Technological and Genomic Analysis of Roles of the Cell-Envelope Protease PrtS in Yoghurt Starter Development

 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Technological Properties of S. thermophilus and Lb. bulgaricus
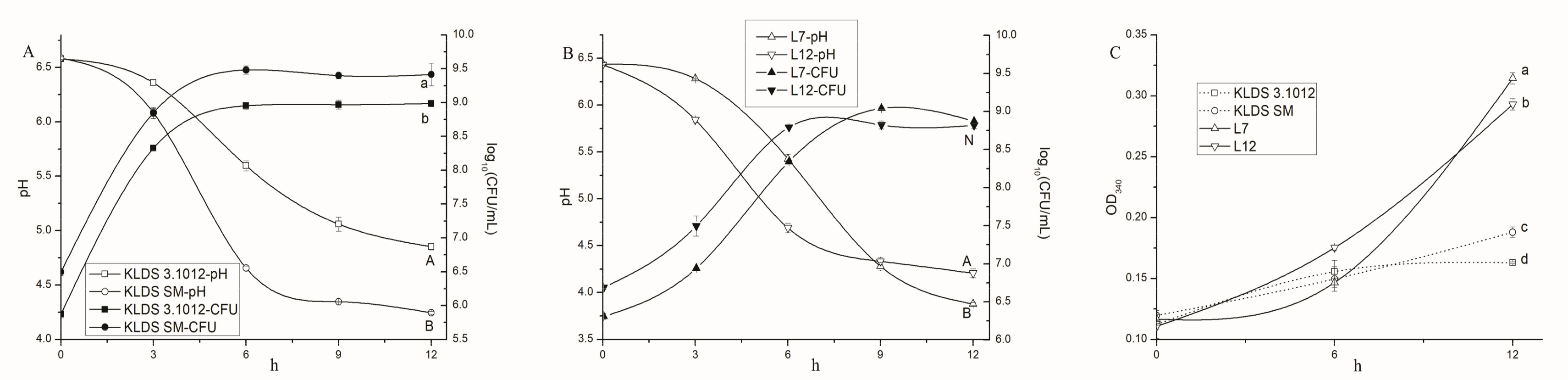
2.1.1. Acid Production and Growth
2.1.2. Proteolytic Activity
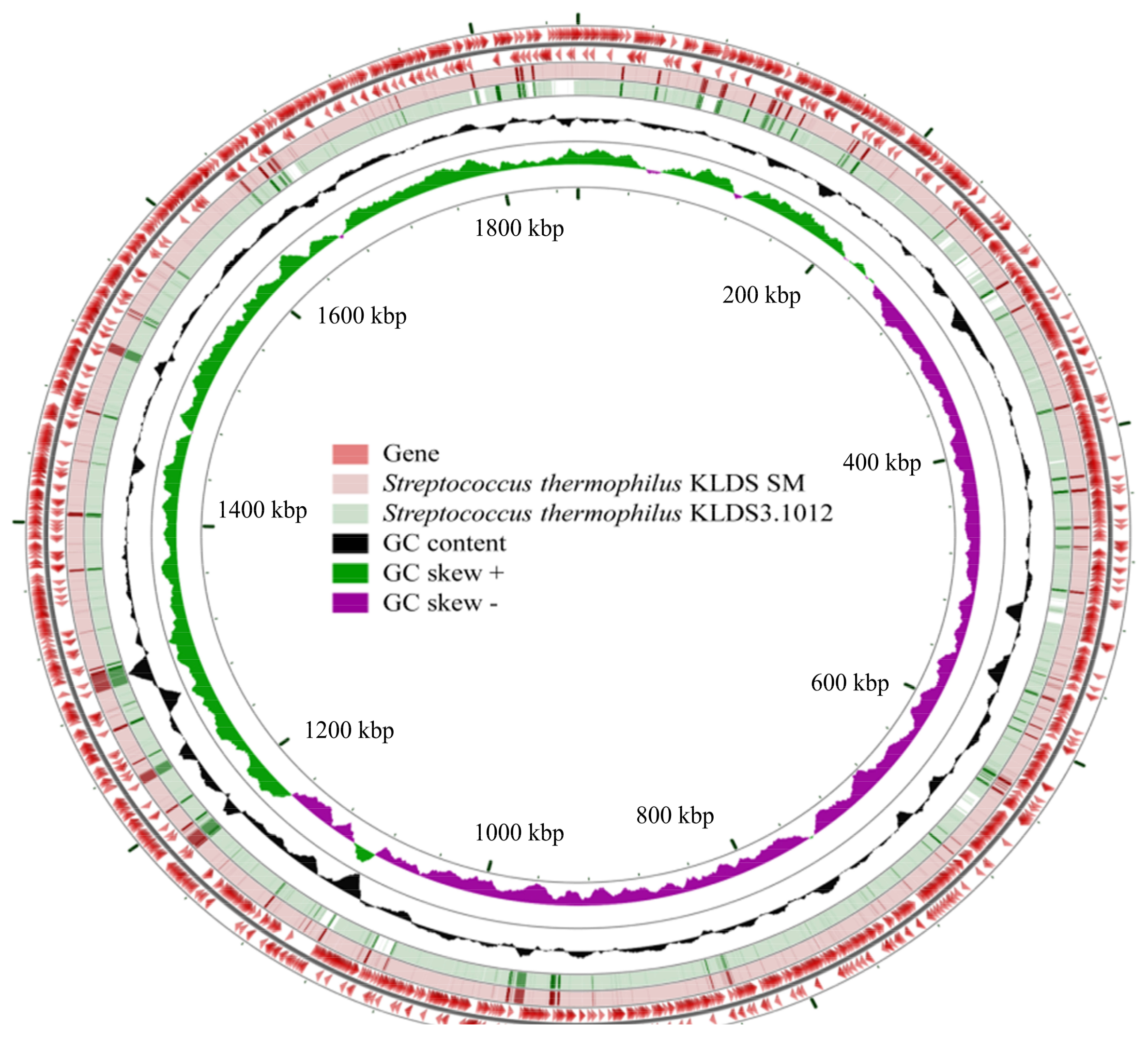
2.2. General Features of Genomes of S. thermophilus
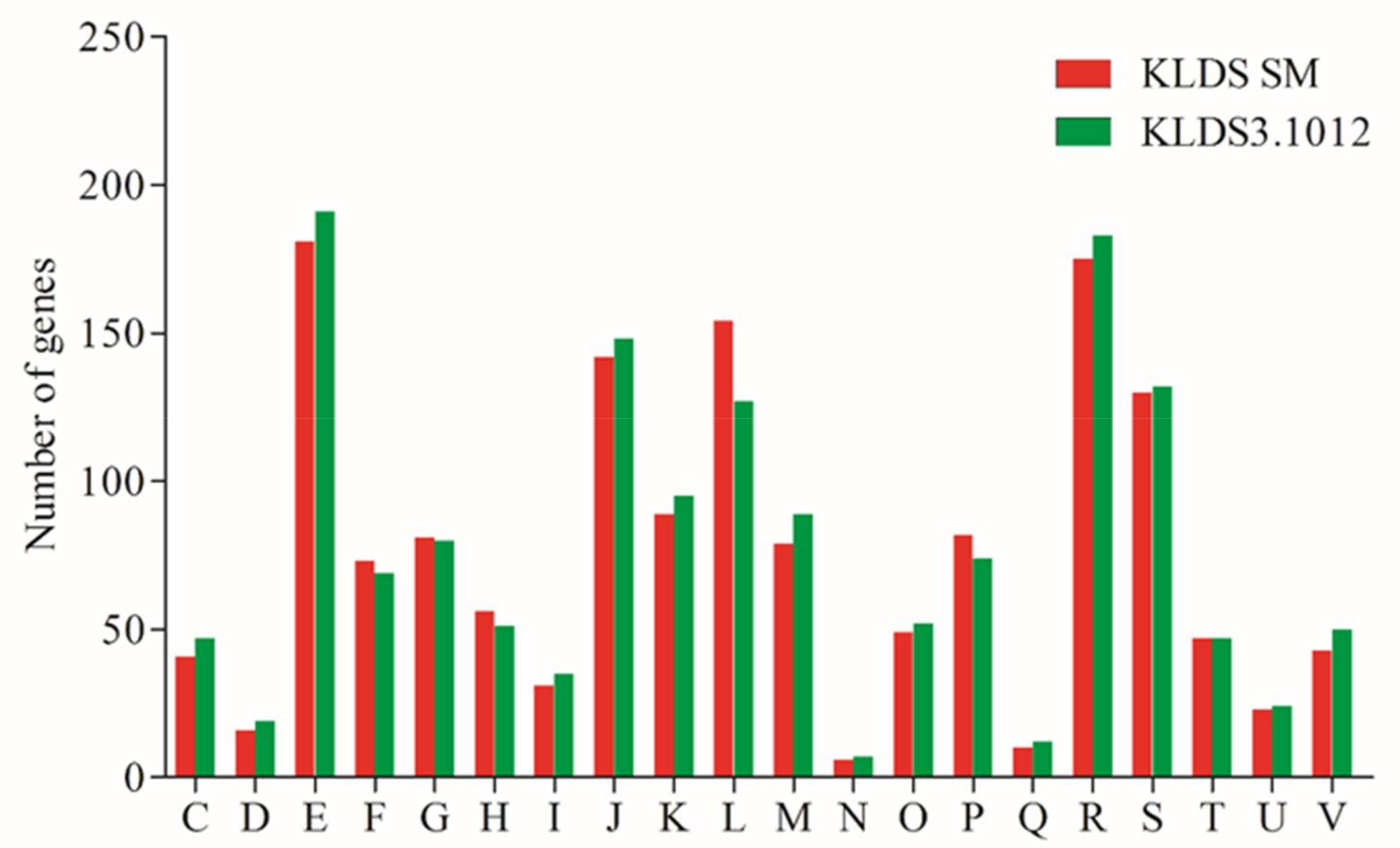
2.3. Proteolytic System and Amino Acid Metabolism of S. thermophilus
2.4. Special prtS Gene Analysis
2.5. Co-Culture of S. thermophilus and Lb. bulgaricus
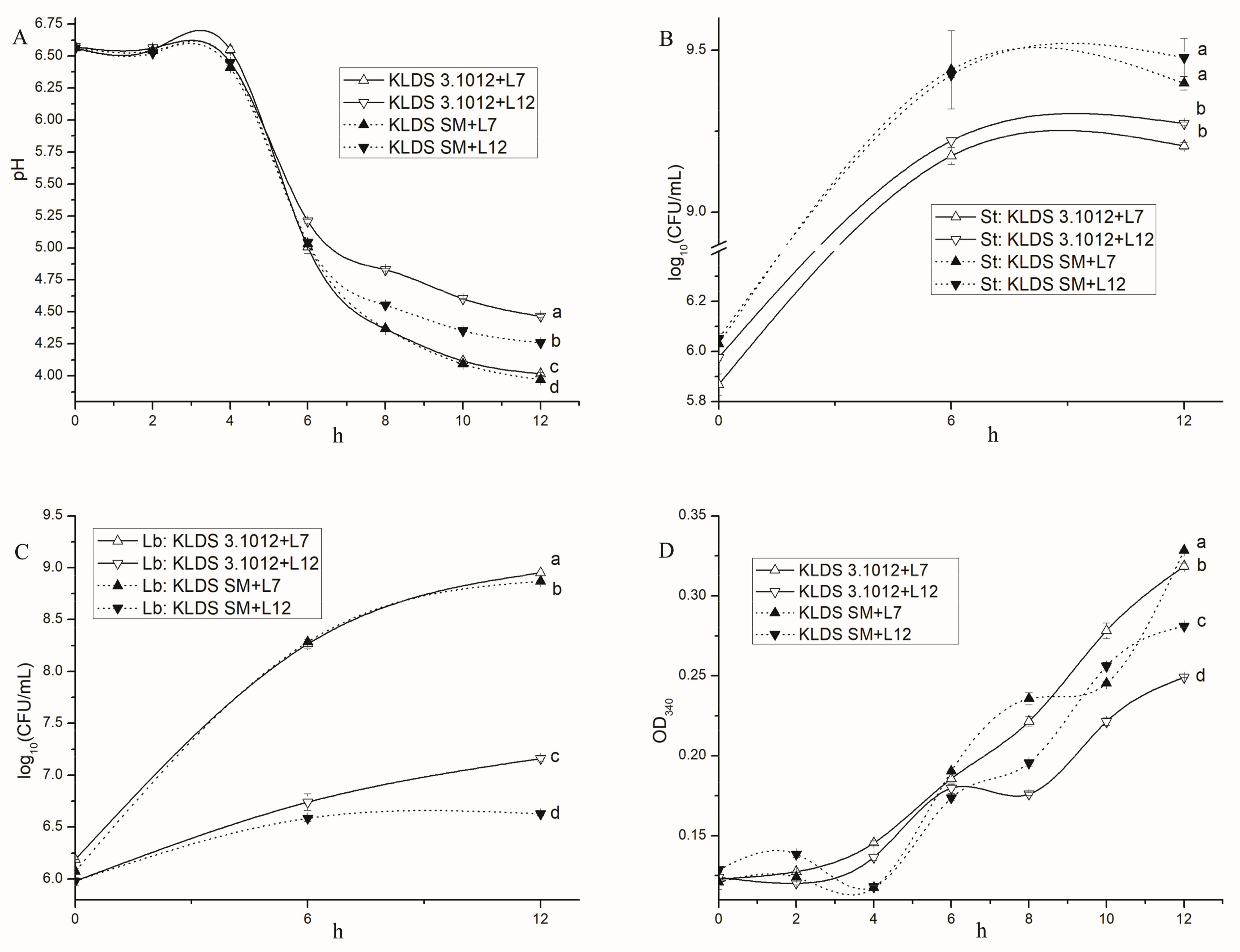
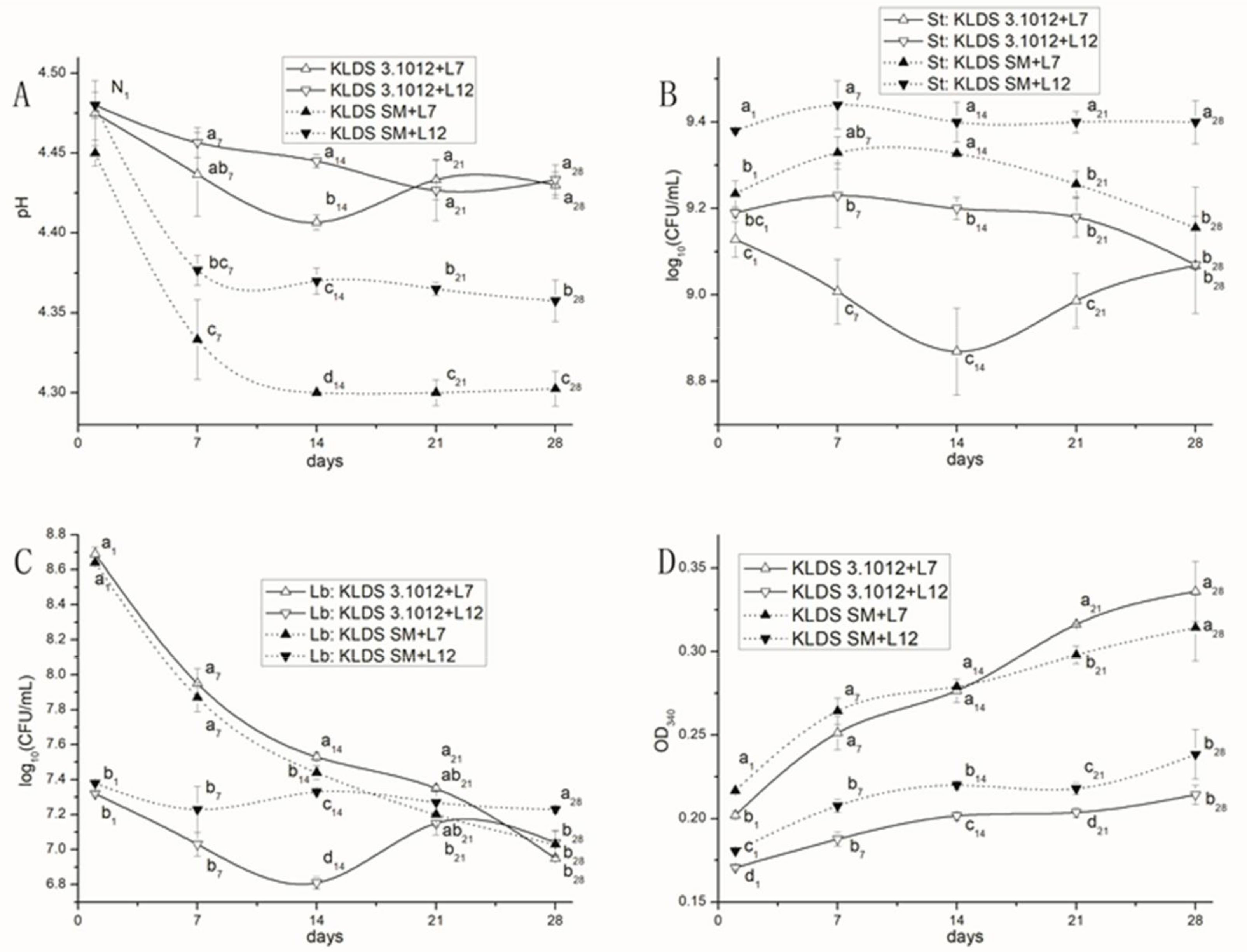
2.5.1. Acid Production
2.5.2. Bacterial Growth
2.5.3. Proteolysis
2.5.4. Effects of PrtS and PrtB on the Proto-Cooperation
2.6. Cold Storage of Yoghurts
2.6.1. Post-Acidification
2.6.2. Survivals of Yoghurt Strains
2.6.3. Proteolysis
3. Materials and Methods
3.1. Strains and Culture Conditions
3.2. Phenotype Terms of Single Strains
3.2.1. Enumeration of Viable Strains and pH Determination
3.2.2. Proteolysis
3.3. Genome Sequencing of S. thermophilus and Analysis in Silico
3.4. Detection of prtS in S. thermophilus
3.5. Co-Culture
3.5.1. Preparation of Starter Cultures
3.5.2. Fermentation and Cold Storage of Yoghurt
3.6. Data Treatment
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ongol, M.P.; Sawatari, Y.; Ebina, Y.; Sone, T.; Tanaka, M.; Tomita, F.; Yokota, A.; Asano, K. Yoghurt fermented by Lactobacillus delbrueckii subsp. bulgaricus H+-ATPase-defective mutants exhibits enhanced viability of Bifidobacterium breve during storage. Int. J. Food Microbiol. 2007, 116, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Tamime, A.Y.; Robinson, R.K. Microbiology of yoghurt and “bio”starter cultures. In Yoghurt: Science and Technology; Woodhead Publishing: Abington, Cambridge, UK, 1999; pp. 407–449. [Google Scholar]
- Zourari, A.; Accolas, J.P.; Desmazeaud, M.J. Metabolism and biochemical characteristics of yogurt bacteria. A review. Dairy Sci. Technol. 1992, 72, 1–34. [Google Scholar] [CrossRef]
- Shahbal, S.; Hemme, D.; Desmazeaud, M. High cell wall-associated proteinase activity of some Streptococcus thermophilus strains (H-strains) correlated with a high acidification rate in milk. Dairy Sci. Technol. 1991, 71, 351–357. [Google Scholar] [CrossRef]
