Calcium Polyphosphate Nanoparticles Act as an Effective Inorganic Phosphate Source during Osteogenic Differentiation of Human Mesenchymal Stem Cells

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
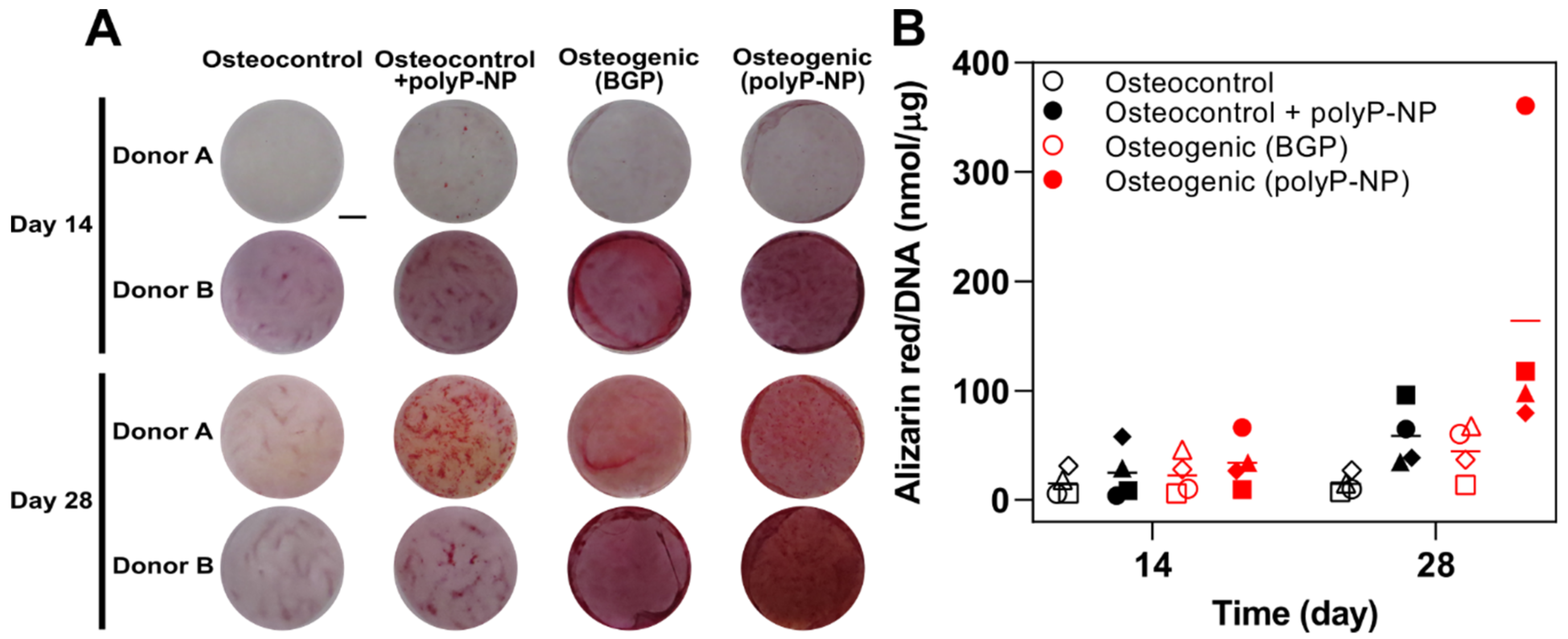
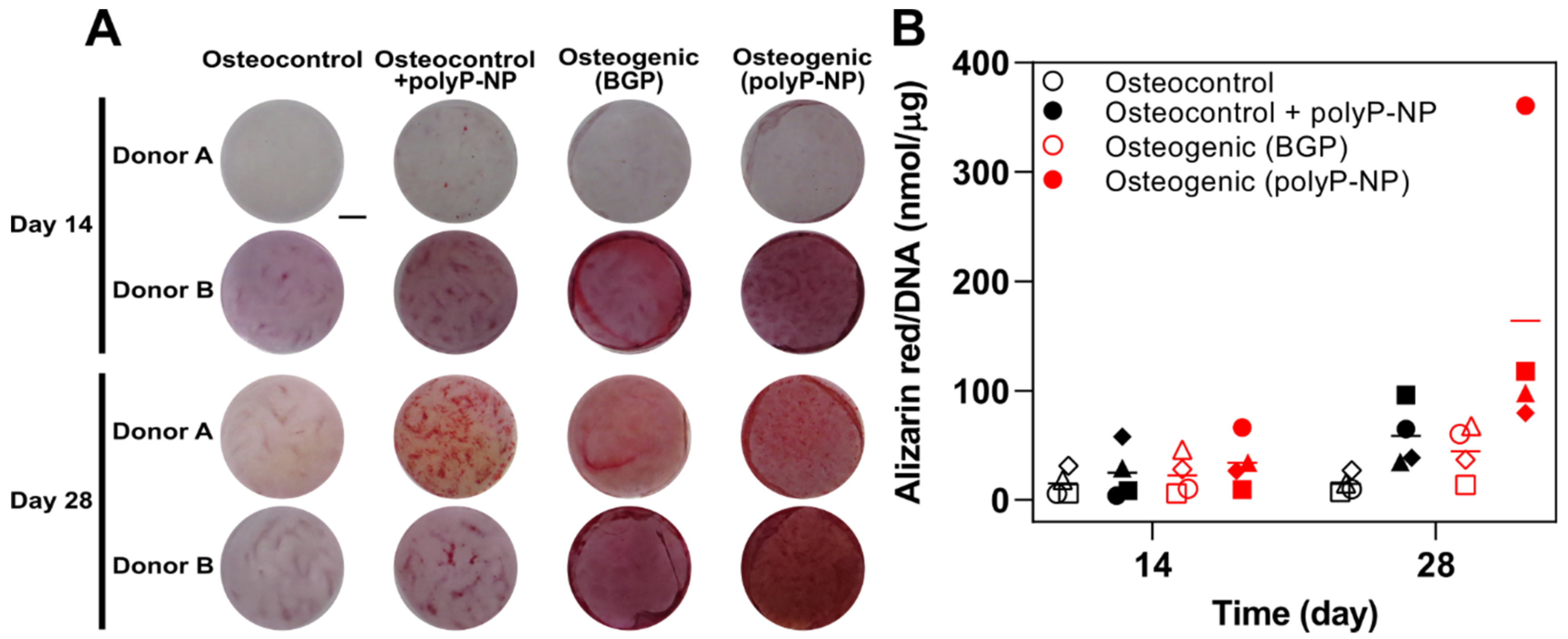
2.1. Inorganic PolyP-NP can Function As an Effective phosphate Source for Matrix Mineralisation during Osteogenic Differentiation of Human BM-MSCs
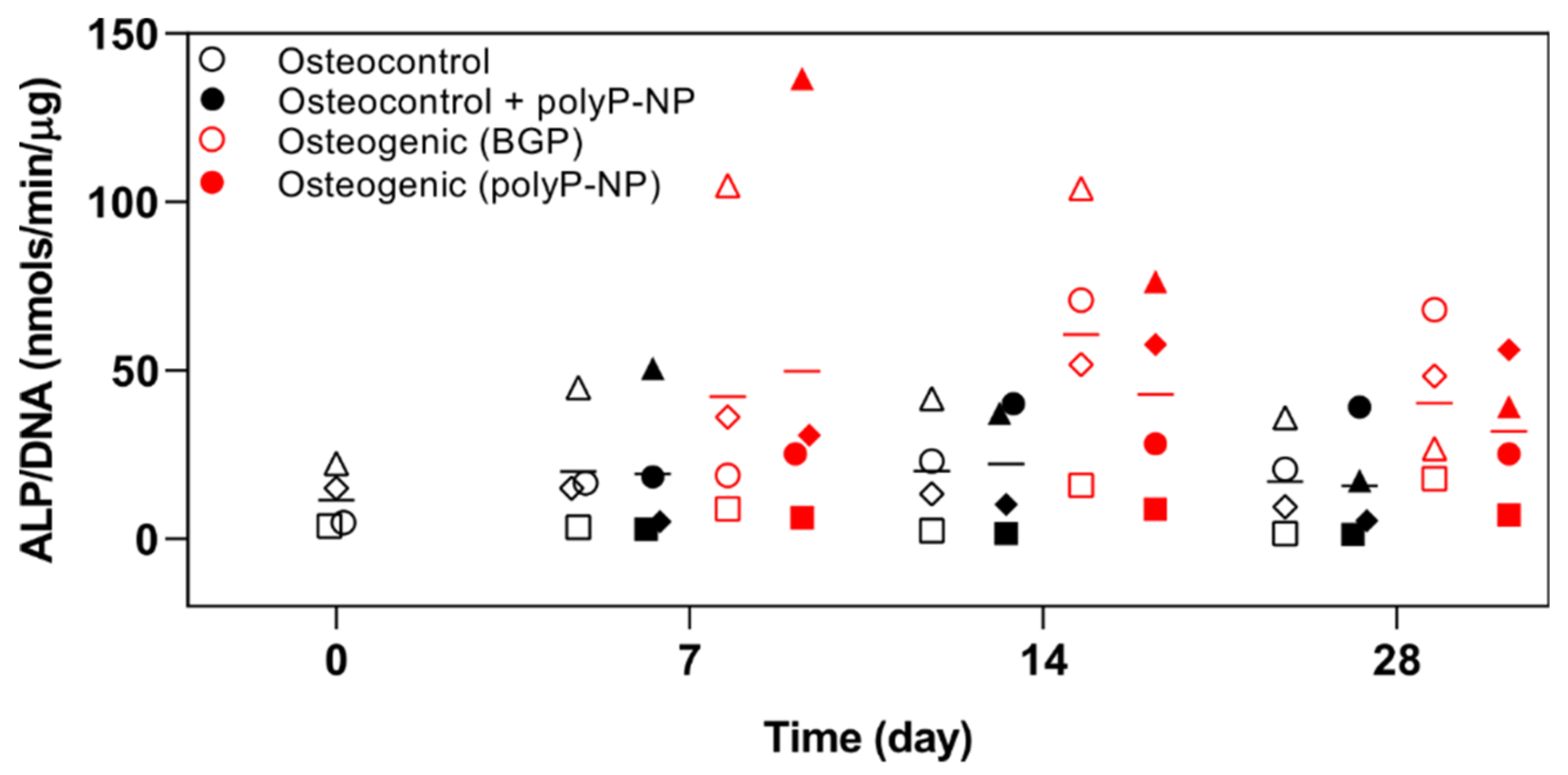
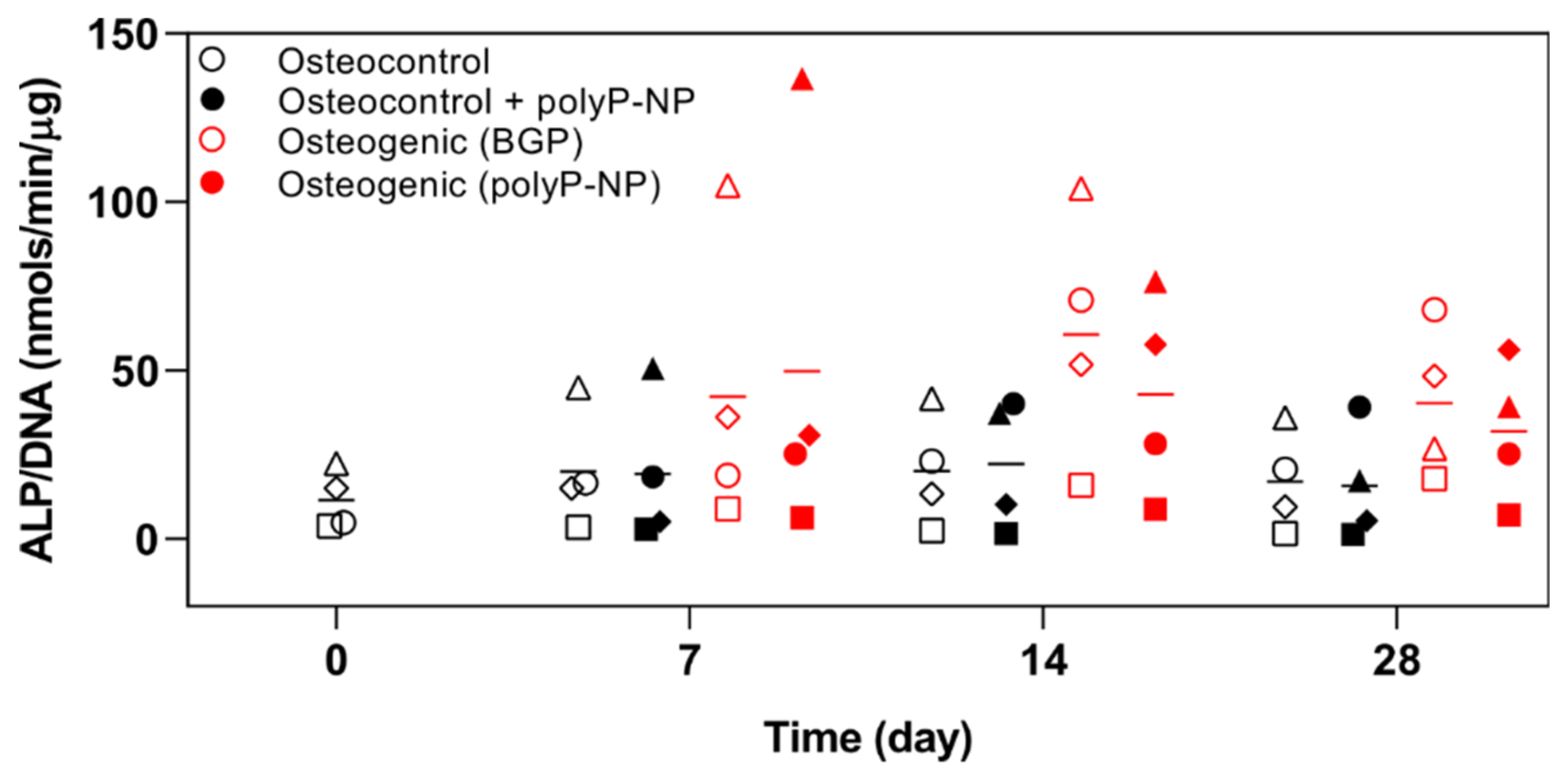
2.2. PolyP-NP Treatment Results in Comparable Effects on ALP Activity to BGP during Osteogenic Differentiation of hBM-MSCs
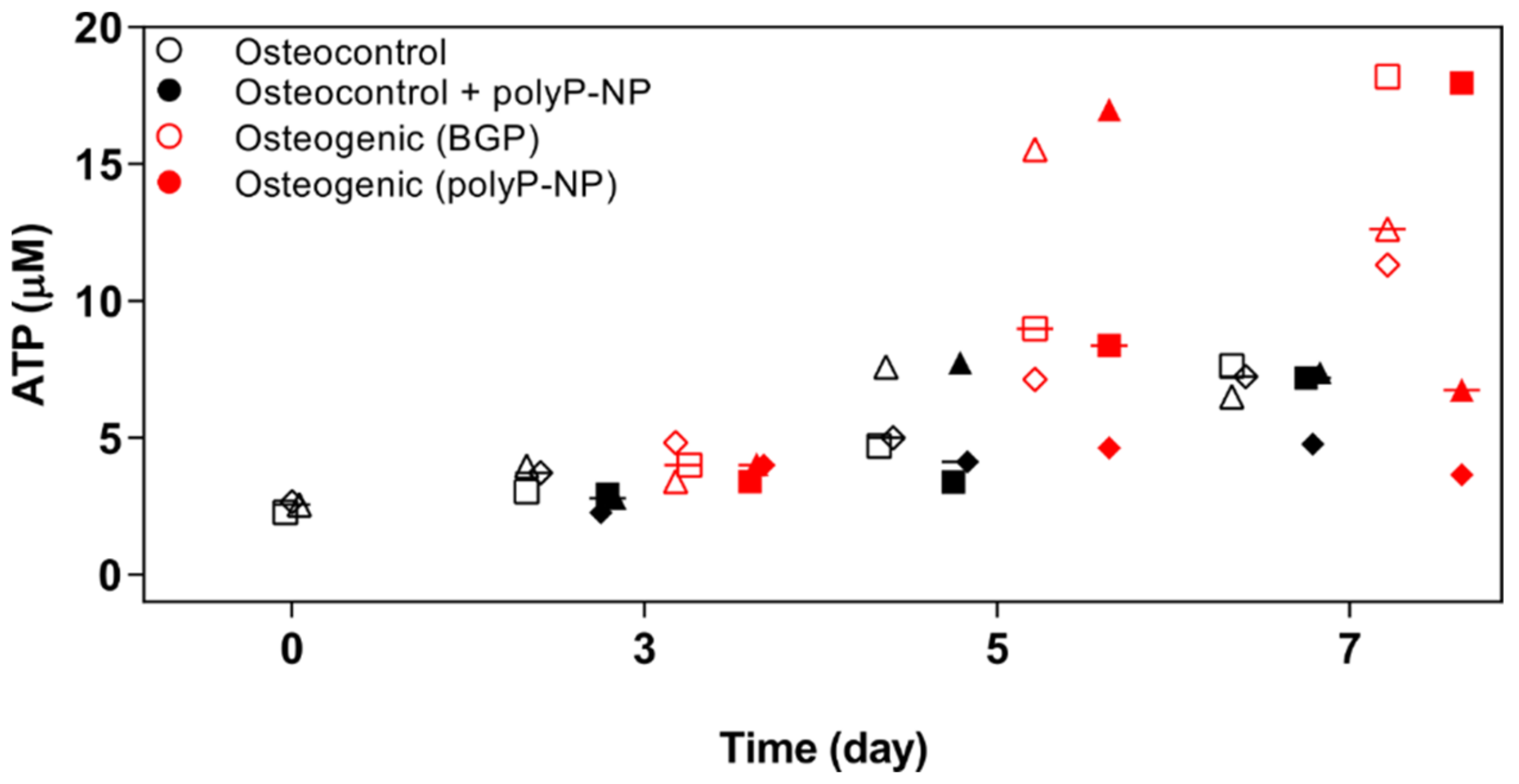
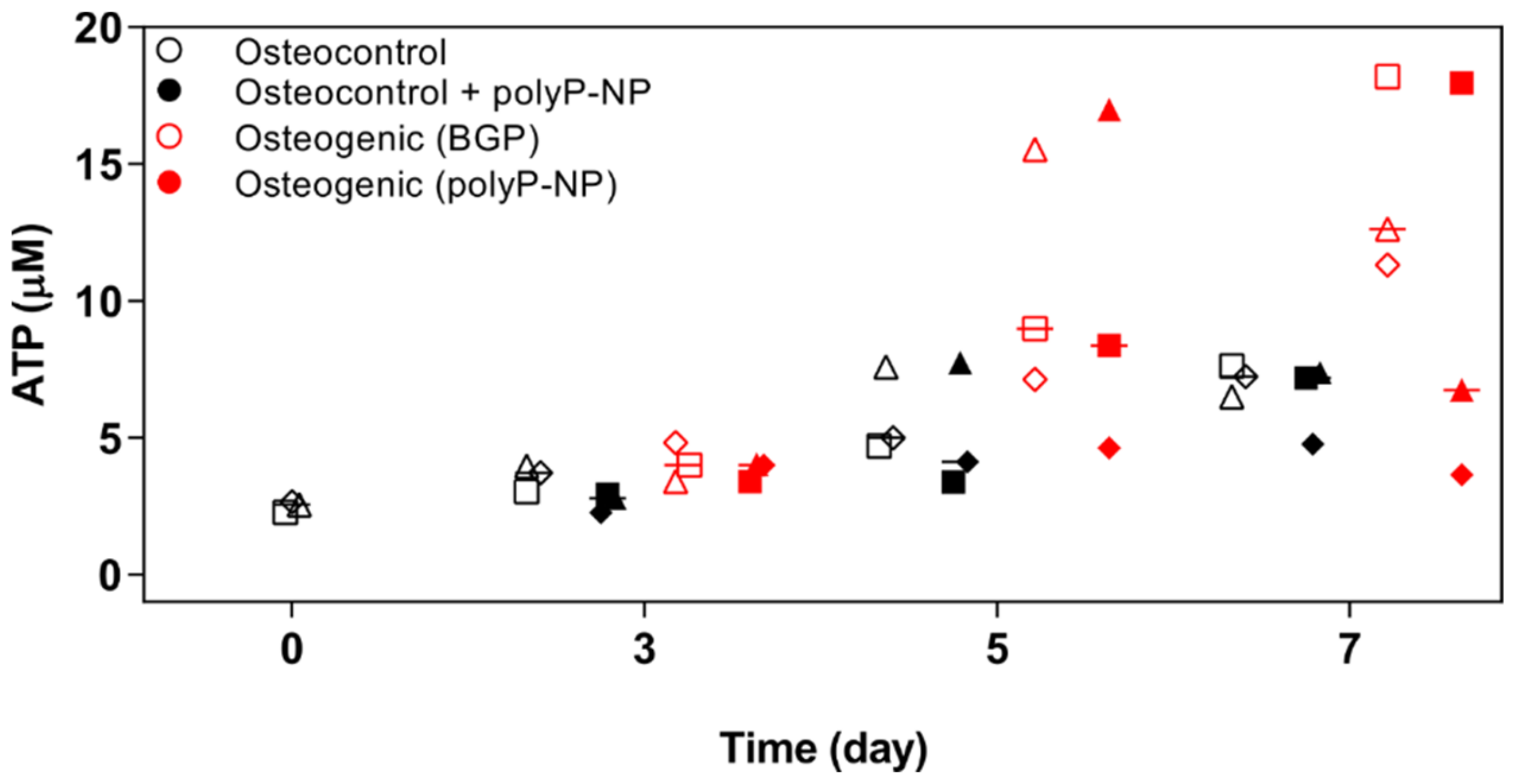
2.3. Osteogenic Differentiation of BM-MSCs, Supported by Either BGPor polyP-NP, Is Associated with Increased Intracellular Levels of ATP at Early TIME Points
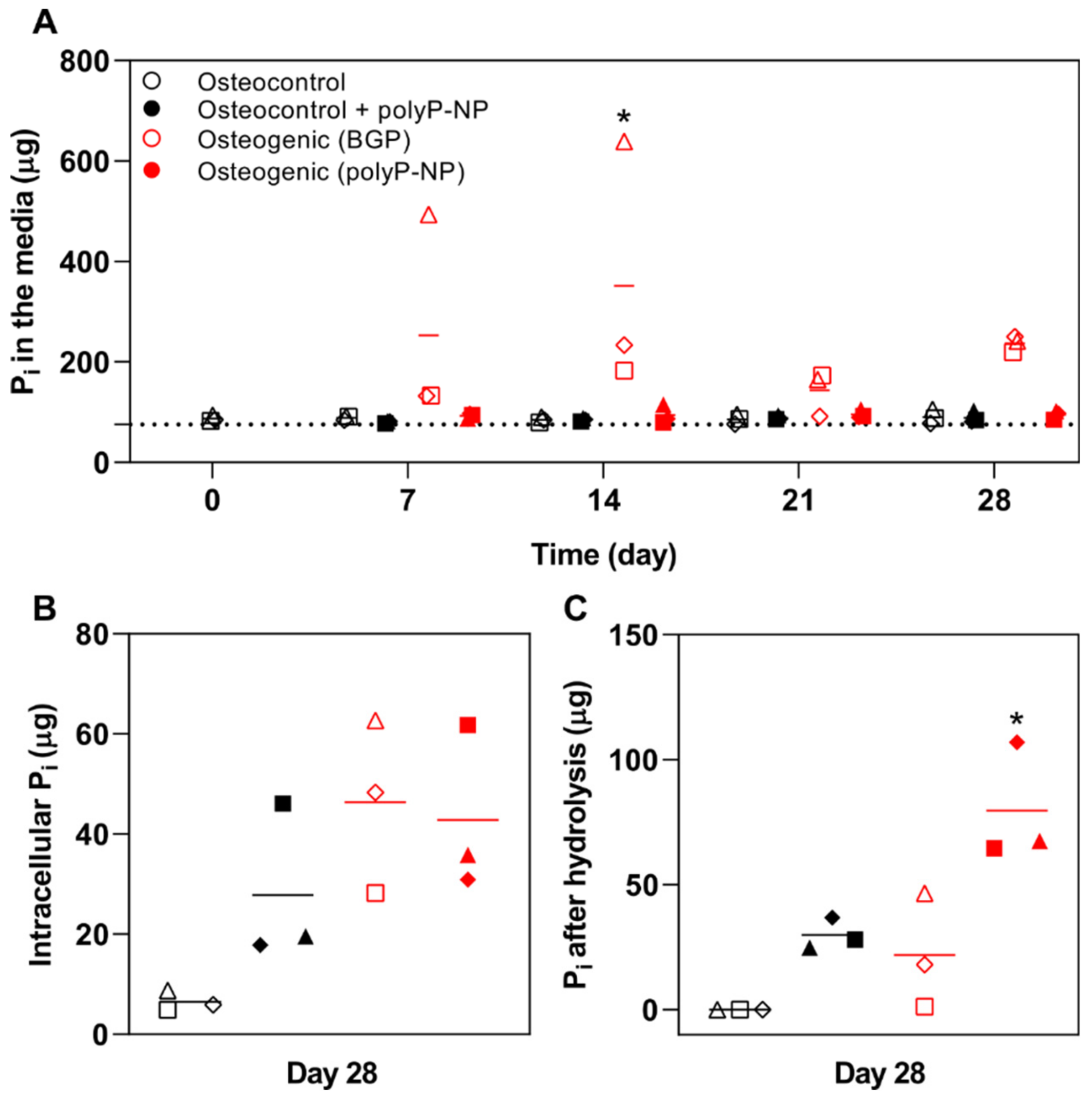
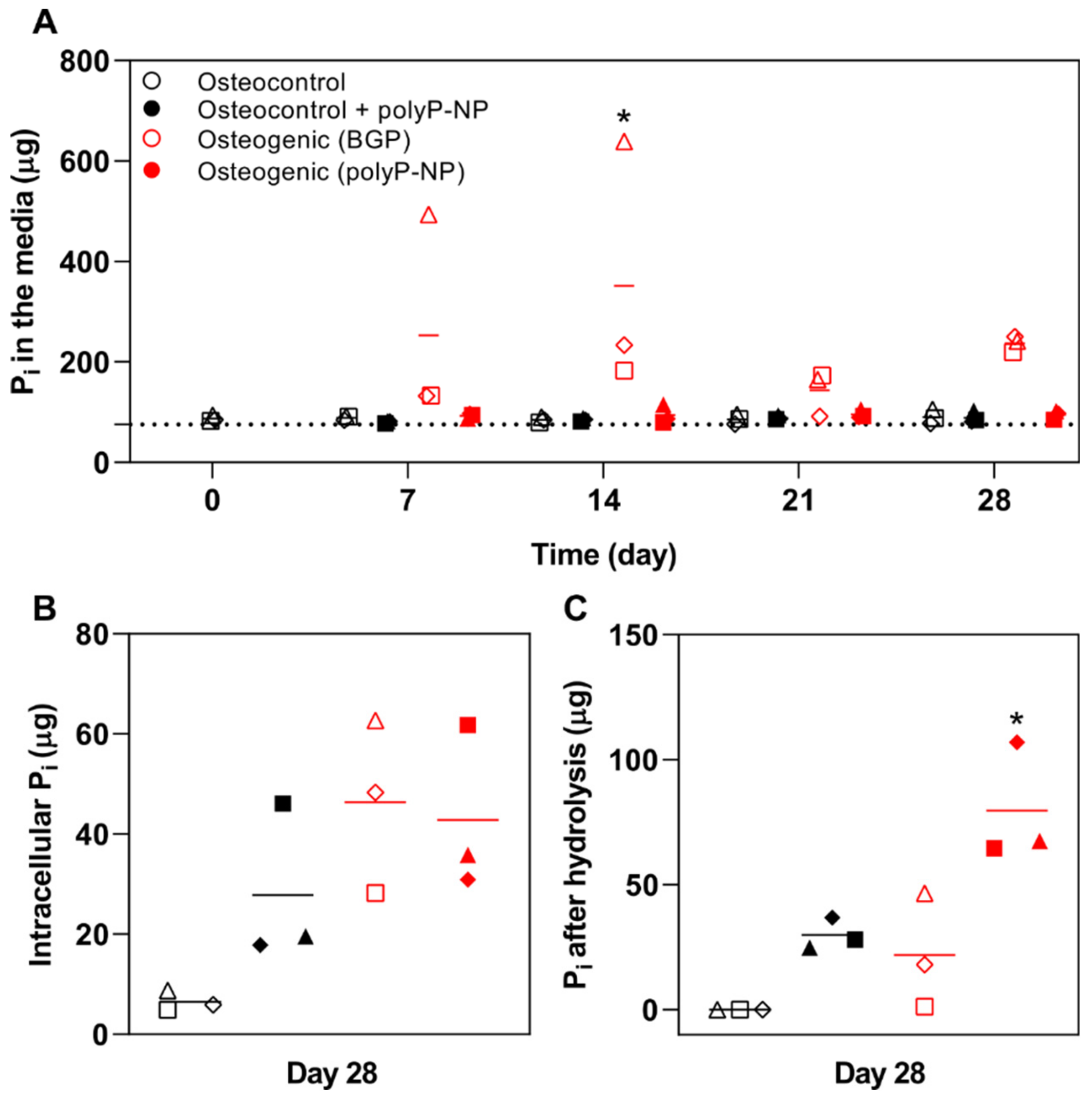
2.4. BGP Results in High Free Phosphate Fluctuations in the Medium
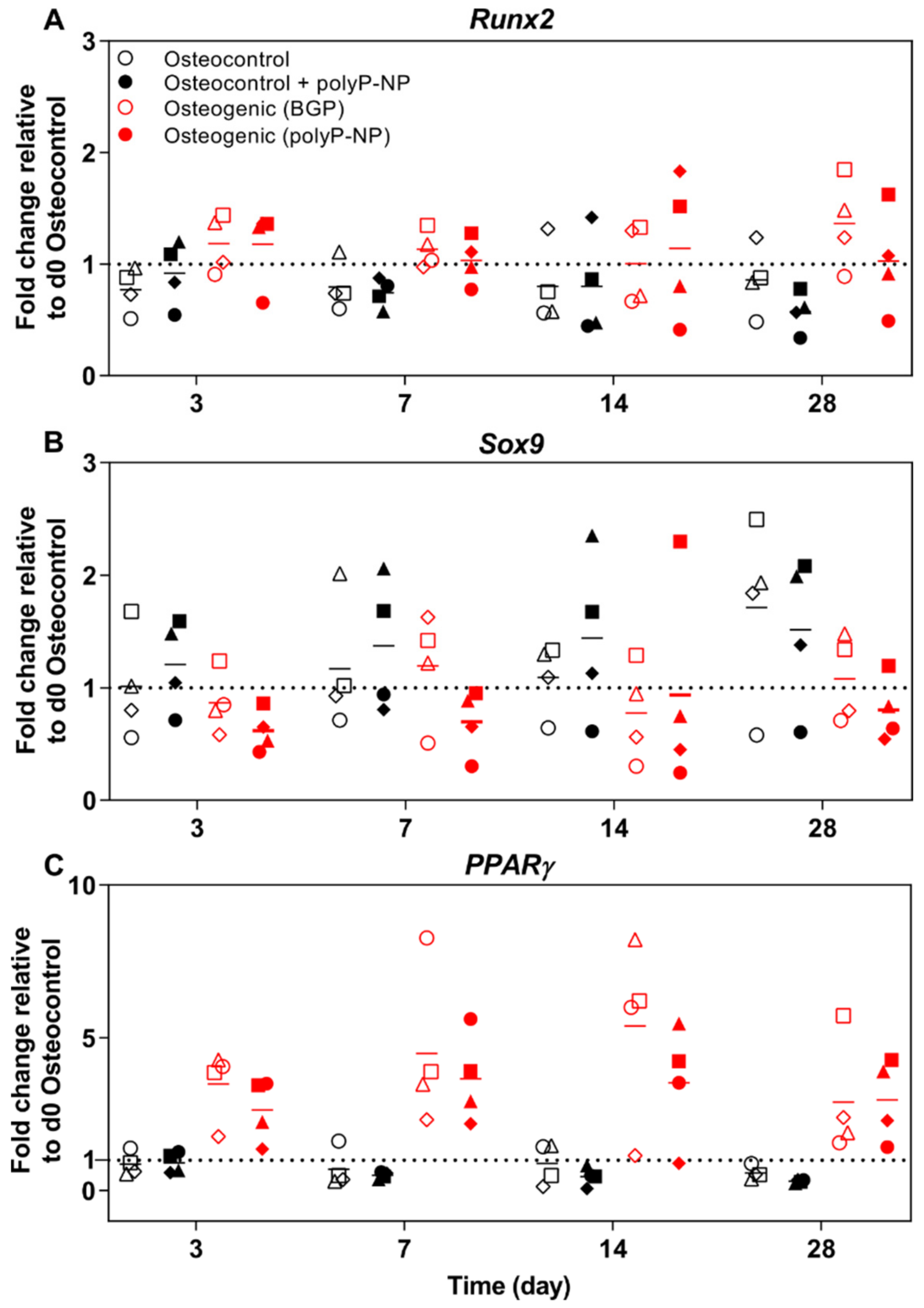
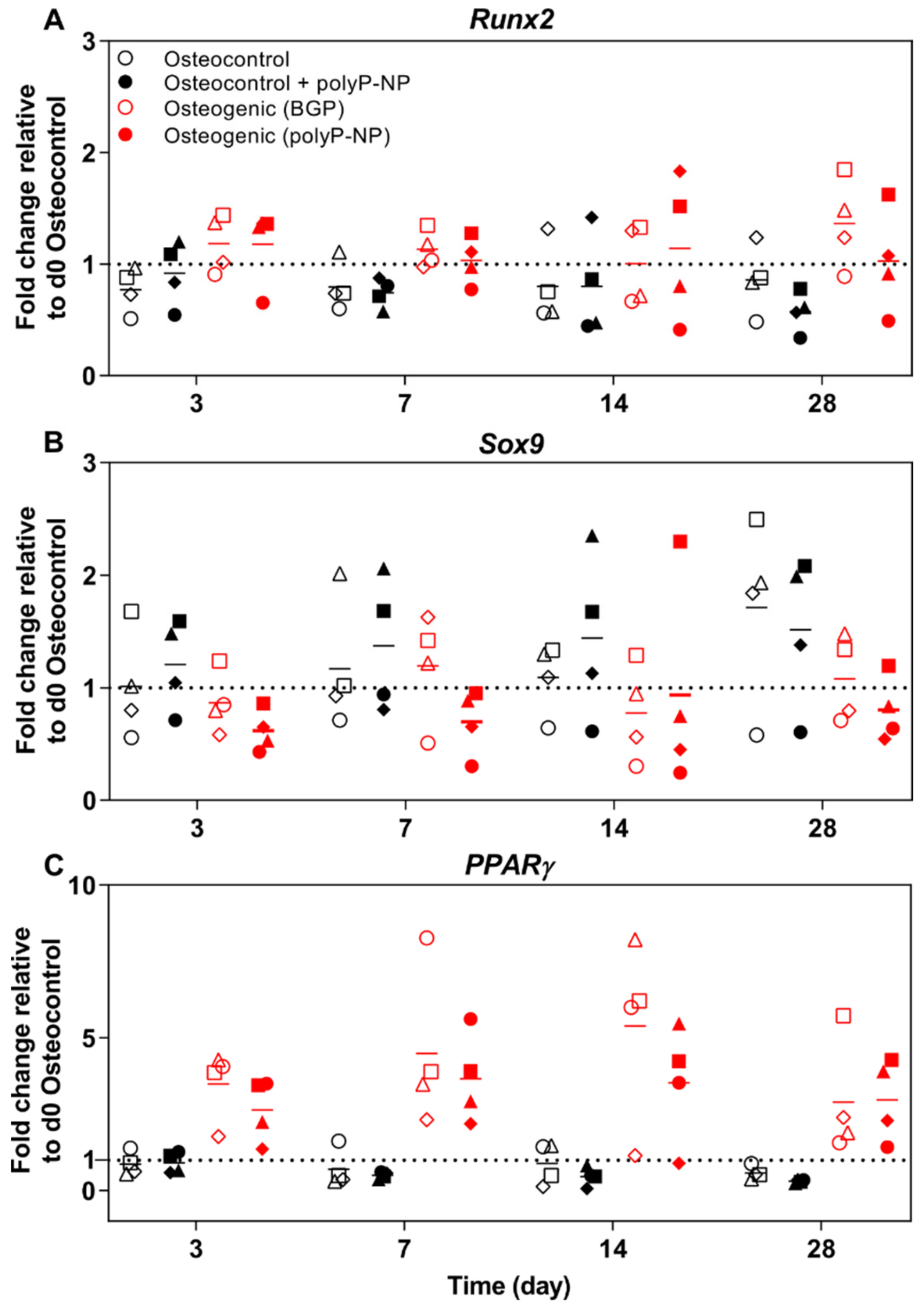
2.5. PolyP-NP Induce Similar Changes to BGP-Containing Osteogenic Media in the Expression of BM-MSC-Relevant Transcription Factors
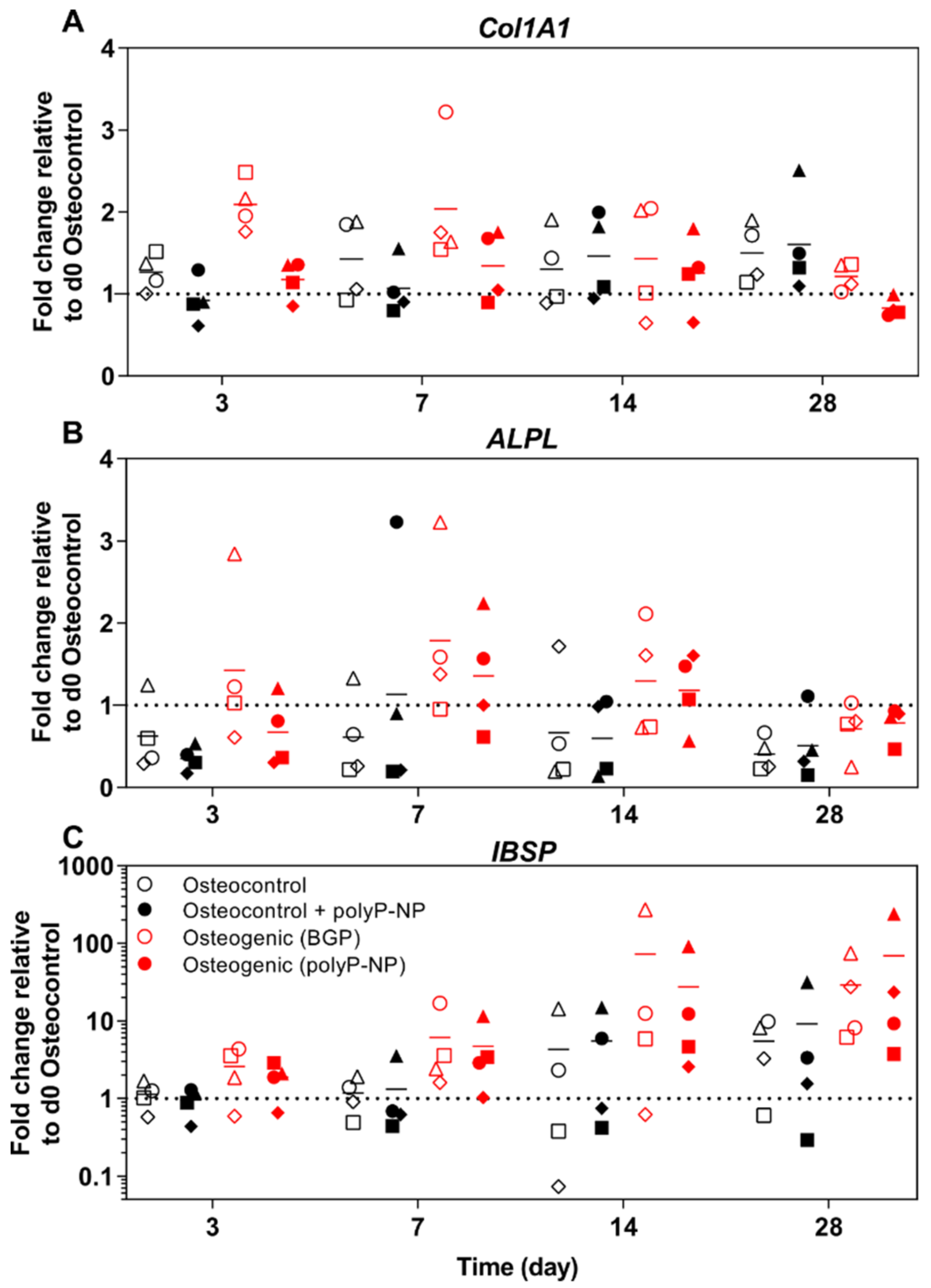
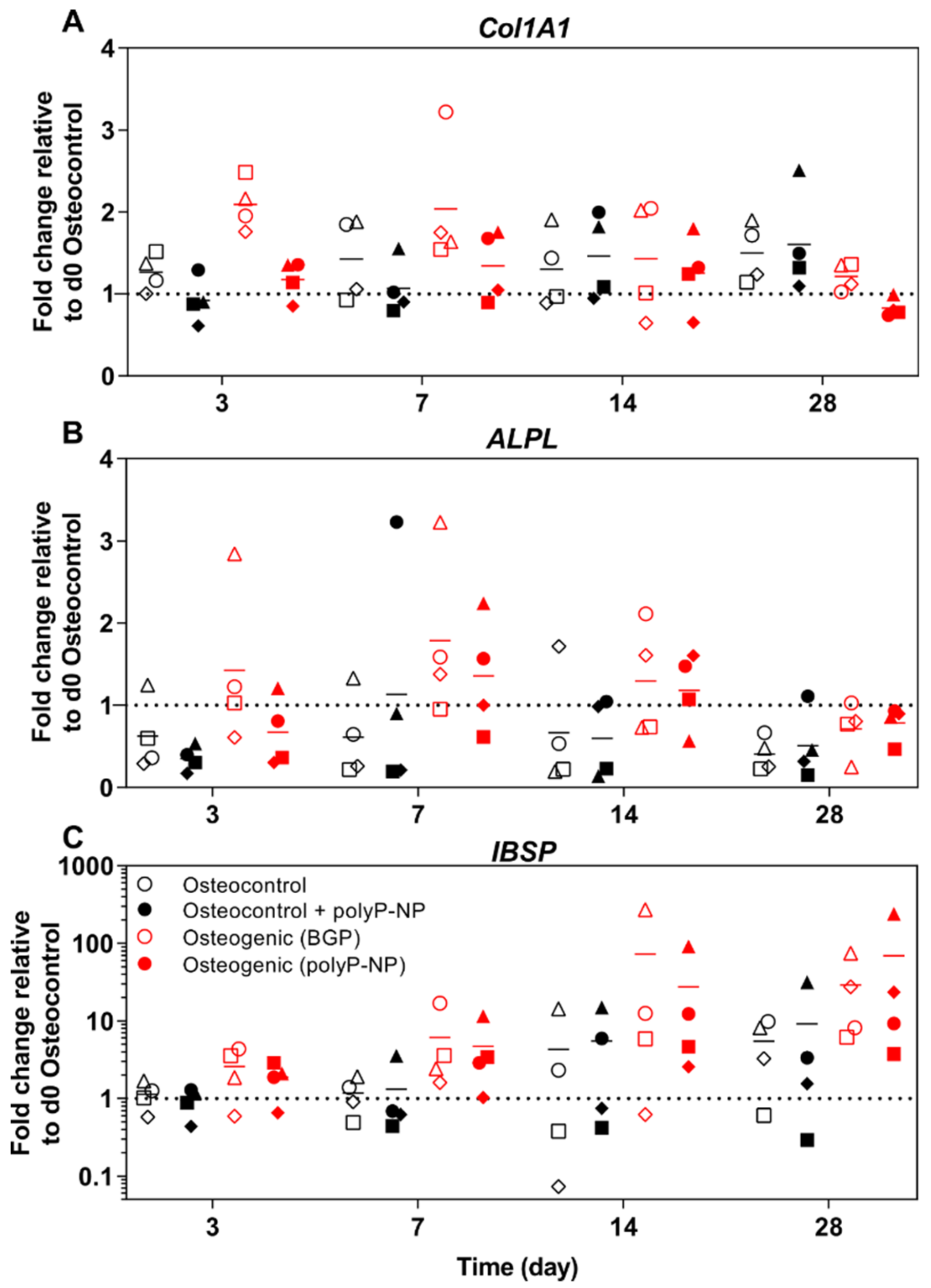
2.6. PolyP-NP-Supplementation of Ostegenic Medium Supports Expression of Early, Intermediate and Late Osteogenic Markers Comparable to BGP-Supplemented Osteogenic Media
3. Discussion
4. Materials and Methods
