Molecular Responses of Maize Shoot to a Plant Derived Smoke Solution

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
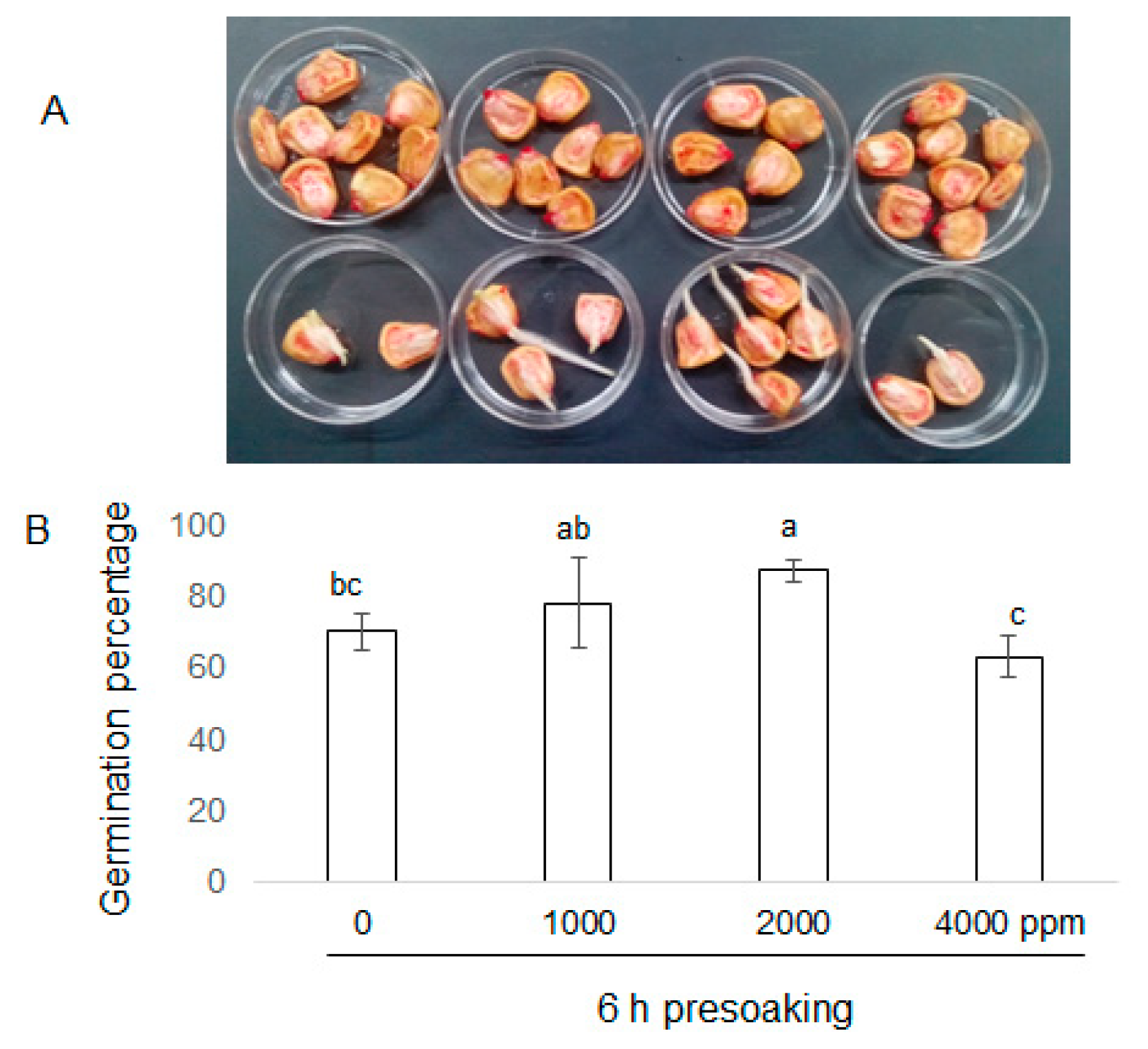
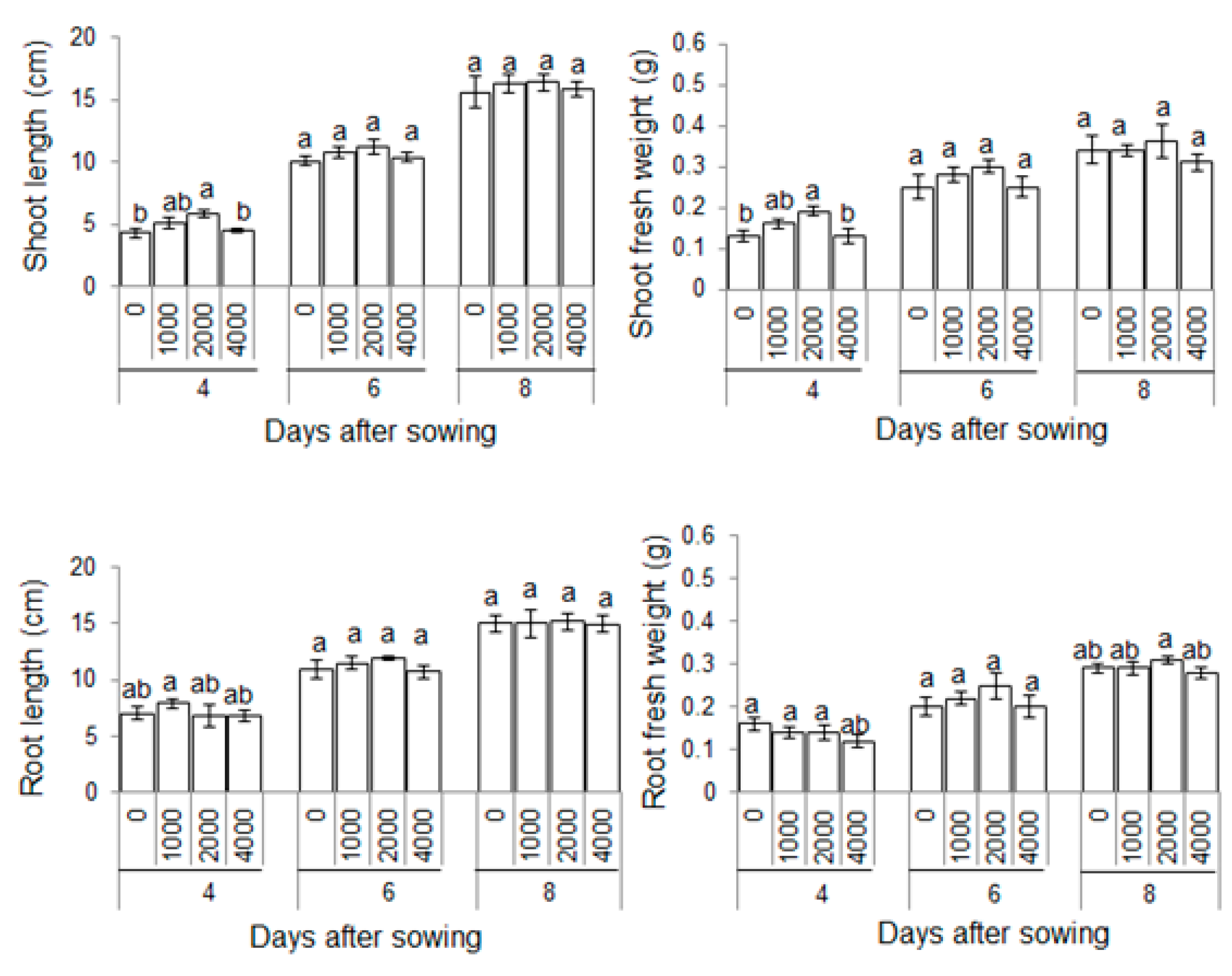
2.1. Morphological Effects of Plant-Derived Smoke on Maize Growth
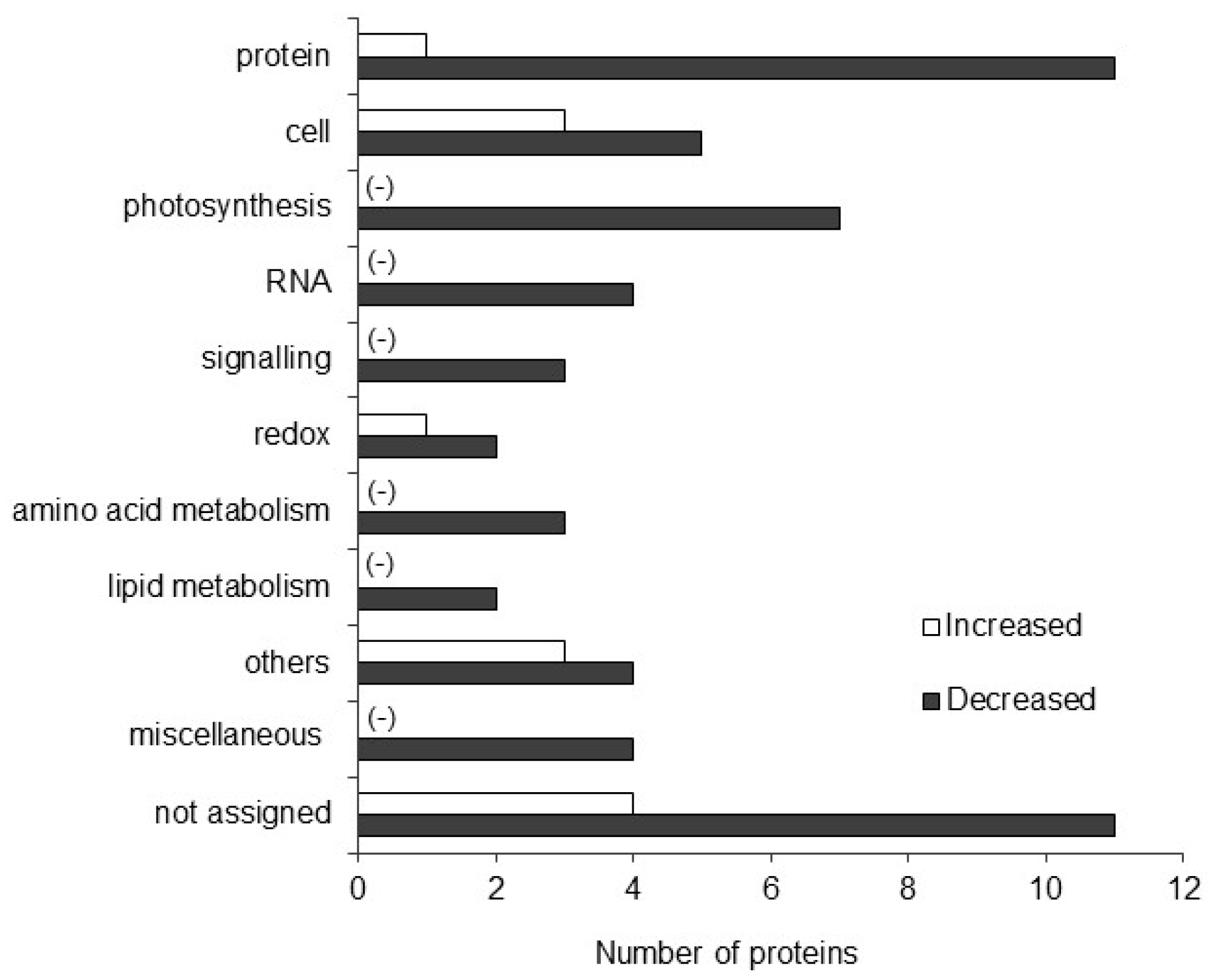
2.2. Functional Classification of Differentially Abundant Proteins in Maize Treated with Plant-Derived Smoke
2.3. Pathway Analysis of Identified Proteins in Maize Treated with Plant-Derived Smoke
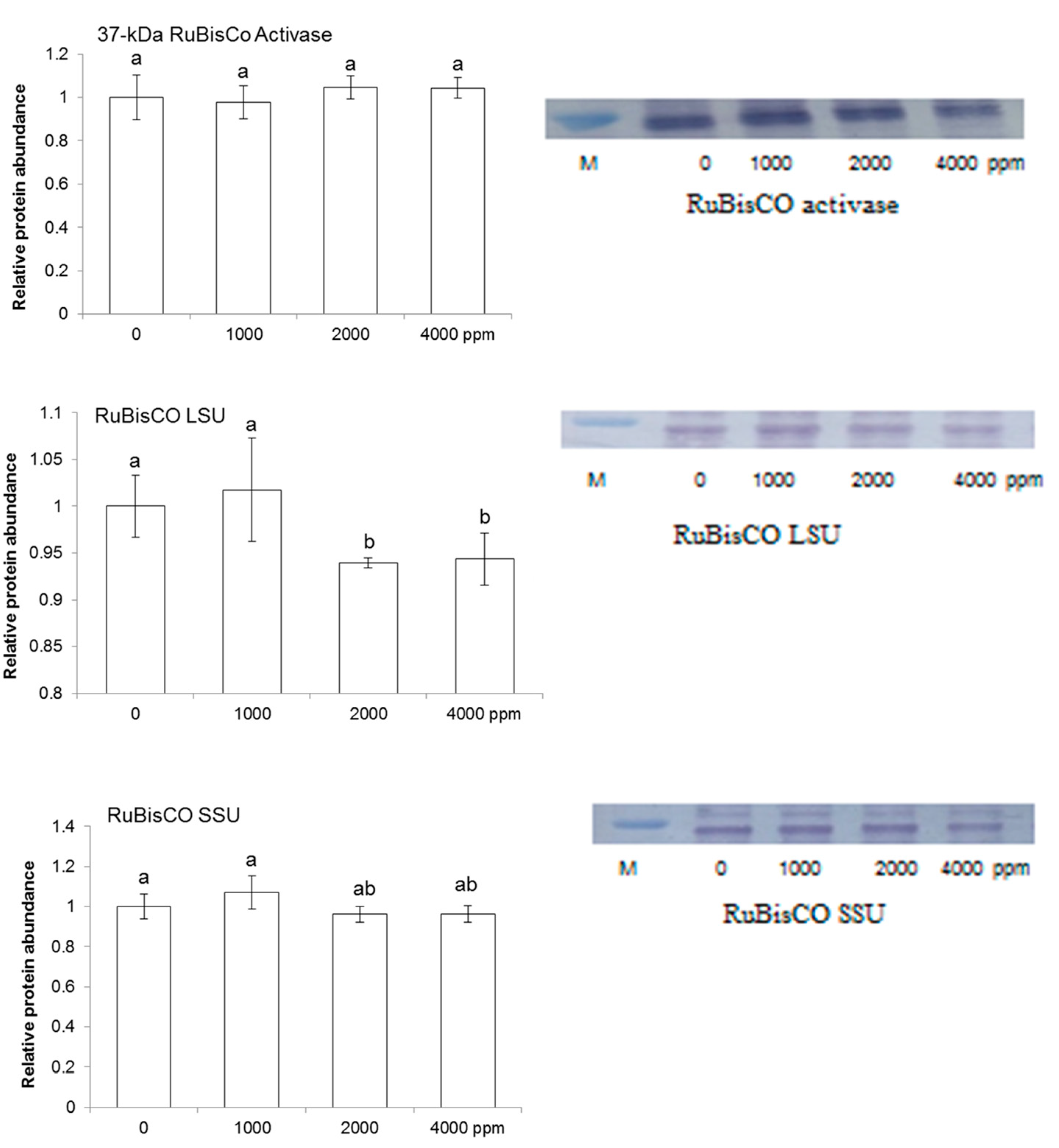
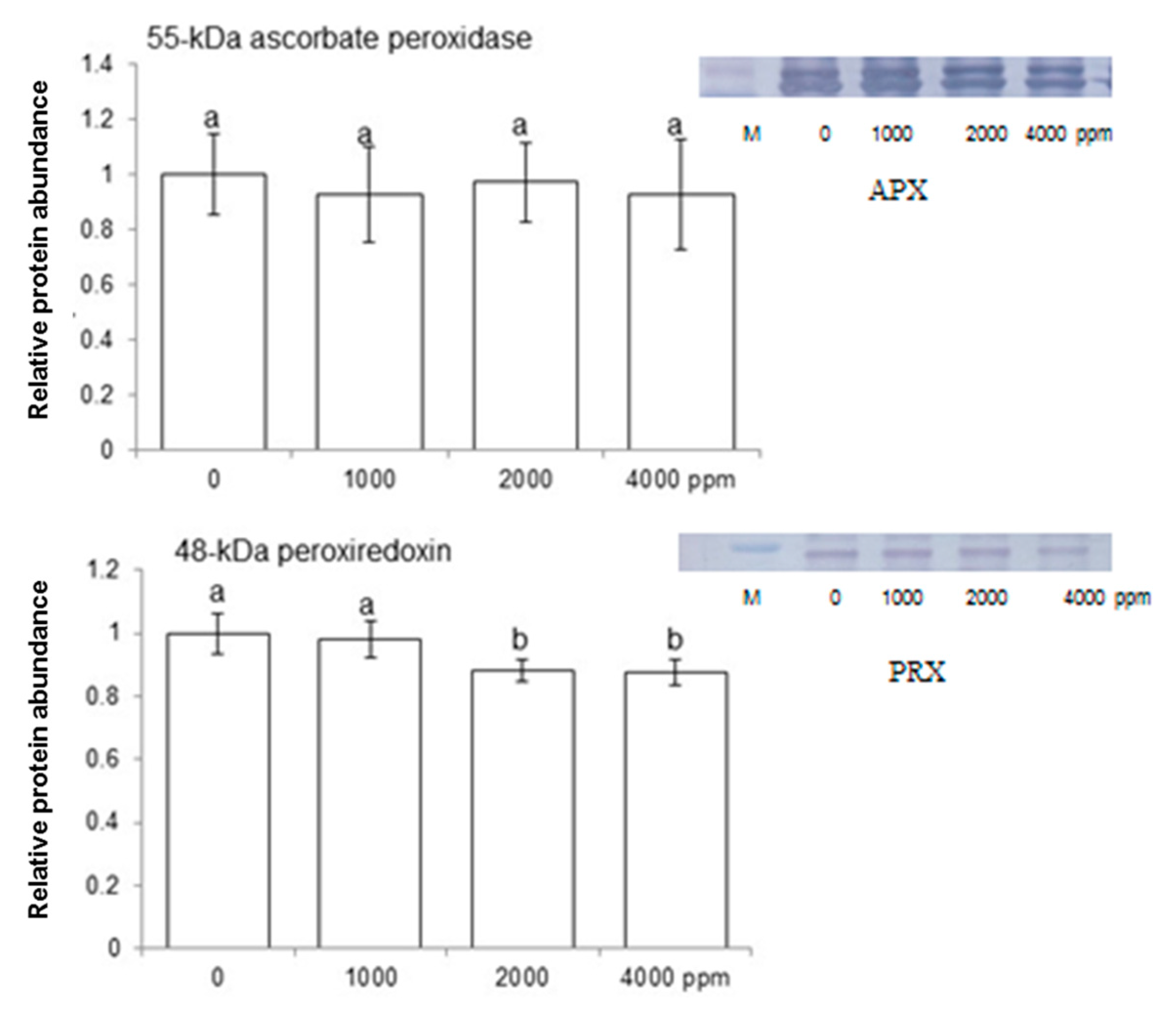
2.4. Immuno-Blot Analysis of Proteins Involved in Redox Homeostasis and Photosynthesis
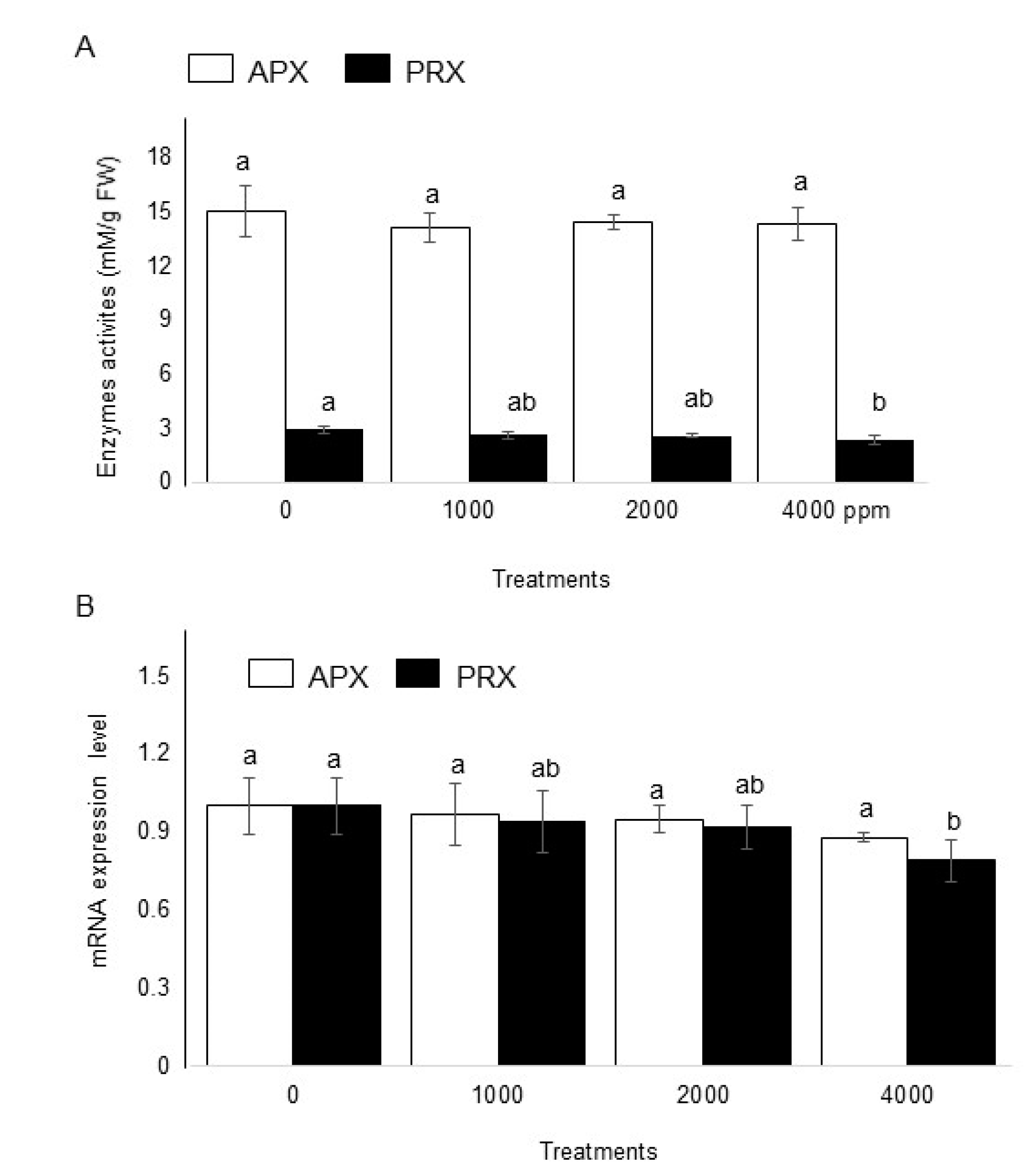
2.5. Effect of Plant-Derived Smoke on Enzymatic Activities and Gene Expression of Maize
3. Discussion
3.1. Effect of Plant-Derived Smoke on Morphology of Maize
3.2. Positive Effect of Plant-Derived Smoke on Cytoskeleton Related Proteins in Maize
3.3. Effect of Plant-Derived Smoke on Photosynthetic Proteins in Maize
3.4. Effect of Plant-Derived Smoke on Scavenging Activity through the Ascorbate/Glutathione Pathway in Maize
3.5. Effect of Plant-Derived Smoke on Enzymatic Activities of Maize
3.6. Effect of Plant-Derived Smoke on Expression of Ascorbate Peroxidase and Peroxiredoxin in Maize
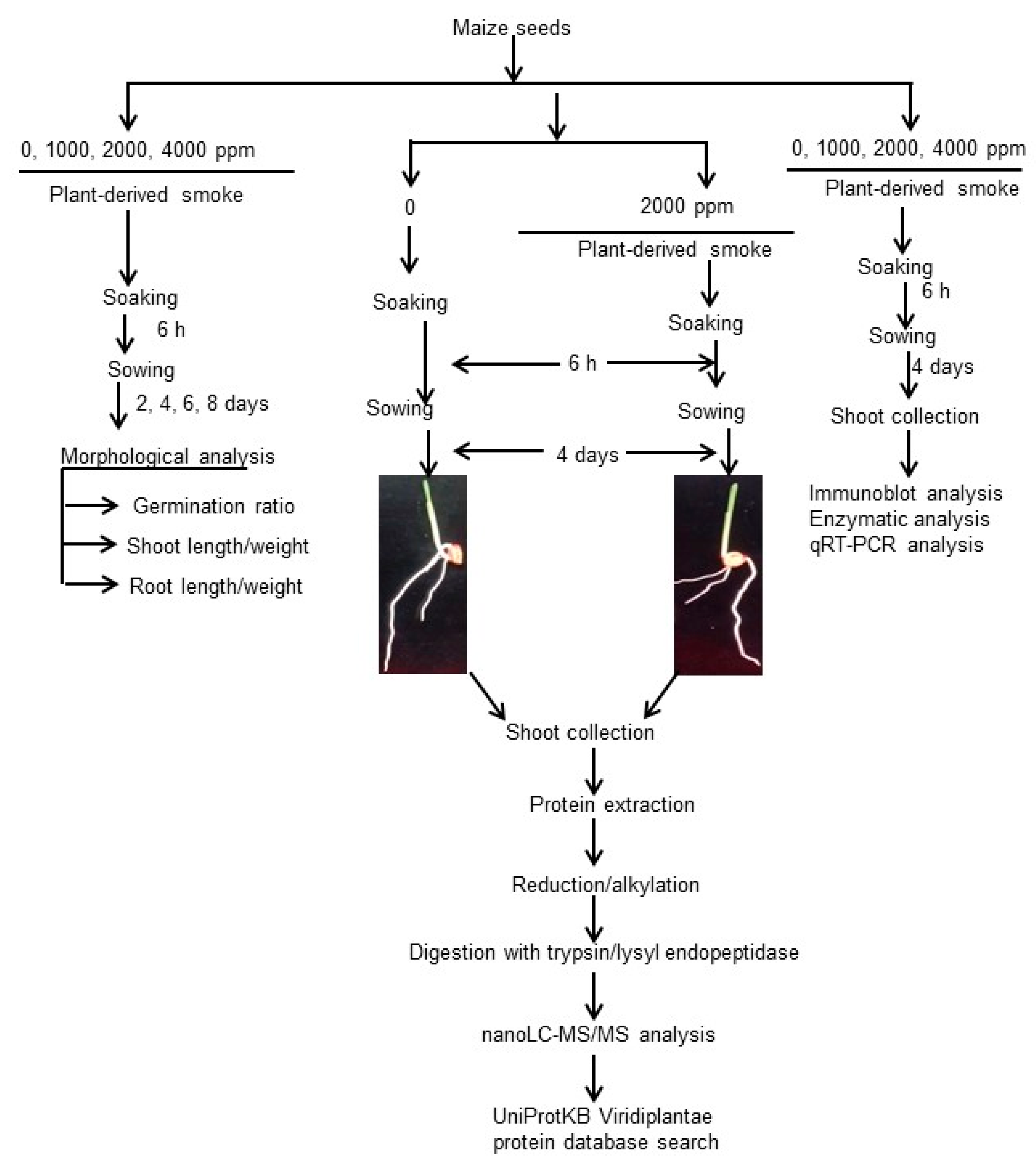
4. Materials and Methods
4.1. Preparation of Plant-Derived Smoke Solution
4.2. Plant Material and Treatment
4.3. Protein Extraction
4.4. Protein Enrichment, Reduction, Alkylation, and Digestion
4.5. Measurement of Protein and Peptide Concentrations
4.6. Protein Identification Using NanoLC-MS/MS
4.7. MS Data Analysis
4.8. Differential Analysis of Proteins Using MS Data
4.9. Functional Categorization
4.10. Immunoblotting Analysis
4.11. Enzymatic Analysis
4.12. RNA Extraction and Reverse Transcription Polymerase Chain Reaction Analysis
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| FDR | False discovery rate |
| LC | Liquid Chromatography |
| RuBisCO | Ribulose-1,5-bisphosphate carboxylase/oxygenase |
| MS | Mass Spectrometry |
References
- Lawrence, C.J.; Walbot, V. Translational genomics for bioenergy production from fuel stock grasses: Maize as the model species. Plant Cell 2007, 19, 2091–2094. [Google Scholar] [CrossRef]
- Morari, F.; Meggio, F.; Lunardon, A.; Scudiero, E.; Forestan, C.; Farinati, S.; Varotto, S. Time course of biochemical, physiological, and molecular responses to field-mimicked conditions of drought, salinity, and recovery in two maize lines. Front. Plant Sci. 2015, 6, 314. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, H.; Wu, L.; Warburton, M.; Yan, J. Genome-wide association studies in maize: Praise and stargaze. Mol. Plant 2017, 10, 359–374. [Google Scholar] [CrossRef]
- Chia, J.M.; Song, C.; Bradbury, P.J.; Costich, D.; Leon, N.; Doebley, J.; Elshire, R.J.; Gaut, B.; Geller, L.; Glaubitz, J.C.; et al. Maize HapMap2 identifies extant variation from a genome in flux. Nat. Genet. 2012, 44, 803–807. [Google Scholar] [CrossRef]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 Maize Genome: Complexity, Diversity, and Dynamics. Science 2009, 326, 1112–1114. [Google Scholar] [CrossRef] [PubMed]
- Hake, S.; Ross-Ibarra, J. The Natural History of Model Organisms: Genetic, evolutionary and plant breeding insights from the domestication of maize. eLife 2015, 4, e05861. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, S.D.; Dixon, K.W.; Hopper, S.D.; Lambers, H.; Turner, S.R. Little evidence for fire-adapted plant traits in Mediterranean climate regions. Trends Plant Sci. 2011, 16, 69–76. [Google Scholar] [CrossRef]
