Gap Junction Channels of Innexins and Connexins: Relations and Computational Perspectives

 ,
,  , , and
, , and 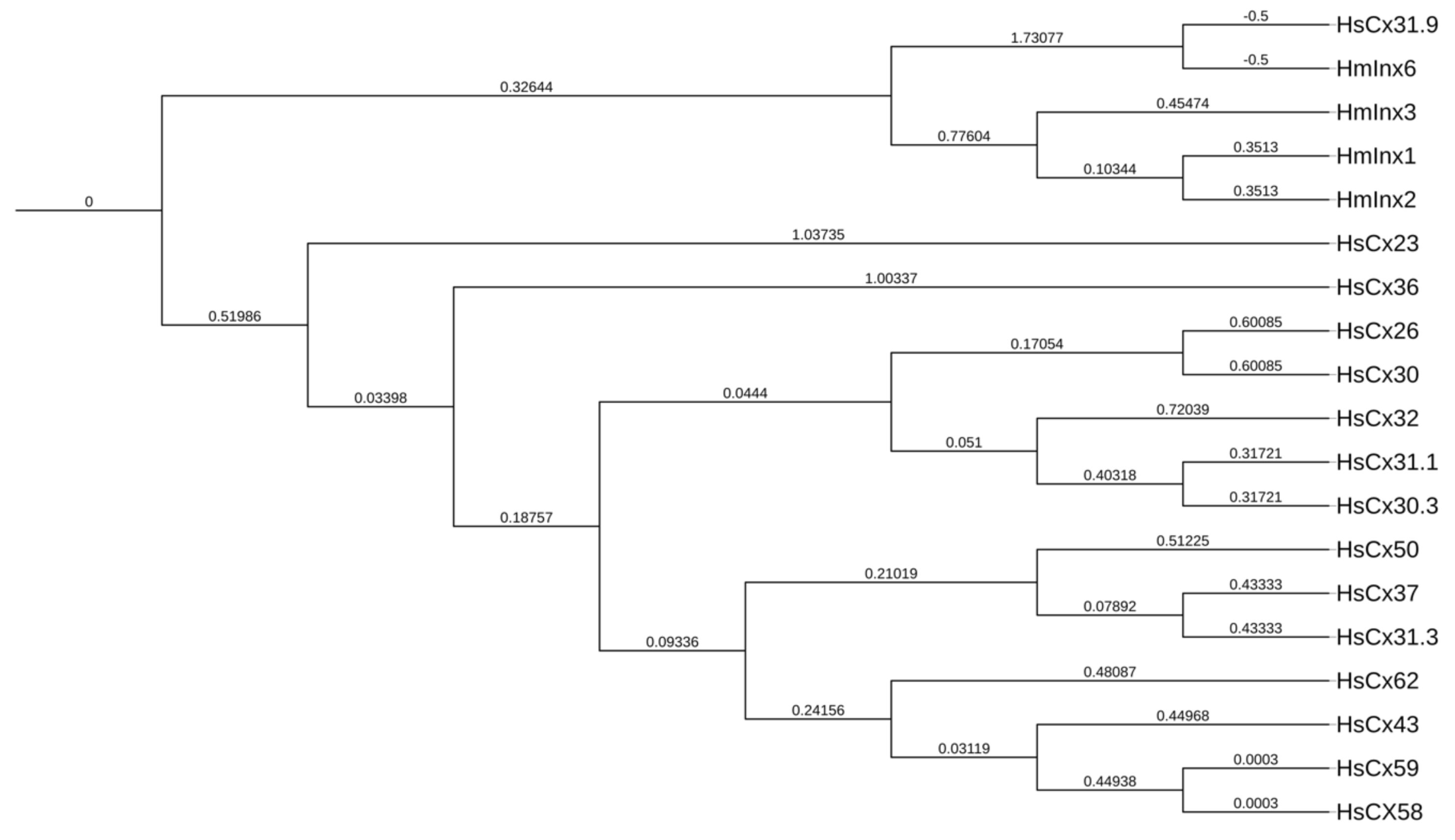{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. The Leech Nervous System: A Chain of Possibilities
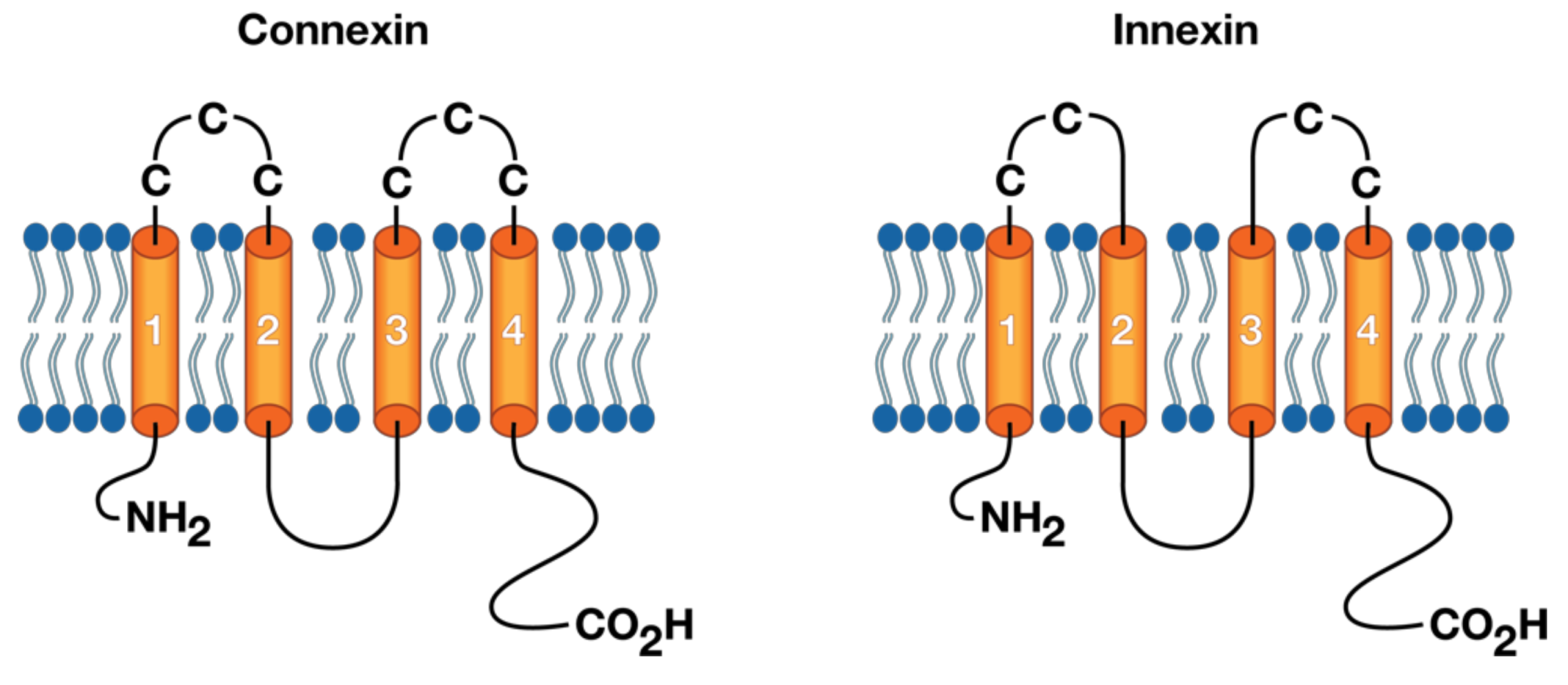
3. The Molecular Structure of Connexins/Innexins
4. The Molecular Structure of Gap Junction Proteins
5. Inhibition of Gap Junction Communication
5.1. Chemical Mechanisms
5.2. RNA Interference
5.3. Anti-Peptide Antibodies
5.4. Antisense Oligonucleotides
5.5. Mimetic Peptides
6. Computational Models
7. Discussion and Future Work
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GJs | Gap junctions |
| CJC | Gap junction communication |
| RNAi | RNA interference |
| dsRNA | Double-stranded RNA |
| ApepA | Anti-peptide antibodies |
| ASOs | Antisense oligonucleotides |
| MPs | Mimetic peptides |
| SVM | Support vector machine |
References
- Scemes, E.; Spray, D.C.; Meda, P. Connexins, pannexins, innexins: Novel roles of “hemi-channels”. Pflug. Arch. Eur. J. Physiol. 2009, 457, 1207–1226. [Google Scholar] [CrossRef]
- Panchin, Y.V. Evolution of gap junction proteins—The pannexin alternative. J. Exp. Biol. 2005, 1415–1419. [Google Scholar] [CrossRef] [PubMed]
- Dykes, I.M. Molecular Basis of Gap Junctional Communication in the CNS of the Leech Hirudo medicinalis. J. Neurosci. 2004, 24, 886–894. [Google Scholar] [CrossRef]
- Oshima, A.; Matsuzawa, T.; Nishikawa, K.; Fujiyoshi, Y. Oligomeric structure and functional characterization of caenorhabditis elegans innexin-6 gap junction protein. J. Biol. Chem. 2013, 288, 10513–10521. [Google Scholar] [CrossRef]
- Nielsen, M.S.; Axelsen, L.N.; Sorgen, P.L.; Verma, V.; Delmar, M.; Holstein-Rathlou, N.H. Gap junctions. Compr. Physiol. 2012, 2, 1981–2035. [Google Scholar] [CrossRef]
- Lohman, A.W.; Isakson, B.E. Differentiating connexin hemichannels and pannexin channels in cellular ATP release. FEBS Lett. 2014, 588, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Baranova, A.; Ivanov, D.; Petrash, N.; Pestova, A.; Skoblov, M.; Kelmanson, I.; Shagin, D.; Nazarenko, S.; Geraymovych, E.; Litvin, O.; et al. The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics 2004, 83, 706–716. [Google Scholar] [CrossRef]
- Yen, M.R.; Saier, M.H. Gap junctional proteins of animals: The innexin/pannexin superfamily. Prog. Biophys. Mol. Biol. 2007, 94, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Fushiki, D.; Hamada, Y.; Yoshimura, R.; Endo, Y. Phylogenetic and bioinformatic analysis of gap junction-related proteins, innexins, pannexins and connexins. Biomed. Res. 2010, 31, 133–142. [Google Scholar] [CrossRef]
- Locovei, S.; Wang, J.; Dahl, G. Activation of pannexin 1 channels by ATP through P2Y receptors and by cytoplasmic calcium. FEBS Lett. 2006, 580, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, C.; Lechner, H.; Lo, B.; Knieps, M.; Herrmann, S.; Famulok, M.; Bauer, R.; Hoch, M. eteromerization of Innexin Gap Junction Proteins Regulates Epithelial Tissue Organization in Drosophila. Mol. Biol. Cell 2006, 17, 1676–1685. [Google Scholar] [CrossRef]
- Baker, M.W.; Macagno, E.R. Gap Juntion Proteins and the Wiring (Rewiring) of Neural Circuits. Dev. Neurobiol. 2016, 1–23. [Google Scholar] [CrossRef]
- Yazdani, N.; Firme, C.P.; Macagno, E.R.; Baker, M.W. Expression of a dominant negative mutant innexin in identified neurons and glial cells reveals selective interactions among gap junctional proteins. Dev. Neurobiol. 2013, 73, 571–586. [Google Scholar] [CrossRef]
- Nielsen, P.A.; Beahm, D.L.; Giepmans, B.N.; Baruch, A.; Hall, J.E.; Kumar, N.M. Molecular cloning, functional expression, and tissue distribution of a novel human gap junction-forming protein, connexin-31.9. Interaction with zona occludens protein-1. J. Biol. Chem. 2002, 277, 38272–38283. [Google Scholar] [CrossRef]
