Improved Anticancer Effect of Recombinant Protein izTRAIL Combined with Sorafenib and Peptide iRGD


 ,
,  , ,
, ,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
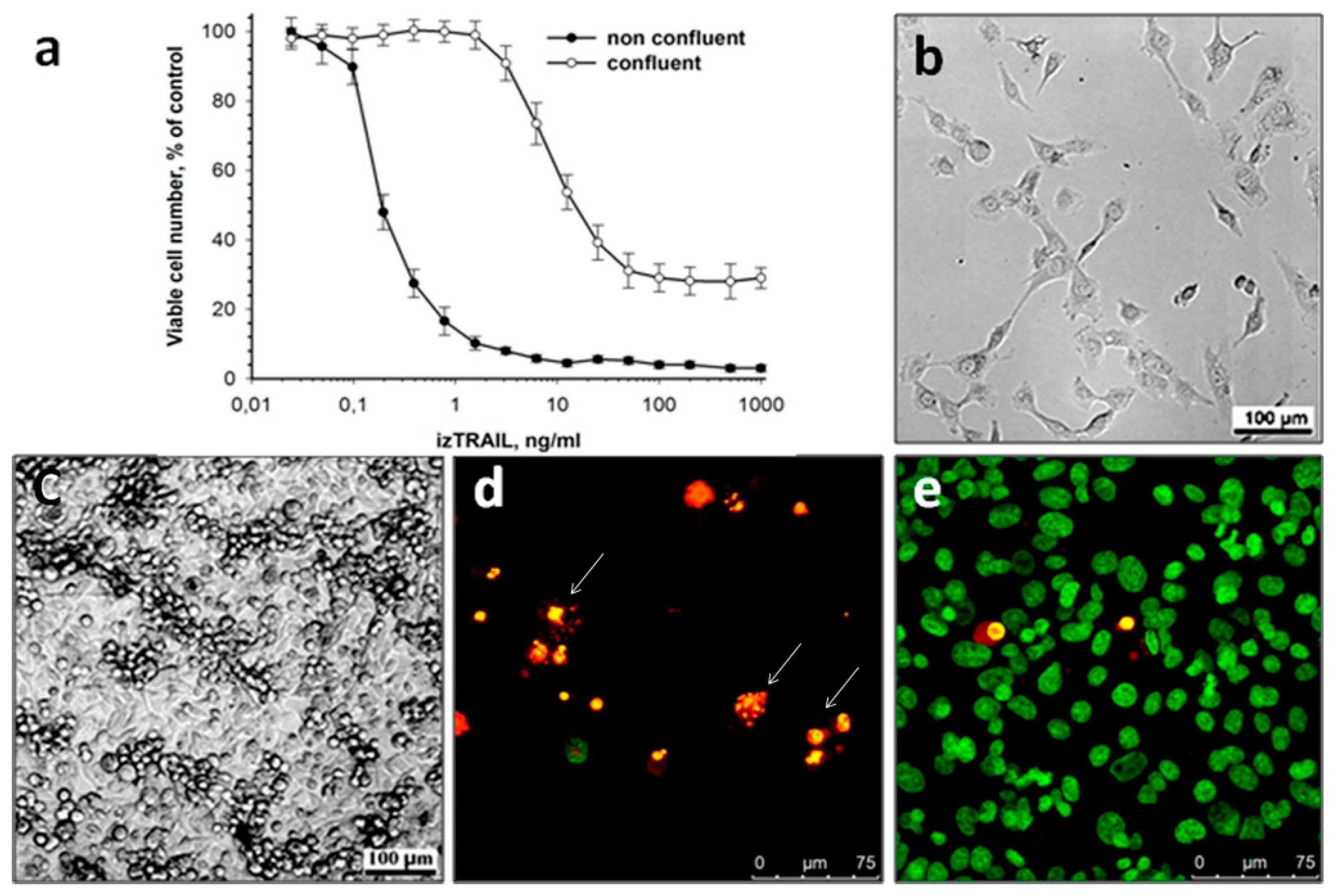
2.1. Increase in Resistance of Cancer Cells HT-1080 to izTRAIL in Confluent Cultures
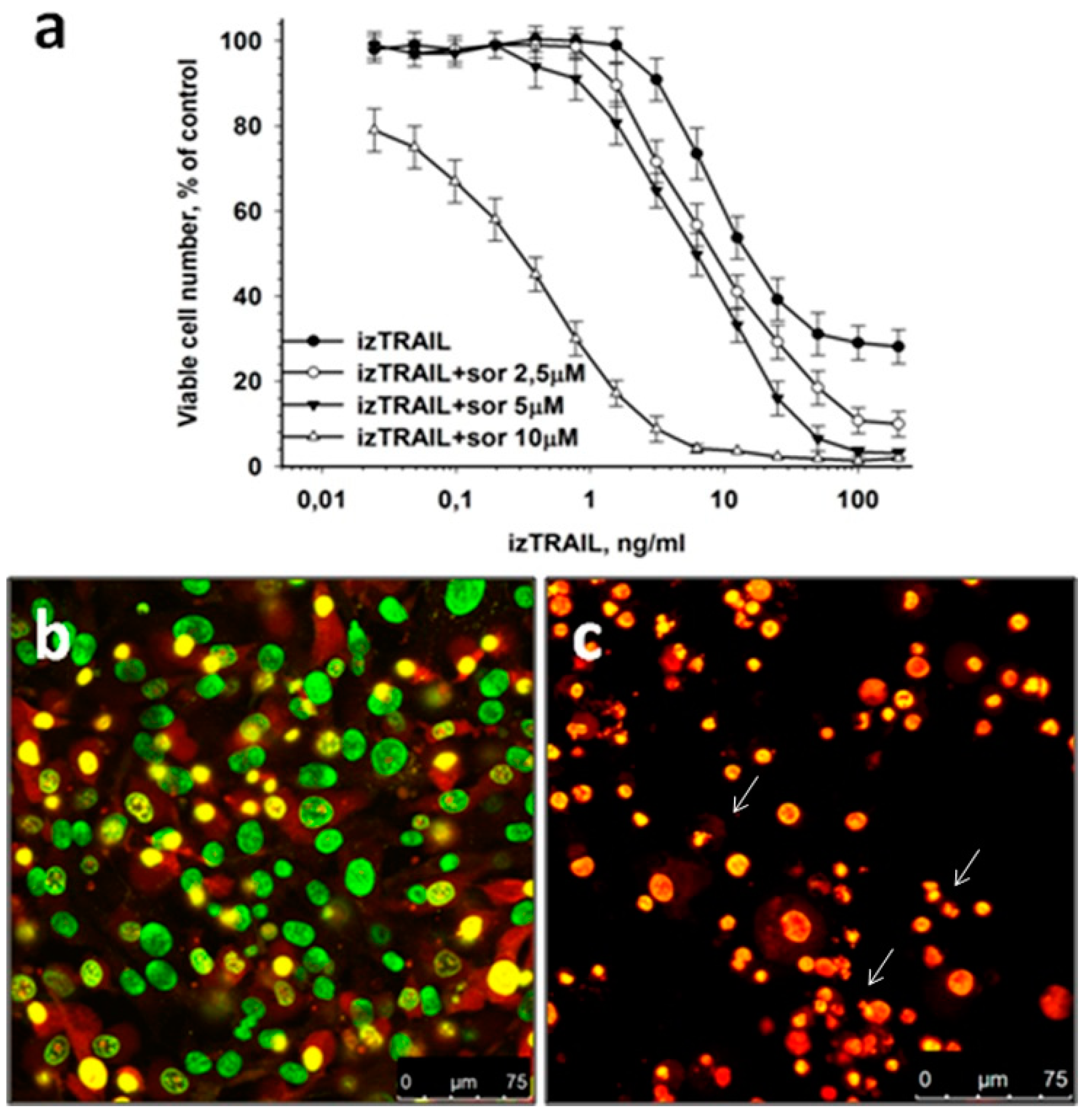
2.2. Suppression of Confluence-Mediated TRAIL Resistance of HT-1080 Cells by Sorafenib
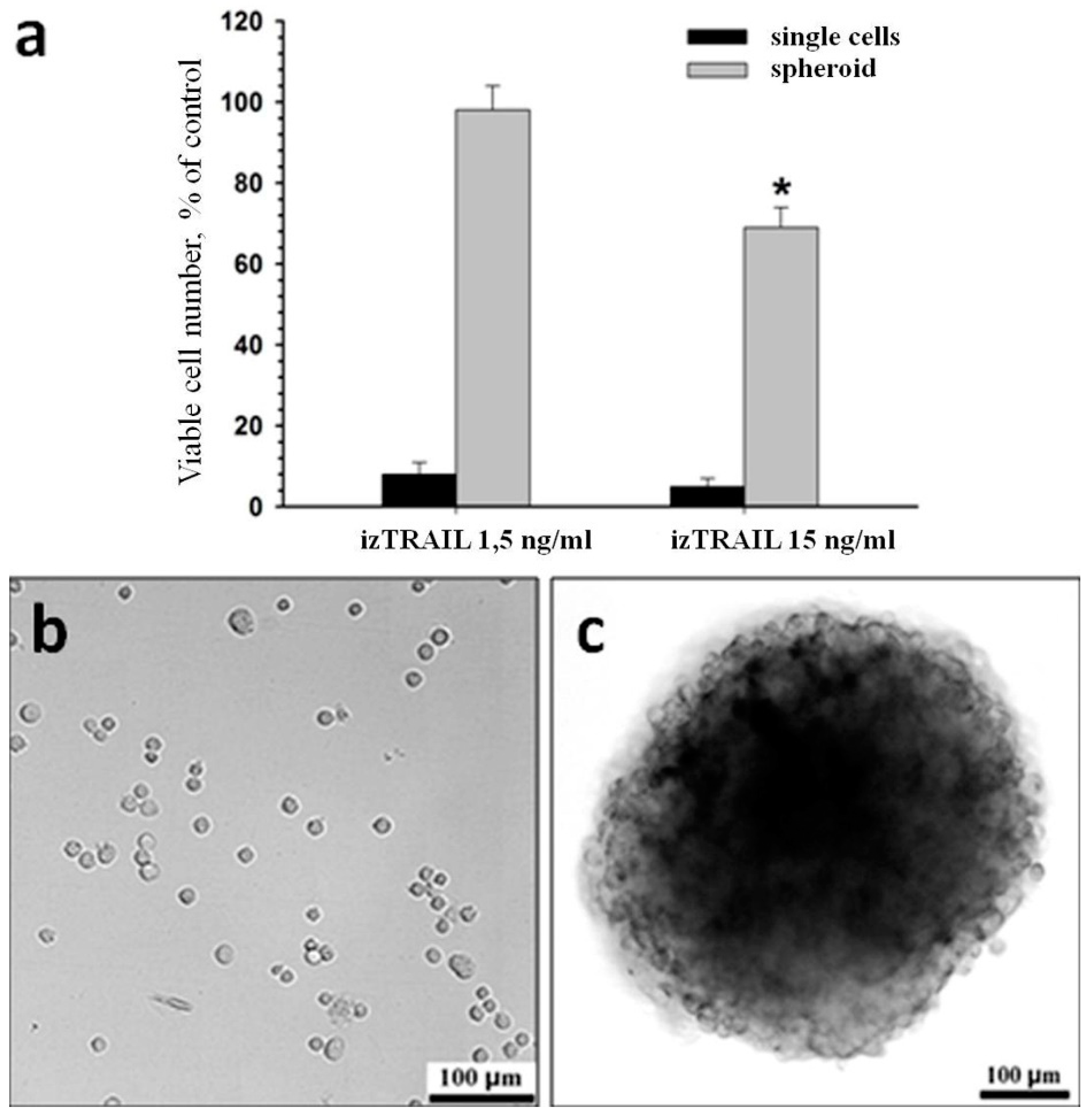
2.3. TRAIL Resistance of HT-1080 Cells Acquired in Spheroids
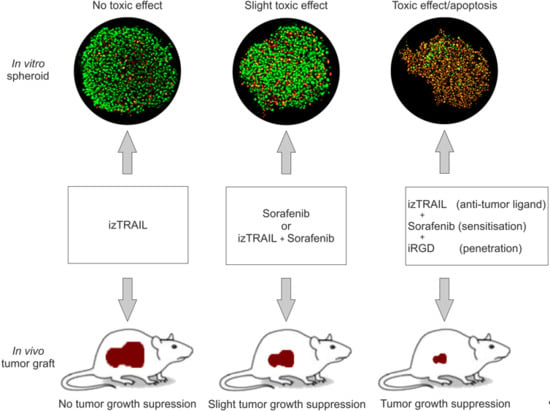
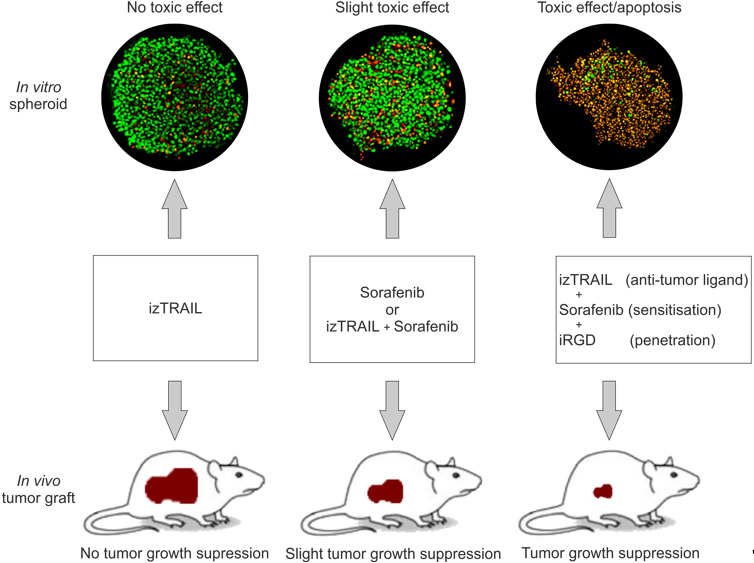
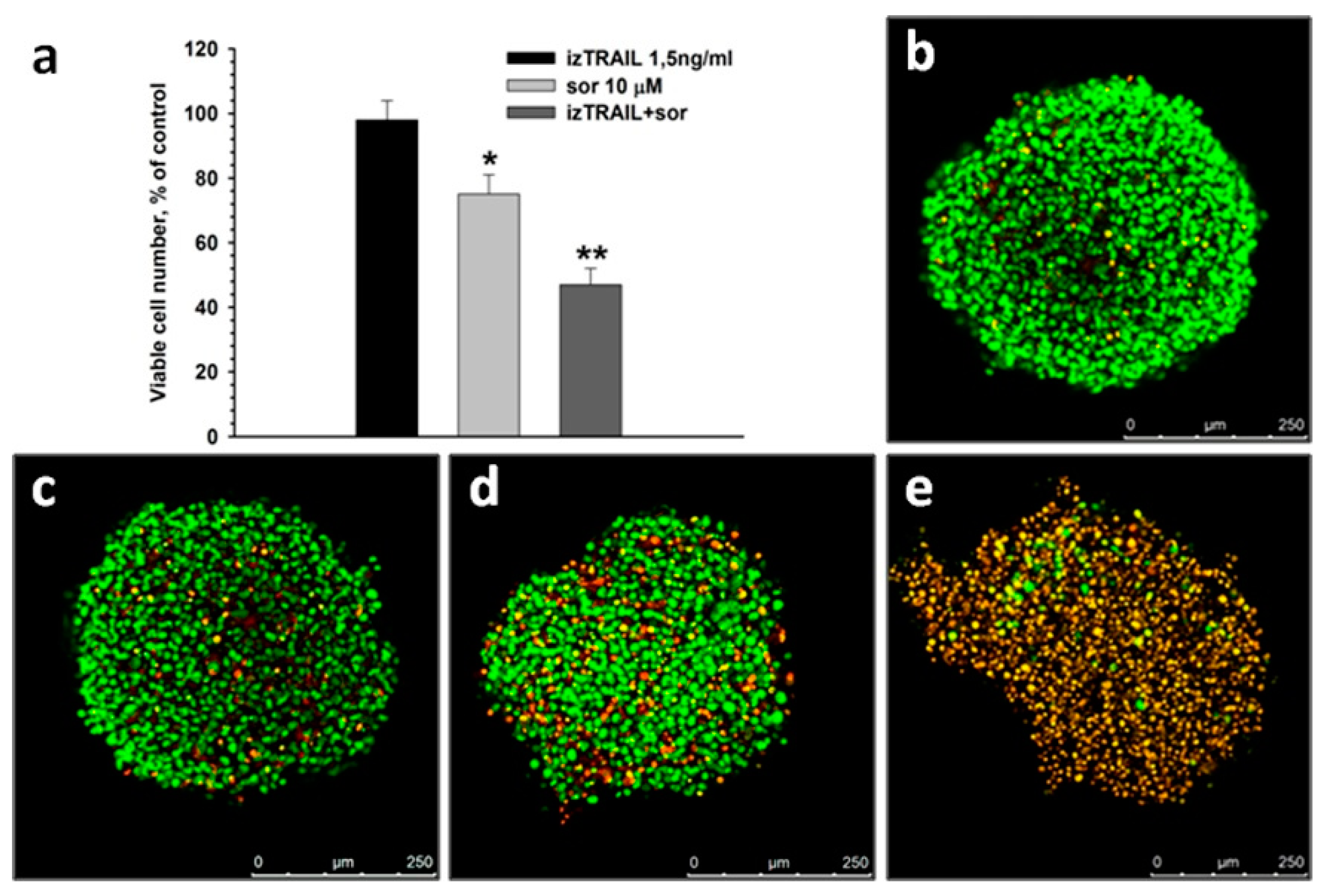
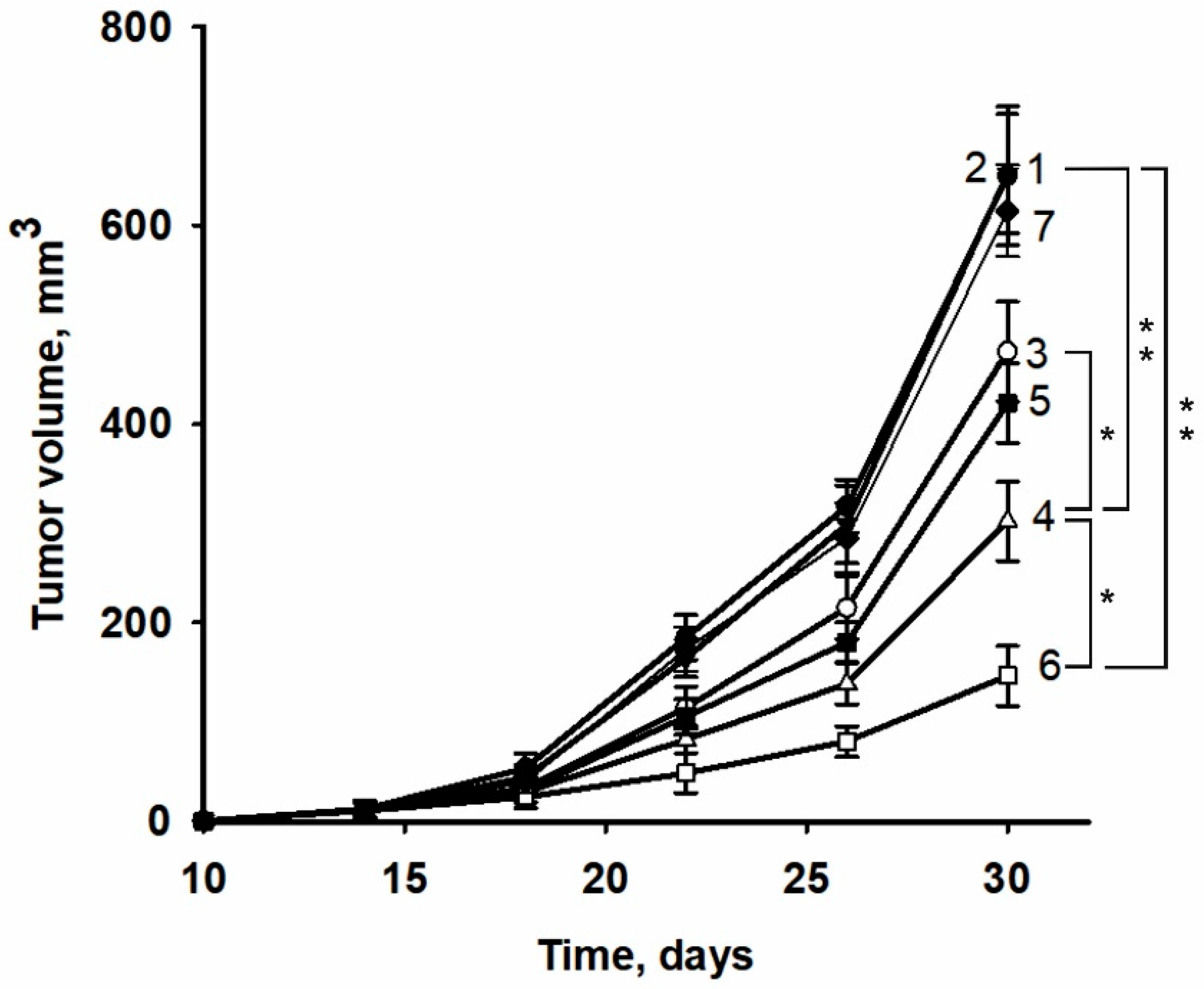
2.4. Improvement of Anticancer Effect of the Recombinant Protein izTRAIL Combined with Sorafenib and the Peptide iRGD
3. Discussion
4. Materials and Methods
4.1. Protocol of izTRAIL Preparation
4.2. Synthesis of iRGD Peptide
4.3. Cell Culture
4.4. Formation of Spheroids
4.5. Cytotoxicity Assay
4.6. Assessment of Apoptotic and Mitotic Cells by Fluorescence Microscopy