- Nelson, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Regulation of seed germination and seedling growth by chemical signals from burning vegetation. Annu. Rev. Plant Biol. 2012, 63, 107–130. [Google Scholar] [CrossRef] [PubMed]
- Flematti, G.R.; Waters, M.T.; Scaffidi, A.; Merritt, D.M.; Ghisalberti, E.L.; Kingsley, W.; Dixon, K.W.; Smith, S.M. Karrikin and cyanohydrin smoke signals provide clues to new endogenous plant signaling compounds. Mol. Plant 2013, 6, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.G.; Light, M.E.; van Staden, J. Plant derived smoke: Old technology with possibilities for economic applications in agriculture and horticulture. S. Afr. J. Bot. 2011, 77, 972–979. [Google Scholar] [CrossRef]
- Dixon, K.W.; Roche, S.; Pate, J.S. The promotive effect of smoke derived from burnt vegetation on seed germination of western Australian plants. Oecologia 1995, 101, 185–192. [Google Scholar] [CrossRef] [PubMed]
- van Staden, J.; Sparg, S.G.; Kulkarni, M.G.; Light, M.E. Post germination effects of the smoke-derived compound 3-methyl-2H-furo[2,3-c]pyran-2-one, and its potential as a preconditioning agent. Field Crop Res. 2006, 98, 98–105. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Street, R.A.; van Staden, J. Germination and seedling growth requirements for propagation of Dioscorea dregeana (Kunth) Dur. and Schinz-A tuberous medicinal plant. S. Afr. J. Bot. 2007, 73, 131–137. [Google Scholar] [CrossRef]
- Mavi, K.; Light, M.E.; Demir, I.; van Staden, J.; Yasar, F. Positive effect of smoke-derived butenolide priming on melon seedling emergence and growth. New Zeal. J. Crop Hort. 2010, 38, 147–155. [Google Scholar] [CrossRef]
- Moreira, B.; Tormo, J.; Estrelles, E.; Pausas, J.G. Disentangling the role of heat and smoke as germination cues in Mediterranean Basin flora. Ann. Bot. 2010, 105, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Papenfus, H.B.; Kulkarni, M.G.; Posta, M.; van Staden, J. Effect of smoke derivatives on in vitro pollen germination and pollen tube elongation of species from different plant families. Plant Biol. 2015, 17, 825–830. [Google Scholar] [CrossRef]
- Gardner, M.J.; Dalling, K.J.; Light, M.E.; Jager, A.K.; van Staden, J. Does smoke substitute for red light in the germination of light-sensitive lettuce seeds by affecting gibberellin metabolism? S. Afr. J. Bot. 2001, 67, 636–640. [Google Scholar] [CrossRef]
- Egerton-Warburton, L.M. A smoke-induced alteration of the sub-testa cuticle in seeds of the post-fire recruiter, Emmenanthe penduliflora Benth (Hydrophyllaceae). J. Exp. Bot. 1998, 49, 1317–1327. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Amoo, S.O.; Kandari, L.S.; van Staden, J. Seed germination and phytochemical evaluation in seedlings of Aloe arborescens Mill. Plant Biosyst. 2013, 148, 460–466. [Google Scholar] [CrossRef]
- Zhou, J.; Da Silva, J.A.T.; Ma, G. Effects of smoke water and karrikin on seed germination of 13 species growing in China. Cent. Eur. J. Biol. 2014, 9, 1108–1116. [Google Scholar] [CrossRef]
- Jamil, M.; Kanwal, M.; Aslam, M.M.; Shakir, K.S.; Malook, I.; Tu, J.; Rehman, S. Effect of plant-derived smoke priming on physiological and biochemical characteristics of rice under salt stress condition. Aust. J. Crop Sci. 2014, 8, 159–170. [Google Scholar]
- Kamran, M.; Latif, K.A.; Waqas, M.; Imran, Q.M.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Kim, M.-J.; Lee, I.J. Effects of plant-derived smoke on the growth dynamics of Barnyard Grass (Echinochloa crus-galli). Acta Agric. Scand. Sect. B Soil Plant Sci. 2014, 64, 121–128. [Google Scholar] [CrossRef]