- White, T.; Srinivas, M.; Ripps, H.; Trovato-Salinaro, A.; Condorelli, D.; Bruzzone, R. Virtual cloning, functional expression, and gating analysis of human connexin31.9. Am. J. Physiol. Cell Physiol. 2002, 283, C960–C970. [Google Scholar] [CrossRef]
- Kreuzberg, M.; Söhl, G.; Kim, J.S.; Verselis, V.; Willecke, K.; Bukauskas, F. Functional properties of mouse connexin30.2 expressed in the conduction system of the heart. Circ. Res. 2005, 96, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Bukauskas, F.; Kreuzberg, M.; Rackauskas, M.; Bukauskiene, A.; Bennett, M.; Verselis, V.; Willecke, K. Properties of mouse connexin 30.2 and human connexin 31.9 hemichannels: Implications for atrioventricular conduction in the heart. Proc. Natl. Acad. Sci. USA 2006, 103, 9726–9733. [Google Scholar] [CrossRef] [PubMed]
- Sohl, G.; Willecke, K. Gap junctions and the connexin protein family. Cardiovasc. Res. 2004, 62, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Eastman, S.D.; Chen, T.H.; Falk, M.M.; Mendelson, T.C.; Iovine, M.K. Phylogenetic analysis of three complete gap junction gene families reveals lineage-specific duplications and highly supported gene classes. Genomics 2006, 87, 265–274. [Google Scholar] [CrossRef]
- Cruciani, V.; Mikalsen, S.O. Evolutionary selection pressure and family relationships among connexin genes. Biol. Chem. 2007, 388, 253–264. [Google Scholar] [CrossRef]
- Tao, W.; Zhang, S.; Turenchalk, G.S.; Stewart, R.A.; John, M.A.R.S.; Chen, W.; Xu, T. Human homologue of the Drosophila melanogaster lats tumour suppressor modulates CDC2 activity. Nature 1999, 21, 177–181. [Google Scholar] [CrossRef]
- Stebbings, L.A.; Todman, M.G.; Phelan, P.; Bacon, J.P.; Davies, J.A. Two Drosophila Innexins Are Expressed in Overlapping Domains and Cooperate to Form Gap-Junction Channels. Mol. Biol. Cell 2000, 11, 2459–2470. [Google Scholar] [CrossRef] [PubMed]
- Starich, T.; Sheehan, M.; Jadrich, J.; Shaw, J. Innexins in C. elegans. Cell Commun. Adhes. 2001, 8, 311–314. [Google Scholar] [CrossRef]
- Leake, L.D. The leech as a scientific tool. Endeavour 1983, 7, 88–93. [Google Scholar] [CrossRef]
- Wagenaar, D.A. A classic model animal in the 21st century: Recent lessons from the leech nervous system. J. Exp. Biol. 2015, 218, 3353–3359. [Google Scholar] [CrossRef]
- Sargent, P.B.; Nicholls, G. Extrasynaptic Receptors on Cell Bodies of Neurons in Central Nervous System of the Leech. J. Neurophysiol. 1977, 40. [Google Scholar] [CrossRef]
- Leake, L.D.; Crowe, R.; Burnstock, G. Localisation of substance P-, somatostatin-, vasoactive intestinal polypeptide- and met-enkephalin-immunoreactive nerves in the peripheral and central nervous systems of the leech (Hirudo medicinalis). Cell Tisue Res. 1986, 345–351. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Sihra, T.S.; Sanchez-Prieto, J. Calcium-Dependent and-Independent Release of Glutamate from Synaptosomes Monitored by Continuous Fluorometry. J. Neurochem. 1987, 49, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Kristan, W.B.; Stent, G.S.; Ort, C.A. Neuronal Control of Swimming in the Medicinal Leech. J. Comp. Physiol. A 1974, 119, 97–119. [Google Scholar] [CrossRef]
- Lent, C.M.; Dickinson, M.H. Serotonin integrates the feeding behavior of the medicinal leech. J. Comp. Physiol. A 1984, 457–471. [Google Scholar] [CrossRef]
- Misell, L.M.; Shaw, B.K.; Kristan, W.B., Jr. Behavioral hierarchy in the medicinal leech, Hirudo medicinalis: Feeding as a dominant behavior. Behav. Brain Res. 1998, 90, 13–21. [Google Scholar] [CrossRef]
