Interaction among Calcium Diet Content, PTH (1-34) Treatment and Balance of Bone Homeostasis in Rat Model: The Trabecular Bone as Keystone

 , , ,
, , , 
Abstract
1. Introduction
2. Results
2.1. Body Weight
2.2. Histology and Histomorphometry
2.2.1. Vertebral body (L5)
2.2.2. Femoral Mid-Diaphysis
2.2.3. Femoral Distal Metaphysis
2.3. Sclerostin–Immunohistochemical Analysis
2.4. Serum Biochemical Analysis
3. Discussion
4. Materials and Methods
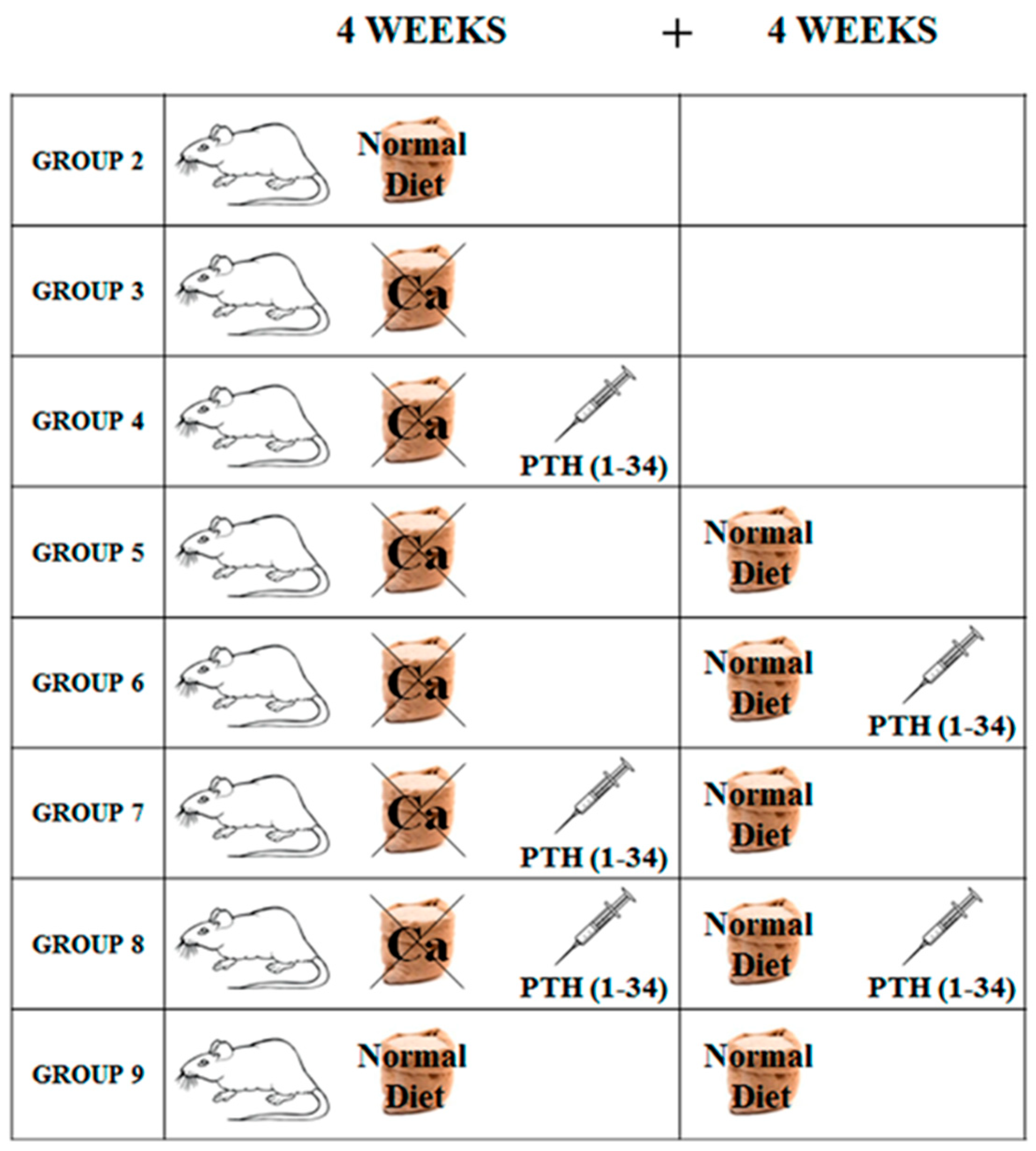
4.1. Experimental Animals and Treatment
- Group 1 (baseline, n = 3): sacrificed after 7 days of acclimation;
- Group 2 (control, n = 5): fed normal diet and natural water ad libitum for 4 weeks (T1);
- Group 3 (n = 5): fed calcium-deprived diet and distilled water ad libitum for 4 weeks (T1);
- Group 4 (n = 5): fed calcium-deprived diet and distilled water ad libitum for 4 weeks with concomitant administration of PTH (1-34) 40 μg/kg/day (T1);
- Group 5 (n = 5): fed calcium-deprived diet and distilled water ad libitum for 4 weeks (T1) and successive normal diet restoration and natural water ad libitum for 4 weeks (T2);
- Group 6 (n = 5): fed calcium-deprived diet and distilled water ad libitum for 4 weeks (T1) and successive normal diet restoration and natural water ad libitum for 4 weeks with concomitant administration of PTH (1-34) 40 μg/kg/day (T2);
- Group 7 (n = 5): fed calcium-deprived diet and distilled water ad libitum for 4 weeks with concomitant administration of PTH (1-34) 40 μg/kg/day (T1) and successive normal diet restoration and natural water ad libitum for 4 weeks (T2);
- Group 8 (n = 5): fed calcium-deprived diet and distilled water ad libitum for 4 weeks with concomitant administration of PTH (1-34) 40 μg/kg/day (T1) and successive normal diet restoration and natural water ad libitum for 4 weeks with concomitant administration of PTH (1-34) 40 μg/kg/day (T2);
- Group 9 (control, n = 5): fed normal diet and natural water ad libitum for 8 weeks (T2).
4.2. Histology and Histomorphometry
- In vertebral body and femoral distal metaphysis: trabecular bone volume (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N), and trabecular separation (Tb.Sp);
- In anterolateral and posterior sides of the vertebral body: cortical bone thickness (Ct.Th);
- In femoral mid-diaphysis and distal metaphysis: cortical bone area (Ct.B.Ar).
4.3. Immunohistochemical Analysis
4.4. Serum Biochemical Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kleerekoper, M.; Villanueva, A.R.; Stanciu, J.; Rao, D.S.; Parfitt, A.M. The role of three-dimensional trabecular microstructure in the pathogenesis of vertebral compression fractures. Calcif. Tissue Int. 1985, 37, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.L.; Slemenda, C.W.; Johnston, C.C., Jr. The contribution of bone loss to postmenopausal osteoporosis. Osteoporos. Int. 1990, 1, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, D.B.; Recker, R.R.; Gallagher, J.C.; Vaswani, A.S.; Aloia, J.F. A comparison of iliac bone histomorphometric data in post-menopausal osteoporotic and normal subjects. Bone Miner. 1990, 11, 217–235. [Google Scholar] [CrossRef]