- Aremu, A.O.; Bairu, M.W.; Finnie, J.F.; van Staden, J. Stimulatory role of smoke-water and karrikinolide on the photosynthetic pigment and phenolic contents of micro-propagated ‘Williams’ bananas. Plant Growth Regul. 2012, 67, 271–279. [Google Scholar] [CrossRef]
- Chiwocha, S.D.S.; Dixon, K.W.; Flematti, G.R.; Ghisalberti, E.L.; Merritt, D.J.; Nelson, D.C.; Riseborough, J.M.; Smith, S.M.; Stevens, J.C. Karrikins: A new family of plant growth regulators in smoke. Plant Sci. 2009, 177, 252–256. [Google Scholar] [CrossRef]
- Chou, Y.-F.; Cox, R.D.; Wester, D.B. Smoke water and heat shock influence germination of shortgrass prairie species. Rangel. Ecol. Manag. 2012, 65, 260–267. [Google Scholar] [CrossRef]
- Soós, V.; Sebestyén, E.; Juhász, A.; Pintér, J.; Light, M.E.; van Staden, J.; Balázs, E. Stress-related genes define essential steps in the response of maize seedlings to smoke-water. Funct. Integr. Genom. 2009, 9, 231–242. [Google Scholar] [CrossRef]
- Aslam, M.M.; Jamil, M.; Khatoon, A.; Hendawy, S.E.; Suhaibani, N.A.; Malook, I.; Rehman, S. Physiological and biochemical responses of maize (Zea mays L.) to plant derived smoke solution. Pak. J. Bot. 2017, 49, 435–443. [Google Scholar]
- Sparg, S.G.; Kulkarni, M.G.; van Staden, J. Aerosol smoke and smoke-water stimulation of seedling vigor of a commercial maize cultivar. Crop Sci. 2006, 46, 1336–1340. [Google Scholar] [CrossRef]
- Abu, Y.; Romo, J.T.; Bai, Y.; Coulman, V. Priming seeds in aqueous smoke solutions to improve seed germination and biomass production of perennial forage species. Can. J. Plant Sci. 2016, 96, 551–563. [Google Scholar] [CrossRef]
- Light, M.E.; Gardner, M.J.; van Staden, J. Dual regulation of seed germination by smoke solutions. Plant Growth Regul. 2002, 37, 135–141. [Google Scholar] [CrossRef]
- Aremu, A.O.; Plackova, L.; Novak, O.; Stirk, W.A.; Dolezal, K.; van Staden, J. Cytokinin profiles in ex vitro acclimatized Eucomis autumnalis plants pre-treated with smoke-derived karrikinolide. Plant Cell Rep. 2016, 35, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. Identification of alkyl substituted 2H-furo[2,3-c]pyran-2-ones as germination stimulants present in smoke. J. Agric. Food Chem. 2009, 57, 9475–9480. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.G.; Sparg, S.G.; van Staden, J. Germination and post-germination response of Acacia seeds to smoke-water and butenolide, a smoke-derived compound. J. Arid Environ. 2007, 69, 177–187. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Ascough, G.D.; Verschaeve, L.; Baeten, K.; Arruda, M.P.; van Staden, J. Effect of smoke-water and a smoke-isolated butenolide on the growth and genotoxicity of commercial onion. Sci. Hort. 2010, 124, 434–439. [Google Scholar] [CrossRef]
- Abdollahi, M.R.; Mehrshad, B.; Moosavi, S.S. Effect of method of seed treatment with plant derived smoke solutions on germination and seedling growth of milk thistle (Silybum marianum L.). Seed Sci. Technol. 2011, 39, 225–229. [Google Scholar] [CrossRef]
- Staiger, C.J. Signalling to the actin cytoskeleton in plants. Annu. Rev. Plant Biol. 2000, 51, 257–288. [Google Scholar] [CrossRef] [PubMed]
- Schmelzer, E. Cell polarization, a crucial process in fungal defense. Trends Plant Sci. 2002, 7, 411–415. [Google Scholar] [CrossRef]
- Sun, T.; Li, S.; Ren, H. Profilin as a regulator of the membrane-actin cytoskeleton interface in plant cells. Front. Plant Sci. 2013. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007, 35, 61–65. [Google Scholar] [CrossRef]
- von Witsch, N.; Baluska, F.C.; Staiger, J.; Volkmann, D. Profilin is associated with the plasma membrane in microspores and pollen. Eur. J. Cell Biol. 1998, 77, 303–312. [Google Scholar] [CrossRef]