- Dykes, I.M.; Macagno, E.R. Molecular characterization and embryonic expression of innexins in the leech Hirudo medicinalis. Dev. Genes Evol. 2006, 216, 185–197. [Google Scholar] [CrossRef]
- Kandarian, B.; Sethi, J.; Wu, A.; Baker, M.; Yazdani, N.; Kym, E.; Sanchez, A.; Edsall, L.; Gaasterland, T.; Macagno, E. The medicinal leech genome encodes 21 innexin genes: Different combinations are expressed by identified central neurons. Dev. Genes Evol. 2012, 222, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Firme, C.P.; Natan, R.G.; Yazdani, N.; Macagno, E.R.; Baker, M.W. Ectopic Expression of Select Innexins in Individual Central Neurons Couples Them to Pre-Existing Neuronal or Glial Networks That Express the Same Innexin. J. Neurosci. 2012, 32, 14265–14270. [Google Scholar] [CrossRef]
- Szabo, T.M. Transient Electrical Coupling Delays the Onset of Chemical Neurotransmission at Developing Synapses. J. Neurosci. 2004, 24, 112–120. [Google Scholar] [CrossRef]
- Marin-Burgin, A.; Eisenhart, F.J.; Kristan, W.B., Jr.; French, K.A. Embryonic electrical connections appear o prefigure a behavioral circuit in the leech CNS. J. Comp. Physiol. A 2006, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Kristan, W.B.; Wittenberg, G.; Nusbaum, M.P.; Stern-Tomlinson, W. Multifunctional interneurons in behavioral circuits of the medicinal leech. Experientia 1988, 44, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Beyer, E.C.; Paul, D.L.; Goodenough, D.A. Connexin Family of Gap Junction Proteins. J. Membr. Biol. 1990, 194, 187–194. [Google Scholar] [CrossRef]
- Harris, A.L. Voltage-sensing and Substate Rectification: Moving Parts of Connexin Channels. J. Gen. Physiol. 2002, 119. [Google Scholar] [CrossRef]
- Iglesias, R.; Spray, D.C.; Scemes, E. Mefloquine Blockade of Pannexin1 Currents: Resolution of a Conflict. Cell Commun. Adhes. 2010, 16, 131–137. [Google Scholar] [CrossRef]
- Kumar, N.M.; Gilula, N.B. The Gap Juntion Communication Channel. Cell 1996, 84, 381–388. [Google Scholar] [CrossRef]
- Foote, C.I.; Zhou, L.; Zhu, X.; Nicholson, B.J. The Pattern of Disulfide Linkages in the Extracellular Loop Regions of Connexin 32 Suggests a Model for the Docking Interface of Gap Junctions. J. Cell Biol. 1998, 140, 1187–1198. [Google Scholar] [CrossRef] [PubMed]
- Sosinsky, G.E.; Nicholson, B.J. Structural organization of gap junction channels. Biochim. Biophys. Acta 2005, 1711, 99–125. [Google Scholar] [CrossRef]
- Oshima, A.; Tani, K.; Fujiyoshi, Y. Atomic structure of the innexin-6 gap junction channel determined by cryo-EM. Nat. Commun. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Unwin, P.N.T.; Zamphigi, G. Structure of the juntion between communicating cells. Nature 1980, 283, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.J.; Hand, G.M.; Engels, A.; Sosinsky, G.E. Conformational changes in surface structures of isolated connexin 26 gap junctions. EMBO J. 2002, 21, 3598–3607. [Google Scholar] [CrossRef]
- Bennett, B.C.; Purdy, M.D.; Baker, K.A.; Acharya, C.; Mcintire, W.E.; Stevens, R.C.; Zhang, Q.; Harris, A.L.; Abagyan, R.; Yeager, M. An electrostatic mechanism for Ca2+-mediated regulation of gap junction channels. Nat. Commun. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Rivkin, S.; Tang, Q.; Verselis, V.K.; Bargiello, T.A. Determinants of Gating Polarity of a Connexin 32 Hemichannel. Biophys. J. 2004, 87, 912–928. [Google Scholar] [CrossRef] [PubMed]
- Bargiello, T.A.; Tang, Q.; Oh, S.; Kwon, T. Voltage-dependent conformational changes in connexin channels. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1807–1822. [Google Scholar] [CrossRef]
- Skerrett, I.M.; Williams, J.B. A structural and functional comparison of gap junction channels composed of onnexins and innexins. Dev. Neurobiol. 2017, 77, 522–547. [Google Scholar] [CrossRef]
