Function and Transcriptional Regulation of Bovine TORC2 Gene in Adipocytes: Roles of C/EBPγ, XBP1, INSM1 and ZNF263

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
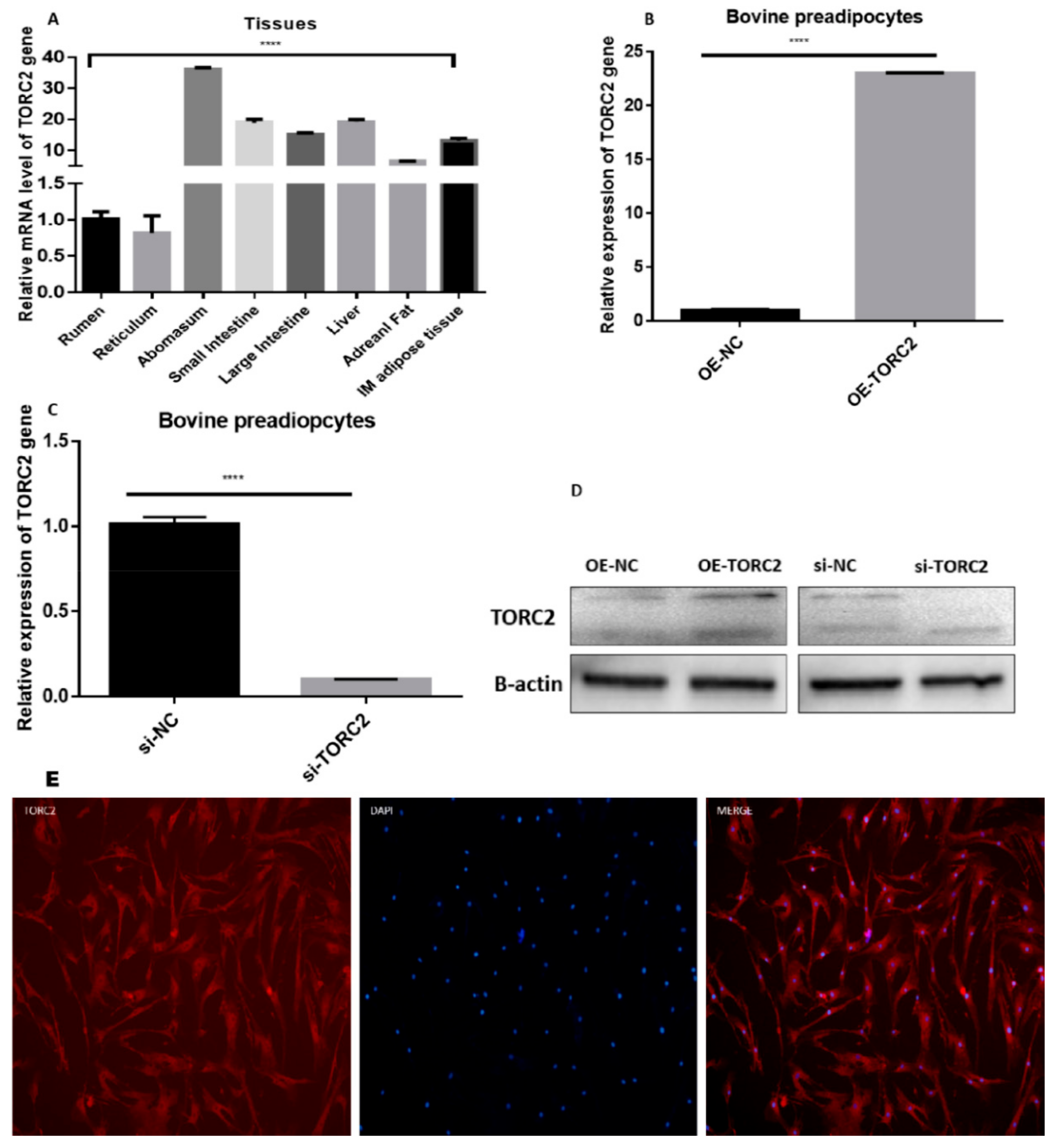
2.1. Transfection Efficiency, Tissues and Cellular Expression of TORC2 Gene
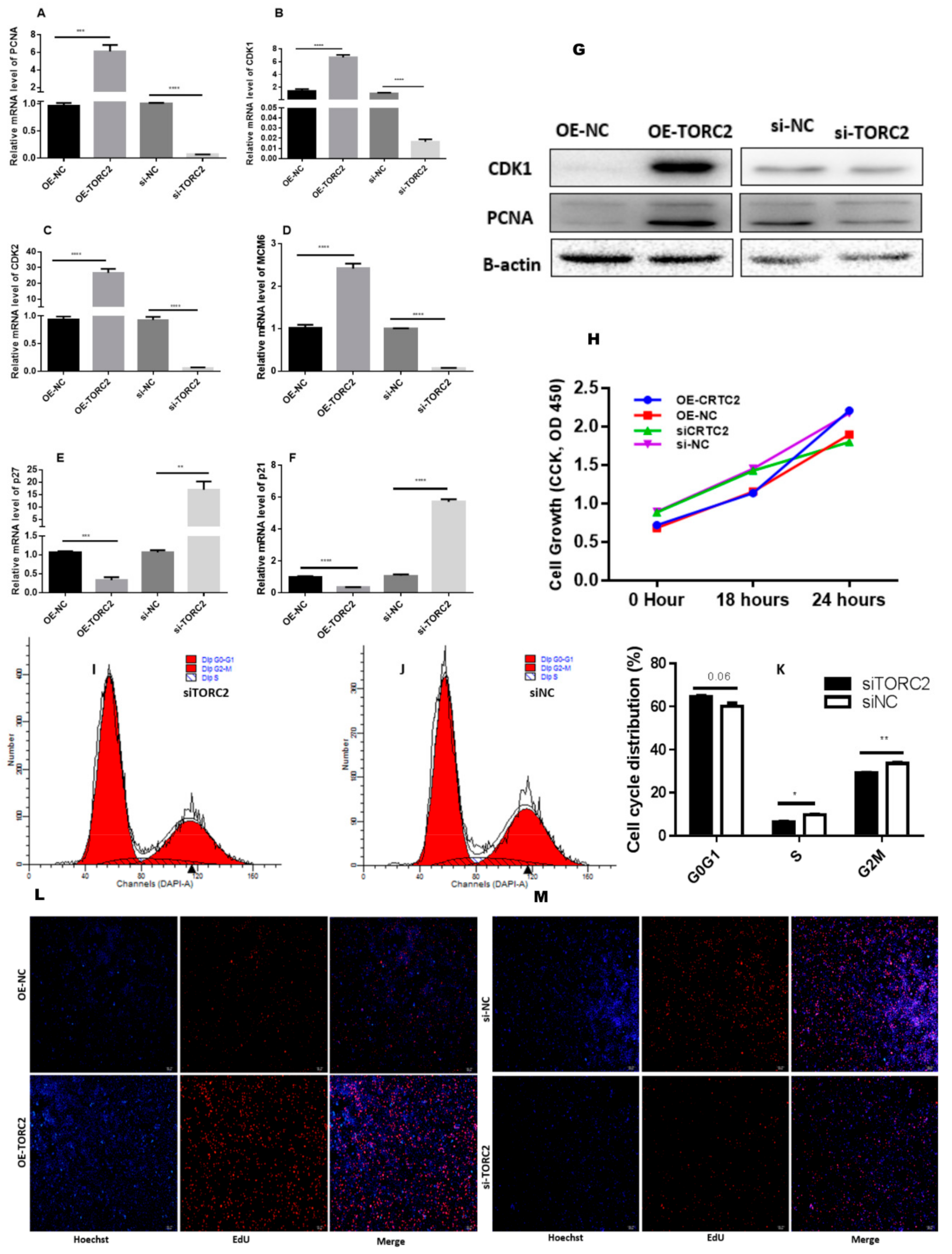
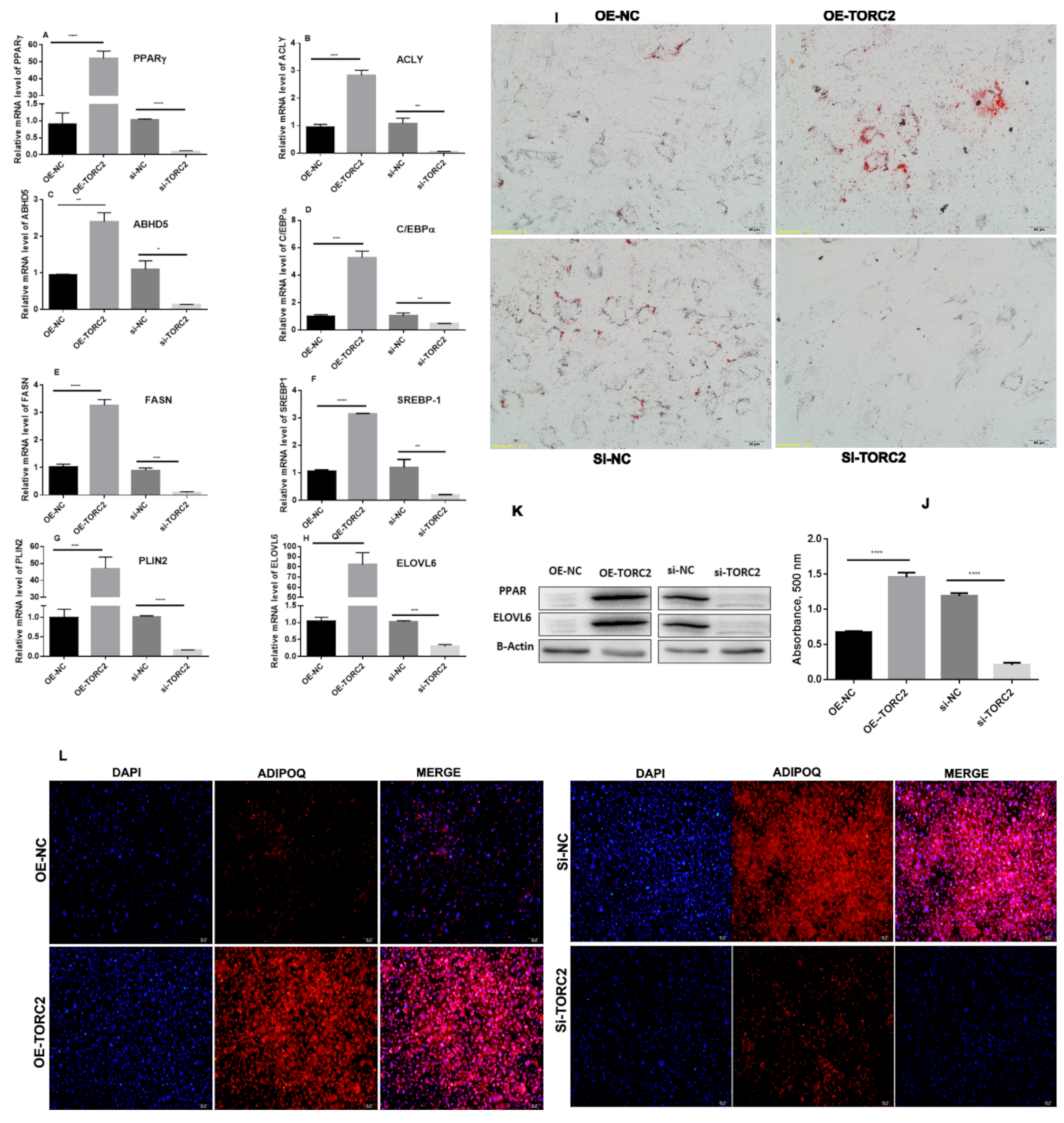
2.2. TORC2 Enhance Adipocyte Differentiation
2.3. Identification of Transcription Start Site (TSS) of the TORC2 Gene
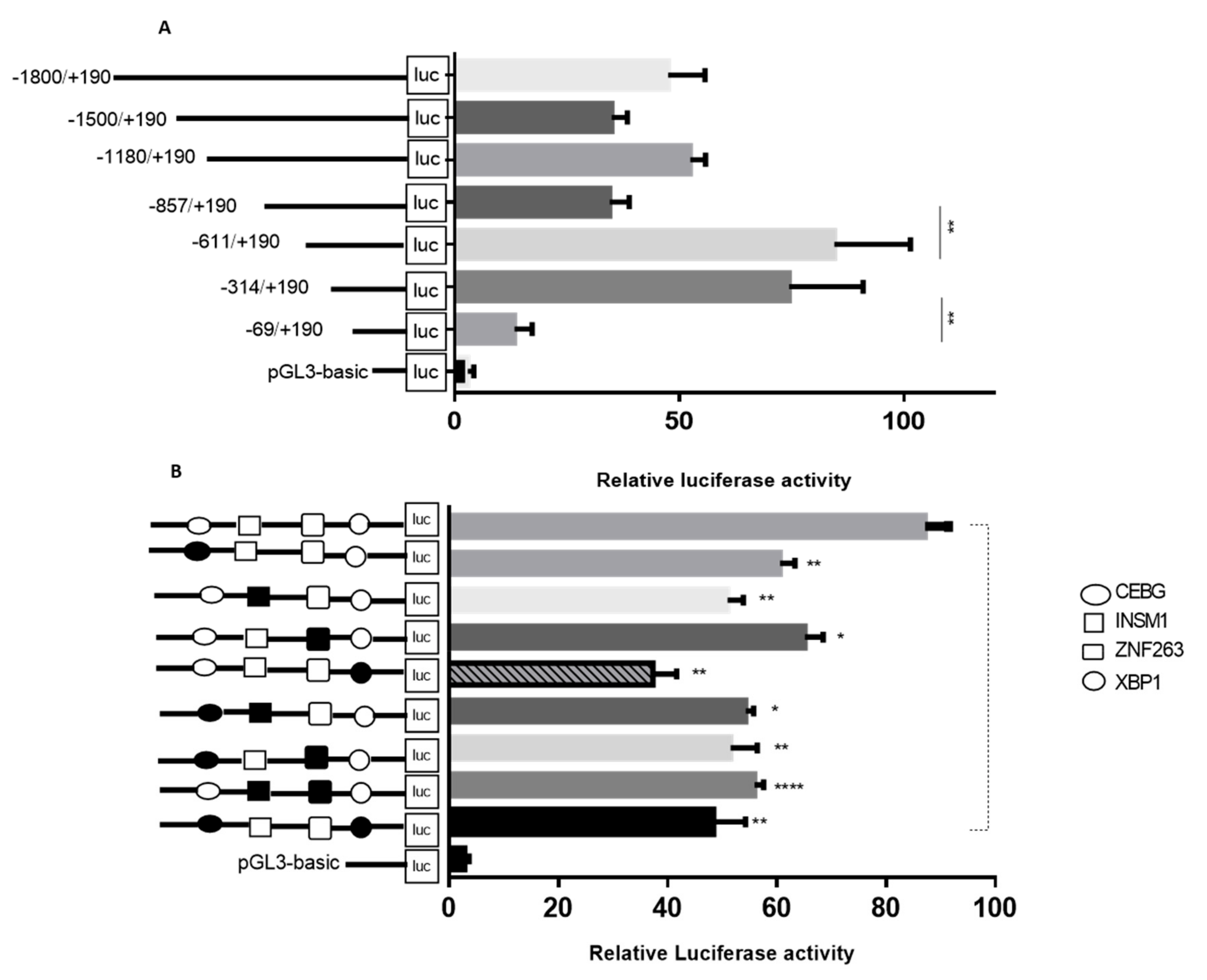
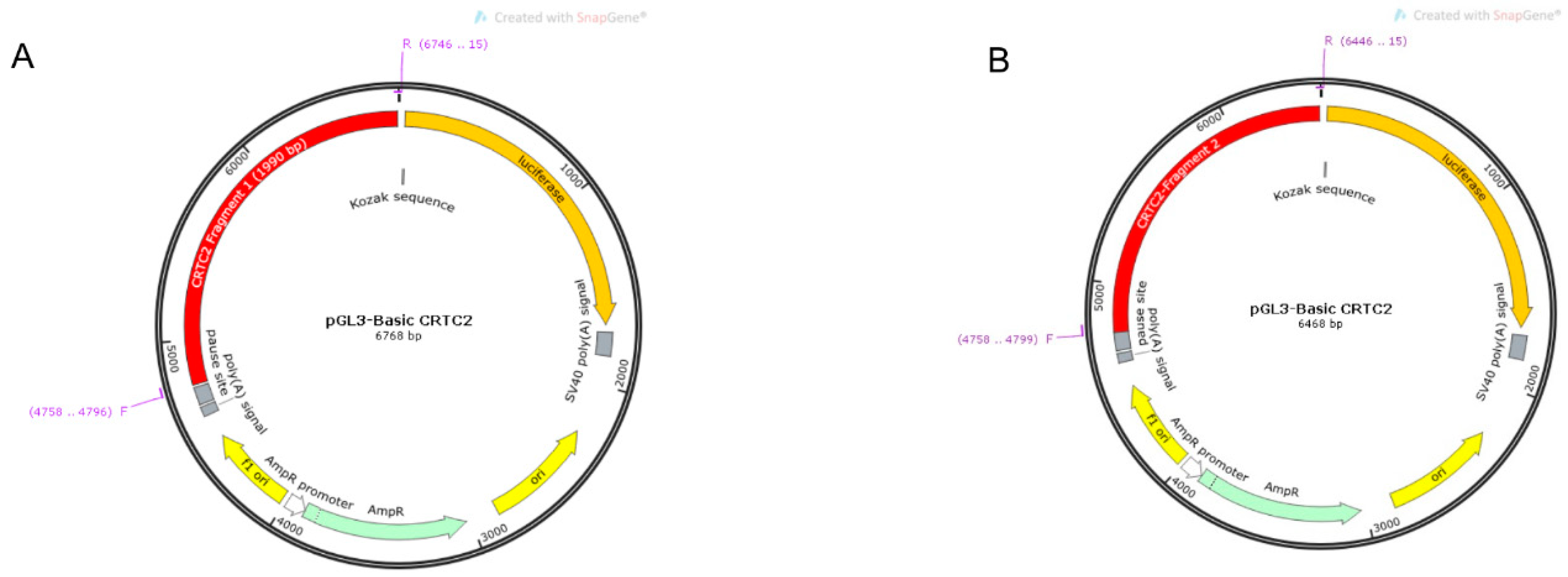
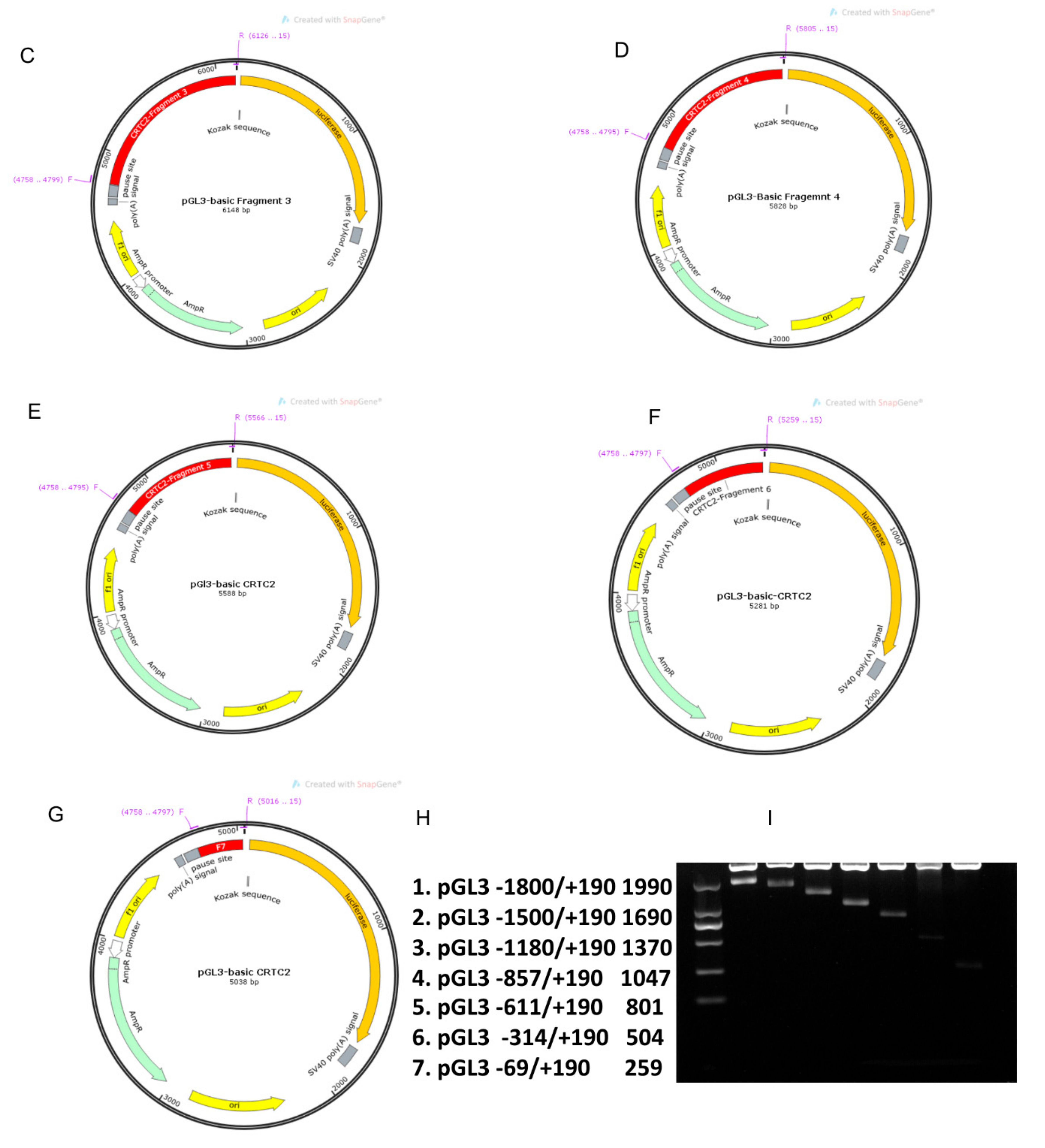
2.4. Identification of Core Promoter Region of TORC2 Gene
2.5. Roles of C/EBPγ, XBP1, INSM1 and ZNF263 in Transcriptional Regulation of TORC2 Gene
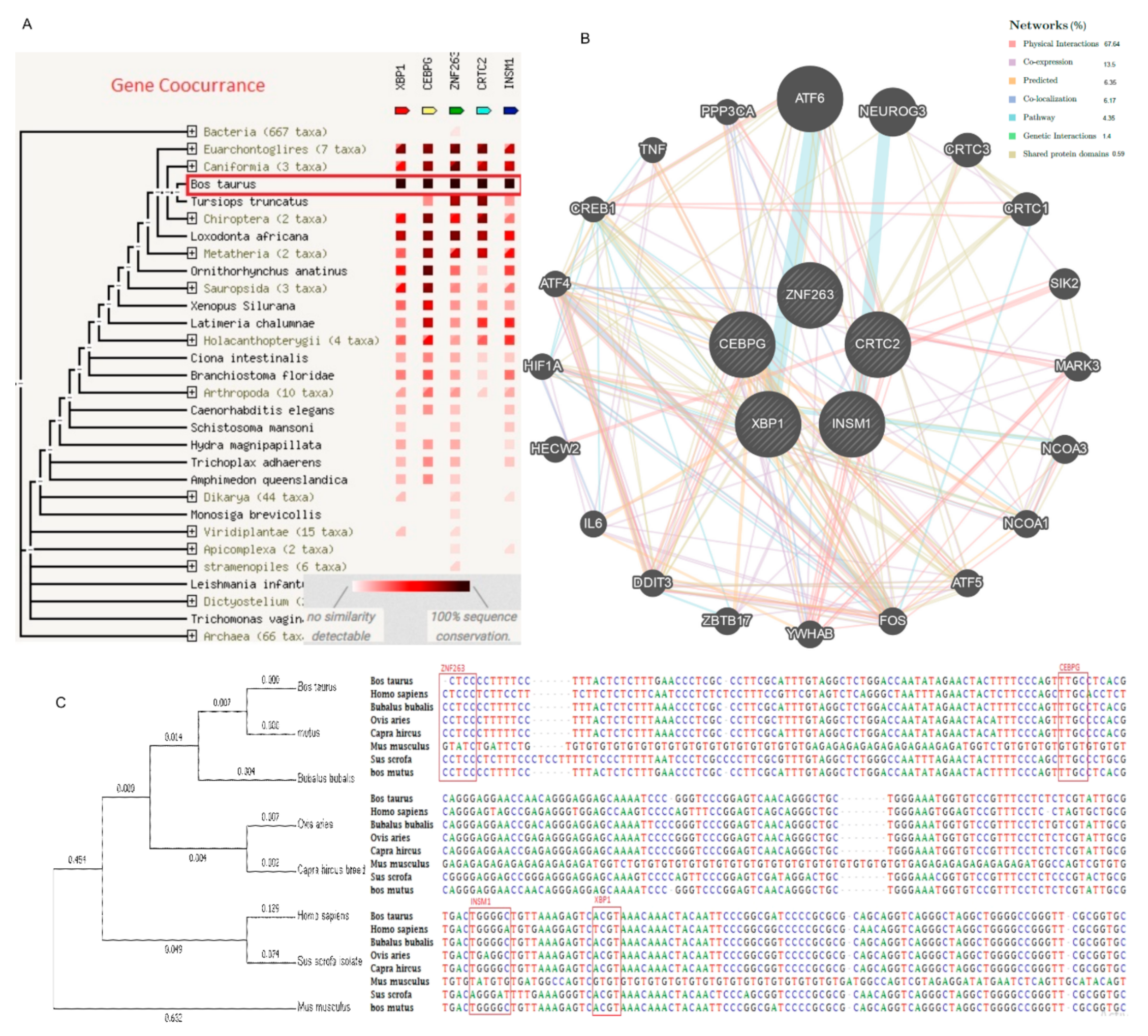
2.6. Genetic Interaction with Transcription Factors
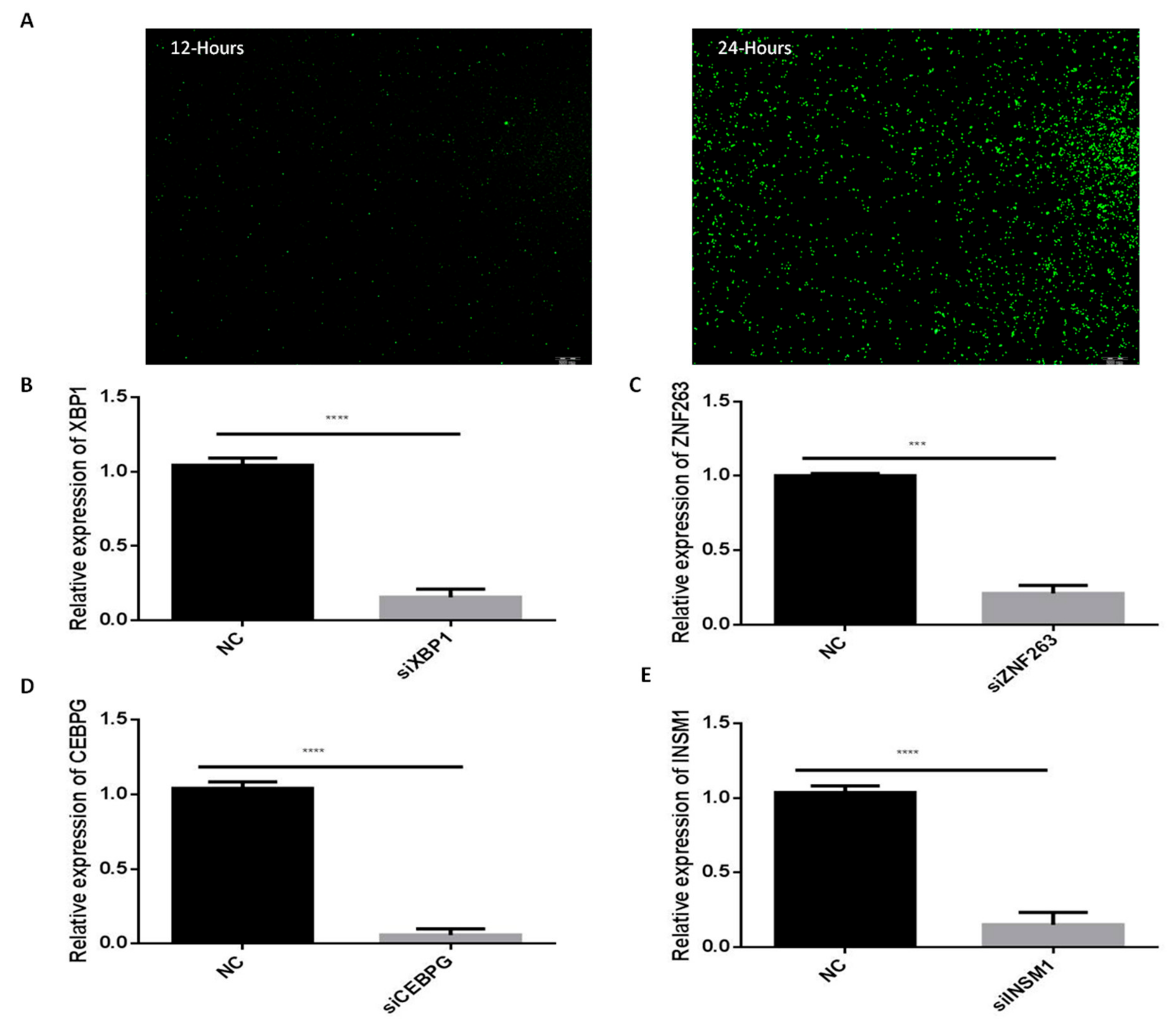
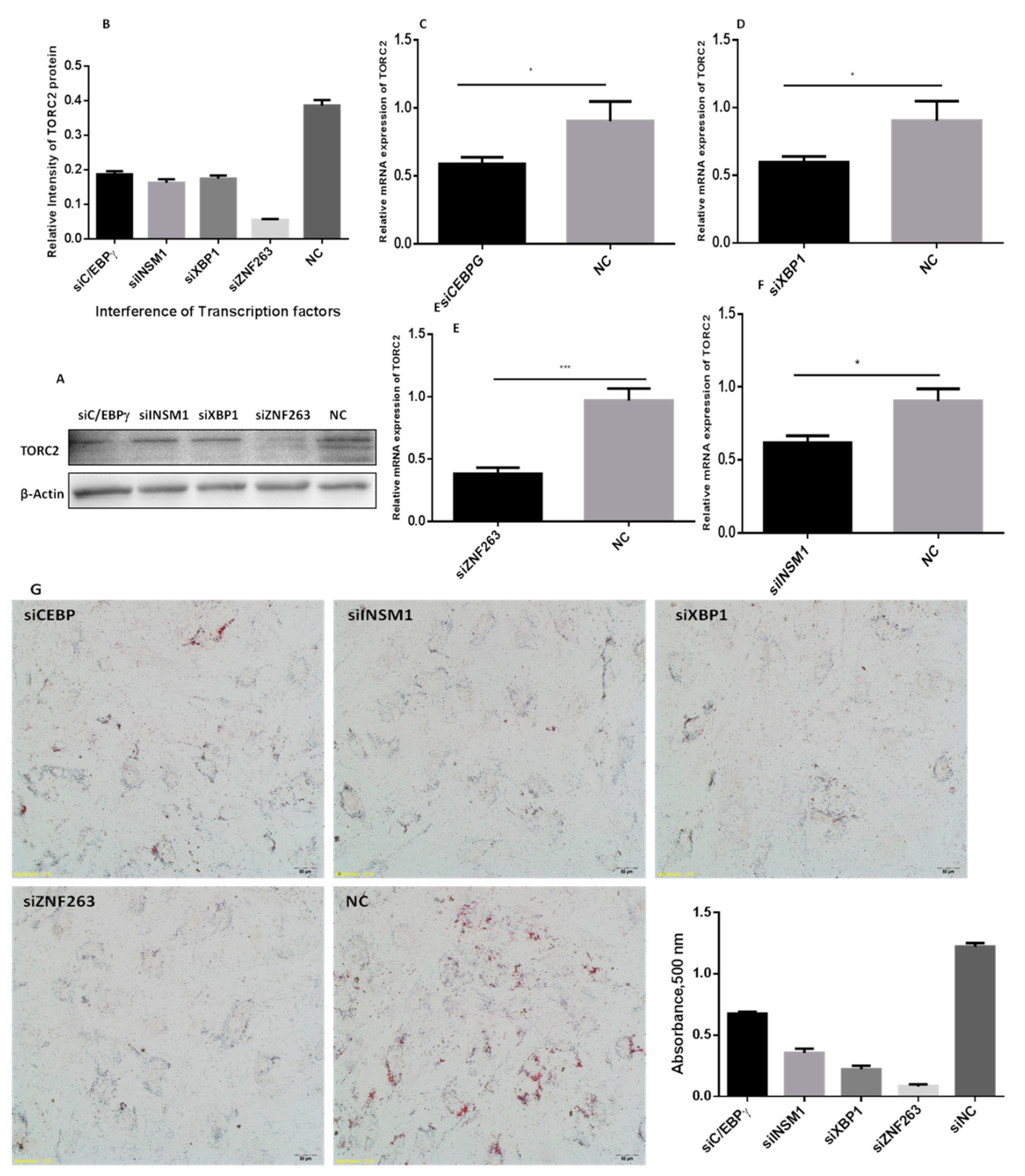
2.7. Silencing of C/EBPγ, ZNF263, XBP1 and INSM1 Transcription Factors
2.8. Oil Red O Staining
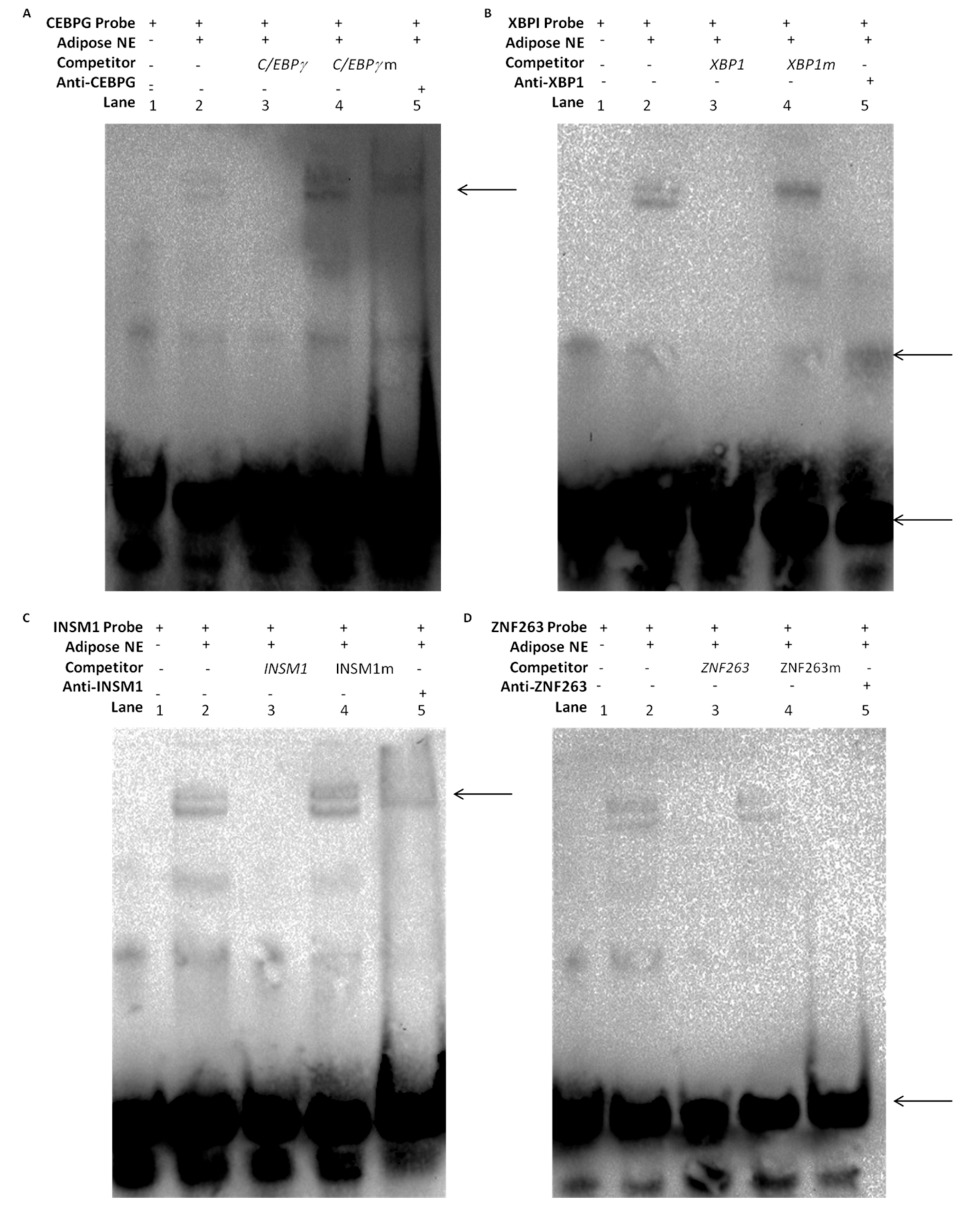
2.9. DNA-Protein Interaction through EMSAs
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Tissue Collection and mRNA Expression
4.3. Isolation of Bovine Primary Preadipocytes
4.4. Cell Culture and Immunofluorescence
4.5. EdU Proliferation Assay
4.6. Cell Cycle Assay through Flow Cytometry
4.7. CCK-8 Assay
4.8. 5′-Rapid Amplification of cDNA Ends (RACE)
4.9. DNA Extraction and Amplification of TORC2 Gene Promoter
4.10. Cloning of TORC2 Gene Promoter
4.11. Cell Culture and Transient Transfection
4.12. Mutagenesis in Transcription Factor Binding Sites
4.13. C/BEPγ, XBP1, ZNF263 and INSM1 Knockdown
4.14. Western Blot Analysis
4.15. Cell Differentiation and Oil Red O Staining
4.16. EMSAs (Electrophoretic Mobility Shift Assays)
4.17. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cai, H.; Li, M.; Sun, X.; Plath, M.; Li, C.; Lan, X.; Lei, C.; Huang, Y.; Bai, Y.; Qi, X.; et al. Global Transcriptome Analysis During Adipogenic Differentiation and Involvement of Transthyretin Gene in Adipogenesis in Cattle. Front. Genet. 2018, 9, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Martins, T.S.; Sanglard, L.M.P.; Silva, W.; Chizzotti, M.L.; Renno, L.N.; Serao, N.V.L.; Silva, F.F.; Guimaraes, S.E.F.; Ladeira, M.M.; Dodson, M.V.; et al. Molecular Factors Underlying the Deposition of Intramuscular Fat and Collagen in Skeletal Muscle of Nellore and Angus Cattle. PLoS ONE 2015, 10, e0139943. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, E.; Gotoh, T.; Ebara, F.; Xu, J.X.; Viergutz, T.; Nurnberg, G.; Maak, S.; Wegner, J. Cellular conditions for intramuscular fat deposition in Japanese Black and Holstein steers. Meat Sci. 2011, 89, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Nagai, K.; Nagata, Y.; Doronbekov, K.; Nishimura, S.; Yoshioka, S.; Fujita, T.; Shiga, K.; Miyake, T.; Taniguchi, Y.; et al. Exploration of Genes Showing Intramuscular Fat Deposition-Associated Expression Changes in Musculus Longissimus Muscle. Anim. Genet. 2006, 37, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Gondro, C.; van der Werf, J.; Kim, N.K.; Lim, D.J.; Park, E.W.; Oh, S.J.; Gibson, J.P.; Thompson, J.M. Use of a bovine genome array to identify new biological pathways for beef marbling in Hanwoo (Korean Cattle). BMC Genom. 2010, 11, 623–624. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.Y.; Chen, K.L.; Li, H.X.; Zhou, G.H. The adipokine Chemerin induces lipolysis and adipogenesis in bovine intramuscular adipocytes. Mol. Cell. Biochem. 2016, 418, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.M.; Ren, J.L.; Chen, L.; Zhang, X.; Cheng, M.L.; Li, W.Z.; Zhang, Y.Y.; Gao, S.Z. Differential Expression of Lipid Metabolism Related Genes in Porcine Muscle Tissue Leading to Different Intramuscular Fat Deposition. Lipids 2009, 44, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Lafontan, M.; Langin, D. Lipolysis and lipid mobilization in human adipose tissue. Prog. Lipid Res. 2009, 48, 275–295. [Google Scholar] [CrossRef]
- Altarejos, J.Y.; Montminy, M. CREB and the CRTC co-activators: Sensors for hormonal and metabolic signals. Nat. Rev. Mol. Cell Biol. 2011, 12, 141–151. [Google Scholar] [CrossRef]
- Screaton, R.A.; Conkright, M.D.; Katoh, Y.; Best, J.L.; Canettieri, G.; Jeffries, S.; Guzman, E.; Niessen, S.; Yates, J.R., III; Takemori, H.; et al. The CREB coactivator TORC2 functions as a calcium and cAMP sensitive coincidence detector. Cell 2004, 1, 61–74. [Google Scholar] [CrossRef]
- Lay, J.L.; Tuteja, G.; White, P.; Dhir, R.; Ahima, R.; Kaestner, K.H. CRTC2(TORC2) contributes to the transcriptional response to fasting in the liver but is not required for the maintenance of glucose homeostasis. Cell Metab. 2009, 10, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Samarajeewa, N.U.; Docanto, M.M.; Simpson, E.R.; Brown, K.A. CREB-Regulated Transcription Co-Activator Family Stimulates Promoter II-Driven Aromatase Expression in Preadipocytes. Horm. Cancer 2013, 4, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Hallenborg, P.; Siersbæk, M.; Barrio-Hernandez, I.; Nielsen, R.; Kristiansen, K.; Mandrup, S.; Grøntved, L.; Blagoev, B. MDM2 facilitates adipocyte differentiation through CRTC-mediated activation of STAT3. Cell Death Dis. 2016, 7, e2289–e2291. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.H.; Flechner, L.; Qi, L.; Zhang, X.; Screaton, R.A.; Jeffries, S.; Hedrick, S.; Xu, W.; Boussouar, F.; Brindle, P.K.; et al. The CREB coactivator TORC2 is a key regulator of fasting glucose metabolism. Nature 2005, 437, 1109–1111. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.; Kang, S. Diet change and exercise enhance protein expression of CREB, CRTC 2 and lipolitic enzymes in adipocytes of obese mice. Lipids Health Dis. 2016, 15, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Saberi, M.; Bjelica, D.; Schenk, S.; Imamura, T.; Bandyopadhyay, G.; Li, P.P.; Jadhar, V.; Vargeese, C.; Wang, W.M.; Bowman, K.; et al. Novel liver-specific TORC2 siRNA corrects hyperglycemia in rodent models of type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2009, 297, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Micah, J.; Hill, S.S.; James, H. Segars, and Tomoshige Kinocorresponding author. CRTC2 Is a Coactivator of GR and Couples GR and CREB in the Regulation of Hepatic Gluconeogenesis. Mol. Endocrinol. 2016, 30, 104–117. [Google Scholar] [CrossRef]
- Canettieri, G.; Koo, S.H.; Berdeaux, R.; Heredia, J.; Hedrick, S.; Zhang, X.; Montminy, M. Dual role of the coactivator TORC2 in modulating hepatic glucose output and insulin signaling. Cell Metab. 2005, 2, 331–338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dentin, R.; Hedrick, S.; Xie, J.X.; Yates, J.R., III; Montminy, M. Hepatic Glucose Sensing via the CREB Coactivator CRTC2. Science 2008, 319, 1402–1405. [Google Scholar] [CrossRef] [PubMed]
- Dentin, R.; Liu, Y.; Koo, S.H.; Hedrick, S.; Vargas, T.; Heredia, J.; Yates, J.R., III; Montminy, M. Insulin modulates gluconeogenesis by inhibition of the coactivator TORC2. Nature 2007, 449, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.-S.; Ryu, D.; Lee, M.-W.; Hong, S.; Koo, S.-H. Adiponectin and thiazolidinedione targets CRTC2 to regulate hepatic gluconeogenesis. Exp. Mol. Med. 2009, 41, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.V.; Accili, D. Hormonal regulation of hepatic glucose production in health and disease. Cell Metab. 2011, 14, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest. 2002, 1125–1131. [Google Scholar] [CrossRef]
- Evans, R.M.; Barish, G.D.; Wang, Y.X. PPARs and the complex journey to obesity. Nat. Med. 2004, 355–361. [Google Scholar] [CrossRef]
- Imai, T.; Takakuwa, R.; Marchand, S.; Dentz, E.; Bornert, J.M.; Messaddeq, N.; Wendling, O.; Mark, M.; Desvergne, B.; Wahli, W.; et al. Peroxisome proliferator-activated receptor gamma is required in mature white and brown adipocytes for their survival in the mouse. Proc. Natl. Acad. Sci. USA 2004, 101, 4543–4547. [Google Scholar] [CrossRef] [PubMed]
- Handschin, C.; Spiegelman, B.M. The role of exercise and PGC1alpha in inflammation and chronic disease. Nature 2008, 463–469. [Google Scholar] [CrossRef]
