Fucoxanthin-Rich Brown Algae Extract Improves Male Reproductive Function on Streptozotocin-Nicotinamide-Induced Diabetic Rat Model



Abstract

1. Introduction
2. Results
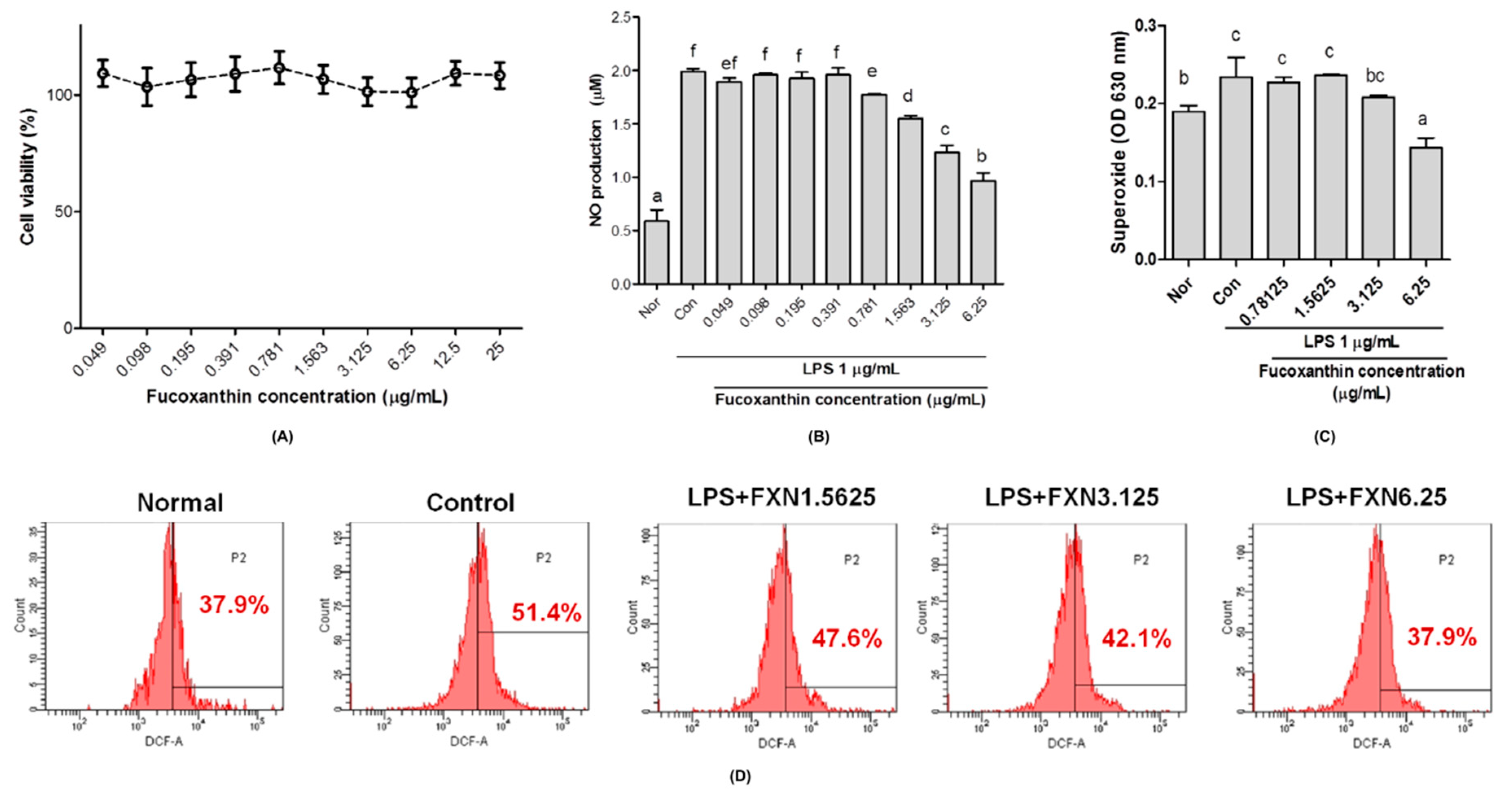
2.1. Effects of Fucoxanthin on Cells Viability, Nitric Oxide, and Reactive Oxygen Species Productions in RAW 264.7 Macrophage Cells
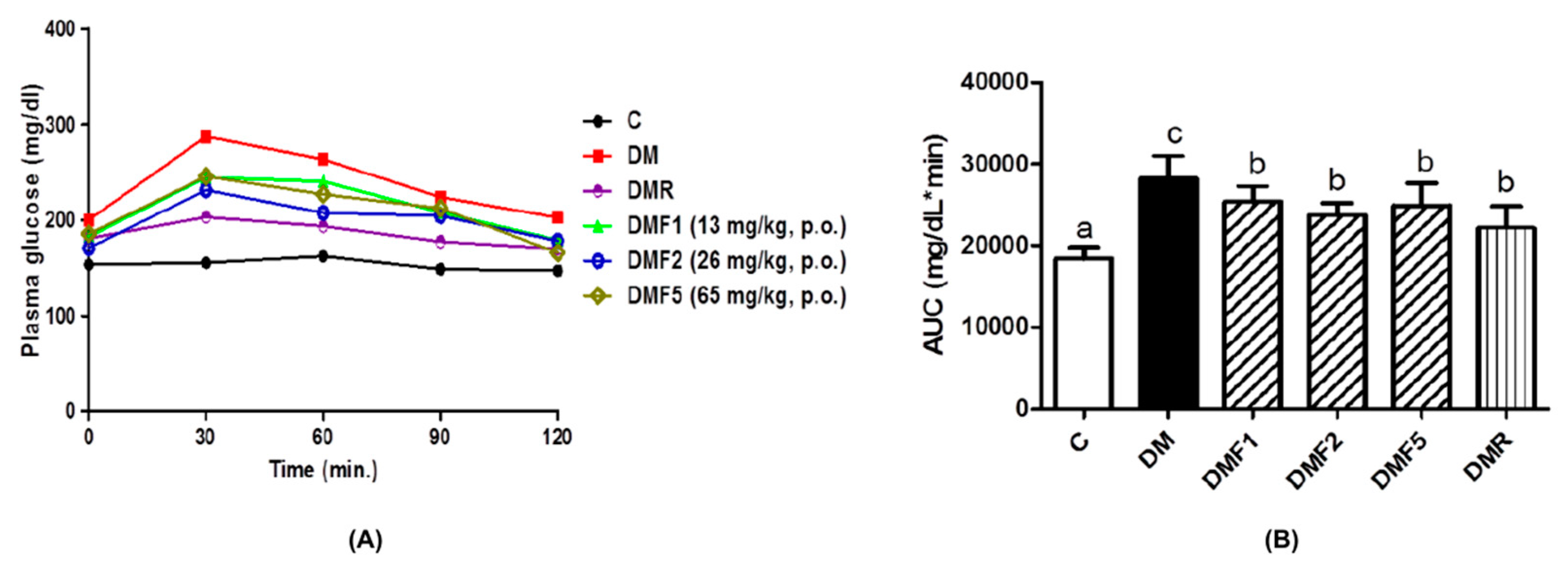
2.2. Effects of Fucoxanthin on Plasma Glucose, Insulin Level, and HOMA-IR
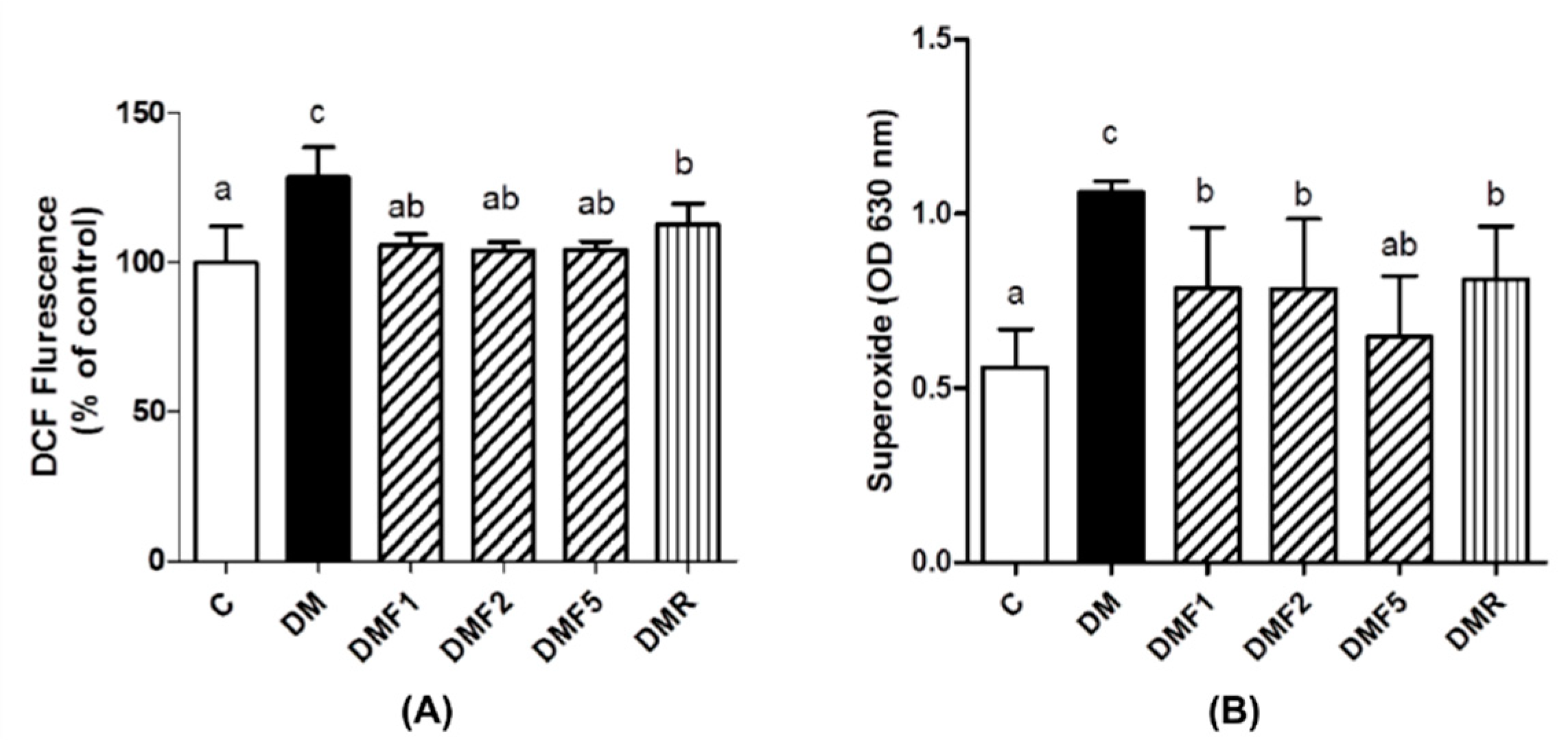
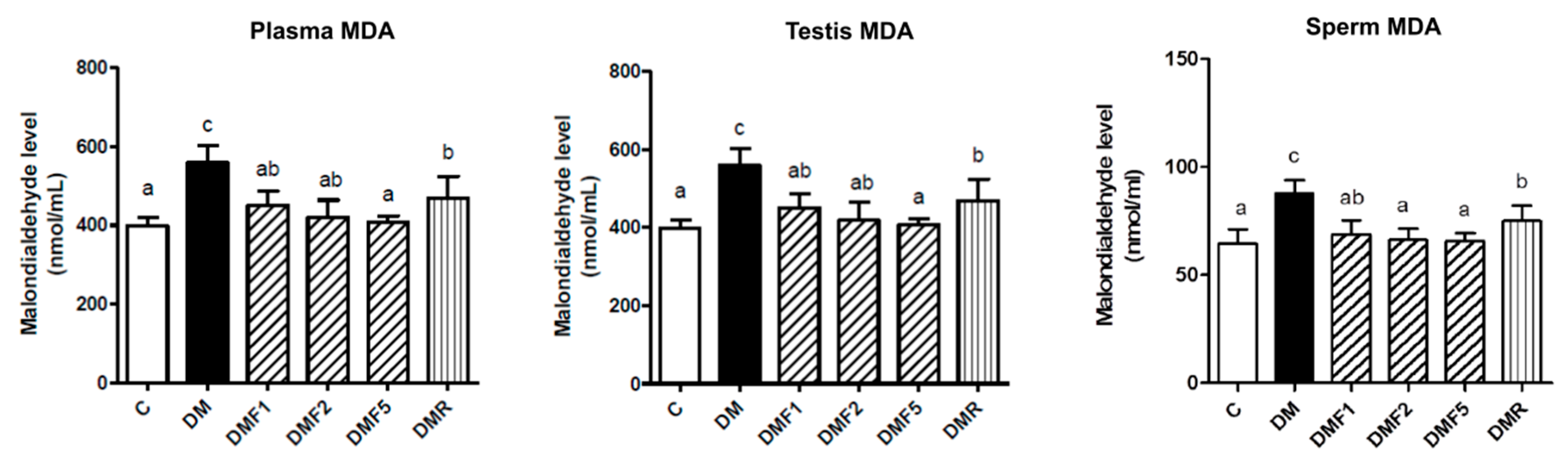
2.3. Effects of Fucoxanthin on Enzymatic Antioxidant and Oxidative Stress Level of Rats
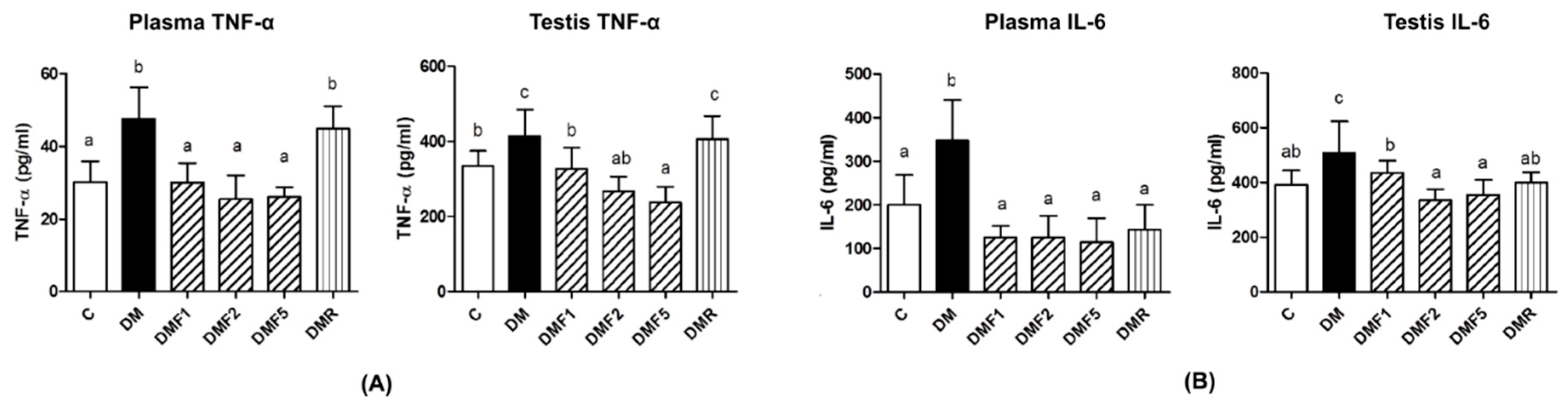
2.4. Effects of Fucoxanthin on Proinflammatory Cytokines
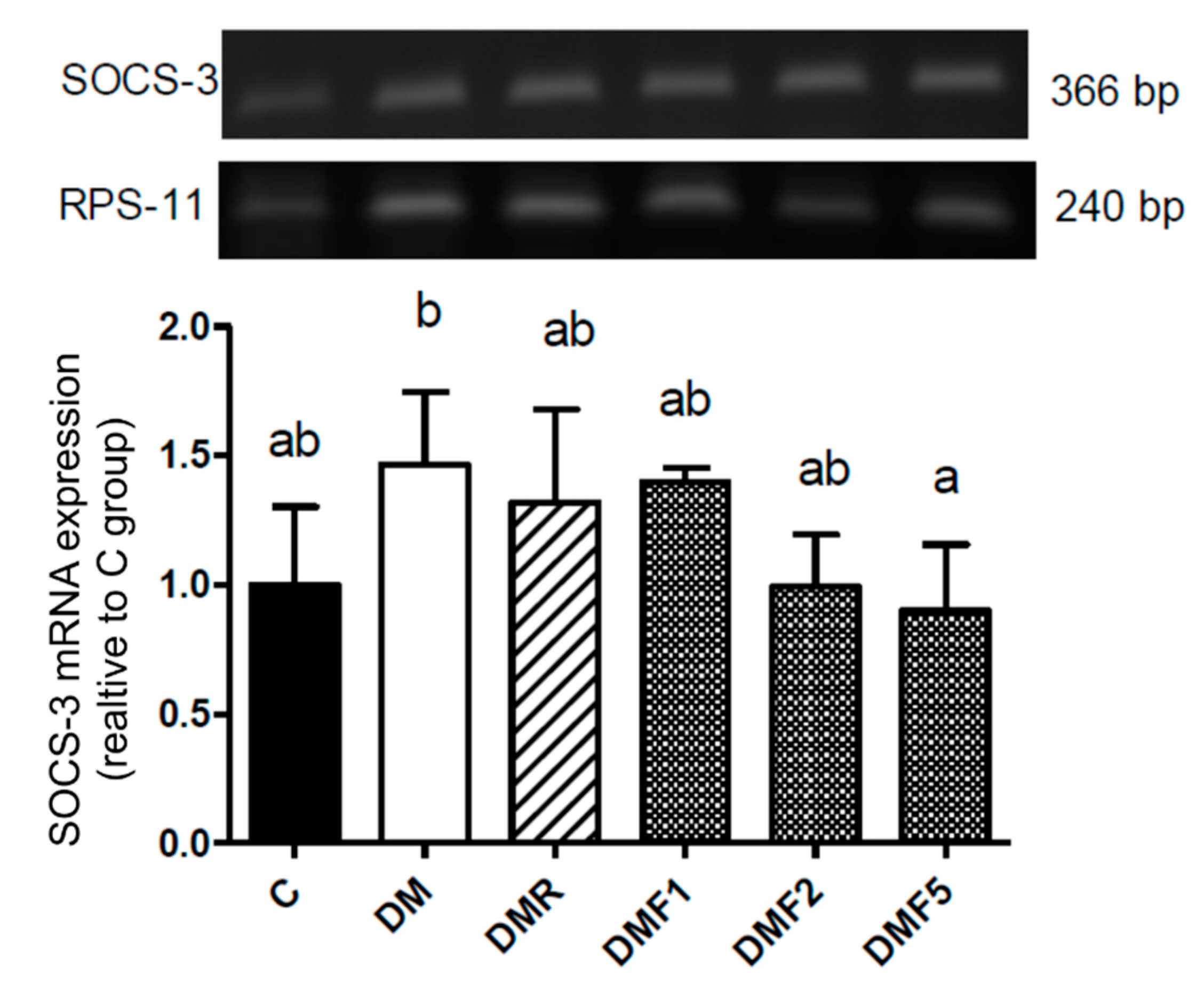
2.5. Effects of Fucoxanthin on SOCS-3 mRNA Expression
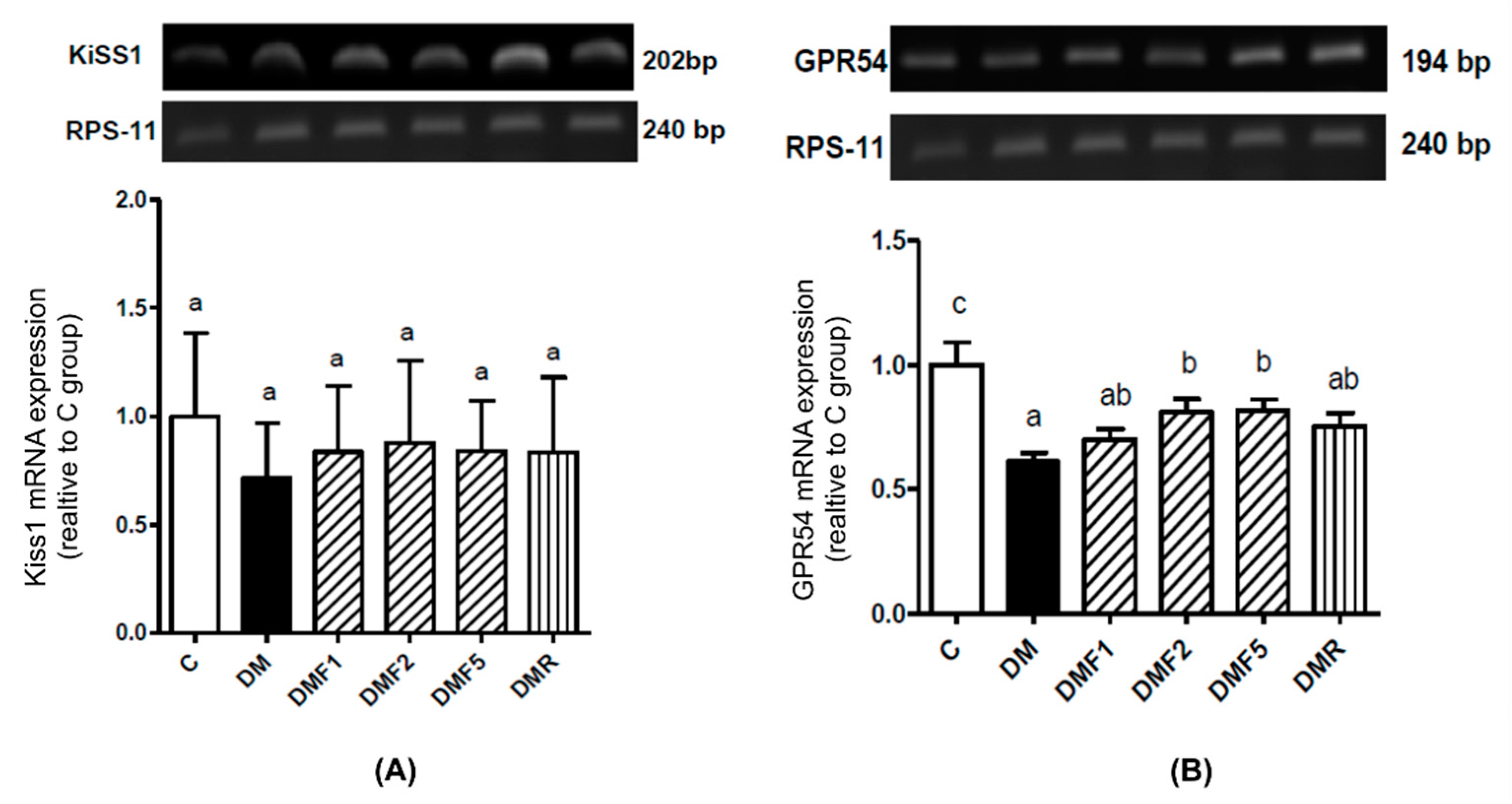
2.6. Effects of Fucoxanthin on Kiss1 and GPR54 mRNA Expression
2.7. Effect of Fucoxanthin on Plasma Reproductive Hormones
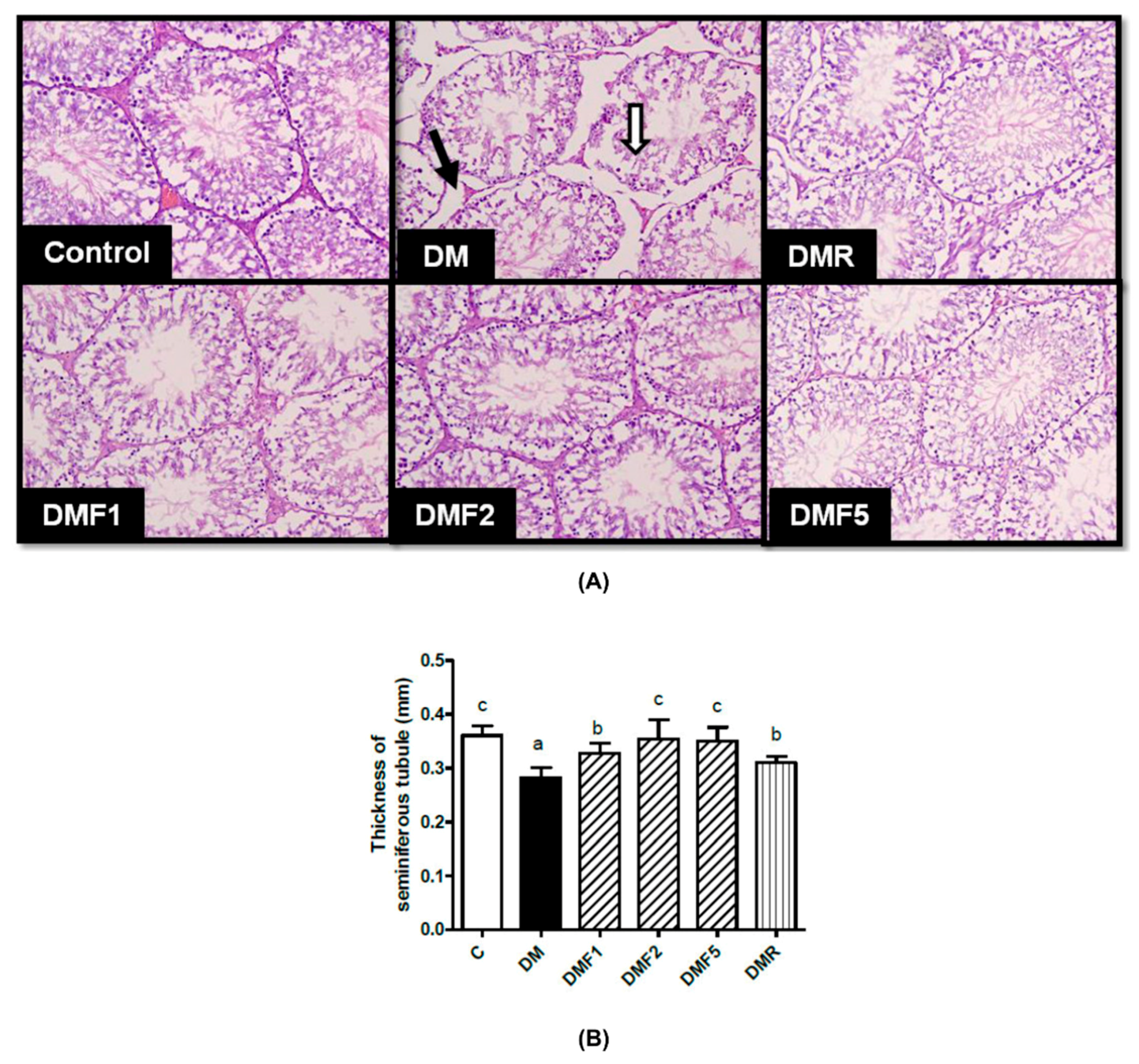
2.8. Effects of Fucoxanthin on Sperm Properties and Testicular Histology
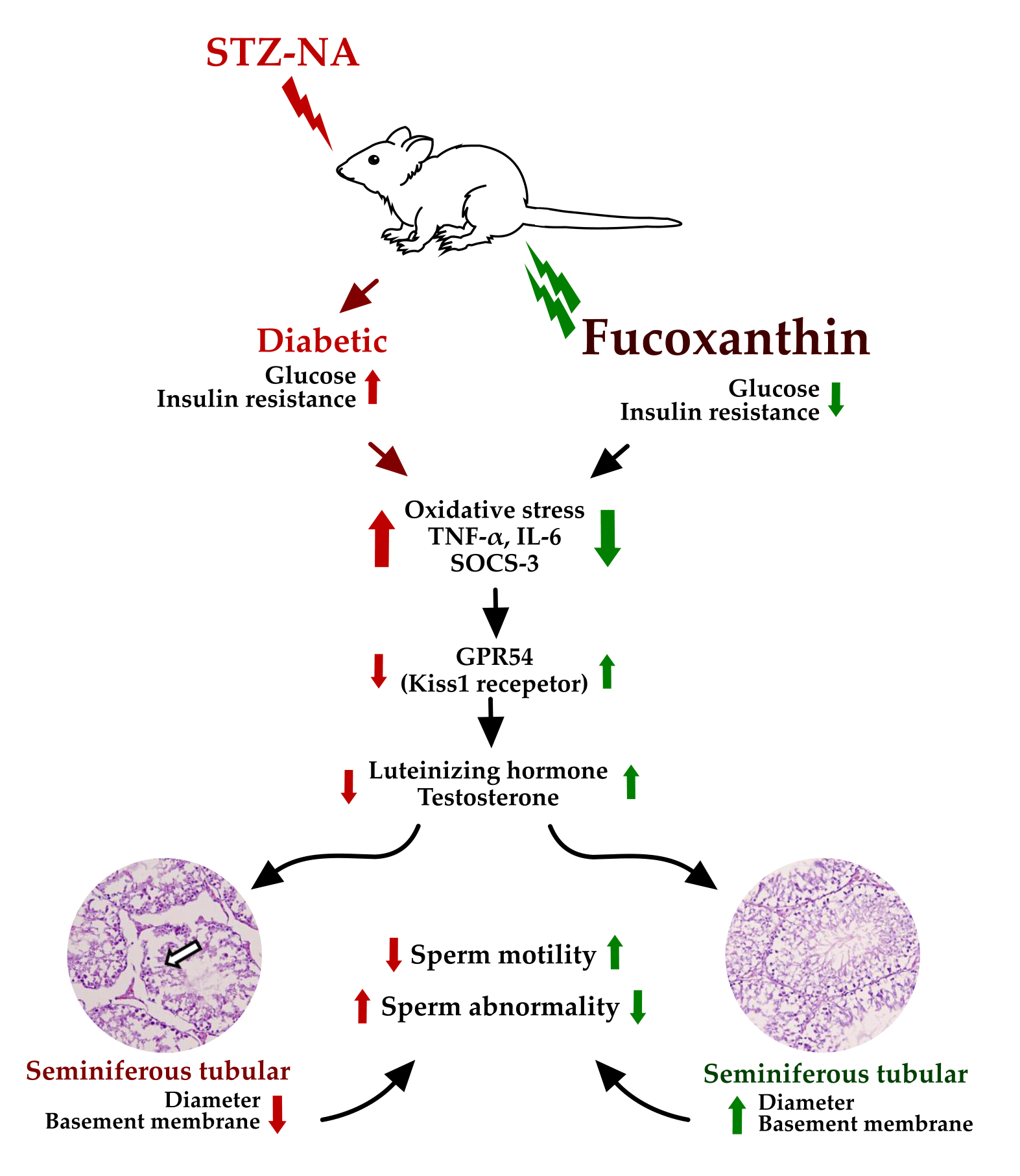
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. RAW 264.7 Macrophage Cells Study
4.2.1. Cell Culture
4.2.2. Cells Viability and Nitric Oxide Assay
4.2.3. Reactive Oxygen Species (ROS) Assay In Vitro
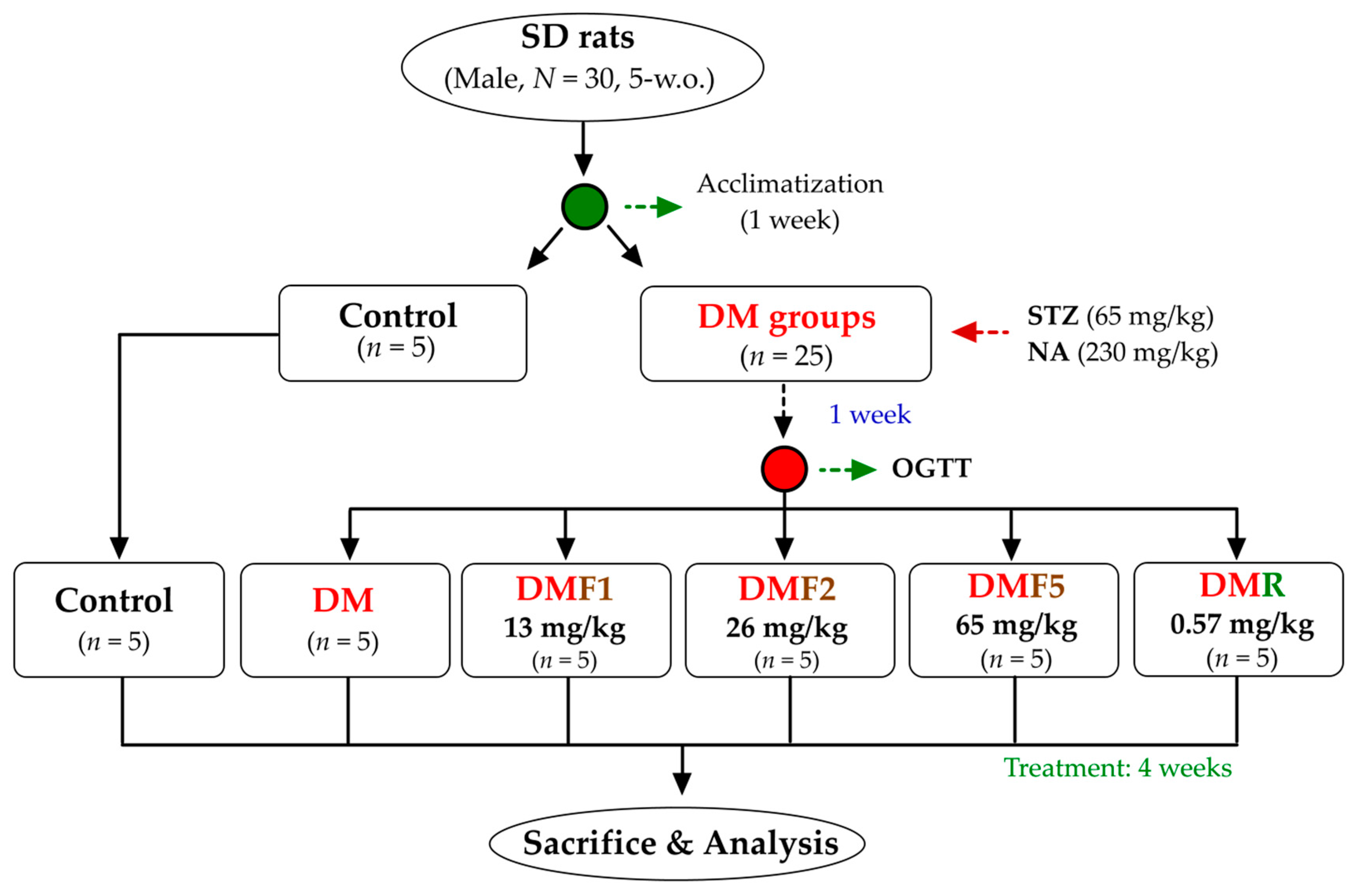
4.3. Animal Experiment
4.4. Blood Collection, Supernatant Homogenized Tissue, and Sperm Cells Preparation
4.5. Reactive Oxygen Species Analysis
4.6. Glucose, Insulin, Proinflammatory Cytokines, and Hormones Analysis
4.7. Kiss1, GPR54, and SOCS-3 mRNA Expression Analysis
4.8. Sperm Count and Morphology Assay
4.9. Testis Histopathological Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, P.; Fiaschi-Taesch, N.M.; Vasavada, R.C.; Scott, D.K.; García-Ocaña, A.; Stewart, A.F. Diabetes mellitus—Advances and challenges in human β-cell proliferation. Nat. Rev. Endocrinol. 2015, 11, 201. [Google Scholar] [CrossRef]
- Aynalem, S.B.; Zeleke, A.J. Prevalence of diabetes mellitus and its risk factors among individuals aged 15 years and above in Mizan-Aman town, southwest Ethiopia, 2016: A cross sectional study. Int. J. Endocrinol. 2018, 2018, 1–7. [Google Scholar] [CrossRef]
- AlSharari, S.D.; Al-Rejaie, S.S.; Abuohashish, H.M.; Aleisa, A.M.; Parmar, M.Y.; Ahmed, M.M. Ameliorative potential of morin in streptozotocin-induced neuropathic pain in rats. Trop. J. Pharm. Res. 2014, 13, 1429–1436. [Google Scholar] [CrossRef]