- Peck, W.A.; Burckhardt, P.; Christiansen, C. Consensus development conference: Diagnosis, prophylaxis, and treatment of osteoporosis. Am. J. Med. 1993, 94, 646–650. [Google Scholar]
- Heaney, R.P. Calcium, dairy products and osteoporosis. J. Am. Coll. Nutr. 2000, 19, 83S–99S. [Google Scholar] [CrossRef] [PubMed]
- Recker, R.R.; Barger-Lux, M.J. The elusive concept of bone quality. Curr. Osteoporos. Rep. 2004, 2, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, C.; Ferretti, M.; Bertoni, L.; Cavani, F.; Resca, E.; Casolari, B.; Carnevale, G.; Zavatti, M.; Montanari, C.; Benelli, A.; et al. Influence of ferutinin on bone metabolism in ovariectomized rats. I: Role in preventing osteoporosis. J. Bone Miner. Metab. 2009, 27, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.; Bertoni, L.; Cavani, F.; Zavatti, M.; Resca, E.; Carnevale, G.; Benelli, A.; Zanoli, P.; Palumbo, C. Influence of ferutinin on bone metabolism in ovariectomized rats. II: Role in recovering osteoporosis. J. Anat. 2010, 217, 48–56. [Google Scholar] [CrossRef]
- Kim, T.H.; Jung, J.W.; Ha, B.G.; Hong, J.M.; Park, E.K.; Kim, H.J.; Kim, S.Y. The effects of luteolin on osteoclast differentiation, function in vitro and ovariectomy-induced bone loss. J. Nutr. Biochem. 2011, 22, 8–15. [Google Scholar] [CrossRef]
- Cavani, F.; Ferretti, M.; Carnevale, G.; Bertoni, L.; Zavatti, M.; Palumbo, C. Effects of different doses of ferutinin on bone formation/resorption in ovariectomized rats. J. Bone Miner. Metab. 2012, 30, 619–629. [Google Scholar] [CrossRef]
- Prentice, A. Diet, nutrition and the prevention of osteoporosis. Pub. Health Nutr. 2004, 7, 227–243. [Google Scholar] [CrossRef]
- Nieves, J.W. Osteoporosis: The role of micronutrients. Am. J. Clin. Nutr. 2005, 81, 1232S–1239S. [Google Scholar] [CrossRef] [PubMed]
- Recker, R.R.; Davies, K.M.; Hinders, S.M.; Heaney, R.P.; Stegman, M.R.; Kimmel, D.B. Bone gain in young adult women. JAMA 1992, 268, 2403–2408. [Google Scholar] [CrossRef] [PubMed]
- Broadus, A.E. Physiological function of calcium, magnesium, and phosphorus and mineral ion balance. In Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism; Favus, M.J., Ed.; Raven Press: New York, NY, USA, 1993; pp. 41–46. [Google Scholar]
- Mohamad, N.V.; Ima-Nirwana, S.; Chin, K.Y. Effect of tocotrienol from Bixa orellana (annatto) on bone microstructure, calcium content, and biomechanical strength in a model of male osteoporosis induced by buserelin. Drug Des. Devel. Ther. 2018, 12, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Smerdely, P. Hypocalcaemia after denosumab in older people following fracture. Osteoporos. Int. 2017, 28, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Arjmandi, B.H.; Khalil, D.A.; Hollis, B.W. Ipriflavone, a synthetic phytoestrogen, enhances intestinal calcium transport in vitro. Calcif. Tissue Int. 2000, 67, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C. Nutrition and metabolism of vitamins. In Nutritional Biochemistry and Metabolism with Clinical Applications, 2nd ed.; Linder, M.C., Ed.; Elsevier: New York, NY, USA, 1991. [Google Scholar]