- Wang, Y.G.; Inoue, H.; Ravnskjaer, K.; Viste, K.; Miller, N.; Liu, Y.; Hedrick, S.; Vera, L.; Montminy, M. Targeted disruption of the CREB coactivator Crtc2 increases insulin sensitivity. Proc. Natl. Acad. Sci., USA. 2010, 107, 3087–3092. [Google Scholar] [CrossRef]
- Jiang, Z.; Kunej, T.; Michal, J.J.; Gaskins, C.T.; Reeves, J.J.; Busboom, J.R.; Dovc, P.; Wright , R.W., Jr. Significant associations of the mitochondrial transcription factor A promoter polymorphisms with marbling and subcutaneous fat depth in Wagyu x Limousin F2 crosses. Biochem. Biophys. Res. Commun. 2005, 334, 516–523. [Google Scholar] [CrossRef]
- Oury, M.P.; Picard, B.; Briand, M.; Blanquet, J.P.; Dumont, R. Interrelationships between meat quality traits, texture measurements and physicochemical characteristics of M. rectus abdominis from Charolais heifers. Meat Sci. 2009, 83, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Huang, J.; Zulfiqar, A.; Li, R.; Xi, Y.; Zhang, M.; Dang, R.; Lan, X.; Chen, H.; Ma, Y.; et al. Population structure and ancestry of Qinchuan cattle. Anim. Genet. 2018, 49, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Raza, S.H.A.; Zhang, J.; Gui, L.; Rahman, S.U.; Khan, R.; Hosseini, S.M.; Kaleri, H.A.; Zan, L. Polymorphism in promoter of SIX4 gene shows association with its transcription and body measurement traits in Qinchuan cattle. Gene 2018, 656, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Mei, C.; Wang, H.; Liao, Q.; Khan, R.; Raza, S.H.A.; Zhao, C.; Wang, H.; Cheng, G.; Tian, W.; Li, Y.; et al. Genome-wide analysis reveals the effects of artificial selection on production and meat quality traits in Qinchuan cattle. Genomics 2018. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Gui, L.; Khan, R.; Schreurs, N.M.; Xiaoyu, W.; Wu, S.; Mei, C.; Wang, L.; Ma, X.; Wei, D.; et al. Association between FASN gene polymorphisms ultrasound carcass traits and intramuscular fat in Qinchuan cattle. Gene 2018, 645, 55–59. [Google Scholar] [CrossRef]
- Guo, H.; Raza, S.H.A.; Schreurs, N.M.; Khan, R.; Wei, D.; Wang, L.; Zhang, S.; Zhang, L.; Wu, S.; Ullah, I.; et al. Genetic variants in the promoter region of the KLF3 gene associated with fat deposition in Qinchuan cattle. Gene 2018, 672, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Jung, C.H.; Jeon, T.I.; Ha, T.Y. MicroRNA-146b promotes adipogenesis by suppressing the SIRT1-FOXO1 cascade. EMBO Mol. Med. 2013, 5, 1602–1612. [Google Scholar] [CrossRef]
- Choi, S.; Kim, W.; Chung, J. Drosophila salt-inducible kinase (SIK) regulates starvation resistance through cAMP-response element-binding protein (CREB)-regulated transcription coactivator (CRTC). J. Biol. Chem. 2011, 286, 2658–2664. [Google Scholar] [CrossRef]