- Manjarrez-Marmolejo, J.; Franco-Pérez, J. Gap Junction Blockers: An Overview of their Effects on Induced Seizures in Animal Models. Curr. Neuropharmacol. 2016, 759–771. [Google Scholar] [CrossRef]
- Todd, K.L.; Kristan, W.B.; French, K.A. Gap Junction Expression Is Required for Normal Chemical Synapse Formation. J. Neurosci. 2010, 30, 15277–15285. [Google Scholar] [CrossRef]
- Dahl, G. Gap Junction—Mimetic Peptides do Work, but in Unexpected Ways. Cell Commun. Adhes. 2009, 9061. [Google Scholar] [CrossRef]
- Nogi, T.; Levin, M. Characterization of innexin gene expression and functional roles of gap-junctional communication in planarian regeneration. Dev. Biol. 2005, 287, 314–335. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.; Simon, S.; Ramón, F. Interaction of anesthetics with electrical synapses. Nature 1980, 286, 498–500. [Google Scholar] [CrossRef]
- Allaire, K.M.; Watson, G.M. Rho participates in chemoreceptor-induced changes in morphology to hair bundle mechanoreceptors of the sea anemone, Nematostella vectensis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 165, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Tse, G.; Yeo, J.M.; Tse, V.; Kwan, J.; Sun, B. Gap junction inhibition by heptanol increases ventricular rhythmogenicity by reducing conduction velocity without affecting repolarization properties or myocardial refractoriness in Langendorff-perfused mouse hearts. Mol. Med. Rep. 2016, 14, 4069–4074. [Google Scholar] [CrossRef] [PubMed]
- Samuels, S.E.; Lipitz, J.B.; Wang, J.; Dahl, G.; Muller, K.J. Arachidonic acid closes innexin/pannexin hannels and thereby inhibits microglia cell movement to a nerve injury. Dev. Neurobiol. 2013, 73, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Mire, P.; Nasse, J.; Venable-thibodeaux, S. Gap junctional communication in the vibration-sensitive response of sea anemones. Hear. Res. 2000, 144, 109–123. [Google Scholar] [CrossRef]
- Adler, E.L.; Woodruff, R.I.; Chester, W. Varied Effects of 1-Octanol on Gap Junctional Communication Between Ovarian Epithelial Cells and Oocytes of Oncopeltus fasciatus, Hyalophora cecropia, and Drosophila melanogaster. Arch. Insect Biochem. Physiol. 2000, 43, 22–32. [Google Scholar] [CrossRef]
- Anderson, K.L.; Woodruff, R.I. A gap junctionally transmitted epithelial cell signal regulates endocytic yolk uptake in Oncopeltus fasciatus. Dev. Biol. 2001, 239, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.A.; Woodruff, R.I. Calmodulin transmitted through gap junctions stimulates endocytic incorporation of yolk precursors in insect oocytes. Dev. Biol. 2004, 271, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, R.; Suetsugu, T.; Endo, Y. Serotonergic transmission and gap junctional coupling in proventricular muscle cells in the American cockroach, Periplaneta americana. J. Insect Physiol. 2017, 99, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Talaverón, R.; Fernández, P.; Escamilla, R.; Pastor, A.M.; Hayley, S. Neural progenitor cells isolated from the subventricular zone present hemichannel activity and form functional gap junctions with glial cells. Front. Cell. Neurosci. 2015, 9, 1–10. [Google Scholar] [CrossRef]
- Kagiava, A.; Theophilidis, G.; Sargiannidou, I.; Kyriacou, K.; Kleopa, K.A. Oxaliplatin-induced neurotoxicity is mediated through gap junction channels and hemichannels and can be prevented by octanol. Neuropharmacology 2015, 97, 289–305. [Google Scholar] [CrossRef]