- Dogan, Y.; Akarsu, S.; Ustundag, B.; Yilmaz, E.; Gurgoze, M.K. Serum il-1β, il-2, and il-6 in insulin-dependent diabetic children. Mediat. Inflamm. 2006, 2006, 1–6. [Google Scholar] [CrossRef]
- Poitout, V.; Robertson, R.P. Minireview: Secondary β-cell failure in type 2 diabetes—A convergence of glucotoxicity and lipotoxicity. Endocrinology 2002, 143, 339–342. [Google Scholar] [CrossRef]
- Abd El-Twab, S.M.; Mohamed, H.M.; Mahmoud, A.M. Taurine and pioglitazone attenuate diabetes-induced testicular damage by abrogation of oxidative stress and up-regulation of the pituitary–gonadal axis. Can. J. Physiol. Pharmacol. 2016, 94, 651–661. [Google Scholar] [CrossRef]
- Aksu, I.; Baykara, B.; Kiray, M.; Gurpinar, T.; Sisman, A.R.; Ekerbicer, N.; Tas, A.; Gokdemir-Yazar, O.; Uysal, N. Serum igf-1 levels correlate negatively to liver damage in diabetic rats. Biotech. Histochem. 2013, 88, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Dhindsa, S.; Chandel, A.; Chaudhuri, A. Hypogonadotropic hypogonadism in men with type 2 diabetes. Postgrad. Med. 2015, 121, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Agbaje, I.M.; Rogers, D.A.; McVicar, C.M.; McClure, N.; Atkinson, A.B.; Mallidis, C.; Lewis, S.E.M. Insulin dependant diabetes mellitus: Implications for male reproductive function. Hum. Reprod. 2007, 22, 1871–1877. [Google Scholar] [CrossRef]
- Marín-Peñalver, J.J.; Martín-Timón, I.; Sevillano-Collantes, C.; Cañizo-Gómez, F.J.D. Update on the treatment of type 2 diabetes mellitus. World J. Diabetes 2016, 7, 354–395. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical review of antidiabetic drugs: Implications for type 2 diabetes mellitus management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Tsukui, T.; Sashima, T.; Hosokawa, M.; Miyashita, K. Seaweed carotenoid, fucoxanthin, as a multi-functional nutrient. Asia Pac. J. Clin. Nutr. 2008, 17, 196–199. [Google Scholar] [PubMed]
- Maeda, H. Nutraceutical effects of fucoxanthin for obesity and diabetes therapy: A review. J. Oleo Sci. 2015, 64, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Shnimizu, M.; Moriwaki, H. Cancer chemoprevention by carotenoids. Molecules 2012, 17, 3202–3242. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, N.; Gemello, E.; Gammone, M.; de Girolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A treasure from the sea. Mar. Drugs 2012, 10, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, G. Marine carotenoids and oxidative stress. Mar. Drugs 2012, 10, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.-L.; Kao, N.-J.; Hu, J.-Y.; Wu, C.-S. Fucoxanthin-rich brown algae extract decreases inflammation and attenuates colitis-associated colon cancer in mice. J. Food Nutr. Res. 2016, 4, 137–147. [Google Scholar]
- Wei, T.; Chen, C.; Hou, J.; Xin, W.; Mori, A. Nitric oxide induces oxidative stress and apoptosis in neuronal cells. Biochim. Biophys. Acta Mol. Cell Res. 2000, 1498, 72–79. [Google Scholar] [CrossRef]
- Pierini, D.; Bryan, N.S. Nitric oxide availability as a marker of oxidative stress. In Advanced Protocols in Oxidative Stress III; Humana Press: New York, NY, USA, 2015; pp. 63–71. [Google Scholar]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2008, 15, 252–259. [Google Scholar] [CrossRef]
- Wang, X.; Cui, Y.-J.; Qi, J.; Zhu, M.-M.; Zhang, T.-L.; Cheng, M.; Liu, S.-M.; Wang, G.-C. Fucoxanthin exerts cytoprotective effects against hydrogen peroxide-induced oxidative damage in l02 cells. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Kaneko, M.; Nagamine, T.; Nakazato, K.; Mori, M. The anti-apoptotic effect of fucoxanthin on carbon tetrachloride-induced hepatotoxicity. J. Toxicol. Sci. 2013, 38, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Masiello, P.; Broca, C.; Gross, R.; Roye, M.; Manteghetti, M.; Hillaire-Buys, D.; Novelli, M.; Ribes, G. Experimental niddm: Development of a new model in adult rats administered streptozotocin and nicotinamide. Diabetes 1998, 47, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Olokoba, A.B.; Obateru, O.A.; Olokoba, L.B. Type 2 diabetes mellitus: A review of current trends. Oman Med. J. 2012, 27, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Gayoso-Diz, P.; Otero-González, A.; Rodriguez-Alvarez, M.X.; Gude, F.; García, F.; De Francisco, A.; Quintela, A.G. Insulin resistance (homa-ir) cut-off values and the metabolic syndrome in a general adult population: Effect of gender and age: Epirce cross-sectional study. BMC Endocr. Disord. 2013, 13, 47. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 1–31. [Google Scholar] [CrossRef]
