Human Wharton’s Jelly-Derived Mesenchymal Stem Cells Minimally Improve the Growth Kinetics and Cardiomyocyte Differentiation of Aged Murine Cardiac c-kit Cells in In Vitro without Rejuvenating Effect


 , , ,
, , ,
Abstract
1. Introduction
2. Results
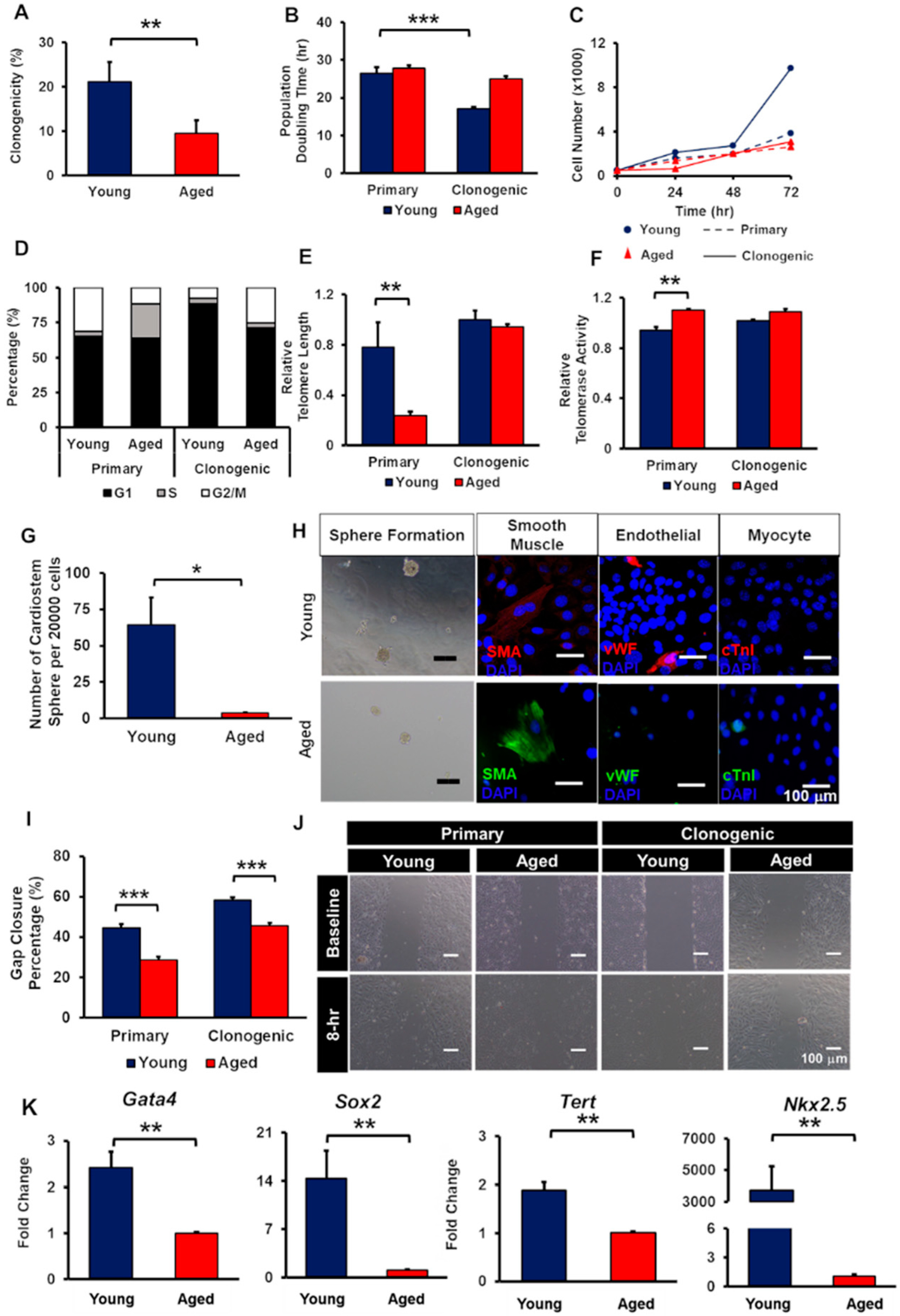
2.1. Isolation and Characterisation of CCs
2.2. Characterisations of CCs Isolated from Young and Aged Mice
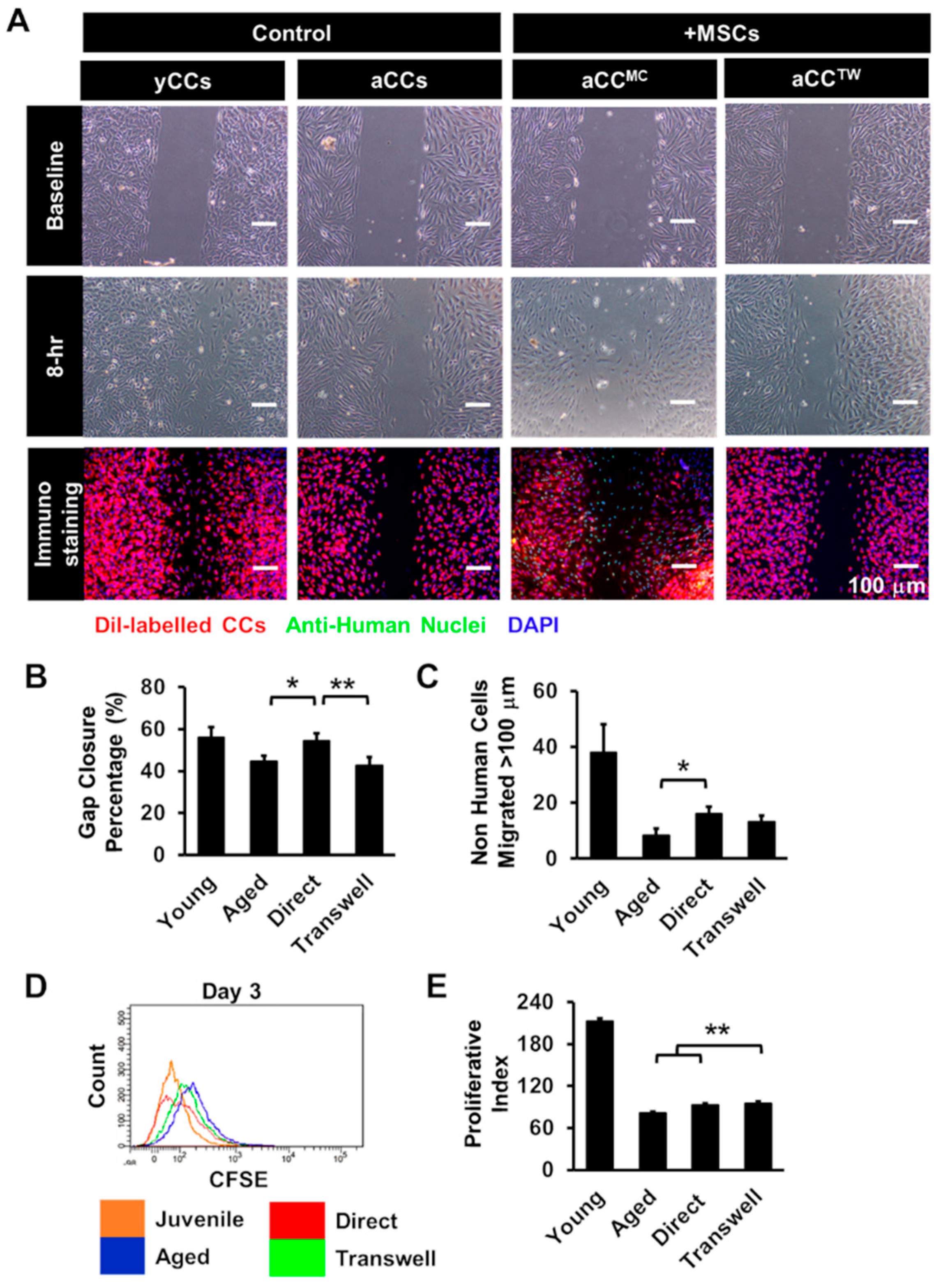