- Han, J.; Li, E.; Chen, L.; Zhang, Y.; Wei, F.; Liu, J.; Deng, H.; Wang, Y. The CREB coactivator CRTC2 controls hepatic lipid metabolism by regulating SREBP1. Nature 2015, 524, 243–246. [Google Scholar] [CrossRef]
- He, L.; Huang, N.; Li, H.; Tian, J.; Zhou, X.; Li, T.; Yao, K.; Wu, G.; Yin, Y. AMPK/alpha-Ketoglutarate Axis Regulates Intestinal Water and Ion Homeostasis in Young Pigs. J. Agric. Food Chem. 2017, 65, 2287–2298. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Yin, L.; Wang, C.; Wen, X. Adenovirus-mediated expression of SIK1 improves hepatic glucose and lipid metabolism in type 2 diabetes mellitus rats. PLoS One 2019, 14, e0210930. [Google Scholar] [CrossRef] [PubMed]
- Hogan, M.F.; Ravnskjaer, K.; Matsumura, S.; Huising, M.O.; Hull, R.L.; Kahn, S.E.; Montminy, M. Hepatic Insulin Resistance Following Chronic Activation of the CREB Coactivator CRTC2. J. Biol. Chem. 2015, 290, 25997–26006. [Google Scholar] [CrossRef] [PubMed]
- Goodman, M.N.; McElaney, M.A.; Ruderman, N.B. Adaptation to prolonged starvation in the rat: Curtailment of skeletal muscle proteolysis. Am. J. Physiol. 1981, 241, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Cahill, G.F., Jr. Fuel metabolism in starvation. Annu. Rev. Nutr. 2006, 26, 1–22. [Google Scholar] [CrossRef]
- Wang, Z.; Takemori, H.; Halder, S.K.; Nonaka, Y.; Okamoto, M. Cloning of a novel kinase (SIK) of the SNF1/AMPK family from high salt diet-treated rat adrenal. FEBS Lett. 1999, 135–139. [Google Scholar] [CrossRef]
- Sanosaka, M.; Fujimoto, M.; Ohkawara, T.; Nagatake, T.; Itoh, Y.; Kagawa, M.; Kumagai, A.; Fuchino, H.; Kunisawa, J.; Naka, T.; et al. Salt-inducible kinase 3 deficiency exacerbates lipopolysaccharide-induced endotoxin shock accompanied by increased levels of pro-inflammatory molecules in mice. Immunology 2015, 268–278. [Google Scholar] [CrossRef]
- Malm, H.A.; Mollet, I.G.; Berggreen, C.; Orho-Melander, M.; Esguerra, J.L.; Goransson, O.; Eliasson, L. Transcriptional regulation of the miR-212/miR-132 cluster in insulin-secreting beta-cells by cAMP-regulated transcriptional co-activator 1 and salt-inducible kinases. Mol. Cell. Endocrinol. 2016, 23–33. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell. Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Conkright, M.D.; Canettieri, G.; Screaton, R.; Guzman, E.; Miraglia, L.; Hogenesch, J.B.; Montminy, M. TORCs: Transducers of regulated CREB activity. Mol. Cell 2003, 12, 413–423. [Google Scholar] [CrossRef]
- Ravnskjaer, K.; Kester, H.; Liu, Y.; Zhang, X.; Lee, D.; Yates, J.R., III; Montminy, M. Cooperative interactions between CBP and TORC2 confer selectivity to CREB target gene expression. EMBO J. 2007, 26, 2880–2889. [Google Scholar] [CrossRef]
