, 216, 249–253. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Ghazy, A.-H.M.; Salem, A.M.; Ghazy, M.A.; Abdel-Monsef, M.M. Purification and characterization of glucose-6-phosphate dehydrogenase from camel liver. Enzyme Res. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Santiago, M.; Ramírez-Sarmiento, C.A.; Zamora, R.A.; Parra, L.P. Discovery, Molecular Mechanisms, and Industrial Applications of Cold-Active Enzymes. Front. Microbiol. 2016, 7, 1–32. [Google Scholar] [CrossRef]
- Hu, W.; Zhi, L.; Zhuo, M.-Q.; Zhu, Q.-L.; Zheng, J.-L.; Chen, Q.-L.; Gong, Y.; Liu, C.-X. Purification and characterization of glucose 6-phosphate dehydrogenase (G6PD) from grass carp (Ctenopharyngodon idella) and inhibition effects of several metal ions on G6PD activity in vitro. Fish Physiol. Biochem. 2013, 39, 637–647. [Google Scholar] [CrossRef]
- Comakli, V.; Akkemik, E.; Ciftci, M.; Kufrevioglu, O.I. Purification and characterization of glucose 6-phosphate dehydrogenase enzyme from rainbow trout (Oncorhynchus mykiss) liver and investigation of the effects of some metal ions on enzyme activity. Toxicol. Ind. Health 2015, 31, 403–411. [Google Scholar] [CrossRef]
- Honjoh, K.-i.; Mimura, A.; Kuroiwa, E.; Hagisako, T.; Suga, K.; Shimizu, H.; Dubey, R.S.; Miyamoto, T.; Hatano, S.; Iio, M. Purification and characterization of two isoforms of glucose 6-phosphate dehydrogenase (G6PDH) from Chlorella vulgaris C-27. Biosci. Biotechnol. Biochem. 2003, 67, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Holzwarth, G.; Doty, P. The Ultraviolet Circular Dichroism of Polypeptides1. J. Am. Chem. Soc. 1965, 87, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Cappellini, M.D.; Fiorelli, G. Glucose-6-phosphate dehydrogenase deficiency. The Lancet 2008, 371, 64–74. [Google Scholar] [CrossRef]
- Feller, G.; Gerday, C. Psychrophilic enzymes: molecular basis of cold adaptation. Cell. Mol. Life Sci. 1997, 53, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Lonhienne, T.; Gerday, C.; Feller, G. Psychrophilic enzymes: revisiting the thermodynamic parameters of activation may explain local flexibility. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2000, 1543, 1–10. [Google Scholar] [CrossRef]
- Struvay, C.; Feller, G. Optimization to low temperature activity in psychrophilic enzymes. Int J Mol Sci 2012, 13, 11643–11665. [Google Scholar]
- Haghighi, B.; Aghatabar, A.; Shahsavari, G. Glucose 6-phosphate dehydrogenase from Streptomyces aureofaciens: Ligand-induced conformational chang. Iran J Sci Technol. 2005, 29, 71–78. [Google Scholar]
- STEINBACH, R.A.; SAHM, H.; SCHÜTTE, H. Purification and Regulation of Glucose-6-Phosphate Dehydrogenase from Obligate Methanol-Utilizing Bacterium Methylomonas M15. Eur. J. Biochem. 1978, 87, 409–415. [Google Scholar]
- Kurlandsky, S.B.; Hilburger, A.C.; Levy, H.R. Glucose-6-phosphate dehydrogenase from Leuconostoc mesenteroides: ligand-induced conformational changes. Arch. Biochem. Biophys. 1988, 264, 93–102. [Google Scholar]
- Miki, T.; Tsujimoto, Y.; MIYASE, S.; Sugiyama, K.-I.; Izawa, S.; Inoue, Y.; Kimura, A. Oxidative stress response in yeast: purification and some properties of oxidative stress-inducible glucose-6-phosphate dehydrogenase from Hansenula mrakii. Biosci. Biotechnol. Biochem. 1996, 60, 966–970. [Google Scholar] [CrossRef]
- Sandoval, J.M.; Arenas, F.A.; Vasquez, C.C. Glucose-6-phosphate dehydrogenase protects Escherichia coli from tellurite-mediated oxidative stress. PLoS One 2011, 6, e25573. [Google Scholar]