2.3. Co-culture with MSCs Can Improve the Growth Kinetics of aCCs
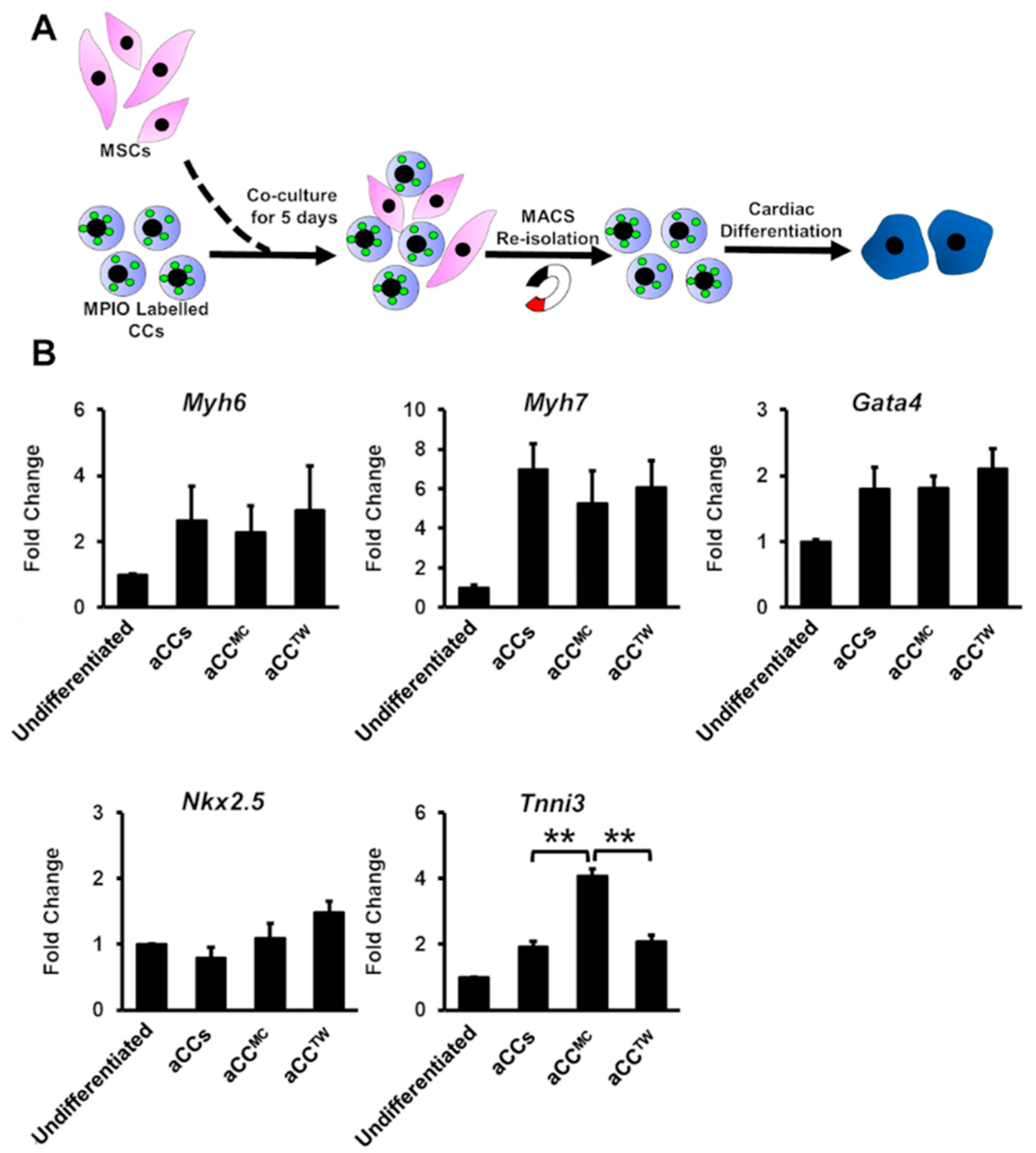
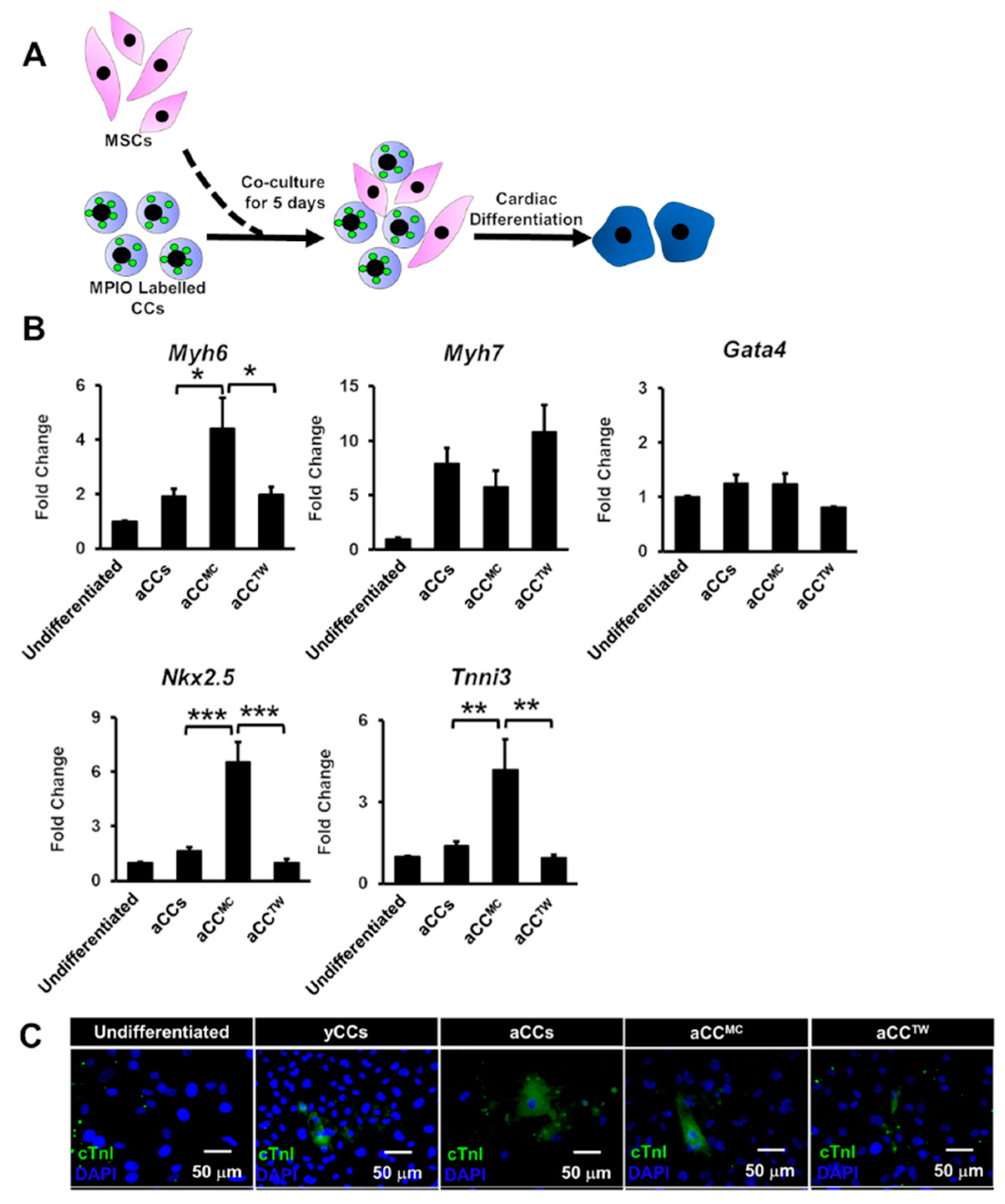
2.4. aCC Cardiomyocyte Differentiation in MSC Co-Culture
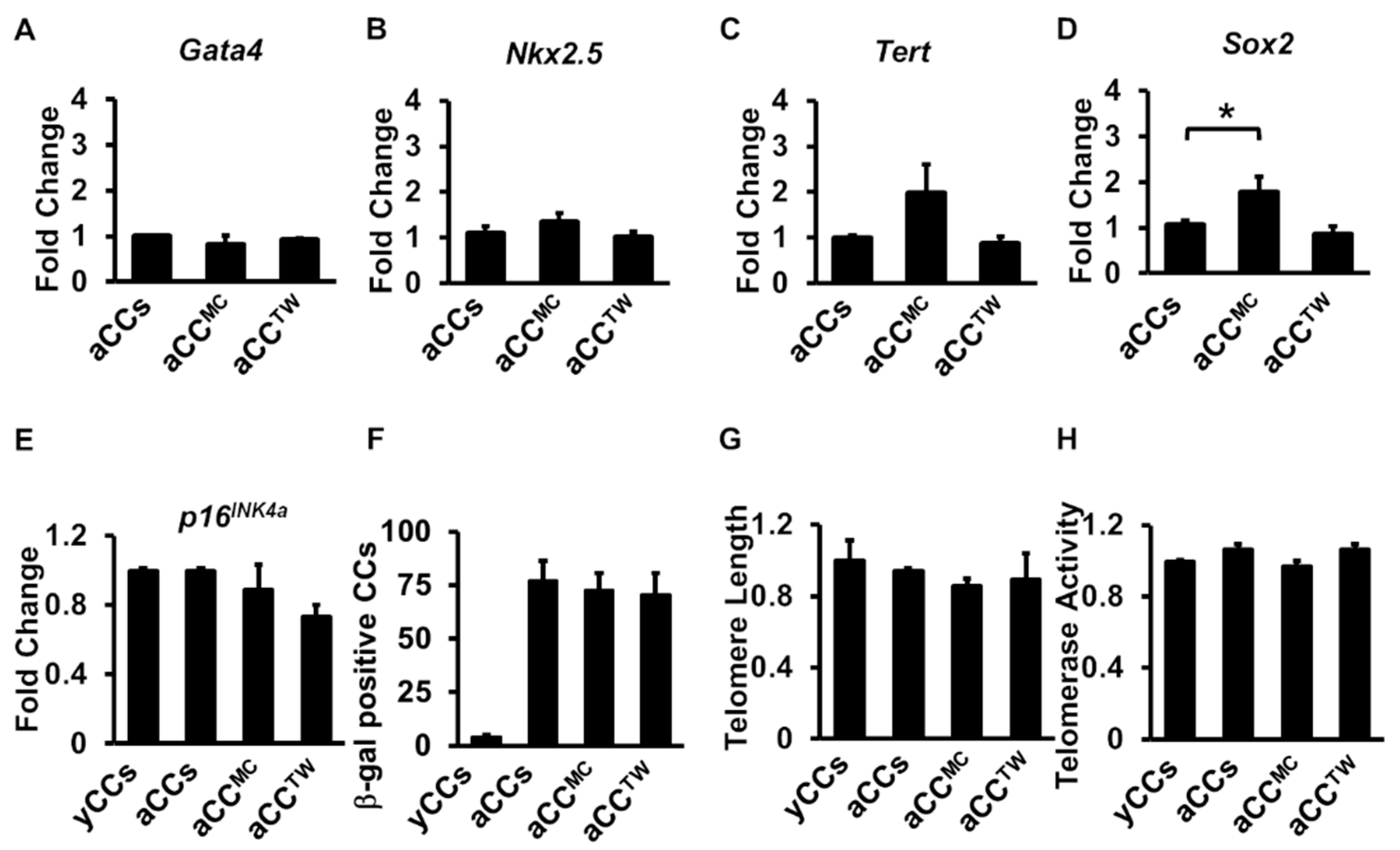
2.5. Effects of MSCs Co-Culture on aCC Stemness, Telomere Length, and Telomerase Activity
3. Discussion
4. Materials and Methods
4.1. Isolation and Characterisation of Cardiac c-kit Cells (CCs) from Young and Aged Mice
4.2. Isolation and Characterisation of Wharton’s Jelly-Derived Mesenchymal Stem Cells
4.3. CC-MSC Co-Culture
4.4. Magnetic Particles Iron Oxide (MPIO) Labelling
4.5. DiI Labelling
4.6. Flow Cytometry
4.7. Immunofluorescence Labelling
4.8. CardioStem Sphere Formation and Trilineage Differentiation
4.9. Chemical-Induced CC Cardiac Differentiation
4.10. RNA Extraction and cDNA Conversion
4.11. DNA Extraction
4.12. Protein Extraction
4.13. Quantitative Real-Time PCR
4.14. Telomere Length Assessment
4.15. Real-Time Quantitative Telomeric Repeat Amplification Protocol (RQ-TRAP)
4.16. Proliferation Assessment of Aged CCs
4.17. Senescence Assay
4.18. Viability Assay
4.19. Migration
4.20. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Fact Sheet-Cardiovascular diseases (CVDs). 2016. Available online: http://www.who.int/mediacentre/factsheets/fs2317/en/ (accessed on 9 March 2018)2016 09 March; pp viewed 09 March 2018.