4.1. Preparation of Calcium-PolyP-NP
4.2. Cell Isolation and Culture
4.3. Osteogenic Differentiation
4.4. RNA Isolation and qRT-PCR
4.5. Alizarin Red Staining and Quantification
4.6. Quantification of ALP Activity and DNA Content
4.7. Quantification of ATP Content
4.8. Phosphate Quantification
4.9. Statistics
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA2P | L-ascorbic acid 2-phosphate sesquimagnesium salt hydrate |
| ALP | Alkaline phosphatase |
| ANOVA | Analysis of variance |
| ATP | Adenosine triphosphate |
| BCP | 1-Bromo-3-chloropropane |
| BGP | Β-Glycerolphosphate disodium salt hydrate |
| BSP | Bone sialoprotein |
| Dexa | Dexamethasone |
| FBS | Fetal bovine serum |
| hBM-MSCs | Human bone marrow-derived mesenchymal stem cells |
| HUVEC | Human umbilical vein endothelial cells |
| LG-DMEM | Low glucose (1 g/L) Dulbecco’s Modified Eagle’s Medium |
| PBS | Phosphate-buffered saline |
| PEN/STREP | Penicillin/Streptomycin |
| Pi | Free phosphate |
| polyP-NP | Polyphosphate nanoparticles |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| qRT-PCR | Quantitative reverse transcription PCR |
| Runx2 | Runt-related transcription factor 2 |
| Sox9 | Sex determining region Y box 9 |
| TBS | Tris buffered saline |
References
- Jaiswal, N.; Haynesworth, S.E.; Caplan, A.I.; Bruder, S.P. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J. Cell Biochem. 1997, 64, 295–312. [Google Scholar] [CrossRef]