4.7. Cancer Xenograft Model and Protocols of Anticancer Treatment
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gonzalvez, F.; Ashkenazi, A. New insights into apoptosis signaling by Apo2L/TRAIL. Oncogene 2010, 29, 4752–4765. [Google Scholar] [CrossRef] [PubMed]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, V.P.; Stylianopoulos, T.; Boucher, Y.; Jain, R.K. Delivery of molecular and nanoscale medicine to tumors: Transport barriers and strategies. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.; Shahrokh, Z.; Marsters, S.; Achilles, K.; Shih, D.; Mounho, B.; Hillan, K.; Totpal, K.; DeForge, L.; Schow, P.; et al. Differential hepatocyte toxicity of recombinant Apo2L/TRAIL versions. Nat. Med. 2001, 7, 383–385. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.W.; Frew, A.J.; Smyth, M.J. The TRAIL apoptotic pathway in cancer onset, progression and therapy. Nat. Rev. Cancer 2008, 8, 782–798. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Pai, R.C.; Fong, S.; Leung, S.; Lawrence, D.A.; Marsters, S.A.; Blackie, C.; Chang, L.; McMurtrey, A.E.; Hebert, A.; et al. Safety and antitumor activity of recombinant soluble Apo2 ligand. J. Clin. Invest. 