- Welp, H.; Spieker, T.; Erren, M.; Scheld, H.H.; Baba, H.A.; Stypmann, J. Sex mismatch in heart transplantation is associated with increased number of severe rejection episodes and shorter long-term survival. Transplant. Proc. 2009, 41, 2579–2584. [Google Scholar] [CrossRef] [PubMed]
- Kilic, A.; Emani, S.; Sai-Sudhakar, C.B.; Higgins, R.S.D.; Whitson, B.A. Donor selection in heart transplantation. J. Thorac. Dis. 2014, 6, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Bolli, R.; Chugh, A.R.; D’Amario, D.; Loughran, J.H.; Stoddard, M.F.; Ikram, S.; Beache, G.M.; Wagner, S.G.; Leri, A.; Hosoda, T.; et al. Cardiac stem cells in patients with ischaemic cardiomyopathy (SCIPIO): Initial results of a randomised phase 1 trial. Lancet (Lond. Engl.) 2011, 378, 1847–1857. [Google Scholar] [CrossRef]
- Chugh, A.R.; Beache, G.M.; Loughran, J.H.; Mewton, N.; Elmore, J.B.; Kajstura, J.; Pappas, P.; Tatooles, A.; Stoddard, M.F.; Lima, J.A.; et al. Administration of cardiac stem cells in patients with ischemic cardiomyopathy: the SCIPIO trial: surgical aspects and interim analysis of myocardial function and viability by magnetic resonance. Circulation 2012, 126, S54–S64. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yan, G.; He, W.; Liu, Z.; Xu, H.; Ma, G. The influence of disease and age on human cardiac stem cells. Ann. Clin. Biochem. 2014, 51, 582–590. [Google Scholar] [CrossRef]
- Cesselli, D.; Beltrami, A.P.; D’Aurizio, F.; Marcon, P.; Bergamin, N.; Toffoletto, B.; Pandolfi, M.; Puppato, E.; Marino, L.; Signore, S.; et al. Effects of age and heart failure on human cardiac stem cell function. Am. J. Pathol. 2011, 179, 349–366. [Google Scholar] [CrossRef]
- Williams, A.R.; Hatzistergos, K.E.; Addicott, B.; McCall, F.; Carvalho, D.; Suncion, V.; Morales, A.R.; Da Silva, J.; Sussman, M.A.; Heldman, A.W.; et al. Enhanced Effect of Human Cardiac Stem Cells and Bone Marrow Mesenchymal Stem Cells to Reduce Infarct Size and Restore Cardiac Function after Myocardial Infarction. Circulation 2012, 127, 213–223. [Google Scholar] [CrossRef]
- Smith, A.J.; Lewis, F.C.; Aquila, I.; Waring, C.D.; Nocera, A.; Agosti, V.; Nadal-Ginard, B.; Torella, D.; Ellison, G.M. Isolation and characterization of resident endogenous c-Kit+ cardiac stem cells from the adult mouse and rat heart. Nat. Protoc. 2014, 9, 1662–1681. [Google Scholar] [CrossRef]
- He, J.Q.; Vu, D.M.; Hunt, G.; Chugh, A.; Bhatnagar, A.; Bolli, R. Human cardiac stem cells isolated from atrial appendages stably express c-kit. PLoS ONE 2011, 6, e27719. [Google Scholar] [CrossRef] [PubMed]
- Smits, A.M.; van Vliet, P.; Metz, C.H.; Korfage, T.; Sluijter, J.P.; Doevendans, P.A.; Goumans, M.J. Human cardiomyocyte progenitor cells differentiate into functional mature cardiomyocytes: an in vitro model for studying human cardiac physiology and pathophysiology. Nat. Protoc. 2009, 4, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Gambini, E.; Pompilio, G.; Biondi, A.; Alamanni, F.; Capogrossi, M.C.; Agrifoglio, M.; Pesce, M. C-kit+ cardiac progenitors exhibit mesenchymal markers and preferential cardiovascular commitment. Cardiovasc. Res. 2011, 89, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.Y.; Ng, W.H.; Umar Fuaad, M.Z.; Ng, C.T.; Ramasamy, R.; Lim, V.; Yong, Y.K.; Tan, J.J. Mesenchymal stem cells facilitate cardiac differentiation in Sox2-expressing cardiac C-kit cells in coculture. J. Cell. Biochem. 2018, 120, 9104–9116. [Google Scholar] [CrossRef] [PubMed]
- Sanada, F.; Kim, J.; Czarna, A.; Chan, N.Y.; Signore, S.; Ogorek, B.; Isobe, K.; Wybieralska, E.; Borghetti, G.; Pesapane, A.; et al. c-Kit-positive cardiac stem cells nested in hypoxic niches are activated by stem cell factor reversing the aging myopathy. Circ. Res. 2014, 114, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Hosoyama, T.; Murakami, J.; Samura, M.; Ueno, K.; Kurazumi, H.; Suzuki, R.; Mikamo, A.; Hamano, K. Age-related increase in Wnt inhibitor causes a senescence-like phenotype in human cardiac stem cells. Biochem. Biophys. Res. Commun. 2017, 487, 653–659. [Google Scholar] [CrossRef]
- Nakamura, T.; Hosoyama, T.; Kawamura, D.; Takeuchi, Y.; Tanaka, Y.; Samura, M.; Ueno, K.; Nishimoto, A.; Kurazumi, H.; Suzuki, R.; et al. Influence of aging on the quantity and quality of human cardiac stem cells. Sci. Rep. 2016, 6, 22781. [Google Scholar] [CrossRef]
- Avolio, E.; Gianfranceschi, G.; Cesselli, D.; Caragnano, A.; Athanasakis, E.; Katare, R.; Meloni, M.; Palma, A.; Barchiesi, A.; Vascotto, C.; et al. Ex vivo molecular rejuvenation improves the therapeutic activity of senescent human cardiac stem cells in a mouse model of myocardial infarction. Stem Cells (Dayt. Ohio) 2014, 32, 2373–2385. [Google Scholar] [CrossRef]