- Rajagopal, S.N.; Sandine, W.E. Associative Growth and Proteolysis of Streptococcus thermophilus and Lactobacillus bulgaricus in Skim Milk1. J. Dairy Sci. 1990, 73, 894–899. [Google Scholar] [CrossRef]
- Kunji, E.R.S.; Mierau, I.; Hagting, A.; Poolman, B.; Konings, W.N. The proteolytic systems of lactic acid bacteria. Antonie van Leeuwenhoek 1996, 70, 187–221. [Google Scholar] [CrossRef] [PubMed]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Ferranti, P.; Smacchi, E.; Goffredi, F.; Addeo, F. Production of angiotensin-I-converting-enzyme-inhibitory peptides in fermented milks started by Lactobacillus delbrueckii subsp. bulgaricus SS1 and Lactococcus lactis subsp. cremoris FT4. Appl. Environ. Microbiol. 2000, 66, 3898–3904. [Google Scholar] [CrossRef] [PubMed]
- Miclo, L.; Roux, E.; Genay, M.; Brusseaux, E.; Poirson, C.; Jameh, N.; Perrin, C.; Dary, A. Variability of hydrolysis of β-, αs1-, and αs2-caseins by 10 strains of Streptococcus thermophilus and resulting bioactive peptides. J. Agric. Food Chem. 2012, 60, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Espla, M.D.; Garault, P.; Monnet, V.; Rul, F. Streptococcus thermophilus cell wall-anchored proteinase: Release, purification, and biochemical and genetic characterization. Appl. Environ. Microbiol. 2000, 66, 4772–4778. [Google Scholar] [CrossRef] [PubMed]
- Shihata, A.; Shah, N.P. Proteolytic profiles of yogurt and probiotic bacteria. Int. Dairy J. 2000, 10, 401–408. [Google Scholar] [CrossRef]
- Delorme, C.; Bartholini, C.; Bolotine, A.; Ehrlich, S.D.; Renault, P. Emergence of a cell wall protease in the Streptococcus thermophilus population. Appl. Environ. Microbiol. 2010, 76, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Settachaimongkon, S.; Nout, M.J.; Antunes, F.E.; Hettinga, K.A.; Vervoort, J.M.; van Hooijdonk, T.C.; Zwietering, M.H.; Smid, E.J.; van Valenberg, H.J. Influence of different proteolytic strains of Streptococcus thermophilus in co-culture with Lactobacillus delbrueckii subsp. bulgaricus on the metabolite profile of set-yoghurt. Int. J. Food Microbiol. 2014, 177, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Dandoy, D.; Fremaux, C.; De Frahan, M.H.; Horvath, P.; Boyaval, P.; Hols, P.; Fontaine, L. The fast milk acidifying phenotype of Streptococcus thermophilus can be acquired by natural transformation of the genomic island encoding the cell-envelope proteinase PrtS. Microb. Cell Fact. 2011, 10, S21. [Google Scholar] [CrossRef] [PubMed]