1999, 104, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Meyer, E.; Debatin, K.M. Inhibition of TRAIL-induced apoptosis by Bcl-2 overexpression. Oncogene 2002, 21, 2283–2294. [Google Scholar] [CrossRef]
- MacFarlane, M. TRAIL-induced signalling and apoptosis. Toxicol. Lett. 2003, 139, 89–97. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; Marshall, J.R.; Messing, J.A.; Hsu, J.W.; King, M.R. TRAIL-mediated apoptosis in breast cancer cells cultured as 3D spheroids. PLoS ONE 2014, 9, e111487. [Google Scholar] [CrossRef]
- Yang, T.M.; Barbone, D.; Fennell, D.A.; Broaddus, V.C. Bcl-2 family proteins contribute to apoptotic resistance in lung cancer multicellular spheroids. Am. J. Respir. Cell Mol. Biol. 2009, 41, 14–23. [Google Scholar] [CrossRef]
- Fadeev, R.S.; Chekanov, A.V.; Dolgikh, N.V.; Akatov, V.S. Multikinase inhibitor sorafenib and HDAC inhibitor suberoylanilide hydroxamic acid suppress confluent resistance of cancer cells to recombinant protein izTRAIL. Biofizika 2012, 57, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, K.N.; Teesalu, T.; Karmali, P.P.; Kotamraju, V.R.; Agemy, L.; Greenwald, D.R.; Ruoslahti, E. Coadministration of a tumor-penetrating peptide enhances the efficacy of cancer drugs. Science 2010, 328, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Teesalu, T.; Sugahara, K.N.; Kotamraju, V.R.; Ruoslahti, E. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration. Proc. Natl. Acad. Sci. USA 2009, 106, 16157–16162. [Google Scholar] [CrossRef] [PubMed]
- Ricci, M.S.; Kim, S.H.; Ogi, K.; Plastaras, J.P.; Ling, J.; Wang, W.; Jin, Z.; Liu, Y.Y.; Dicker, D.T.; Chiao, P.J.; et al. Reduction of TRAIL-induced Mcl-1 and cIAP2 by c-Myc or sorafenib sensitizes resistant human cancer cells to TRAIL-induced death. Cancer Cell 2007, 12, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.U.; Wilson, S.M.; Abayasiriwardana, K.S.; Collins, R.; Fjellbirkeland, L.; Xu, Z.; Jablons, D.M.; Nishimura, S.L.; Broaddus, V.C. A novel in vitro model of human