- Schack, L.M.; Noack, S.; Winkler, R.; Wissmann, G.; Behrens, P.; Wellmann, M.; Jagodzinski, M.; Krettek, C.; Hoffmann, A. The phosphate source influences gene expression and quality of mineralization during in vitro osteogenic differentiation of human mesenchymal stem cells. PLoS ONE 2013, 8, e65943. [Google Scholar] [CrossRef] [PubMed]
- Cassella, J.P.; Garrington, N.; Stamp, T.C.; Ali, S.Y. An electron probe X-ray microanalytical study of bone mineral in osteogenesis imperfecta. Calcif. Tissue Int. 1995, 56, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Bellows, C.G.; Heersche, J.N.; Aubin, J.E. Inorganic phosphate added exogenously or released from beta-glycerophosphate initiates mineralization of osteoid nodules in vitro. Bone Miner. 1992, 17, 15–29. [Google Scholar] [CrossRef]
- Gronowicz, G.; Woodiel, F.N.; McCarthy, M.B.; Raisz, L.G. In vitro mineralization of fetal rat parietal bones in defined serum-free medium: Effect of beta-glycerol phosphate. J. Bone Miner. Res. 1989, 4, 313–324. [Google Scholar] [CrossRef]
- Beck, G.R., Jr. Inorganic phosphate as a signaling molecule in osteoblast differentiation. J. Cell Biochem. 2003, 90, 234–243. [Google Scholar] [CrossRef]
- Foster, B.L.; Nociti, F.H., Jr.; Swanson, E.C.; Matsa-Dunn, D.; Berry, J.E.; Cupp, C.J.; Zhang, P.; Somerman, M.J. Regulation of cementoblast gene expression by inorganic phosphate in vitro. Calcif. Tissue Int. 2006, 78, 103–112. [Google Scholar] [CrossRef]
- Fatherazi, S.; Matsa-Dunn, D.; Foster, B.L.; Rutherford, R.B.; Somerman, M.J.; Presland, R.B. Phosphate regulates osteopontin gene transcription. J. Dent. Res. 2009, 88, 39–44. [Google Scholar] [CrossRef]