- Takano, K.; Ogawa, M.; Kawabe, K.; Moriyama, M. Inhibition of Gap Junction Elevates Glutamate Uptake in Cultured Astrocytes. Neurochem. Res. 2017. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Lett. Nat. 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Wang, J.; Ma, M.; Locovei, S.; Keane, R.W.; Dahl, G. Modulation of membrane channel currents by gap junction protein mimetic peptides: Size matters. Am. J. Physiol. Cell Physiol. 2007, 293, C1112–C1119. [Google Scholar] [CrossRef]
- Talbot, J.; Brion, R.; Lamora, A.; Mullard, M.; Morice, S.; Heymann, D.; Verrecchia, F. Connexin43 intercellular communication drives the early differentiation of human bone marrow stromal cells into osteoblasts. J. Cell. Physiol. 2018, 233, 946–957. [Google Scholar] [CrossRef]
- O’Donnell, J.J., III; Birukova, A.A.; Beyer, E.C.; Birukov, K.G. Gap junction protein connexin43 exacerbates lung vascular permeability. PLoS ONE 2014, 9, e100931. [Google Scholar] [CrossRef]
- Osbourne, A.; Calway, T.; Broman, M.; McSharry, S.; Earley, J.; Kim, G.H. Downregulation of connexin43 by microRNA-130a in cardiomyocytes results in cardiac arrhythmias. J. Mol. Cell. Cardiol. 2014, 74, 53–63. [Google Scholar] [CrossRef]
- Gärtner, C.; Ziegelhöffer, B.; Kostelka, M.; Stepan, H.; Mohr, F.W.; Dhein, S. Knock-down of endothelial connexins impairs angiogenesis. Pharmacol. Res. 2012, 65, 347–357. [Google Scholar] [CrossRef]
- Anava, S.; Saad, Y.; Ayali, A. The role of gap junction proteins in the development of neural network functional topology. Insect Mol. Biol. 2013, 22, 457–472. [Google Scholar] [CrossRef]
- Li, M.W.; Wang, J.; Zhao, Y.O.; Fikrig, E. Innexin AGAP001476 is critical for mediating anti-Plasmodium responses in Anopheles mosquitoes. J. Biol. Chem. 2014, 289, 24885–24897. [Google Scholar] [CrossRef]
- Calkins, T.L.; Piermarini, P.M. Pharmacological and Genetic Evidence for Gap Junctions as Potential New Insecticide Targets in the Yellow Fever Mosquito, Aedes aegypti. PLoS ONE 2015, 1–15. [Google Scholar] [CrossRef]
- Calkins, T.L.; Piermarini, P.M. A Blood Meal Enhances Innexin mRNA Expression in the Midgut, Malpighian Tubules, and Ovaries of the Yellow Fever Mosquito Aedes aegypti. Insects 2017, 8, 122. [Google Scholar] [CrossRef]
- Schulz, R.; Görge, P.M.; Görbe, A.; Ferdinandy, P.; Lampe, P.D.; Leybaert, L. Connexin 43 is an emerging therapeutic target in ischemia/reperfusion injury, cardioprotection and neuroprotection. Pharmacol. Ther. 2015, 153, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, M.A.; Kar, R.; Gu, S.; Jiang, J.X. Neuropharmacology Antibodies targeting extracelular domain of connexins for studies of hemichannels. Neuropharmacology 2013, 75, 525–532. [Google Scholar] [CrossRef]
- Bastide, B.; Briand, J.P.; Gros, D. Effect of Antipeptide Antibodies Directed against Three Domains of Connexin43 on the Gap Junctional Permeability of Cultured Heart Cells. J. Membr. Biol. 1996, 253, 243–253. [Google Scholar] [CrossRef]
- Hertzberg, E.L.; Sa, J.C.; Corpina, R.A.; Roy, C.; Kessler, J.A. Use of Antibodies in the Analysis of Connexin 43 Turnover and Phosphorylation. Acad. Press 2000, 20, 129–139. [Google Scholar] [CrossRef]
- Sosinsky, G.E.; Solan, J.L.; Gaietta, G.M.; Ngan, L.; Lee, G.J.; Mackey, M.R.; Lampe, P.D. The C-terminus of connexin43 adopts different conformations in the Golgi and gap junction as detected with structure-specific antobodies. Biochem. J. 2007, 408, 375–385. [Google Scholar] [CrossRef]
- Clair, C.; Combettes, L.; Pierre, F.; Sansonetti, P.; Tran, G.; Nhieu, V. Extracellular-loop peptide antibodies reveal a predominant hemichannel organization of connexins in polarized intestinal cells. Exp. Cell Res. 2008, 4. [Google Scholar] [CrossRef]
- Bohrmann, J.; Zimmermann, J. Gap junctions in the ovary of Drosophila melanogaster: Localization of innexins 1, 2, 3 and 4 and evidence for intercellular communication via innexin-2 containing channels. BMC Dev. Biol. 2008, 8, 1–12. [Google Scholar] [CrossRef]