- Wang, L.; Tran, A.B.; Nociti, F.H., Jr.; Thumbigere-Math, V.; Foster, B.L.; Krieger, C.C.; Kantovitz, K.R.; Novince, C.M.; Koh, A.J.; McCauley, L.K.; et al. PTH and vitamin D repress DMP1 in cementoblasts. J. Dent. Res. 2015, 94, 1408–1416. [Google Scholar] [CrossRef] [PubMed]
- Hock, J.M.; Gera, I. Effects of continuous and intermittent administration and inhibition of resorption on the anabolic response of bone to parathyroid hormone. J. Bone Miner. Res. 1992, 7, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Burr, D.B.; Hirano, T.; Turner, C.H.; Hotchkiss, C.; Brommage, R.; Hock, J.M. Intermittently administered human parathyroid hormone (1–34) treatment increases intracortical bone turnover and porosity without reducing bone strength in the humerus of ovariectomized cynomolgus monkeys. J. Bone Miner. Res. 2001, 16, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Almagro, M.I.; Roman-Blas, J.A.; Bellido, M.; Castañeda, S.; Cortez, R.; Herrero-Beaumont, G. PTH [1-34] enhances bone response around titanium implants in a rabbit model of osteoporosis. Clin. Oral Implants Res. 2013, 24, 1027–1034. [Google Scholar] [CrossRef]
- Oswald, A.J.; Berg, J.; Milne, G.; Ralston, S.H. Teriparatide treatment of severe osteoporosis reduces the risk of vertebral fractures compared with standard care in routine clinical practice. Calcif. Tissue Int. 2014, 94, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Saidak, Z.; Le Henaff, C.; Azzi, S.; Marty, C.; Marie, P.J. Low-dose PTH increases osteoblast activity via decreased Mef2c/Sost in senescent osteopenic mice. J. Endocrinol. 2014, 223, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Takao-Kawabata, R.; Isogai, Y.; Takakura, A.; Shimazu, Y.; Sugimoto, E.; Nakazono, O.; Ikegaki, I.; Kuriyama, H.; Tanaka, S.; Oda, H.; et al. Three-times-weekly administration of teriparatide improves vertebral and peripheral bone density, microarchitecture, and mechanical properties without accelerating bone resorption in ovariectomized rats. Calcif. Tissue Int. 2015, 97, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.L.; Zeng, Q.; Donley, D.W.; Ste-Marie, L.G.; Gallagher, J.C.; Dalsky, G.P.; Marcus, R.; Eriksen, E.F. Teriparatide increases bone formation in modeling and remodeling osteons and enhances IGF-II immunoreactivity in postmenopausal women with osteoporosis. J. Bone Miner. Res. 2006, 21, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T. Intermittent parathyroidhormone therapy to increase bone formation. Jt. Bone Spine 2006, 73, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Jilka, R.L. Molecular and cellular mechanisms of the anabolic effect of intermittent PTH. Bone 2007, 40, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, R.; Zhou, H.; Cosman, F.; Nieves, J.; Dempster, D.W.; Hodsman, A.B. Effects of a one-month treatment with PTH (1-34) on bone formation on cancellous, endocortical, and periosteal surfaces of the human ilium. J. Bone Miner. Res. 2007, 22, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Kusuzaki, K.; Kageyama, N.; Shinjo, H.; Takeshita, H.; Murata, H.; Hashiguchi, S.; Ashihara, T.; Hirasawa, Y. Development of bone canaliculi during bone repair. Bone 2000, 27, 655–659. [Google Scholar] [CrossRef]