- Shim, S.-Y.; Kim, H.-S. Oxidative stress and the antioxidant enzyme system in the developing brain. Korean J. Pediatrics 2013, 56, 107. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.-J.; Seo, Y.-J.; Pan, C.-H.; Lee, O.-H.; Kim, K.-J.; Lee, B.-Y. Fucoxanthin suppresses lipid accumulation and ros production during differentiation in 3t3-l1 adipocytes. Phytother. Res. 2016, 30, 1802–1808. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Turner, T.T.; Lysiak, J.J. Oxidative stress: A common factor in testicular dysfunction. J. Androl. 2008, 29, 488–498. [Google Scholar] [CrossRef]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Mens Health 2014, 32, 1–17. [Google Scholar] [CrossRef]
- Gonzalez, Y.; Herrera, M.T.; Soldevila, G.; Garcia-Garcia, L.; Fabián, G.; Pérez-Armendariz, E.M.; Bobadilla, K.; Guzmán-Beltrán, S.; Sada, E.; Torres, M. High glucose concentrations induce tnf-α production through the down-regulation of cd33 in primary human monocytes. BMC Immunol. 2012, 13, 19. [Google Scholar] [CrossRef]
- Navarro-Gonzalez, J.F.; Mora-Fernandez, C. The role of inflammatory cytokines in diabetic nephropathy. J. Am. Soc. Nephrol. 2008, 19, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Atta, M.; Almadaly, E.; El-Far, A.; Saleh, R.; Assar, D.; Al Jaouni, S.; Mousa, S. Thymoquinone defeats diabetes-induced testicular damage in rats targeting antioxidant, inflammatory and aromatase expression. Int. J. Mol. Sci. 2017, 18, 919. [Google Scholar] [CrossRef]
- Rieusset, J.; Bouzakri, K.; Chevillotte, E.; Ricard, N.; Jacquet, D.; Bastard, J.P.; Laville, M.; Vidal, H. Suppressor of cytokine signaling 3 expression and insulin resistance in skeletal muscle of obese and type 2 diabetic patients. Diabetes 2004, 53, 2232–2241. [Google Scholar] [CrossRef]
- Colledge, W.H. Gpr54 and kisspeptins. In Orphan g Protein-Coupled Receptors and Novel Neuropeptides; Springer: Berlin, Germany, 2008; pp. 117–143. [Google Scholar]
- Sarchielli, E.; Comeglio, P.; Squecco, R.; Ballerini, L.; Mello, T.; Guarnieri, G.; Idrizaj, E.; Mazzanti, B.; Vignozzi, L.; Gallina, P.; et al. Tumor necrosis factor α impairs kisspeptin signaling in human gonadotropin-releasing hormone primary neurons. J. Clin. Endocrinol. Metab. 2016, 102, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Dao, H.; Wang, M.; Heston, A.; Garcia, K.M.; Sangal, A.; Dowling, A.R.; Faulkner, L.D.; Molitor, S.C.; Elias, C.F.; et al. Insulin and leptin signaling interact in the mouse kiss1 neuron during the peripubertal period. PLoS ONE 2015, 10, e0121974. [Google Scholar] [CrossRef] [PubMed]