- Courtin, P.; Monnet, V.; Rul, F. Cell-wall proteinases PrtS and PrtB have a different role in Streptococcus thermophilus/Lactobacillus bulgaricus mixed cultures in milk. Microbiology 2002, 148, 3413–3421. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Erkus, O.; Okuklu, B.; Yenidunya, A.F.; Harsa, S. High genetic and phenotypic variability of Streptococcus thermophilus strains isolated from artisanal Yuruk yoghurts. LWT Food Sci. Technol. 2014, 58, 348–354. [Google Scholar] [CrossRef]
- Pescuma, M.; Hebert, E.M.; Bru, E.; Font, D.V.G.; Mozzi, F. Diversity in growth and protein degradation by dairy relevant lactic acid bacteria species in reconstituted whey. J. Dairy Res. 2012, 79, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Moller, C.; Bockelmann, W.; Ammann, A.; Heller, K.J. Production of yoghurt with mild taste by a Lactobacillus delbrueckii subsp. bulgaricus mutant with altered proteolytic properties. Biotechnol. J. 2007, 2, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Hols, P.; Hancy, F.; Fontaine, L.; Grossiord, B.; Prozzi, D.; Leblond-Bourget, N.; Decaris, B.; Bolotin, A.; Delorme, C.; Dusko Ehrlich, S.; et al. New insights in the molecular biology and physiology of Streptococcus thermophilus revealed by comparative genomics. FEMS Microbiol. Rev. 2005, 29, 435–463. [Google Scholar] [CrossRef] [PubMed]
- Urshev, Z.; Ninovanikolova, N.; Ishlimova, D.; Pashovabaltova, K.; Michaylova, M.; Savova, T. Selection and characterization of naturally occurring high acidification rate Streptococcus thermophilus strains. Infect. Immun. 2014, 28, 899–903. [Google Scholar] [CrossRef]
- Goh, Y.J.; Goin, C.; O’Flaherty, S.; Altermann, E.; Hutkins, R. Specialized adaptation of a lactic acid bacterium to the milk environment: The comparative genomics of Streptococcus thermophilus LMD-9. Microb. Cell Fact. 2011, 10 (Suppl. 1), S22. [Google Scholar] [CrossRef] [PubMed]