- Feller, U.; Anders, I.; Mae, T. Rubiscolytics: Fate of Rubisco after its enzymatic function in a cell is terminated. J. Exp. Bot. 2008, 59, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Bota, J.; Medrano, H.; Flexas, J. Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? New Phytol. 2004, 162, 671–681. [Google Scholar] [CrossRef]
- Khodadadi, E.; Fakheria, B.A.; Aharizad, S.; Emamjomeha, A.; Norouzic, M.; Komatsu, S. Leaf proteomics of drought-sensitive and -tolerant genotypes of fennel. Biochim. Biophys. Acta 2017, 1865, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Tezara, W.; Mitchell, V.; Driscoll, S.P.; Lawlor, D.W. Effects of water deficit and its interaction with CO2 supply on the biochemistry and physiology of photosynthesis in sunflower. J. Exp. Bot. 2002, 53, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- Baldrianová, J.; Černý, M.; Novák, J.; Jedelský, P.L.; Divíšková, E.; Brzobohatý, B. Arabidopsis proteome responses to the smoke-derived growth regulator karrikins. J. Proteom. 2015, 120, 7–20. [Google Scholar] [CrossRef]
- Sanahuja, G.; Farré, G.; Bassie, L.; Zhu, C.; Christou, P.; Capell, T. Ascorbic acid synthesis and metabolism in maize are subject to complex and genotype-dependent feedback regulation during endosperm development. Biotechnol. J. 2013, 8, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Su, D.; Lei, B.; Wang, F.; Geng, W.; Pan, G.; Cheng, F. Transcriptional profile of genes involved in ascorbate-glutathione cycle in senescing leaves for an early senescence leaf (esl) rice mutant. J. Plant Physiol. 2015, 176, 1–15. [Google Scholar] [CrossRef]
- Pandey, P.; Singh, J.; Achary, V.M.; Reddy, M.K. Redox homeostasis via gene families of ascorbate-glutathione pathway. Front. Environ. Sci. 2015, 3, 25. [Google Scholar] [CrossRef]
- Selote, D.S.; Chopra, P.K. Drought acclimation confers oxidative stress tolerance by inducing co-ordinated antioxidant defense at cellular and subcellular level in leaves of wheat seedlings. Physiol. Plant 2006, 127, 494–506. [Google Scholar] [CrossRef]
- Kausar, R.; Hossain, Z.; Makino, T.; Komatsu, S. Characterization of ascorbate peroxidase in soybean under flooding and drought stresses. Mol. Biol. Rep. 2012, 39, 10573–10579. [Google Scholar] [CrossRef]
- Scandalios, J.G. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Ascorbate peroxidase: A hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Baier, M.; Dietz, K.J. Chloroplasts as source and target of cellular redox regulation: A discussion on chloroplast redox signals in the context of plant physiology. J. Exp. Bot. 2005, 56, 1449–1462. [Google Scholar] [CrossRef] [PubMed]
- Waheed, M.A.; Jamil, M.; Khan, M.D.; Shakir, S.K.; Rehman, S.U. Effect of plant-derived smoke solutions on physiological and biochemical attributes of maize (Zea mays L.) under salt stress. Pak. J. Bot. 2016, 48, 1763–1774. [Google Scholar]
- Naranjo, B.; Diaz-Espejo, A.; Lindahl, M.; Cejudo, F.J. Type-f thioredoxins have a role in the short-term activation of carbon metabolism and their loss affects growth under short-day conditions in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 1951–1964. [Google Scholar] [CrossRef]
- Bharwana, S.A.; Ali, S.; Farooq, M.A.; Iqbal, N.; Abbas, F.; Ahmad, M.S.A. Alleviation of lead toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes suppressed lead uptake and oxidative stress in cotton. J. Bioremed. Biodeg. 2013, 4, 187. [Google Scholar] [CrossRef]
- El-Gaied, L.F.; Abu El-Heba, G.A.; El-Sherif, N.A. Effect of growth hormones on some antioxidant parameters and gene expression in tomato. GM Crops Food 2013, 4, 67–73. [Google Scholar] [CrossRef]