- Schoeller, E.L.; Schon, S.; Moley, K.H. The effects of type 1 diabetes on the hypothalamic, pituitary and testes axis. Cell Tissue Res. 2012, 349, 839–847. [Google Scholar] [CrossRef]
- Abbara, A.; Ratnasabapathy, R.; Jayasena, C.N.; Dhillo, W.S. The effects of kisspeptin on gonadotropin release in non-human mammals. In Kisspeptin Signaling in Reproductive Biology; Springer: New York, NY, USA, 2013; Volume 784, pp. 63–87. [Google Scholar]
- Kilarkaje, N.; Al-Hussaini, H.; Al-Bader, M.M. Diabetes-induced DNA damage and apoptosis are associated with poly (adp ribose) polymerase 1 inhibition in the rat testis. Eur. J. Pharmacol. 2014, 737, 29–40. [Google Scholar] [CrossRef]
- Sisman, A.R.; Kiray, M.; Camsari, U.M.; Evren, M.; Ates, M.; Baykara, B.; Aksu, I.; Guvendi, G.; Uysal, N. Potential novel biomarkers for diabetic testicular damage in streptozotocin-induced diabetic rats: Nerve growth factor beta and vascular endothelial growth factor. Dis. Markers 2014, 2014, 1–7. [Google Scholar] [CrossRef]
- Orman, D.; Vardi, N.; Ates, B.; Taslidere, E.; Elbe, H. Aminoguanidine mitigates apoptosis, testicular seminiferous tubules damage, and oxidative stress in streptozotocin-induced diabetic rats. Tissue Cell 2015, 47, 284–290. [Google Scholar] [CrossRef]
- La Vignera, S.; Condorelli, R.A.; Di Mauro, M.; Lo Presti, D.; Mongioì, L.M.; Russo, G.; Calogero, A.E. Reproductive function in male patients with type 1 diabetes mellitus. Andrology 2015, 3, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Mayerson, A.B.; Hundal, R.S.; Dufour, S.; Lebon, V.; Befroy, D.; Cline, G.W.; Enocksson, S.; Inzucchi, S.E.; Shulman, G.I.; Petersen, K.F. The effects of rosiglitazone on insulin sensitivity, lipolysis, and hepatic and skeletal muscle triglyceride content in patients with type 2 diabetes. Diabetes 2002, 51, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shen, X. Effect of rosiglitazone on inflammatory cytokines and oxidative stress after intensive insulin therapy in patients with newly diagnosed type 2 diabetes. Diabetol. Metab. Syndr. 