- Tada, H.; Nemoto, E.; Foster, B.L.; Somerman, M.J.; Shimauchi, H. Phosphate increases bone morphogenetic protein-2 expression through cAMP-dependent protein kinase and ERK1/2 pathways in human dental pulp cells. Bone 2011, 48, 1409–1416. [Google Scholar] [CrossRef]
- Leyhausen, G.; Lorenz, B.; Zhu, H.; Geurtsen, W.; Bohnensack, R.; Muller, W.E.G.; Schröder, H.C. Inorganic polyphosphate in human osteoblast-like cells. J. Bone Miner. Res. 1998, 13, 803–812. [Google Scholar] [CrossRef]
- Usui, Y.; Uematsu, T.; Uchihashi, T.; Takahashi, M.; Takahashi, M.; Ishizuka, M.; Doto, R.; Tanaka, H.; Komazaki, Y.; Osawa, M.; et al. Inorganic polyphosphate induces osteoblastic differentiation. J. Dent. Res. 2010, 89, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Hacchou, Y.; Uematsu, T.; Ueda, O.; Usui, Y.; Uematsu, S.; Takahashi, M.; Uchihashi, T.; Kawazoe, Y.; Shiba, T.; Kurihara, S.; et al. Inorganic polyphosphate: A possible stimulant of bone formation. J. Dent. Res. 2007, 86, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Kawazoe, Y.; Katoh, S.; Onodera, Y.; Kohgo, T.; Shindoh, M.; Shiba, T. Activation of the FGF signaling pathway and subsequent induction of mesenchymal stem cell differentiation by inorganic polyphosphate. Int. J. Biol. Sci. 2008, 4, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, D.; Tomita, T.; Kuroda, S.; Higuchi, C.; Kato, S.; Shiba, T.; Nakagami, H.; Morishita, R.; Yoshikawa, H. Inorganic polyphosphate differentiates human mesenchymal stem cells into osteoblastic cells. J. Bone Miner. Metab. 