mesothelioma for studying tumor biology and apoptotic resistance. Am. J. Respir. Cell Mol. Biol. 2005, 33, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Transport of molecules, particles, and cells in solid tumors. Annu. Rev. Biomed. Eng. 1999, 1, 241–263. [Google Scholar] [CrossRef]
- Hambley, T.W.; Hait, W.N. Is anticancer drug development heading in the right direction? Cancer Res. 2009, 69, 1259–1262. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, J.; Ji, Y.; Lu, J.; Chan, R.; Meng, H. Targeted drug delivery using iRGD peptide for solid cancer treatment. Mol. Syst. Des. Eng. 2017, 4, 370–379. [Google Scholar] [CrossRef]
- Hu, C.; Chen, X.; Huang, Y.; Che, Y. Co-administration of iRGD with peptide HPRP-A1 to improve anticancer activity and membrane penetrability. Sci. Rep. 2018, 8, 2274. [Google Scholar] [CrossRef]
- Suzuki, K.; Hiroaki, H.; Kohda, D.; Tanaka, T. An isoleucine zipper peptide forms a native-like triple stranded coiled coil in solution. Protein Eng. 1998, 11, 1051–1055. [Google Scholar] [CrossRef]
- Ganten, T.M.; Koschny, R.; Sykora, J.; Schulze-Bergkamen, H.; Buchler, P.; Haas, T.L.; Schader, M.B.; Untergasser, A.; Stremmel, W.; Walczak, H. Preclinical differentiation between apparently safe and potentially hepatotoxic applications of TRAIL either alone or in combination with chemotherapeutic drugs. Clin. Cancer Res. 2006, 12, 2640–2646. [Google Scholar] [CrossRef] [PubMed]
- Rodionov, I.L.; Baru, M.B.; Ivanov, V.T. A swellographic approach to monitoring continuous-flow solid-phase peptide synthesis. Pept. Res. 1992, 5, 119–125. [Google Scholar] [PubMed]
- Rodionov, I.L.; Peshenko, I.A.; Baidakova, L.K.; Ivanov, V.T. Swellographic study of peptide resin swelling behavior during solid phase peptide synthesis. Int. J. Pept. Res. Ther. 2007, 13, 161–171. [Google Scholar] [CrossRef]
- Stewart, J.; Young, J. Solid Phase Peptide Synthesis; Academic Press: San Diego, IL, USA, 1984. [Google Scholar]
- Friedrich, J.; Seidel, C.; Ebner, R.; Kunz-Schughart, L.A. Spheroid-based drug screen: Considerations and practical approach. Nat. Protoc. 