- Güiza, J.; Barría, I.; Sáez, J.C.; Vega, J.L. Innexins: Expression, Regulation, and Functions. Front. Physiol. 2018, 9, 1414. [Google Scholar] [CrossRef] [PubMed]
- Evers, M.M.; Toonen, L.J.A.; Roon-mom, W.M.C.V. Antisense oligonucleotides in therapy for neurodegenerative disorders. Adv. Drug Deliv. Rev. 2015, 87, 90–103. [Google Scholar] [CrossRef]
- Schoch, K.M.; Miller, T.M. Review Antisense Oligonucleotides: Translation from Mouse Models to Human Neurodegenerative Diseases. Neuron 2017, 94, 1056–1070. [Google Scholar] [CrossRef]
- Rinaldi, C.; Wood, M.J.A. Antisense oligonucleotides: The next disorders. Nat. Rev. Neurol. 2017. [Google Scholar] [CrossRef]
- Green, C.R.; Law, L.; Lin, J.S.; Becker, D.L. Spatiotemporal Depletion of connexins Using Antisense Oligonucleotides. Methods Mol. Biol. 2001, 154, 175–185. [Google Scholar] [CrossRef]
- Danesh-Meyer, H.V.; Huang, R.; Nicholson, L.F.B.; Green, C.R. Connexin43 antisense oligodeoxynucleotide treatment down-regulates the inflammatory response in an in vitro interphase organotypic culture model of optic nerve ischaemia. J. Clin. Neurosci. 2008, 15, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Grupcheva, C.N.; Laux, W.T.; Rupenthal, I.D.; Mcghee, J.; Mcghee, C.N.J.; Green, C.R. Improved Corneal Wound Healing through Modulation Connexin43-Specific Antisense Oligodeoxynucleotides. Investig. Ophthalmol. Vis. Sci. 2012, 1130–1138. [Google Scholar] [CrossRef]
- Dahl, G.; Werner, R.; Levine, E.; Rabadan-diehl, C. Mutational analysis of gap junction formation. Biophys. J. 1992, 62, 172–182. [Google Scholar] [CrossRef]
- Dahl, G.; Nonner, W.; Werner, R. Attempts to define functional domains of gap junction proteins with synthetic peptides. Biophys. J. 1994, 67, 1816–1822. [Google Scholar] [CrossRef]
- Kwak, B.R.; Jongsma, H.J. Selective inhibition of gap junction channel activity by synthetic peptides. J. Physiol. 1999, 516, 679–685. [Google Scholar] [CrossRef]
- Martin, P.E.M.; Wall, C.; Griffith, T.M. Effects of connexin-mimetic peptides on gap junction functionality and connexin expression in cultured vascular cells. Br. J. Pharmacol. 2005, 144, 617–627. [Google Scholar] [CrossRef]
- Wang, Z. Big data mining powers fungal research: Recent advances in fission yeast systems biology approaches. Curr. Genet. 2016. [Google Scholar] [CrossRef]
- De Castro, L. Fundamentals of natural computing: An overview. Phys. Life Rev. 2006, 4, 1–36. [Google Scholar] [CrossRef]
- Mondal, A.; Moreno, A.P. Heteromultimeric gap-junction channel permeance: Directional fluxes simulated using a Brownian dynamics model. Biophys. J. 2010, 98, 94a–95a. [Google Scholar] [CrossRef][Green Version]
- Kwon, T.; Harris, A.L.; Rossi, A.; Bargiello, T.A. Molecular dynamics simulations of the Cx26 hemichannel: Evaluation of structural models with Brownian dynamics. J. Gen. Physiol. 2011, 138, 475–493. [Google Scholar] [CrossRef]
- Mondal, A.; Appadurai, D.A.; Nazem, W.; Sachse, F.B.; Moreno, A.P. Computational Simulations of Asymmetric Fluxes of Large Molecules Through Gap Junction Channel Pores. J. Theor. Biol. 2016. [Google Scholar] [CrossRef]
- Bargiello, T.A.; Oh, S.; Tang, Q.; Bargiello, N.K.; Dowd, T.L.; Kwon, T. Gating of Connexin Channels by transjunctional-voltage: Conformations and models of open and closed states. Biochim. Biophys. Acta Biomembr. 2018, 1860, 22–39. [Google Scholar] [CrossRef]
- Janjua, K.; Garyantes, T.; Baron, B. Identification of Gap Junction Blockers Using Automated Fluorescence Microscopy Imaging. J. Biomol. Screen. 2003, 8, 489–499. [Google Scholar] [CrossRef]