- Skripitz, R.; Andreassen, T.T.; Aspenberg, P. Strong effect of PTH (1-34) on regenerating bone: A time sequence study in rats. Acta Orthop. Scand. 2000, 71, 619–624. [Google Scholar] [CrossRef]
- Chiba, S.; Okada, K.; Lee, K.; Segre, G.V.; Neer, R.M. Molecular analysis of defect healing in rat diaphyseal bone. J. Vet. Med. Sci. 2001, 63, 603–608. [Google Scholar] [CrossRef]
- Karp, J.M.; Sarraf, F.; Shoichet, M.S.; Davies, J.E. Fibrin-filled scaffolds for bone-tissue engineering: An in vivo study. J. Biomed. Mater Res. A 2004, 71, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.C.; Fernandes, R.G.; Carvalh, Y.R.; Balducci, I.; Faig-Leite, H. Bone healing in drill hole defects in spontaneously hypertensive male and female rats’ femurs. A histological and histometric study. Arq. Bras. Cardiol. 2007, 88, 93–97. [Google Scholar]
- Oana, L.; Miclaus, V.; Oros, D.; Almasan, H.; Ratiu, C.; Rus, V.; Pestea, C.; Ober, C.; Crecan, C.; Bel, L. Experimental method of inducing a bone defect in rat and histological monitoring of the evolution of the healing processes. Ann. RSCB 2008, 16, 111–114. [Google Scholar]
- Jilka, R.L.; O’Brien, C.A.; Ali, A.A.; Roberson, P.K.; Weinstein, R.S.; Manolagas, S.C. Intermittent PTH stimulates periosteal bone formation by actions on post-mitotic preosteoblasts. Bone 2009, 44, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Tagil, M.; McDonald, M.M.; Morse, A.; Peacock, L.; Mikulec, K.; Amanat, N.; Godfrey, C.; Little, D.G. Intermittent PTH (1-34) does not increase union rates in open rat femoral fractures and exhibits attenuated anabolic effects compared to closed fractures. Bone 2010, 46, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Noda, H.; Joyashiki, E.; Nakagawa, C.; Asanuma, K.; Hayasaka, A.; Kato, M.; Nanami, M.; Inada, M.; Miyaura, C.; et al. The optimal duration of PTH (1–34) infusion is one hour per day to increase bone mass in rats. Biol. Pharm. Bull. 2016, 39, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.; Cavani, F.; Smargiassi, A.; Roli, L.; Palumbo, C. Mineral and skeletal homeostasis influence the manner of bone loss in metabolic osteoporosis due to calcium-deprived diet in different sites of rat vertebra and femur. BioMed. Res. Int. 2015, 2015, ID-304178. [Google Scholar] [CrossRef] [PubMed]
- Orwoll, E.S.; Scheele, W.H.; Paul, S.; Adami, S.; Syversen, U.; Diez-Perez, A.; Kaufman, J.M.; Clancy, A.D.; Gaich, G.A. The effect of teriparatide [human parathyroid hormone (1–34)] therapy on bone density in men with osteoporosis. J. Bone Miner. Res. 2003, 18, 9–17. [Google Scholar] [CrossRef]
- Tanaka, S.; Kuroda, T.; Sugimoto, T.; Nakamura, T.; Shiraki, M. Changes in bone mineral density, bone turnover markers, and vertebral fracture risk reduction with once weekly teriparatide. Curr. Med. Res. Opin. 2014, 30, 931–936. [Google Scholar] [CrossRef]