2009, 4, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Saotome, K.; Morita, H.; Umeda, M. Cytotoxicity test with simplified crystal violet staining method using microtitre plates and its application to injection drugs. Toxicol. In Vitro 1989, 3, 317–321. [Google Scholar] [CrossRef]
- Solovieva, M.E.; Solovyev, V.V.; Kudryavtsev, A.A.; Trizna, Y.A.; Akatov, V.S. Vitamin B12b enhances the cytotoxicity of dithiothreitol. Free Radic. Biol. Med. 2008, 44, 1846–1856. [Google Scholar] [CrossRef]
- Akatov, V.S.; Korystov, Y.N.; Kublik, L.N.; Levitman, M.K.; Shaposhnikova, V.V.; Lezhnev, E.L. Antitumor effect of acute hyperoxia. Exp. Oncol. 2001, 23, 123–125. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fadeev, R.; Chekanov, A.; Solovieva, M.; Bezborodova, O.; Nemtsova, E.; Dolgikh, N.; Fadeeva, I.; Senotov, A.; Kobyakova, M.; Evstratova, Y.; et al. Improved Anticancer Effect of Recombinant Protein izTRAIL Combined with Sorafenib and Peptide iRGD. Int. J. Mol. Sci. 2019, 20, 525. https://doi.org/10.3390/ijms20030525