- Poquet, I.; Saint, V.; Seznec, E.; Simoes, N.; Bolotin, A.; Gruss, A. HtrA is the unique surface housekeeping protease in Lactococcus lactis and is required for natural protein processing. Mol. Microbiol. 2000, 35, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, J.B.; Zala, H.P.; Nathani, N.M.; Sajnani, M.; Joshi, C.G. Genome-wide analysis of a potent functional dairy starter bacterium Streptococcus thermophilus MTCC 5460: A comprehensive study of its dairy Niche adaptive features. Curr. Sci. India 2017, 113, 2292. [Google Scholar] [CrossRef]
- Christensen, J.E.; Dudley, E.G.; Pederson, J.A.; Steele, J.L. Peptidases and amino acid catabolism in lactic acid bacteria. Antonie van Leeuwenhoek 1999, 76, 217–246. [Google Scholar] [CrossRef] [PubMed]
- Letort, C.; Nardi, M.; Garault, P.; Monnet, V.; Juillard, V. Casein utilization by Streptococcus thermophilus results in a diauxic growth in milk. Appl. Environ. Microbiol. 2002, 68, 3162–3165. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Xu, T.; Qu, X.; Hu, T.; Jiang, X.; Zhao, C. New Insights into Various Production Characteristics of Streptococcus thermophilus Strains. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Abu-Tarboush, H.M. Comparison of Associative Growth and Proteolytic Activity of Yogurt Starters in Whole Milk from Camels and Cows. J. Dairy Sci. 1996, 79, 366–371. [Google Scholar] [CrossRef]
- Brasca, M.; Hogenboom, J.A.; Morandi, S.; Rosi, V.; D’Incecco, P.; Silvetti, T.; Pellegrino, L. Proteolytic Activity and Production of gamma-Aminobutyric Acid by Streptococcus thermophilus Cultivated in Microfiltered Pasteurized Milk. J. Agric. Food Chem. 2016, 64, 8604–8614. [Google Scholar] [CrossRef] [PubMed]
- Sieuwerts, S.; Molenaar, D.; van Hijum, S.A.; Beerthuyzen, M.; Stevens, M.J.; Janssen, P.W.; Ingham, C.J.; de Bok, F.A.; de Vos, W.M.; van Hylckama, V.J. Mixed-culture transcriptome analysis reveals the molecular basis of mixed-culture growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl. Environ. Microbiol. 2010, 76, 7775–7784. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.; Shah, N.P. Cell growth and proteolytic activity of Lactobacillus acidophilus, Lactobacillus helveticus, Lactobacillus delbrueckii ssp. bulgaricus, and Streptococcus thermophilus in milk as affected by supplementation with peptide fractions. Int. J. Food Sci. Nutr. 2014, 65, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Zheng, H.; Hao, P.; Konno, T.; Yu, Y.; Kume, H.; Oda, M.; Ji, Z.S. A model of proteolysis and amino acid biosynthesis for Lactobacillus delbrueckii subsp. bulgaricus in whey. Curr. Microbiol. 2012, 65, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Courtin, P.; Rul, F. Interactions between microorganisms in a simple ecosystem: Yogurt bacteria as a study model. Dairy Sci. Technol. 2004, 84, 125–134. [Google Scholar] [CrossRef]