- Pasternak, T.P.; Potters, G.; Caubergs, R.; Jansen, M.A.K. Complementary interactions between oxidative stress and auxins control plant growth responses at plant, organ, and cellular level. J. Exp. Bot. 2005, 56, 1991–2001. [Google Scholar] [CrossRef]
- Correa-Aragunde, N.; Foresi, N.; Delledonne, M.; Lamattina, L. Auxin induces redox regulation of ascorbate peroxidase 1 activity by S-nitrosylation/denitrosylation balance resulting in changes of root growth pattern in Arabidopsis. J. Exp. Bot. 2013, 64, 3339–3349. [Google Scholar] [CrossRef]
- Hernández, J.A.; Ferrer, M.A.; Jiménez, A.; Ros-Barceló, A.; Sevilla, F. Antioxidant systems and O2−/H2O2 production in the apoplast of Pisum sativum L. leaves: Its relation with NaCl-induced necrotic lesions in minor veins. Plant Physiol. 2001, 127, 817–831. [Google Scholar] [CrossRef]
- Diaz-Vivancos, P.; Faize, M.; Barba-Espin, G.; Faize, L.; Petri, C.; Hernández, J.A.; Burgos, L. Ectopic expression of cytosolic superoxide dismutase and ascorbate peroxidase leads to salt stress tolerance in transgenic plums. Plant Biotechnol. J. 2013, 11, 976–985. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta 2012, 1819, 120–128. [Google Scholar] [CrossRef]
- de Lange, J.H.; Boucher, C. Auto ecological studies on Audinia capitate (Bruniaceaae), plant-derived smoke as a germination cue. S. Afr. J. Bot. 1990, 56, 188–202. [Google Scholar] [CrossRef]
- Tieu, A.; Dixon, K.A.; Sivasithamparam, K.; Plummer, J.A. Germination of four species of native Western Australian plant using plant-derived smoke. Aust. J. Bot. 1999, 47, 207–219. [Google Scholar] [CrossRef]
- Komatsu, S.; Han, C.; Nanjo, Y.; Altaf-Un-Nahar, M.; Wang, K.; He, D.; Yang, P. Label-free quantitative proteomic analysis of abscisic acid effect in early-stage soybean under flooding. J. Proteome Res. 2013, 12, 4769–4784. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, Z.; Washburn, M.P.; Florens, L. Effect of dynamic exclusion duration on spectral count based quantitative proteomics. Anal. Chem. 2009, 81, 6317–6326. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Siniteyn, P.; Carlson, A.; Hein, Y.; Gieger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of proteomics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.A.; Côté, R.G.; Csordas, A.; Dianes, J.A.; Fabregat, A.; Foster, J.M.; Griss, J.; Alpi, E.; Birim, M.; Contell, J.; et al. The proteomics identifications (PRIDE) database and associated tools: Status in 2013. Nucleic Acids Res. 2013, 41, D1063–D1069. [Google Scholar] [CrossRef] [PubMed]
- Okuda, S.; Watanabe, Y.; Moriya, Y.; Kawano, S.; Yamamoto, T.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Araki, N.; Yoshizawa, A.C.; et al. jPOSTrepo: An international standard data repository for proteomes. Nucleic Acids Res. 2017, 45, D1107–D1111. [Google Scholar] [CrossRef]
- Usadel, B.; Nagel, A.; Thimm, O.; Redestig, H.; Blaesing, O.E.; Palacios-Rofas, N.; Selbig, J.; Hannemann, J.; Piques, M.C.; Steinhauser, D.; et al. Extension of the visualization tool MapMan to allow statistical analysis of arrays, display of corresponding genes and comparison with known responses. Plant Physiol. 2005, 138, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Usadel, B.; Poree, F.; Nagel, A.; Lohse, M.; Czedik-Eysenberg, A.; Stitt, M. A guide to using MapMan to visualize and compare Omics data in plants: A case study in the crop species, maize. Plant Cell Environ. 2009, 32, 1211–1229. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Yamamoto, A.; Nakamura, T.; Nouri, M.Z.; Nanjo, Y.; Nishizawa, K.; Furukawa, K. Comprehensive analysis of mitochondria in roots and hypocotyls of soybean under stress using proteomics and metabolomics techniques. J. Proteome Res. 2011, 10, 3993–4004. [Google Scholar] [CrossRef]