2019, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- Bach, R.G.; Brooks, M.M.; Lombardero, M.; Genuth, S.; Donner, T.W.; Garber, A.; Kennedy, L.; Monrad, E.S.; Pop-Busui, R.; Kelsey, S.F.; et al. Rosiglitazone and outcomes for patients with diabetes mellitus and coronary artery disease in the bypass angioplasty revascularization investigation 2 diabetes (bari 2d) trial. Circulation 2013, 128, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Nissen, S.E.; Wolski, K. Effect of rosiglitazone on the risk of myocardial infarction and death from cardiovascular causes. N. Engl. J. Med. 2007, 356, 2457–2471. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.L.; Travaglini, M.T. A review of rosiglitazone in type 2 diabetes mellitus. Pharmacotherapy 2001, 21, 1082–1099. [Google Scholar] [CrossRef] [PubMed]
- Van de Loosdrecht, A.A.; Beelen, R.H.J.; Ossenkoppele, G.J.; Broekhoven, M.G.; Langenhuijsen, M.M.A.C. A tetrazolium-based colorimetric mtt assay to quantitate human monocyte mediated cytotoxicity against leukemic cells from cell lines and patients with acute myeloid leukemia. J. Immunol. Methods 1994, 174, 311–320. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15n]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Abbasihormozi, S.; Shahverdi, A.; Kouhkan, A.; Cheraghi, J.; Akhlaghi, A.A.; Kheimeh, A. Relationship of leptin administration with production of reactive oxygen species, sperm DNA fragmentation, sperm parameters and hormone profile in the adult rat. Arch. Gynecol. Obstet. 2012, 287, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Kim, J.W.; Cha, Y.N.; Kim, C. A quantitative nitroblue tetrazolium assay for determining intracellular superoxide anion production in phagocytic cells. J. Immunoass. Immunochem. 2006, 27, 31–44. [Google Scholar] [CrossRef]
- Bartoli, E.; Fra, G.P.; Schianca, G.P.C. The oral glucose tolerance test (ogtt) revisited. Eur. J. Intern. Med. 2011, 22, 8–12. [Google Scholar] [CrossRef]
- Yu, Z.; Kastenmüller, G.; He, Y.; Belcredi, P.; Möller, G.; Prehn, C.; Mendes, J.; Wahl, S.; Roemisch-Margl, W.; Ceglarek, U.; et al. Differences between human plasma and serum metabolite profiles. PLoS ONE 2011, 6, e21230. [Google Scholar] [CrossRef]
- Sudirman, S.; Hsu, Y.-H.; He, J.-L.; Kong, Z.-L. Dietary polysaccharide-rich extract from eucheuma cottonii modulates the inflammatory response and suppresses colonic injury on dextran sulfate sodium-induced colitis in mice. PLoS ONE 2018, 13, e0205252. [Google Scholar] [CrossRef]
- Abdelali, A.; Al-Bader, M.; Kilarkaje, N. Effects of trans-resveratrol on hyperglycemia-induced abnormal spermatogenesis, DNA damage and alterations in poly (adp-ribose) polymerase signaling in rat testis. Toxicol. Appl. Pharmacol. 2016, 311, 61–73. [Google Scholar] [CrossRef]