- Donkor, O.N.; Henriksson, A.; Vasiljevic, T.; Shah, N.P. Effect of acidification on the activity of probiotics in yoghurt during cold storage. Int. Dairy J. 2006, 16, 1181–1189. [Google Scholar] [CrossRef]
- Glusac, J.; Stijepic, M.; Durdevic-Milosevic, D.; Milanovic, S.; Kanuric, K.; Vukic, V. Growth and viability of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus in traditional yoghurt enriched by honey and whey protein concentrate. Iran. J. Vet. Res. 2015, 16, 249–254. [Google Scholar] [PubMed]
- Galia, W.; Perrin, C.; Genay, M.; Dary, A. Variability and molecular typing of Streptococcus thermophilus strains displaying different proteolytic and acidifying properties. Int. Dairy J. 2009, 19, 89–95. [Google Scholar] [CrossRef]
- Prasanna, P.H.P.; Grandison, A.S.; Charalampopoulos, D. Microbiological, chemical and rheological properties of low fat set yoghurt produced with exopolysaccharide (EPS) producing Bifidobacterium strains. Food Res. Int. 2013, 51, 15–22. [Google Scholar] [CrossRef]
- Ramchandran, L.; Shah, N.P. Characterization of functional, biochemical and textural properties of synbiotic low-fat yogurts during refrigerated storage. LWT Food Sci. Technol. 2010, 43, 819–827. [Google Scholar] [CrossRef]
- Germani, A.; Luneia, R.; Nigro, F.; Vitiello, V.; Donini, L.M.; Del, B.V. The yogurt amino acid profile’s variation during the shelf-life. Ann. Ig. 2014, 26, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Ragout, A.; Siñeriz, F. Influence of dilution rate on the morphology and technological properties of Lactobacillus delbrueckii subsp. bulgaricus. Appl. Microbiol. Biot. 1994, 41, 461–464. [Google Scholar] [CrossRef]
- Oliveira, R.P.D.S.; Torres, B.R.; Perego, P.; Oliveira, M.N.D.; Converti, A. Co-metabolic models of Streptococcus thermophilus in co-culture with Lactobacillus bulgaricus or Lactobacillus acidophilus. Biochem. Eng. J. 2012, 62, 62–69. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Li, B.; Ding, X.; Evivie, S.E.; Jin, D.; Meng, Y.; Huo, G.; Liu, F. Short communication: Genomic and phenotypic analyses of exopolysaccharides produced by Streptococcus thermophilus KLDS SM. J. Dairy Sci. 2018, 101, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhu, Z.; Fu, L.; Niu, B.; Li, W. WebMGA: A customizable web server for fast metagenomic sequence analysis. BMC Genom. 2011, 12, 444. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, S.A.; Roberts, R.F. Development of a growth medium suitable for exopolysaccharide production by Lactobacillus delbrueckii ssp. bulgaricus RR. Int. J. Food Microbiol. 1998, 40, 87–92. [Google Scholar] [CrossRef]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Effects of various sugars added to growth and drying media upon thermotolerance and survival throughout storage of freeze-dried Lactobacillus delbrueckii ssp. bulgaricus. Biotechnol. Prog. 2004, 20, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; De Antoni, G.L.; Añón, M.C. Effect of calcium on the cryopreservation of Lactobacillus bulgaricus in different freezing media. Cryobiology 1990, 27, 336–342. [Google Scholar] [CrossRef]
- Ramchandran, L.; Shah, N.P. Growth, proteolytic, and ACE-I activities of Lactobacillus delbrueckii ssp. bulgaricus and Streptococcus thermophilus and rheological properties of low-fat yogurt as influenced by the addition of Raftiline HP. J. Food Sci. 2008, 73, M368–M374. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Encoded Proteins | Genes of Strains | |
|---|---|---|
| KLDS SM | KLDS3.1012 | |
| Exported serine protease HrtA | A9497_06005 | AKL23_RS09565 |
| Protease maturation protein foldase PrsA | A9497_08370 | AKL23_RS02295 |
| Cell envelope serine proteinase PrtS | A9497_00420 | − |
| d-alanyl-d-alanine carboxypeptidase | A9497_06565, ψA9497_06560, ψA9497_01445 | AKL23_RS00470, ψAKL23_RS09800 |
| Specificity | Genes of Strains | Product | |
|---|---|---|---|
| KLDS SM | KLDS3.1012 | ||
| Oligopeptide | A9497_03170 A9497_03160 | AKL23_RS06760 AKL23_RS06745 | OppA; Substrate-binding protein |
| A9497_03155 | AKL23_RS06740 | OppB; Permease protein | |
| A9497_03150 | AKL23_RS06735 | OppC; Permease protein | |
| A9497_03145 | AKL23_RS06730 | OppD; ATP-binding protein | |
| A9497_03140 | AKL23_RS06725 | OppF; ATP-binding protein | |
| Amino acid | A9497_00725 A9497_04145 | AKL23_RS04220 AKL23_RS07755 | ATP-binding cassette (ABC) transporter ATP-binding protein |