- Chong, J.J.; Chandrakanthan, V.; Xaymardan, M.; Asli, N.S.; Li, J.; Ahmed, I.; Heffernan, C.; Menon, M.K.; Scarlett, C.J.; Rashidianfar, A.; et al. Adult cardiac-resident MSC-like stem cells with a proepicardial origin. Cell Stem Cell 2011, 9, 527–540. [Google Scholar] [CrossRef]
- Hsiao, L.C.; Perbellini, F.; Gomes, R.S.; Tan, J.J.; Vieira, S.; Faggian, G.; Clarke, K.; Carr, C.A. Murine cardiosphere-derived cells are impaired by age but not by cardiac dystrophic dysfunction. Stem Cells Dev. 2014, 23, 1027–1036. [Google Scholar] [CrossRef]
- Tan, J.-J.; Carr, C.A.; Stuckey, D.J.; Ellison, G.M.; Messina, E.; Giacomello, A.; Clarke, K. Isolation and Expansion of Cardiosphere-Derived Stem Cells. Curr. Protoc. Stem Cell Biol. 2011, 16, 2C.3.1–2C.3.12. [Google Scholar] [CrossRef]
- Leong, Y.Y.; Ng, W.H.; Ellison-Hughes, G.M.; Tan, J.J. Cardiac Stem Cells for Myocardial Regeneration: They Are Not Alone. Front. Cardiovasc. Med. 2017, 4, 47. [Google Scholar] [CrossRef]
- Murakami, Y.; Hirata, H.; Miyamoto, Y.; Nagahashi, A.; Sawa, Y.; Jakt, M.; Asahara, T.; Kawamata, S. Isolation of cardiac cells from E8.5 yolk sac by ALCAM (CD166) expression. Mech. Dev. 2007, 124, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Hirata, H.; Murakami, Y.; Miyamoto, Y.; Tosaka, M.; Inoue, K.; Nagahashi, A.; Jakt, L.M.; Asahara, T.; Iwata, H.; Sawa, Y.; et al. ALCAM (CD166) is a surface marker for early murine cardiomyocytes. Cells Tissues Organs 2006, 184, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Gessert, S.; Maurus, D.; Brade, T.; Walther, P.; Pandur, P.; Kuhl, M. DM-GRASP/ALCAM/CD166 is required for cardiac morphogenesis and maintenance of cardiac identity in first heart field derived cells. Dev. Biol. 2008, 321, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Lewis-McDougall, F.C.; Ruchaya, P.J.; Domenjo-Vila, E.; Shin Teoh, T.; Prata, L.; Cottle, B.J.; Clark, J.E.; Punjabi, P.P.; Awad, W.; Torella, D.; et al. Aged-senescent cells contribute to impaired heart regeneration. Aging Cell 2019, 18, e12931. [Google Scholar] [CrossRef] [PubMed]
- Subramani, B.; Subbannagounder, S.; Palanivel, S.; Ramanathanpullai, C.; Sivalingam, S.; Yakub, A.; SadanandaRao, M.; Seenichamy, A.; Pandurangan, A.K.; Tan, J.J.; et al. Generation and characterization of human cardiac resident and non-resident mesenchymal stem cell. Cytotechnology 2016, 68, 2061–2073. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oldershaw, R.; Owens, W.A.; Sutherland, R.; Linney, M.; Liddle, R.; Magana, L.; Lash, G.E.; Gill, J.H.; Richardson, G.; Meeson, A. Human Cardiac-Mesenchymal Stem Cell-Like Cells, a Novel Cell Population with Therapeutic Potential. Stem Cells Dev. 2019, 28, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Garikipati, V.N.S.; Singh, S.P.; Mohanram, Y.; Gupta, A.K.; Kapoor, D.; Nityanand, S. Isolation and characterization of mesenchymal stem cells from human fetus heart. PLoS ONE 2018, 13, e0192244. [Google Scholar] [CrossRef]
- Castaldi, A.; Dodia, R.M.; Orogo, A.M.; Zambrano, C.M.; Najor, R.H.; Gustafsson, Å.B.; Heller Brown, J.; Purcell, N.H. Decline in cellular function of aged mouse c-kit(+) cardiac progenitor cells. J. Physiol. 2017, 595, 6249–6262. [Google Scholar] [CrossRef]
- O’Sullivan, R.J.; Karlseder, J. Telomeres: protecting chromosomes against genome instability. Nat. Reviews. Mol. Cell Biol. 2010, 11, 171–181. [Google Scholar] [CrossRef]
- Nugent, C.I.; Lundblad, V. The telomerase reverse transcriptase: components and regulation. Genes Dev. 1998, 12, 1073–1085. [Google Scholar] [CrossRef]
- Osterhage, J.L.; Friedman, K.L. Chromosome end maintenance by telomerase. J. Biol. Chem. 2009, 284, 16061–16065. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Meng, Q.; Li, Y.; Deng, S.; Yu, Z.; Liu, Z.; Zhang, L.; Fan, H. C-Kit Positive Cardiac Stem Cells and Bone Marrow-Derived Mesenchymal Stem Cells Synergistically Enhance Angiogenesis and Improve Cardiac Function After Myocardial Infarction in a Paracrine Manner. J. Card. Fail. 2017, 23, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Karantalis, V.; Suncion-Loescher, V.Y.; Bagno, L.; Golpanian, S.; Wolf, A.; Sanina, C.; Premer, C.; Kanelidis, A.J.; McCall, F.; Wang, B.; et al. Synergistic Effects of Combined Cell Therapy for Chronic Ischemic Cardiomyopathy. J. Am. Coll. Cardiol. 2015, 66, 1990–1999. [Google Scholar] [CrossRef] [PubMed]