Fadeev R, Chekanov A, Solovieva M, Bezborodova O, Nemtsova E, Dolgikh N, Fadeeva I, Senotov A, Kobyakova M, Evstratova Y, et al. Improved Anticancer Effect of Recombinant Protein izTRAIL Combined with Sorafenib and Peptide iRGD. International Journal of Molecular Sciences. 2019; 20(3):525. https://doi.org/10.3390/ijms20030525
Chicago/Turabian StyleFadeev, Roman, Alexey Chekanov, Marina Solovieva, Olga Bezborodova, Elena Nemtsova, Nadezda Dolgikh, Irina Fadeeva, Anatoly Senotov, Margarita Kobyakova, Yana Evstratova, and et al. 2019. "Improved Anticancer Effect of Recombinant Protein izTRAIL Combined with Sorafenib and Peptide iRGD" International Journal of Molecular Sciences 20, no. 3: 525. https://doi.org/10.3390/ijms20030525
APA StyleFadeev, R., Chekanov, A., Solovieva, M., Bezborodova, O., Nemtsova, E., Dolgikh, N., Fadeeva, I., Senotov, A., Kobyakova, M., Evstratova, Y., Yakubovskaya, R., & Akatov, V. (2019). Improved Anticancer Effect of Recombinant Protein izTRAIL Combined with Sorafenib and Peptide iRGD. International Journal of Molecular Sciences, 20(3), 525. https://doi.org/10.3390/ijms20030525