2010, 28, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Tolba, E.; Schröder, H.C.; Wang, S.F.; Glasser, G.; Munoz-Espi, R.; Link, T.; Wang, X.H. A new polyphosphate calcium material with morphogenetic activity. Mater. Lett. 2015, 148, 163–166. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Tolba, E.; Feng, Q.; Schröder, H.C.; Markl, J.S.; Kokkinopoulou, M.; Wang, X. Amorphous Ca2+ polyphosphate nanoparticles regulate the ATP level in bone-like SaOS-2 cells. J. Cell Sci. 2015, 128, 2202–2207. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Wang, S.; Wiens, M.; Neufurth, M.; Ackermann, M.; Relkovic, D.; Kokkinopoulou, M.; Feng, Q.; Schröder, H.C.; Wang, X. Uptake of polyphosphate microparticles in vitro (SaOS-2 and HUVEC cells) followed by an increase of the intracellular ATP pool size. PLoS ONE 2017, 12, e0188977. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Neufurth, M.; Wang, S.; Ackermann, M.; Munoz-Espi, R.; Feng, Q.; Lu, Q.; Schröder, H.C.; Wang, X. Amorphous, smart, and bioinspired polyphosphate nano/microparticles: A biomaterial for regeneration and repair of osteo-articular impairments in-situ. Int. J. Mol. Sci. 2018, 19, 427. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Tolba, E.; Schröder, H.C.; Munoz-Espi, R.; Diehl-Seifert, B.; Wang, X. Amorphous polyphosphate-hydroxyapatite: A morphogenetically active substrate for bone-related SaOS-2 cells in vitro. Acta Biomater. 2016, 31, 358–367. [Google Scholar] [CrossRef]