- Casaleggio, A.; Hines, M.L.; Migliore, M. Computational Model of Erratic Arrhythmias in a Cardiac Cell Network: The Role of Gap Junctions. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Kononenko, I. Machine learning for medical diagnosis: History, state of the art and perspective. Artif. Intell. Med. 2001, 23, 89–109. [Google Scholar] [CrossRef]
- Fooshee, D.; Mood, A.; Gutman, E.; Tavakoli, M.; Urban, G.; Liu, F.; Huynh, N.; van Vranken, D.; Baldi, P. Deep learning for chemical reaction prediction. Mol. Syst. Des. Eng. 2018. [Google Scholar] [CrossRef]
- Wang, J.; Ding, H.; Bidgoli, F.A.; Zhou, B.; Iribarren, C.; Molloi, S.; Baldi, P. Detecting Cardiovascular Disease from Mammograms with Deep Learning. IEEE Trans. Med Imaging 2017, 36, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Baldi, P. Deep Learning in Biomedical Data Science. Annu. Rev. 2018, 181–205. [Google Scholar] [CrossRef]
- Cai, Y.D.; Zhou, G.P.; Chou, K.C. Support Vector Machines for Predicting Membrane Protein Types by Using Functional Domain Composition. Biophys. J. 2003, 84, 3257–3263. [Google Scholar] [CrossRef]
- Guermeur, Y.; Pollastri, G.; Zelus, D. Combining protein secondary structure prediction models with ensemble methods of optimal complexity. Neurocomputing 2004, 56, 305–327. [Google Scholar] [CrossRef]
- Lee, E.Y.; Fulan, B.M.; Wong, G.C.L.; Ferguson, A.L. Mapping membrane activity in undiscovered peptide sequence space using machine learning. Proc. Natl. Acad. Sci. USA 2016, 113. [Google Scholar] [CrossRef]
- Karamzadeh, R.; Karimi-jafari, M.H.; Zarchi, A.S. Machine Learning and Network Analysis of Molecular Dynamics Trajectories Reveal Two Chains of Red/Ox-specific Residue Interactions in Human Protein Disulfide Isomerase. Nat. Sci. Rep. 2017, 1–11. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Langlois, R.E.; Zhao, G.; Lu, H. Kernel-based machine learning protocol for predicting DNA-binding proteins. Nucleic Acids Res. 2005, 33, 6486–6493. [Google Scholar] [CrossRef] [PubMed]
- Ballester, P.J.; Mitchell, J.B.O. A machine learning approach to predicting protein—ligand binding affinity with applications to molecular docking. Bioinformatics 2010, 26, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- King, R.D.; Sternbergt, M.J.E. Machine Learning Approach for the Prediction of Protein Secondary Structure. J. Mol. Biol. 1990, 441, 441–457. [Google Scholar] [CrossRef]
- Nielsen, H.; Brunak, S.; Heijne, G.V. Machine learning approaches for the prediction of signal peptides and other protein sorting signals. Protein Eng. 1999, 12, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Gourdie, R.G.; Green, C.R.; Severs, N.J. Gap junction distribution in adult mammalian myocardium revealed by an anti-peptide antibody and laser scanning confocal microscopy. J. Cell. Sci. 1991, 99 Pt 1, 41–55. [Google Scholar]
- Rawanduzy, A.; Hansen, A.; Hansen, T.W.; Nedergaard, M. Effective reduction of infarct volume by gap junction blockade in a rodent model of stroke. J. Neurosurg. 1997, 87, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Mizoguchi, H.; Doi, Y.; Jin, S.; Noda, M.; Liang, J. Blockade of Gap Junction Hemichannel Suppresses Disease Progression in Mouse Models of Amyotrophic Lateral Sclerosis and Alzheimer’s Disease. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.; King, R.D.; Altmann, T.; Fiehn, O. Application of metabolomics to plant genotype discrimination using statistics and machine learning. Bioinformatics 2002, 18, 241–248. [Google Scholar] [CrossRef]
- Korolev, D.; Balakin, K.V.; Nikolsky, Y.; Kirillov, E.; Ivanenkov, Y.A.; Savchuk, N.P.; Ivashchenko, A.A.; Nikolskaya, T. Modeling of Human Cytochrome P450-Mediated Drug Metabolism Using Unsupervised Machine Learning Approach. J. Med. Chem. 2003, 46, 3631–3643. [Google Scholar] [CrossRef] [PubMed]