- Younglai, E.V.; Holt, D.; Brown, P.; Jurisicova, A.; Casper, R.F. Sperm swim-up techniques and DNA fragmentation. Hum. Reprod. 2001, 16, 1950–1953. [Google Scholar] [CrossRef]
- Placer, Z.A.; Cushman, L.L.; Johnson, B.C. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 1966, 16, 359–364. [Google Scholar] [CrossRef]
- Wyrobek, A.J.; Bruce, W.R. Chemical induction of sperm abnormalities in mice. Proc. Natl. Acad. Sci. USA 1975, 72, 4425–4429. [Google Scholar] [CrossRef]
- Narayana, K.; D’Souza, U.J.A.; Seetharama Rao, K.P. Ribavirin-induced sperm shape abnormalities in wistar rat. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2002, 513, 193–196. [Google Scholar] [CrossRef]
- Taib, I.S.; Budin, S.B.; Ghazali, A.R.; Jayusman, P.A.; Louis, S.R.; Mohamed, J. Fenitrothion induced oxidative stress and morphological alterations of sperm and testes in male sprague-dawley rats. Clinics 2013, 68, 93–100. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | C | DM | DMF1 | DMF2 | DMF5 | DMR |
|---|---|---|---|---|---|---|
| FPG (mg/dL) | 153.25 ± 20.99 a | 211 ± 16.83 c | 180.47 ± 10.99 b | 174.81 ± 8.52 b | 181.55 ± 8.2 b | 182.72 ± 13.97 b |
| Insulin (µg/L) | 2.21 ± 0.75 a | 4.37 ± 0.63 b | 2.81 ± 0.59 a | 2.23 ± 0.87 a | 2.48 ± 0.78 a | 2.46 ± 0.60 a |
| HOMA-IR | 0.82 ± 0.27 a | 2.16 ± 0.39 b | 1.25 ± 0.19 a | 0.95 ± 0.42 a | 1.15 ± 0.39 a | 1.08 ± 0.18 a |
| Activities (U/mg Protein) | C | DM | DMF1 | DMF2 | DMF5 | DMR |
|---|---|---|---|---|---|---|
| Catalase | 38.72 ± 12.38 b | 22.12 ± 4.12 a | 29.57 ± 4.89 ab | 33.57 ± 9.18 b | 38.33 ± 4.55 b | 33.06 ± 9.42 ab |
| SOD | 0.56 ± 0.16 b | 0.12 ± 0.07 a | 0.42 ± 0.20 b | 0.40 ± 0.07 b | 0.42 ± 0.21 b | 0.54 ± 0.13 b |
| GPx | 216.74 ± 44.52 cd | 83.95 ± 59.08 a | 143.56 ± 43.70 ab | 161.83 ± 32.03 bc | 191.44 ± 46.18 bc | 265.01 ± 58.01 d |
| Activities (U/mg Protein) | C | DM | DMF1 | DMF2 | DMF5 | DMR |
|---|---|---|---|---|---|---|
| Catalase | 105.28 ± 8.59 b | 88.38 ± 6.30 a | 95.18 ± 8.45 ab | 94.36 ± 10.83 ab | 99.41 ± 12.68 ab | 89.39 ± 4.49 a |
| SOD | 29.71 ± 3.03 c | 15.35 ± 3.67 a | 16.83 ± 4.42 ab | 17.82 ± 2.71 ab | 22.78 ± 6.64 b | 22.78 ± 4.42 b |
| Hormones (ng/mL) | C | DM | DMF1 | DMF2 | DMF5 | DMR |
|---|---|---|---|---|---|---|
| LH | 0.91 ± 0.18 b | 0.75 ± 0.03 a | 0.96 ± 0.03 b | 1.16 ± 0.14 c | 1.19 ± 0.10 c | 0.85 ± 0.12 ab |