- Hasegawa, T.; Amizuka, N. Bone remodeling and modeling/mini-modeling. Clin. Calcium 2017, 27, 1713–1722. [Google Scholar]
- Kumabe, Y.; Lee, S.Y.; Waki, T.; Iwakura, T.; Takahara, S.; Arakura, M.; Kuroiwa, Y.; Fukui, T.; Matsumoto, T.; Matsushita, T.; et al. Triweekly administration of parathyroid hormone (1–34) accelerates bone healing in a rat refractory fracture model. BMC Musculoskelet. Disord. 2017, 18, 545. [Google Scholar] [CrossRef] [PubMed]
- Canalis, E. Management of endocrine disease: Novel anabolic treatments for osteoporosis. Eur. J. Endocrinol. 2018, 178, R33–R44. [Google Scholar] [CrossRef] [PubMed]
- Liebschner, M.A. Biomechanical considerations of animal models used in tissue engineering of bone. Biomaterials 2004, 25, 1697–1714. [Google Scholar] [CrossRef]
- Ferretti, M.; Palumbo, C.; Marotti, G. Static and dynamic bone osteogenesis: Two different types of bone formation. Anat. Embryol. 2002, 206, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Cavani, F.; Ferretti, M.; Smargiassi, A.; Palumbo, C. PTH (1-34) effects on repairing experimentally drilled holes in rat femur: Novel aspects—qualitative vs. quantitative improvement of osteogenesis. J. Anat. 2017, 230, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Netzer, C.; Distel, P.; Wolfram, U.; Deyhle, H.; Jost, G.F.; Schären, S.; Geurts, J. Comparative analysis of bone structural parameters reveals subchondral cortical plate resorption and increased trabecular bone remodeling in human facet joint osteoarthritis. Int. J. Mol. Sci. 2018, 19, 845. [Google Scholar] [CrossRef] [PubMed]
- Dehority, W.; Halloran, B.P.; Bikle, D.D.; Curren, T.; Kostenuik, P.J.; Wronski, T.J.; Shen, Y.; Rabkin, B.; Bouraoui, A.; Morey-Holton, E. Bone and hormonal changes induced by skeletal unloading in the mature male rat. Am. J. Physiol. 1999, 276, 62–69. [Google Scholar] [CrossRef]
- Iwamoto, J.; Yeh, J.K.; Aloia, J.F. Differential effect of treadmill exercise on three cancellous bone sites in the young growing rat. Bone 1999, 24, 163–169. [Google Scholar] [CrossRef]
- Lecoq, B.; Potrel-Burgot, C.; Granier, P.; Sabatier, J.P.; Marcelli, C. Comparison of bone loss induced in female rats by hindlimb unloading, ovariectomy, or both. Jt. Bone Spine 2006, 73, 189–195. [Google Scholar] [CrossRef]
- Burgers, T.A.; Williams, B.O. Regulation of Wnt/β-catenin signaling within and from osteocytes. Bone 2013, 54, 244–249. [Google Scholar] [CrossRef]
- Goto, S.; Fujita, Y.; Hotta, M.; Sugiyama, A.; Maki, K. Influence of differences in the hardness and calcium content of diets on the growth of craniofacial bone in rats. Angle Orthod. 2015, 85, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Goto, S.; Ichikawa, M.; Hamaguchi, A.; Maki, K. Effect of dietary calcium deficiency and altered diet hardness on the jawbone growth: A micro-CT and bone histomorphometric study in rats. Arch. Oral Biol. 2016, 72, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Weaver, C.M. Calcium. In Present Knowledge in Nutrition, 9th ed.; ILSI Press: Washington, DC, USA, 2006; Volume 1. [Google Scholar]