- Nishizawa, K.; Komatsu, S. Characteristics of soybean 1-cys peroxiredoxin and its behavior in seedlings under flooding stress. Plant Biotechnol. J. 2011, 28, 83–88. [Google Scholar] [CrossRef]
- Hashimoto, M.; Komatsu, S. Proteomic analysis of rice seedlings during cold stress. Proteomics 2007, 7, 1293–1302. [Google Scholar] [CrossRef]
- Komatsu, S.; Masuda, T.; Abe, K. Phosphorylation of a protein (pp56) is related to the regeneration of rice cultured suspension cells. Plant Cell Physiol. 1996, 37, 748–753. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen-peroxide is scavenged by ascorbate-specific peroxidase in spinachchloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Horling, F.; König, J.; Dietz, K.J. Type II peroxiredoxin C, a member of the peroxiredoxin family of Arabidopsis thaliana: Its expression and activity in comparison with other peroxiredoxins. Plant Physiol. Biochem. 2002, 40, 491–499. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Accession | Description | Fold Change | Functional Category | Biological Process | Cellular Component | Molecular Function |
|---|---|---|---|---|---|---|---|
| 1 | B4FTX3 | Profilin | 7.22 | Cell | not assigned | cell part | not assigned |
| 2 | B9RWF4 | Elongation factor 1-alpha | 6.79 | Protein | not assigned | cell part | binding |
| 3 | A0A1D6BQY9 | Tubulin beta chain | 4.91 | Cell | not assigned | not assigned | not assigned |
| 4 | C5XR12 | Uncharacterized protein | 4.89 | Not assigned | not assigned | not assigned | catalytic activity |
| 5 | K7TFA7 | Aldo/keto reductase family protein | 4.17 | Redox | biosynthetic process | cell part | catalytic activity |
| 6 | C9VWQ3 | Actin | 3.99 | Cell | not assigned | not assigned | not assigned |
| 7 | A0A1D6DZR9 | AICARFT/IMPCHase bienzyme | 3.11 | Nucleotide metabolism | not assigned | not assigned | not assigned |
| 8 | C5YEF7 | Uncharacterized protein | 2.91 | Not assigned | not assigned | cell part | adenyl nucleotide binding |
| 9 | B6T289 | Glucan endo-1,3-β-glucosidase 7 | 2.55 | Not assigned | carbohydrate metabolic process | not assigned | catalytic activity |
| 10 | A0A1B6QAM8 | Uncharacterized protein | 2.36 | Major CHO metabolism | not assigned | not assigned | not assigned |
| 11 | B4FTR5 | Uncharacterized protein | 1.95 | Not assigned | cell communication | cell part | not assigned |
| 12 | C8ZK16 | Malic enzyme | 1.37 | Tricarboxylic acid cycle | not assigned | not assigned | not assigned |
| 13 | C0PG78 | Monocopper oxidase-like protein SKU5 | 0.91 | Development | not assigned | not assigned | binding |
| 14 | A0A1D6M373 | Mannosylglycoprotein endo-beta-mannosidase | 0.91 | Miscellaneous | not assigned | not assigned | not assigned |
| 15 | A0A0P0X334 | Os07g0176900 protein | 0.89 | Photosynthesis | not assigned | not assigned | not assigned |
| 16 | B4FH75 | 4-Hydroxy-tetrahydrodipicolinate reductase 2 | −0.51 | Amino acid metabolism | amine biosynthetic process | cell part | binding |
| 17 | B4FAW7 | Histidinol-phosphate aminotransferase 2 | −0.70 | Amino acid metabolism | amine biosynthetic process | not assigned | binding |
| 18 | A0A1D6KWD0 | Lysine–tRNA ligase | −0.85 | Protein | not assigned | not assigned | not assigned |
| 19 | A0A1D6LT87 | Ras-related protein | −0.89 | Signaling | not assigned | not assigned | not assigned |
| 20 | O24574 | RuBisCO small chain | −1.05 | Photosynthesis | biosynthetic process | cell part | carbon-carbon lyase activity |