- Eberhard, C.E.; Fu, A.; Reeks, C.; Screaton, R.A. CRTC2 Is Required for β-Cell Function and Proliferation. Endocrinology 2013, 154, 2308–2317. [Google Scholar] [CrossRef] [PubMed]
- Bittinger, M.A.; McWhinnie, E.; Meltzer, J.; Iourgenko, V.; Latario, B.; Liu, X.; Chen, C.H.; Song, C.; Garza, D.; Labow, M. Activation of cAMP response element-mediated gene expression by regulated nuclear transport of TORC proteins. Curr. Biol. 2004, 14, 2156–2161. [Google Scholar] [CrossRef] [PubMed]
- Busby, S.; Ebright, R.H. Promoter structure, promoter recognition, and transcription activation in prokaryotes. Cell 1994, 79, 743–746. [Google Scholar] [CrossRef]
- Struhl, K. Fundamentally different logic of gene regulation in eukaryotes and prokaryotes. Cell 1999, 98, 1–4. [Google Scholar] [CrossRef]
- Sandelin, A.; Carninci, P.; Lenhard, B.; Ponjavic, J.; Hayashizaki, Y.; Hume, D.A. Mammalian RNA polymerase II core promoters: Insights from genome-wide studies. Nat. Rev. Genet. 2007, 8, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Orlando, U.; Cooke, M.; Maciel, F.C.; Papadopoulos, V.; Podestá, E.J.; Maloberti, P. Characterization of the mouse promoter region of the acyl-CoA synthetase 4 gene: Role of Sp1 and CREB. Mol. Cell. Endocrinol. 2013, 369, 15–36. [Google Scholar] [CrossRef]
- Wei, D.; Li, A.; Zhao, C.; Wang, H.; Mei, C.; Khan, R.; Zan, L. Transcriptional Regulation by CpG Sites Methylation in the Core Promoter Region of the Bovine SIX1 Gene: Roles of Histone H4 and E2F2. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Fang, M.; Park, M.L.; Chamberlain, L.; Xing, W.; Yu, H.; Green, M.R. The CREB Coactivator CRTC2 is a Lymphoma Tumor Suppressor that Preserves Genome Integrity Through Transcription of DNA Mismatch Repair Genes. Cell Rep. 2015, 11, 1350–1357. [Google Scholar] [CrossRef]
- Morgan, H.D.; Santos, F.; Green, K.; Dean, W.; Reik, W. Epigenetic reprogramming in mammals. Hum. Mol. Genet. 2005, 14, 47–58. [Google Scholar] [CrossRef]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wang, Y.; Ning, Y.; Guo, H.; Wang, X.; Zhang, L.; Khan, R.; Cheng, G.; Wang, H.; Zan, L. Genetic Variants in STAT3 Promoter Regions and Their Application in Molecular Breeding for Body Size Traits in Qinchuan Cattle. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Robertson, K.D. Genome Stability, Role of DNA Methylation in Genome Stability; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Yan, C.; Wang, X.; Cao, J.; Wu, M.; Gao, H. CCAAT/enhancer-binding protein gamma is a critical regulator of IL-1beta-induced IL-6 production in alveolar epithelial cells. PLoS ONE 2012, e35492. [Google Scholar] [CrossRef]