| Testosterone | 1.95 ± 0.04 b | 1.02 ± 0.24 a | 2.27 ± 0.93 b | 2.17 ± 1.03 b | 3.21 ± 0.76 c | 1.86 ± 0.34 ab |
| Sperm | C | DM | DMF1 | DMF2 | DMF5 | DMR |
|---|---|---|---|---|---|---|
| Total count (millions) | 495.22 ± 76.70 a | 405.64 ± 100.01 a | 557.60 ± 234.76 a | 493.2 ± 144.32 a | 540.8 ± 100.16 a | 542.00 ± 196.62 a |
| Motility (% total) | 19.23 ± 3.07 b | 11.07 ± 5.03 a | 15.91 ± 8.07 ab | 18.96 ± 6.21 b | 18.74 ± 2.39 b | 10.95 ± 2.95 a |
| Abnormal morphology (% total) | 8.43 ± 4.04 a | 15.76 ± 8.32 b | 8.45 ± 3.22 a | 10.13 ± 4.07 ab | 10.26 ± 1.56 ab | 9.88 ± 4.11 ab |
| mRNA | Primer Sequence | Expected Size (bp) | |
|---|---|---|---|
| Kisss1 | Sense | 5′-TGG CAC CTG TGG TGA ACC CTG AAC-3′ | 202 |
| Anti-sense | 5′-ATC AGG CGA CTG CGG GTG GCA CAC-3′ | ||
| GPR54 | Sense | 5′-TGT GCA AAT TCG TCA ACT ACA TCC-3′ | 194 |
| Anti-sense | 5′-AGC ACC GGG GCG GAA ACA GCT GC-3′ | ||
| SOCS-3 | Sense | 5′-CTG GAG CTG CCC GGG CCA GCC-3′ | 400 |
| Anti-sense | 5′-CAA GGC TGA CCA CAT CTG GG-3′ | ||
| RP-S11 | Sense | 5′-CAT TCA GAC GGA GCG TGC TTA C-3′ | 240 |
| Anti-sense | 5′-TGC ATC TTC ATC TTC GTC AC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, Z.-L.; Sudirman, S.; Hsu, Y.-C.; Su, C.-Y.; Kuo, H.-P. Fucoxanthin-Rich Brown Algae Extract Improves Male Reproductive Function on Streptozotocin-Nicotinamide-Induced Diabetic Rat Model. Int. J. Mol. Sci. 2019, 20, 4485. https://doi.org/10.3390/ijms20184485
Kong Z-L, Sudirman S, Hsu Y-C, Su C-Y, Kuo H-P. Fucoxanthin-Rich Brown Algae Extract Improves Male Reproductive Function on Streptozotocin-Nicotinamide-Induced Diabetic Rat Model. International Journal of Molecular Sciences. 2019; 20(18):4485. https://doi.org/10.3390/ijms20184485
Chicago/Turabian StyleKong, Zwe-Ling, Sabri Sudirman, Yu-Chun Hsu, Chieh-Yu Su, and Hsiang-Ping Kuo. 2019. "Fucoxanthin-Rich Brown Algae Extract Improves Male Reproductive Function on Streptozotocin-Nicotinamide-Induced Diabetic Rat Model" International Journal of Molecular Sciences 20, no. 18: 4485. https://doi.org/10.3390/ijms20184485
APA StyleKong, Z.-L., Sudirman, S., Hsu, Y.-C., Su, C.-Y., & Kuo, H.-P. (2019). Fucoxanthin-Rich Brown Algae Extract Improves Male Reproductive Function on Streptozotocin-Nicotinamide-Induced Diabetic Rat Model. International Journal of Molecular Sciences, 20(18), 4485. https://doi.org/10.3390/ijms20184485