| A9497_00720 A9497_04150 A9497_03405 A9497_03795 | AKL23_RS04215 AKL23_RS07760 AKL23_RS07020 AKL23_RS07405 | ABC transporter permease | |
| A9497_04155 A9497_03360 A9497_03370 A9497_07620 | AKL23_RS07765 AKL23_RS06975 AKL23_RS06985 AKL23_RS06990 | ABC transporter substrate-binding protein | |
| A9497_03375 A9497_03785 A9497_07595 | AKL23_RS07395 AKL23_RS01465 AKL23_RS01490 | ||
| Amino acid | A9497_08630 | AKL23_RS02545 | Transporter |
| A9497_02760 A9497_03620 A9497_09055 | AKL23_RS02000 AKL23_RS02915 AKL23_06320 | Permease | |
| Branched-chain amino acid | A9497_00490 A9497_01930 A9497_07910 A9497_07915 | AKL23_RS01775 AKL23_RS01780 AKL23_RS05525 AKL23_RS08285 | ABC transporter permease |
| A9497_04685 A9497_04690 | AKL23_RS08280 AKL23_RS08285 | Permease | |
| A9497_07905 | AKL23_RS01770 | ABC transporter substrate-binding protein | |
| Glutamine | A9497_01790 A9497_06995 A9497_00730 | AKL23_RS04225 AKL23_RS00860 AKL23_RS05335 | ABC transporter substrate-binding protein |
| A9497_01795 A9497_01800 A9497_09175 | AKL23_RS05340 AKL23_RS05345 AKL23_RS03035 | ABC transporter permease | |
| A9497_03790 A9497_09180 | AKL23_RS03040 AKL23_RS07400 | ABC transporter ATP-binding protein | |
| Methionine | A9497_07635 | AKL23_RS01505 | ABC transporter ATP-binding protein |
| A9497_07640 | AKL23_RS01510 | ABC transporter permease | |
| Polar amino acid | A9497_03800 | AKL23_RS07410 | ABC transporter permease |
| Serine/threonine | A9497_07645 | AKL23_RS01515 | Transporter SstT |
| Threonine | A9497_02865 | AKL23_RS06450 | Transporter RhtB |
| Encoded Proteins | Genes of Strains | |
|---|---|---|
| KLDS SM | KLDS3.1012 | |
| Rhomboid family intramembrane serine protease | A9497_05030 | AKL23_RS08625 |
| Membrane-bound protease2C CAAX family | A9497_04220 A9497_05120 A9497_05125 A9497_05130 A9497_05135 ψA9497_09495 | AKL23_RS07830 AKL23_RS08710 AKL23_RS08715 AKL23_RS08720 AKL23_RS08725 ψAKL23_RS10450 |
| Serine protease | A9497_00565 | AKL23_RS04060 |
| C3-degrading protease | A9497_04215 | AKL23_RS07825 |
| CPBP family intramembrane metalloprotease | A9497_09585 | AKL23_RS03380 |
| Metalloprotease | A9497_07170 | AKL23_RS01030 |
| Putative Zn-dependent protease | A9497_01910 | AKL23_RS05505 |
| ATP-dependent Zn protease | A9497_02810 | AKL23_RS06395 |
| Zinc protease | A9497_00005 | AKL23_RS03520 |