- D’Amico, S.; Collins, T.; Marx, J.C.; Feller, G.; Gerday, C. Psychrophilic microorganisms: challenges for life. EMBO reports 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Smalås, A.O.; Leiros, H.-K.S.; Os, V.; Willassen, N.P. Cold adapted enzymes. Biotechnol. Annu. Rev. 2000, 6, 1–57. [Google Scholar]
- Siddiqui, K.S.; Cavicchioli, R. Cold-adapted enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Boonyuen, U.; Chamchoy, K.; Swangsri, T.; Junkree, T.; Day, N.P.; White, N.J.; Imwong, M. A trade-off between catalytic activity and protein stability determines the clinical manifestations of glucose-6-phosphate dehydrogenase (G6PD) deficiency. Int. J. Biol. Macromol. 2017, 104, 145–156. [Google Scholar]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: a better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: homology modeling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total Protein (mg) | Total Activity (units) | Specific Activity (units/mg) | Yield (%) | Purification (fold) |
|---|---|---|---|---|---|
| Cell extract | 36.4 | 11284 | 310 | 100 | 1.0 |
| HisTrap | 4.2 | 7282.9 | 1717 | 65 | 5.5 |
| Q-Sepharose | 2.4 | 6382.8 | 2700 | 57 | 8.7 |
| Metal Ion | Relative Activity (%) | |
|---|---|---|
| 1 mM | 5 mM | |
| None | 100 | 100 |
| EDTA | 100.8 | 100.8 |
| Ca2+ | 101.2 | 104.5 |
| Mg2+ | 107.3 | 108.0 |
| Ni2+ | 28.6 | ND |
| Zn2+ | 33.0 | 7.0 |
| Fe3+ | 52.6 | 4.8 |
| Mn2+ | 99.0 | 98.8 |
| Cu2+ | 45.8 | 35.2 |
| NADP+-Linked Reaction | ||||||
|---|---|---|---|---|---|---|
| Temp. (°C) | Km (µM) | kcat (min−1) | kcat/Km (min−1·µM−1) | ΔG‡ (kJ·mol−1) | ΔH‡ (kJ·mol−1) | TΔS‡ (kJ·mol−1) |
| 20 | 130 ± 1.5 | 7018 ± 72 | 54.0 | 60.1 | 27.8 | −32.3 |
| 30 | 145 ± 2 | 8081 ± 48 | 55.7 | 61.9 | 27.8 | −34.1 |
| NAD+-Linked Reaction | ||||||
| Temp. (°C) | Km (µM) | kcat (min−1) | kcat/Km (min-1·µM−1) | ΔG‡ (kJ·mol−1) | ΔH‡ (kJ·mol−1) | TΔS‡ (kJ·mol−1) |
| 20 | 675 ± 9 | 1196 ± 18 | 1.80 | 64.4 | 54.1 | −10.3 |
| 30 | 4308 ± 155 | 2782 ± 288 | 0.65 | 64.6 | 54.0 | −10.6 |
| Source | Temp. (°C) | ΔG‡ (kJ·mol−1) | ΔH‡ (kJ·mol−1) | TΔS‡ (kJ·mol−1) | Reference | |
|---|---|---|---|---|---|---|
| Sphingomonas sp. | Psychrophile | 10 | 59.5 | 27.9 | −31.6 | This study |
| Dissostichus mawsoni | Cold-adapted | 0 | 56.1 | 36.8 | −19.4 | [10] |
| Chionodraco hamatus | Cold-adapted | 0 | 56.5 | 40.6 | -15.9 | [10] |
| Human | Mesophile | 0 | 58.2 | 51.9 | -6.3 | [10] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
TranNgoc, K.; Pham, N.; Lee, C.; Jang, S.-H. Cloning, Expression, and Characterization of a Psychrophilic Glucose 6-Phosphate Dehydrogenase from Sphingomonas sp. PAMC 26621. Int. J. Mol. Sci. 2019, 20, 1362. https://doi.org/10.3390/ijms20061362
TranNgoc K, Pham N, Lee C, Jang S-H. Cloning, Expression, and Characterization of a Psychrophilic Glucose 6-Phosphate Dehydrogenase from Sphingomonas sp. PAMC 26621. International Journal of Molecular Sciences. 2019; 20(6):1362. https://doi.org/10.3390/ijms20061362
Chicago/Turabian StyleTranNgoc, Kiet, Nhung Pham, ChangWoo Lee, and Sei-Heon Jang. 2019. "Cloning, Expression, and Characterization of a Psychrophilic Glucose 6-Phosphate Dehydrogenase from Sphingomonas sp. PAMC 26621" International Journal of Molecular Sciences 20, no. 6: 1362. https://doi.org/10.3390/ijms20061362
APA StyleTranNgoc, K., Pham, N., Lee, C., & Jang, S.-H. (2019). Cloning, Expression, and Characterization of a Psychrophilic Glucose 6-Phosphate Dehydrogenase from Sphingomonas sp. PAMC 26621. International Journal of Molecular Sciences, 20(6), 1362. https://doi.org/10.3390/ijms20061362