- Hamidouche, Z.; Hay, E.; Vaudin, P.; Charbord, P.; Schule, R.; Marie, P.J.; Fromigue, O. FHL2 mediates dexamethasone-induced mesenchymal cell differentiation into osteoblasts by activating Wnt/beta-catenin signaling-dependent Runx2 expression. FASEB J. 2008, 22, 3813–3822. [Google Scholar] [CrossRef]
- Shui, C.; Spelsberg, T.C.; Riggs, B.L.; Khosla, S. Changes in Runx2/Cbfa1 expression and activity during osteoblastic differentiation of human bone marrow stromal cells. J. Bone Miner. Res. 2003, 18, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Loebel, C.; Czekanska, E.M.; Bruderer, M.; Salzmann, G.; Alini, M.; Stoddart, M.J. In vitro osteogenic potential of human mesenchymal stem cells is predicted by Runx2/Sox9 ratio. Tissue Eng. Part A 2015, 21, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Hoemann, C.D.; El-Gabalawy, H.; McKee, M.D. In vitro osteogenesis assays: Influence of the primary cell source on alkaline phosphatase activity and mineralization. Pathol. Biol. 2009, 57, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Morita, K.; Hirata, I.; Doi, K.; Kubo, T.; Kato, K.; Tsuga, K. Enhancement of calcification by osteoblasts cultured on hydroxyapatite surfaces with adsorbed inorganic polyphosphate. In Vitro Cell Dev. Biol. Anim. 2018, 54, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.T.; Aoki, T.; Sakoda, M.; Ohta, S.; Ichimura, S.; Ito, T.; Ushida, T.; Furukawa, K.S. Enhancing osteogenic differentiation of MC3T3-E1 