| Aminopeptidase T | A9497_06485 | AKL23_RS00400 |
| Aminopeptidase N | A9497_01225 | AKL23_RS04750 |
| Aminopeptidase C | A9497_07295 | AKL23_RS01165 |
| Xaa-Pro aminopeptidase | A9497_09300 A9497_04560 | AKL23_RS08155 AKL23_RS03160 |
| Tripeptide aminopeptidase | A9497_01685 | AKL23_RS05230 |
| Glutamyl aminopeptidase | A9497_05110 | AKL23_RS08700 |
| Methionine aminopeptidase | A9497_03680 | AKL23_RS07290 |
| d-alanyl-d-alanine carboxypeptidase | A9497_06720 | AKL23_RS00620 |
| Dipeptidase | A9497_01640 ψA9497_00590 | AKL23_RS04085 ψAKL23_RS10110 |
| Xaa-Pro dipeptidyl-peptidase | A9497_04230 | AKL23_RS07840 |
| Metalloendopeptidases | A9497_03000 | AKL23_RS06585 |
| Endopeptidase | A9497_05285 | AKL23_RS08875 |
| Oligoendopeptidase F | A9497_02650, A9497_08360 | AKL23_RS02285 |
| O-sialoglycoprotein endopeptidase | A9497_04050 A9497_07625 | AKL23_RS07660 AKL23_RS01495 |
| Peptidase M16 | A9497_05935 A9497_05940 | AKL23_RS09495 AKL23_RS09500 |
| Peptidase M20 | A9497_08905 ψA9497_07630 | AKL23_RS02765 ψAKL23_RS01500 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, H.; Li, B.; Evivie, S.E.; Sarker, S.K.; Chowdhury, S.; Lu, J.; Ding, X.; Huo, G. Technological and Genomic Analysis of Roles of the Cell-Envelope Protease PrtS in Yoghurt Starter Development. Int. J. Mol. Sci. 2018, 19, 1068. https://doi.org/10.3390/ijms19041068
Tian H, Li B, Evivie SE, Sarker SK, Chowdhury S, Lu J, Ding X, Huo G. Technological and Genomic Analysis of Roles of the Cell-Envelope Protease PrtS in Yoghurt Starter Development. International Journal of Molecular Sciences. 2018; 19(4):1068. https://doi.org/10.3390/ijms19041068
Chicago/Turabian StyleTian, Hui, Bailiang Li, Smith Etareri Evivie, Shuvan Kumar Sarker, Sathi Chowdhury, Jingjing Lu, Xiuyun Ding, and Guicheng Huo. 2018. "Technological and Genomic Analysis of Roles of the Cell-Envelope Protease PrtS in Yoghurt Starter Development" International Journal of Molecular Sciences 19, no. 4: 1068. https://doi.org/10.3390/ijms19041068
APA StyleTian, H., Li, B., Evivie, S. E., Sarker, S. K., Chowdhury, S., Lu, J., Ding, X., & Huo, G. (2018). Technological and Genomic Analysis of Roles of the Cell-Envelope Protease PrtS in Yoghurt Starter Development. International Journal of Molecular Sciences, 19(4), 1068. https://doi.org/10.3390/ijms19041068