TASK-3 Downregulation Triggers Cellular Senescence and Growth Inhibition in Breast Cancer Cell Lines

Abstract

1. Introduction
2. Results
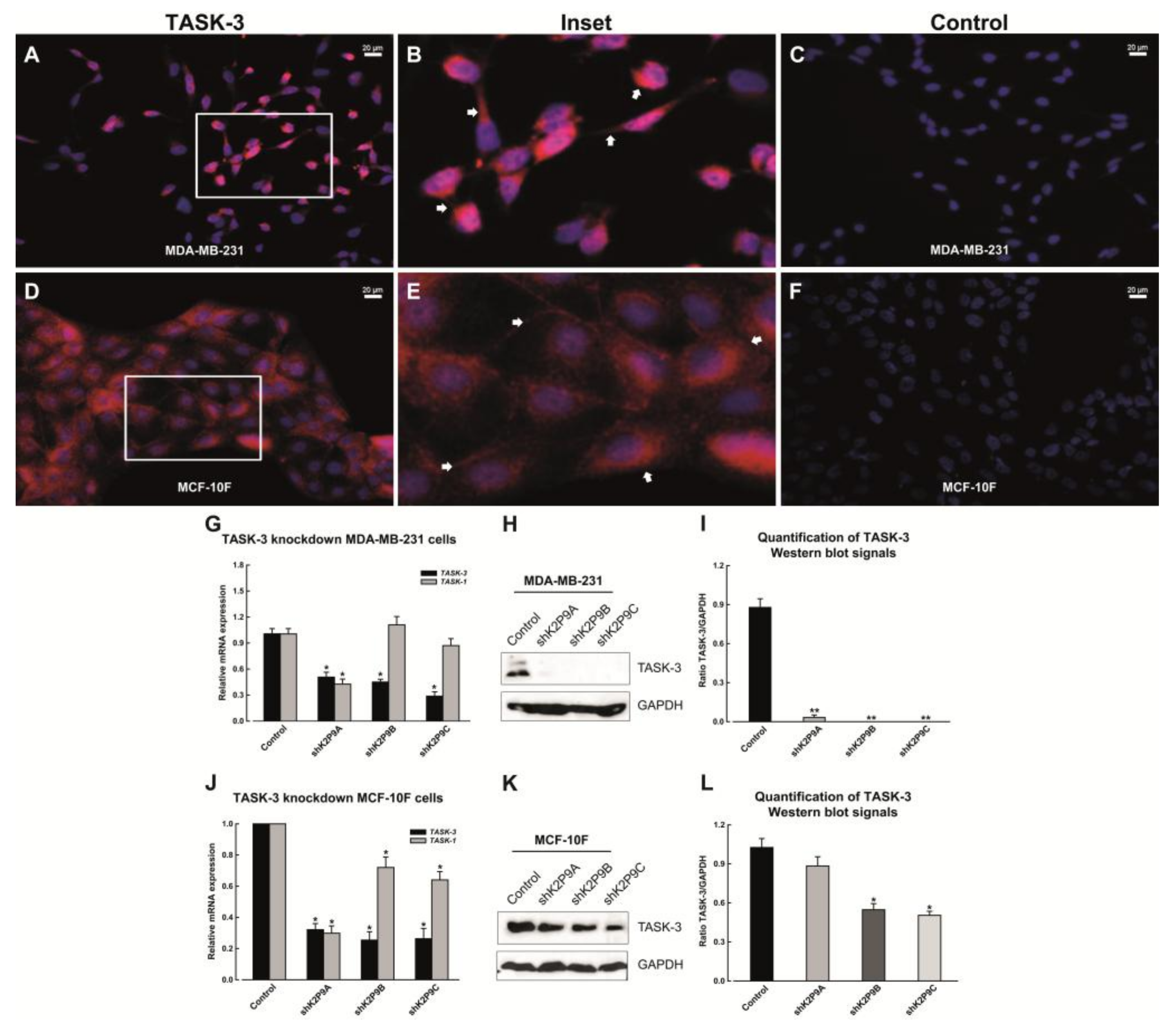