- Conboy, I.M.; Rando, T.A. Heterochronic parabiosis for the study of the effects of aging on stem cells and their niches. Cell Cycle (Georget. Tex.) 2012, 11, 2260–2267. [Google Scholar] [CrossRef]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef]
- Katsimpardi, L.; Litterman, N.K.; Schein, P.A.; Miller, C.M.; Loffredo, F.S.; Wojtkiewicz, G.R.; Chen, J.W.; Lee, R.T.; Wagers, A.J.; Rubin, L.L. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science 2014, 344, 630–634. [Google Scholar] [CrossRef]
- Li, S.H.; Sun, L.; Yang, L.; Li, J.; Shao, Z.; Du, G.Q.; Wu, J.; Weisel, R.D.; Li, R.K. Young Bone-Marrow Sca-1+ Stem Cells Rejuvenate the Aged Heart and Improve Function after Injury through PDGFRbeta-Akt pathway. Sci Rep. 2017, 7, 41756. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, D.; Li, S.; Chang, L.; Zhang, Y.; Liu, R.; Sun, F.; Duan, W.; Du, W.; Wu, Y.; et al. Bone marrow mesenchymal stem cell transplantation retards the natural senescence of rat hearts. Stem Cells Transl. Med. 2015, 4, 494–502. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, W.; Bender, I.; Gong, W.; Kwak, I.-Y.; Yellamilli, A.; Hodges, T.J.; Nemoto, N.; Zhang, J.; Garry, D.J.; et al. Pathologic Stimulus Determines Lineage Commitment of Cardiac C-kit Cells. Circulation 2017, 136, 2359–2372. [Google Scholar] [CrossRef]
- Bruunsgaard, H.; Pedersen, B.K. Age-related inflammatory cytokines and disease. Immunol. Allergy Clin. North. Am. 2003, 23, 15–39. [Google Scholar] [CrossRef]
- Jenny, N.S. Inflammation in aging: cause, effect, or both? Discov. Med. 2012, 13, 451–460. [Google Scholar] [PubMed]
- Quijada, P.; Salunga, H.T.; Hariharan, N.; Cubillo, J.D.; El-Sayed, F.G.; Moshref, M.; Bala, K.M.; Emathinger, J.M.; De La Torre, A.; Ormachea, L.; et al. Cardiac stem cell hybrids enhance myocardial repair. Circ. Res. 2015, 117, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Rahbarghazi, R.; Nassiri, S.M.; Khazraiinia, P.; Kajbafzadeh, A.M.; Ahmadi, S.H.; Mohammadi, E.; Molazem, M.; Zamani-Ahmadmahmudi, M. Juxtacrine and paracrine interactions of rat marrow-derived mesenchymal stem cells, muscle-derived satellite cells, and neonatal cardiomyocytes with endothelial cells in angiogenesis dynamics. Stem Cells Dev. 2013, 22, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Markel, T.A.; Wang, Y.; Herrmann, J.L.; Crisostomo, P.R.; Wang, M.; Novotny, N.M.; Herring, C.M.; Tan, J.; Lahm, T.; Meldrum, D.R. VEGF is critical for stem cell-mediated cardioprotection and a crucial paracrine factor for defining the age threshold in adult and neonatal stem cell function. Am. J. Physiology. Heart Circ. Physiol. 2008, 295, H2308–H2314. [Google Scholar] [CrossRef] [PubMed]
- Mirotsou, M.; Zhang, Z.; Deb, A.; Zhang, L.; Gnecchi, M.; Noiseux, N.; Mu, H.; Pachori, A.; Dzau, V. Secreted frizzled related protein 2 (Sfrp2) is the key Akt-mesenchymal stem cell-released paracrine factor mediating myocardial survival and repair. Proc. Natl. Acad. Sci. USA 2007, 104, 1643–1648. [Google Scholar] [CrossRef]
- Timmers, L.; Lim, S.K.; Arslan, F.; Armstrong, J.S.; Hoefer, I.E.; Doevendans, P.A.; Piek, J.J.; El Oakley, R.M.; Choo, A.; Lee, C.N.; et al. Reduction of myocardial infarct size by human mesenchymal stem cell conditioned medium. Stem Cell Res. 2007, 1, 129–137. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef]
- Callicott, R.J.; Womack, J.E. Real-time PCR assay for measurement of mouse telomeres. Comp. Med. 2006, 56, 17–22. [Google Scholar]
- Wege, H.; Chui, M.S.; Le, H.T.; Tran, J.M.; Zern, M.A. SYBR Green real-time telomeric repeat amplification protocol for the rapid quantification of telomerase activity. Nucleic Acids Res. 2003, 31, E3. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody. | Dilution Factor | Application | Manufacturer |
|---|---|---|---|
| Rabbit Polyclonal Anti-c-kit Antibody (H-300) | 1:50 | ICC/FC | Santa Cruz Biotechnology, Germany (sc-5535) |
| FITC Rat Anti-mouse CD34 Antibody (Clone RAM34) | 1:50 | FC | BD Bioscience, USA (560238) |
| FITC Rat Anti-mouse CD90.2 Antibody (Clone 53-2.1,RUO) | 1:50 | FC | BD Bioscience, USA (553003) |
| FITC Rat Anti-mouse Sca-1 Antibody (Clone D7) | 1:10 | FC | Miltenyi Biotec, Germany (130-102-297) |
| PE Rat Anti-mouse CD140a Antibody (Clone APA5) | 1:10 | FC | Miltenyi Biotec, Germany (130-102-502) |
| PE Rat Anti-mouse CD166 Antibody (Clone eBioALC48) | 1:50 | FC | eBioscience, USA (12-1661-81) |
| PE Rat Anti-mouse CD105 Antibody (Clone MJ7/18) | 1:10 | FC | Miltenyi Biotec, Germany (130-102-548) |
| FITC Rat Anti-mouse CD45 Antibody (Clone 30F11) | 1:10 | FC | Miltenyi Biotec, Germany (130-102-491) |
| Rabbit Polyclonal Anti-GATA-4 Antibody (H-112) | 1:50 | ICC | Santa Cruz Biotechnology, Germany (sc-9053) |