- Parkin, S.E.; Baer, M.; Copeland, T.D.; Schwartz, R.C.; Johnson, P.F. Regulation of CCAAT/enhancer-binding protein (C/EBP) activator proteins by heterodimerization with C/EBPgamma (Ig/EBP). J. Biol. Chem. 2002, 277, 23563–23572. [Google Scholar] [CrossRef] [PubMed]
- Huggins, C.J.; Malik, R.; Lee, S.; Salotti, J.; Thomas, S.; Martin, N.; Quiñones, O.A.; Alvord, W.G.; Olanich, M.E.; Keller, J.R.; et al. C/EBPγ suppresses senescence and inflammatory gene expression by heterodimerizing with C/EBPβ. Mol. Cell. Biol. 2013, 33, 3242–3258. [Google Scholar] [CrossRef] [PubMed]
- Gregor, M.F.; Hotamisligil, G.S. Thematic review series: Adipocyte Biology. Adipocyte stress: The endoplasmic reticulum and metabolic disease. J. Lipid Res. 2007, 48, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.L.; Cheng, D.D.; Zhou, S.M.; Zhu, B.; Hu, T.; Yang, Q.C. Overexpression of X-Box Binding Protein 1 (XBP1) Correlates to Poor Prognosis and Up-Regulation of PI3K/mTOR in Human Osteosarcoma. Int. J. Mol. Sci. 2015, 16, 28635–28646. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Zhang, T.; Saunee, N.A.; Breslin, M.B.; Song, K.J.; Lan, M.S. Functional role of an islet transcription factor, INSM1/IA-1, on pancreatic acinar cell trans-differentiation. J. Cell. Physiol. 2012, 227, 2470–2479. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, C.; Breslin, M.B.; Song, K.J.; Lan, M.S. Extra-nuclear activity of INSM1 transcription factor enhances insulin receptor signaling pathway and Nkx6.1 expression through RACK1 interaction. Cell Signal. 2014, 26, 740–747. [Google Scholar] [CrossRef]
- Angelo, L.; Elena, C.; Giorgia, M.; Diana, Z.; Paola, L.; Paola, C. KRAB-Zinc Finger Proteins: A Repressor Family Displaying Multiple Biological Functions. Curr. Genom. 2013, 14, 268–278. [Google Scholar]
- Frietze, S.; Lan, X.; Jin, V.X.; Farnham, P.J. Genomic targets of the KRAB and SCAN domain-containing zinc finger protein 263. J. Biol. Chem. 2010, 285, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.) 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Meissburger, B.; Perdikari, A.; Moest, H.; Muller, S.; Geiger, M.; Wolfrum, C. Regulation of adipogenesis by paracrine factors from adipose stromal-vascular fraction-a link to fat depot-specific differences. Biochim. Biophys. Acta 2016, 1861, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Zhang, M.; Zheng, Y.; Yan, P. ZBTB16 Overexpression Enhances White Adipogenesis and Induces Brown-Like Adipocyte Formation of Bovine White Intramuscular Preadipocytes. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 48, 2528–2538. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Su, X.; Wang, H.; Yang, W.; Zan, L. Cooperative and Independent Functions of the miR-23a~27a~24-2 Cluster in Bovine Adipocyte Adipogenesis. Int. J. Mol. Sci. 2018, 19, 3957. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.D.; Zan, L.S.; Li, A.N.; Cheng, G.; Li, S.J.; Zhang, Y.R.; Wang, X.Y.; Zhang, Y.Y. Characterization of the promoter region of the bovine long-chain acyl-CoA synthetase 1 gene: Roles of E2F1, Sp1, KLF15, and E2F4. Sci. Rep. 2016, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.W.; Gui, L.S.; Raza, S.H.A.; Zhang, S.; Khan, R.; Wang, L.; Guo, H.F.; Zan, L.S. NRF1 and ZSCAN10 bind to the promoter region of the SIX1 gene and their effects body measurements in Qinchuan cattle. Sci. Rep. 2017, 7, 7867. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wu, L.; Wang, X.; Xin, Y.; Zan, L. Tissue expression analysis, cloning and characterization of the 5′-regulatory region of the bovine FABP3 gene. Mol. Biol. Rep. 2016, 43, 991–998. [Google Scholar] [CrossRef]
- Wang, X.; Khan, R.; Raza, S.H.A.; Li, A.; Zhang, Y.; Liang, C.; Yang, W.; Wu, S.; Zan, L. Molecular characterization of ABHD5 gene promoter in intramuscular preadipocytes of Qinchuan cattle: Roles of Evi1 and C/EBPalpha. Gene 2018, 690, 38–47. [Google Scholar] [CrossRef]
- Junjvlieke, Z.; Mei, C.G.; Khan, R.; Zhang, W.Z.; Hong, J.Y.; Wang, L.; Li, S.J.; Zan, L.S. Transcriptional regulation of bovine elongation of very long chain fatty acids protein 6 in lipid metabolism and adipocyte proliferation. J. Cell. Biochem. 2019, 120, 13932–13943. [Google Scholar] [CrossRef] [PubMed]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, R.; Raza, S.H.A.; Junjvlieke, Z.; Xiaoyu, W.; Garcia, M.; Elnour, I.E.; Hongbao, W.; Linsen, Z. Function and Transcriptional Regulation of Bovine TORC2 Gene in Adipocytes: Roles of C/EBPγ, XBP1, INSM1 and ZNF263. Int. J. Mol. Sci. 2019, 20, 4338. https://doi.org/10.3390/ijms20184338
Khan R, Raza SHA, Junjvlieke Z, Xiaoyu W, Garcia M, Elnour IE, Hongbao W, Linsen Z. Function and Transcriptional Regulation of Bovine TORC2 Gene in Adipocytes: Roles of C/EBPγ, XBP1, INSM1 and ZNF263. International Journal of Molecular Sciences. 2019; 20(18):4338. https://doi.org/10.3390/ijms20184338
Chicago/Turabian StyleKhan, Rajwali, Sayed Haidar Abbas Raza, Zainaguli Junjvlieke, Wang Xiaoyu, Matthew Garcia, Ibrahim Elsaeid Elnour, Wang Hongbao, and Zan Linsen. 2019. "Function and Transcriptional Regulation of Bovine TORC2 Gene in Adipocytes: Roles of C/EBPγ, XBP1, INSM1 and ZNF263" International Journal of Molecular Sciences 20, no. 18: 4338. https://doi.org/10.3390/ijms20184338
APA StyleKhan, R., Raza, S. H. A., Junjvlieke, Z., Xiaoyu, W., Garcia, M., Elnour, I. E., Hongbao, W., & Linsen, Z. (2019). Function and Transcriptional Regulation of Bovine TORC2 Gene in Adipocytes: Roles of C/EBPγ, XBP1, INSM1 and ZNF263. International Journal of Molecular Sciences, 20(18), 4338. https://doi.org/10.3390/ijms20184338