- Parfitt, A.M.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R. Bone histomorphometry: Standardization of nomenclature, symbols, and units. J. Bone Miner. Res. 1987, 2, 595–610. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Weight at TA | Weight at T0 | Weight at T1 | Weight at T2 |
|---|---|---|---|---|
| 2 | 405 ± 32 | 436 ± 35 °°° | 526 ± 47 *** | |
| 3 | 405 ± 32 | 429 ± 37 °° | 528 ± 70 ** | |
| 4 | 397 ± 25 | 422 ± 21 °° | 504 ± 40 ** | |
| 5 | 398 ± 24 | 423 ± 30 °°° | 505 ± 48 ** | 551 ± 65 ## |
| 6 | 400 ± 26 | 417 ± 23 °° | 518 ± 28 ** | 533 ± 30 |
| 7 | 403 ± 29 | 425 ± 29 °° | 507 ± 34 *** | 555 ± 44 ## |
| 8 | 405 ± 28 | 428 ± 36 °° | 503 ± 55 ** | 534 ± 62 ## |
| 9 | 406 ± 28 | 430 ± 29 °°° | 523 ± 54 ** | 559 ± 74 # |
| Group | Ca mg/dL | P mg/dL | OPG ng/mL | BALP ng/mL | Cross Laps ng/mL | PTH (1-84) pg/mL |
|---|---|---|---|---|---|---|
| 2 | 9.74 ± 0.28 | 6.65 ± 0.92 | 0.71 ± 0.04 | 6.90 ± 0.63 | 51.91 ± 19.16 | 54.52 ± 25.85 |
| 3 | 10.01 ± 0.16 | 7.35 ± 0.87 | 0.67 ± 0.10 | 6.84 ± 0.41 | 66.36 ± 17.20 | 78.88 ± 29.79 |
| 4 | 10.12 ± 0.28 | 7.04 ± 0.72 | 0.69 ± 0.05 | 6.63 ± 0.37 | 58.24 ± 20.88 | 62.41 ± 23.35 |
| 5 | 10.15 ± 0.27 | 5.89 ± 0.41 | 0.69 ± 0.07 | 6.51 ± 1.32 | 44.84 ± 11.86 | 58.41 ± 12.65 |
| 6 | 10.34 ± 0.24 | 6.10 ± 0.86 | 0.74 ± 0.10 | 6.97 ± 0.41 | 35.07 ± 4.55 | 80.76 ± 40.22 |
| 7 | 10.20 ± 0.16 | 6.18 ± 0.67 | 0.65 ± 0.06 | 6.41 ± 0.73 | 40.38 ± 22.07 | 112.99 ± 79.19 |
| 8 | 10.15 ± 0.19 | 6.12 ± 0.43 | 0.69 ± 0.04 | 6.69 ± 0.55 | 58.68 ± 42.44 | 72.05 ± 49.77 |
| 9 | 10.25 ± 0.30 | 7.10 ± 1.09 | 0.65 ± 0.03 | 6.07 ± 1.5 | 48.89 ± 15.66 | 63.35 ± 38.47 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferretti, M.; Cavani, F.; Roli, L.; Checchi, M.; Magarò, M.S.; Bertacchini, J.; Palumbo, C. Interaction among Calcium Diet Content, PTH (1-34) Treatment and Balance of Bone Homeostasis in Rat Model: The Trabecular Bone as Keystone. Int. J. Mol. Sci. 2019, 20, 753. https://doi.org/10.3390/ijms20030753
Ferretti M, Cavani F, Roli L, Checchi M, Magarò MS, Bertacchini J, Palumbo C. Interaction among Calcium Diet Content, PTH (1-34) Treatment and Balance of Bone Homeostasis in Rat Model: The Trabecular Bone as Keystone. International Journal of Molecular Sciences. 2019; 20(3):753. https://doi.org/10.3390/ijms20030753
Chicago/Turabian StyleFerretti, Marzia, Francesco Cavani, Laura Roli, Marta Checchi, Maria Sara Magarò, Jessika Bertacchini, and Carla Palumbo. 2019. "Interaction among Calcium Diet Content, PTH (1-34) Treatment and Balance of Bone Homeostasis in Rat Model: The Trabecular Bone as Keystone" International Journal of Molecular Sciences 20, no. 3: 753. https://doi.org/10.3390/ijms20030753
APA StyleFerretti, M., Cavani, F., Roli, L., Checchi, M., Magarò, M. S., Bertacchini, J., & Palumbo, C. (2019). Interaction among Calcium Diet Content, PTH (1-34) Treatment and Balance of Bone Homeostasis in Rat Model: The Trabecular Bone as Keystone. International Journal of Molecular Sciences, 20(3), 753. https://doi.org/10.3390/ijms20030753