cells by immobilizing inorganic polyphosphate onto hyaluronic acid hydrogel. Biomacromolecules 2015, 16, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Wang, S.; Ackermann, M.; Gerich, T.; Neufurth, M.; Wiens, M.; Schröder, H.C.; Wang, X. Biologization of allogeneic bone grafts with polyphosphate: A route to a biomimetic periosteum. Adv. Funct. Mater. 2019, 29, 1905220. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Erices, A.; Conget, P.; Minguell, J.J. Mesenchymal progenitor cells in human umbilical cord blood. Br. J. Haematol. 2000, 109, 235–242. [Google Scholar] [CrossRef]
- Gardner, O.F.; Alini, M.; Stoddart, M.J. Mesenchymal stem cells derived from human bone marrow. Methods Mol. Biol. 2015, 1340, 41–52. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Osteocontrol | Osteocontrol + polyP-NP | Osteogenic (BGP) | Osteogenic (polyP-NP) | |
|---|---|---|---|---|
| LG-DMEM 1 | + | + | + | + |
| PEN/STREP | + | + | + | + |
| FBS | + | + | + | + |
| Dexa | - | - | + | + |
| AA2P | - | - | + | + |
| BGP | - | - | + | - |
| polyP-NP | - | + | - | + |
| Gene | Forward | Reverse | Probe |
| COL1A1 | 5′-CCC TGG AAA GAA TGG AGA TGA T-3′ | 5′-ACT GAA ACC TCT GTG TCC CTT CA-3′ | 5′-CGG GCA ATC CTC GAG CAC CCT-3′ |
| RPLP0 | 5′-TGG GCA AGA ACA CCA TGA TG-3′ | 5′-CGG ATA TGA GGC AGC AGT TTC-3′ | 5′-AGG GCA CCT GGA AAA CAA CCC AGC-3′ |
| RUNX2 | 5′-AGC AAG GTT CAA CGA TCT GAG AT-3′ | 5′-TTT GTG AAG ACG GTT ATG GTC AA-3′ | 5′-TGA AAC TCT TGC CTC GTC CAC TCC G-3′ |
| Gene | Assay on Demand Ref No. (Applied Biosystems) | ||