- Ziliang, Y.D.C.; Lin, Q.; Xin, K.Y.F.; Bing, M.; Lu, G.D.Z.W.C. Prediction of compounds’ biological function (metabolic pathways) based on functional group composition. Mol. Divers. 2008, 131–137. [Google Scholar] [CrossRef]
- Plaimas, K.; Mallm, J.P.; Oswald, M.; Svara, F.; Sourjik, V.; Eils, R.; König, R. Machine learning based analysis on metabolic networks supports high-throughput knockout screens. BMC Syst. Biol. 2008, 11, 1–11. [Google Scholar] [CrossRef]
- Dale, J.M.; Popescu, L.; Karp, P.D. Machine learning methods for metabolic pathway prediction. BMC Bioinform. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Romero, M.; Vázquez-Naya, J.M.; Rabuñal, J.R.; Pita-Fernández, S.; Macenlle, R.; Castro-Alvariño, J.; López-Roses, L.; Ulla, J.L.; Martínez-Calvo, A.V.; Vázquez, S.; et al. Artificial Intelligence Techniques for Colorectal Cancer Drug Metabolism: Ontologies and Complex Networks. Curr. Drug Metab. 2010, 11, 347–368. [Google Scholar] [CrossRef]
- Lima, A.N.; Philot, E.A.; Goulart, G.H.; Paulo, L.; Scott, B.; Maltarollo, V.G. Expert Opinion on Drug Discovery Use of machine learning approaches for novel drug discovery. Expert Opin. Drug Discov. 2016, 11, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.D.; Liu, X.J.; Xu, X.B.; Chou, K.C. Support Vector Machines for Predicting HIV Protease Cleavage Sites in Protein. J. Comput. Chem. 2002, 23, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Li, Y.; Yang, S.L.; Yang, L. Classification of Substrates and Inhibitors of P-Glycoprotein Using Unsupervised Machine Learning Approach. J. Chem. Inf. Model. 2005, 45, 750–757. [Google Scholar] [CrossRef]
- Kinnings, S.L.; Liu, N.; Tonge, P.J.; Jackson, R.M.; Xie, L.; Bourne, P.E. A Machine Learning-Based Method to Improve Docking Scoring Functions and Its Application to Drug Repurposing. J. Chem. Inf. Model. 2011, 408–419. [Google Scholar] [CrossRef]
- Valdebenito, S.; Lou, E.; Baldoni, J.; Okafo, G. The Novel Roles of Connexin Channels and Tunneling Nanotubes in Cancer Pathogenesis. Int. J. Mol. Sci. 2018. [Google Scholar] [CrossRef]
- Vargas, A.A.; Cisterna, B.A.; Saavedra-Leiva, F.; Urrutia, C.; Cea, L.A.; Vielma, A.H.; Gutierrez-Maldonado, S.E.; Martin, A.J.M.; Pareja-Barrueto, C.; Escalona, Y.; et al. On Biophysical Properties and Sensitivity to Gap Junction Blockers of Connexin 39 Hemichannels Expressed in HeLa Cells. Front. Physiol. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Bauer, R.; Löer, B.; Ostrowski, K.; Martini, J.; Weimbs, A.; Lechner, H.; Hoch, M. Intercellular Communication: The Drosophila Innexin Multiprotein Family of Gap Junction Proteins. Chem. Biol. 2005, 12, 515–526. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, A.; Castro, C.; Flores, D.-L.; Gutiérrez, E.; Baldi, P. Gap Junction Channels of Innexins and Connexins: Relations and Computational Perspectives. Int. J. Mol. Sci. 2019, 20, 2476. https://doi.org/10.3390/ijms20102476
Sánchez A, Castro C, Flores D-L, Gutiérrez E, Baldi P. Gap Junction Channels of Innexins and Connexins: Relations and Computational Perspectives. International Journal of Molecular Sciences. 2019; 20(10):2476. https://doi.org/10.3390/ijms20102476
Chicago/Turabian StyleSánchez, Alejandro, Carlos Castro, Dora-Luz Flores, Everardo Gutiérrez, and Pierre Baldi. 2019. "Gap Junction Channels of Innexins and Connexins: Relations and Computational Perspectives" International Journal of Molecular Sciences 20, no. 10: 2476. https://doi.org/10.3390/ijms20102476
APA StyleSánchez, A., Castro, C., Flores, D.-L., Gutiérrez, E., & Baldi, P. (2019). Gap Junction Channels of Innexins and Connexins: Relations and Computational Perspectives. International Journal of Molecular Sciences, 20(10), 2476. https://doi.org/10.3390/ijms20102476