| 21 | Q5I204 | Brain acid soluble protein | −1.08 | RNA | not assigned | cell part | binding |
| 22 | A0A1Q0YQ12 | Oil body-associated protein 1B | −1.25 | Not assigned | not assigned | not assigned | not assigned |
| 23 | A0A1D6I8U5 | Flowering time control protein FPA | −1.31 | RNA | not assigned | not assigned | not assigned |
| 24 | K7UK47 | Clathrin interactor EPSIN | −1.69 | Not assigned | not assigned | not assigned | not assigned |
| 25 | A0A0E0CRT7 | Uncharacterized protein | −1.80 | Cell | not assigned | not assigned | not assigned |
| 26 | A0A0D9XRV3 | Adenosylhomocysteinase | −1.88 | Amino acid metabolism | not assigned | not assigned | not assigned |
| 27 | A0A1D6MQK0 | Rab escort protein 1 | −1.89 | Signaling | not assigned | not assigned | not assigned |
| 28 | B6TBI9 | Pyridoxamine 5-phosphate oxidase | −1.95 | Not assigned | not assigned | not assigned | binding |
| 29 | A0A0E0JEZ9 | Uncharacterized protein | −1.96 | Not assigned | not assigned | not assigned | not assigned |
| 30 | B4FTH5 | Xyloglucan endotransglucosylase/hydrolase | −2.06 | Cell wall | carbohydrate metabolic process | apoplast | catalytic activity |
| 31 | B4FZJ2 | β-Glucosidase 11 | −2.13 | Misce | carbohydrate metabolic process | not assigned | catalytic activity |
| 32 | B6TUP8 | Zinc finger homeodomain protein 1 | −2.21 | RNA | not assigned | not assigned | binding |
| 33 | A0A1D6H8U6 | 3-Oxoacyl-[acyl-carrier-protein] reductase | −2.31 | Lipid metabolism | not assigned | not assigned | not assigned |
| 34 | A0A1D6EZ65 | Uncharacterized protein | −2.38 | Not assigned | not assigned | not assigned | not assigned |
| 35 | A0A1D6GLX4 | DEK domain-containing chromatin associated protein | −2.54 | Not assigned | not assigned | not assigned | not assigned |
| 36 | B6TQH7 | THA4 | −2.63 | Not assigned | cellular process | cell part | protein transporter activity |
| 37 | C5WUG0 | Mitogen-activated protein kinase | −2.75 | Signaling | not assigned | not assigned | adenyl nucleotide binding |
| 38 | A0A0K9PCU0 | RPM1-interacting protein 4 | −2.80 | Not assigned | not assigned | not assigned | not assigned |
| 39 | B6TIG8 | Protein arginine N-methyltransferase 1 | −2.80 | Miscellaneous | cellular macromolecule metabolic process | not assigned | catalytic activity |
| 40 | B6TR82 | Thioredoxin F-type | −2.81 | Redox | biological regulation | not assigned | catalytic activity |
| 41 | K3Y2G0 | Uncharacterized protein | −2.83 | Redox | catabolic process | cell part | not assigned |
| 42 | A0A0D3GFF5 | Uncharacterized protein | −2.85 | Stress | not assigned | not assigned | not assigned |
| 43 | B6TYK8 | Putative uncharacterized protein | −2.87 | Not assigned | not assigned | not assigned | not assigned |
| 44 | A0A1D6FY68 | SIT4 phosphatase-associated | −2.95 | Metal handling | not assigned | not assigned | not assigned |
| 45 | A0A1E5V130 | Uncharacterized protein | −3.01 | Not assigned | not assigned | not assigned | not assigned |
| 46 | A0A0N7KSP9 | Os11g0247300 protein | −3.18 | Cell | not assigned | not assigned | not assigned |
| 47 | A0A1D6JR65 | Alcohol dehydrogenase-like 2 | −3.22 | Miscellaneous | not assigned | not assigned | not assigned |
| 48 | B6SW97 | Putative uncharacterized protein | −3.28 | Protein | not assigned | not assigned | not assigned |
| 49 | B6U581 | Ribosome-like protein | −3.31 | Protein | biosynthetic process | cell part | structural constituent of ribosome |
| 50 | K4BGM2 | Uncharacterized protein | −3.40 | Protein | biological regulation | cell part | binding |
| 51 | B5QSJ9 | Acetyl-coenzyme A carboxylase | −3.53 | Lipid metabolism | biosynthetic process | not assigned | acetyl-CoA carboxylase activity |