| ALPL | Hs00758162_m1 | ||
| IBSP | Hs0017320_m1 | ||
| PPAR-γ | Hs00234592_m1 | ||
| SOX9 | Hs00165814_m1 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phelipe Hatt, L.; Thompson, K.; Müller, W.E.G.; Stoddart, M.J.; Armiento, A.R. Calcium Polyphosphate Nanoparticles Act as an Effective Inorganic Phosphate Source during Osteogenic Differentiation of Human Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 5801. https://doi.org/10.3390/ijms20225801
Phelipe Hatt L, Thompson K, Müller WEG, Stoddart MJ, Armiento AR. Calcium Polyphosphate Nanoparticles Act as an Effective Inorganic Phosphate Source during Osteogenic Differentiation of Human Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2019; 20(22):5801. https://doi.org/10.3390/ijms20225801
Chicago/Turabian StylePhelipe Hatt, Luan, Keith Thompson, Werner E. G. Müller, Martin James Stoddart, and Angela Rita Armiento. 2019. "Calcium Polyphosphate Nanoparticles Act as an Effective Inorganic Phosphate Source during Osteogenic Differentiation of Human Mesenchymal Stem Cells" International Journal of Molecular Sciences 20, no. 22: 5801. https://doi.org/10.3390/ijms20225801
APA StylePhelipe Hatt, L., Thompson, K., Müller, W. E. G., Stoddart, M. J., & Armiento, A. R. (2019). Calcium Polyphosphate Nanoparticles Act as an Effective Inorganic Phosphate Source during Osteogenic Differentiation of Human Mesenchymal Stem Cells. International Journal of Molecular Sciences, 20(22), 5801. https://doi.org/10.3390/ijms20225801