| Rabbit Polyclonal Anti-NKX2.5 Antibody (H-114) | 1:50 | ICC | Santa Cruz Biotechnology, Germany (sc-14033) |
| Rabbit Polyclonal Anti-Ki67 Antibody (SP6) | 1:50 | ICC | Genetex, Germany (GTX16667) |
| Goat Polyclonal Anti-Tryptase Antibody (V-13) | 1:50 | ICC | Santa Cruz Biotechnology, Germany (sc-32473) |
| Goat Polyclonal Anti-Sox2 Antibody (Y-17) | 1:50 | ICC | Santa Cruz Biotechnology, Germany (sc-17320) |
| Rabbit Polyclonal Anti-OCT3/4 Antibody (H-134) | 1:50 | ICC | Santa Cruz Biotechnology, Germany (sc-9081) |
| Mouse Monoclonal Anti-Smooth Mucle Actinin (Clone 5C5) | 1:400 | ICC | Sigma Aldrich, USA (A2172) |
| Rabbit Polyclonal Anti-von Willebrand Factor Antibody | 1:400 | ICC | Dako, USA (A0082) |
| Rabbit Polyclonal anti-Cardiac Troponin I (H-170) | 1:50 | ICC | Santa Cruz Biotechnology, Germany (sc-15368) |
| Alexa Fluor 488 Donkey Anti-rabbit Antibody | 1:500 | ICC | Molecular Probes, CA |
| Alexa Fluor 488 Donkey Anti-goat Antibody | 1:500 | ICC | Molecular Probes, CA |
| Alexa Fluor 568 Donkey Anti-rabbit Antibody | 1:500 | ICC | Molecular Probes, CA |
| Alexa Fluor 568 Donkey Anti-goat Antibody | 1:500 | ICC | Molecular Probes, CA |
| Gene/Accession Number | Primer Sequence (5′-3′) |
|---|---|
| Gata4 NM_008092.3 | Forward: TCTCTGCATGTCCCATACCA Reverse: TGTGTGTGAAGGGGTGAAAA |
| Nkx2.5 NM_008700.2 | Forward: GCTACAAGTGCAAGCGACAG Reverse: GGGTAGGCGTTGTAGCCATA |
| Sox2 NM_011443.3 | Forward: GCGGAGTGGAAACTTTTGTCC Reverse: CGGGAAGCGTGTACTTATCCTT |
| Tert NM_009354.1 | Forward: TGGGTCTCCCCTGTACCAAAT Reverse: GGCCTGTAACTAGCGGACACA |
| Myh6 NM_010856.4 | Forward: AAGGTGAAGGCCTACAAGCG Reverse: GGTCTGCTGGAGAGGTTATTCC |
| Myh7 NM_080728.2 | Forward: GCCAACACCAACCTGTCCAAGTTC Reverse: TGCAAAGGCTCCAGGTCTGAGGGC |
| Tnni3 NM_000353.4 | Forward: TCTGCCAACTACCGAGCCTAT Reverse: CTCTTCTGCCTCTCGTTCCAT |
| p16INK4a NM_009877.2 | Forward: CGCAGGTTCTTGGTCACTGT Reverse: TGTTCACGAAAGCCAGAGCG |
| Gapdh NM_008084.2 | Forward: ACCCAGAAGACTGTGGATGG Reverse: CACATTGGGGGTAGGAACAC |
| Gene | Primer Sequence (5′-3′) |
|---|---|
| Telomere | Forward: CGGTTTGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTT Reverse: GGCTTGCCTTACCCTTACCCTTACCCTTACCCTTACCCT |
| 36B4 | Forward: ACTGGTCTAGGACCCGAGAAG Reverse: TCAATGGTGCCTCTGGAGATT |
| Stage | Time | Temperature |
|---|---|---|
| Holding Stage | 2 min | 95 °C |
| Cycling Stage (For telomere sequence) | ||
| (i) Denaturation | 15 s | 95 °C |
| (ii) Annealing extension | 1 min | 56 °C |
| Cycling Stage (For 36B4 sequence) | ||
| (i) Denaturation | 15 s | 95 °C |
| (ii) Annealing | 20 s | 52 °C |
| (iii) Extension | 30 s | 72 °C |
| Gene | Primer Sequence (5′-3′) |
|---|---|
| TS substrate | AATCCGTCGAGCAGAGTT |
| ACX (Anchored primers) | GCGCGGCTTACCCTTACCCTTACCCTAACC |
| Stage | Time | Temperature |
|---|---|---|
| Holding Stage | 20 min | 25 °C |
| Cycling Stage | ||
| (i) Denaturation | 30 s | 95 °C |
| (ii) Annealing extension | 90 s | 60 °C |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, W.H.; Yong, Y.K.; Ramasamy, R.; Ngalim, S.H.; Lim, V.; Shaharuddin, B.; Tan, J.J. Human Wharton’s Jelly-Derived Mesenchymal Stem Cells Minimally Improve the Growth Kinetics and Cardiomyocyte Differentiation of Aged Murine Cardiac c-kit Cells in In Vitro without Rejuvenating Effect. Int. J. Mol. Sci. 2019, 20, 5519. https://doi.org/10.3390/ijms20225519
Ng WH, Yong YK, Ramasamy R, Ngalim SH, Lim V, Shaharuddin B, Tan JJ. Human Wharton’s Jelly-Derived Mesenchymal Stem Cells Minimally Improve the Growth Kinetics and Cardiomyocyte Differentiation of Aged Murine Cardiac c-kit Cells in In Vitro without Rejuvenating Effect. International Journal of Molecular Sciences. 2019; 20(22):5519. https://doi.org/10.3390/ijms20225519
Chicago/Turabian StyleNg, Wai Hoe, Yoke Keong Yong, Rajesh Ramasamy, Siti Hawa Ngalim, Vuanghao Lim, Bakiah Shaharuddin, and Jun Jie Tan. 2019. "Human Wharton’s Jelly-Derived Mesenchymal Stem Cells Minimally Improve the Growth Kinetics and Cardiomyocyte Differentiation of Aged Murine Cardiac c-kit Cells in In Vitro without Rejuvenating Effect" International Journal of Molecular Sciences 20, no. 22: 5519. https://doi.org/10.3390/ijms20225519
APA StyleNg, W. H., Yong, Y. K., Ramasamy, R., Ngalim, S. H., Lim, V., Shaharuddin, B., & Tan, J. J. (2019). Human Wharton’s Jelly-Derived Mesenchymal Stem Cells Minimally Improve the Growth Kinetics and Cardiomyocyte Differentiation of Aged Murine Cardiac c-kit Cells in In Vitro without Rejuvenating Effect. International Journal of Molecular Sciences, 20(22), 5519. https://doi.org/10.3390/ijms20225519