| 52 | A0A1D6L558 | Plasmodesmata callose-binding protein 2 | −3.67 | Miscellaneous | not assigned | not assigned | not assigned |
| 53 | A0A0K9NNM0 | Cysteine proteinase cathepsin F | −3.69 | Protein | not assigned | not assigned | not assigned |
| 54 | C0P3K6 | Aspartic proteinase A1 | −3.75 | Protein | lipid metabolic process | not assigned | aspartic-type endopeptidase activity |
| 55 | J3LDT9 | Uncharacterized protein | −3.76 | Not assigned | anatomical structure morphogenesis | cell part | not assigned |
| 56 | B9MSV5 | Tubulin alpha chain | −3.77 | Cell | cellular component assembly | cell part | binding |
| 57 | A0A068UVK8 | Chlorophyll a-b binding protein | −3.84 | Photosynthesis | cellular macromolecule metabolic process | cell part | binding |
| 58 | B6SZR1 | Chlorophyll a-b binding protein | −3.97 | Photosynthesis | cellular macromolecule metabolic process | cell part | binding |
| 59 | A0A1D6LJZ2 | 30S ribosomal protein S16 | −4.00 | Protein | not assigned | not assigned | not assigned |
| 60 | B4FV94 | Chlorophyll a-b binding protein | −4.06 | Photosynthesis | cellular macromolecule metabolic process | cell part | binding |
| 61 | E9KIP1 | Photosystem I P700 chlorophyll apoprotein A1 | −4.13 | Photosynthesis | cellular macromolecule metabolic process | cell part | 4 iron, 4 sulfur cluster binding |
| 62 | A0A061EVS4 | Nascent polypeptide-associated complex beta | −4.23 | RNA | biological regulation | not assigned | not assigned |
| 63 | A0A1D6IIC3 | Nuclear transport factor 2 | −4.38 | Protein | not assigned | not assigned | not assigned |
| 64 | A0A1D5WFY2 | Small ubiquitin-related modifier | −4.40 | Protein | not assigned | not assigned | not assigned |
| 65 | A0A1D6PYA1 | 60S ribosomal protein L17 | −4.61 | Protein | not assigned | not assigned | not assigned |
| 66 | Q8H6N0 | Tubulin beta chain | −4.76 | Cell | cellular component assembly | cell part | binding |
| 67 | A0A1D5AHD9 | RuBisCO large chain | −4.79 | Photosynthesis | not assigned | not assigned | not assigned |
| 68 | A0A1D1Z067 | Elongation factor 1-alpha | −6.21 | Protein | not assigned | not assigned | not assigned |
| 69 | A0A097PJF2 | Structural maintenance of chromosomes protein 1 | −9.99 | Cell | cell cycle process | cell part | adenyl nucleotide binding |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, M.M.; Rehman, S.; Khatoon, A.; Jamil, M.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Li, X.; Sunohara, Y.; Matsumoto, H.; et al. Molecular Responses of Maize Shoot to a Plant Derived Smoke Solution. Int. J. Mol. Sci. 2019, 20, 1319. https://doi.org/10.3390/ijms20061319
Aslam MM, Rehman S, Khatoon A, Jamil M, Yamaguchi H, Hitachi K, Tsuchida K, Li X, Sunohara Y, Matsumoto H, et al. Molecular Responses of Maize Shoot to a Plant Derived Smoke Solution. International Journal of Molecular Sciences. 2019; 20(6):1319. https://doi.org/10.3390/ijms20061319
Chicago/Turabian StyleAslam, Muhammad Mudasar, Shafiq Rehman, Amana Khatoon, Muhammad Jamil, Hisateru Yamaguchi, Keisuke Hitachi, Kunihiro Tsuchida, Xinyue Li, Yukari Sunohara, Hiroshi Matsumoto, and et al. 2019. "Molecular Responses of Maize Shoot to a Plant Derived Smoke Solution" International Journal of Molecular Sciences 20, no. 6: 1319. https://doi.org/10.3390/ijms20061319
APA StyleAslam, M. M., Rehman, S., Khatoon, A., Jamil, M., Yamaguchi, H., Hitachi, K., Tsuchida, K., Li, X., Sunohara, Y., Matsumoto, H., & Komatsu, S. (2019). Molecular Responses of Maize Shoot to a Plant Derived Smoke Solution. International Journal of Molecular Sciences, 20(6), 1319. https://doi.org/10.3390/ijms20061319