2.1. Expression of TASK-3 Channels in MDA-MB-231 and MCF-10F Cells
2.2. Short Hairpin RNA-Mediated Knockdown of TASK-3
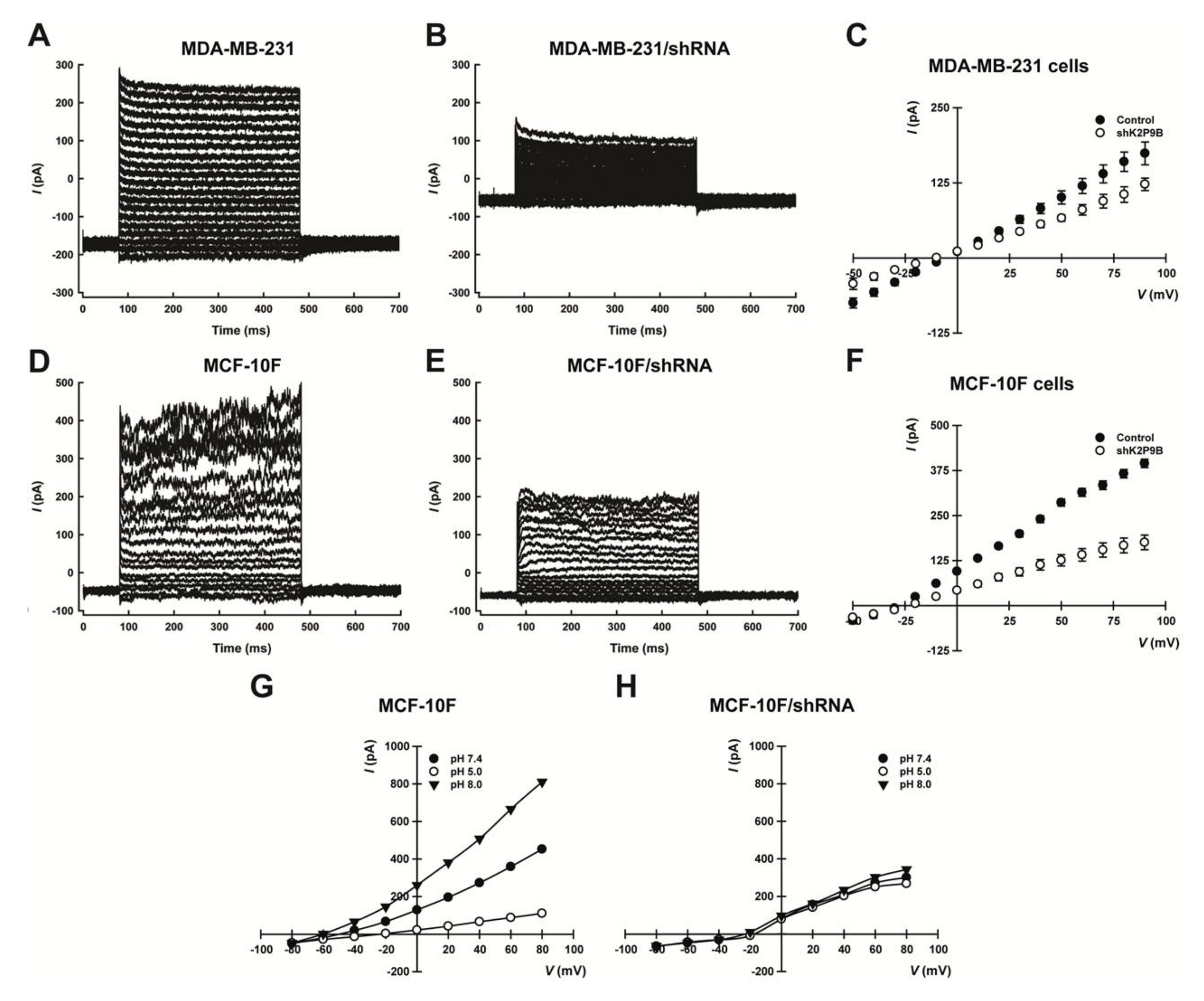
2.3. Electrophysiological Characterization of TASK-3 Knockdown
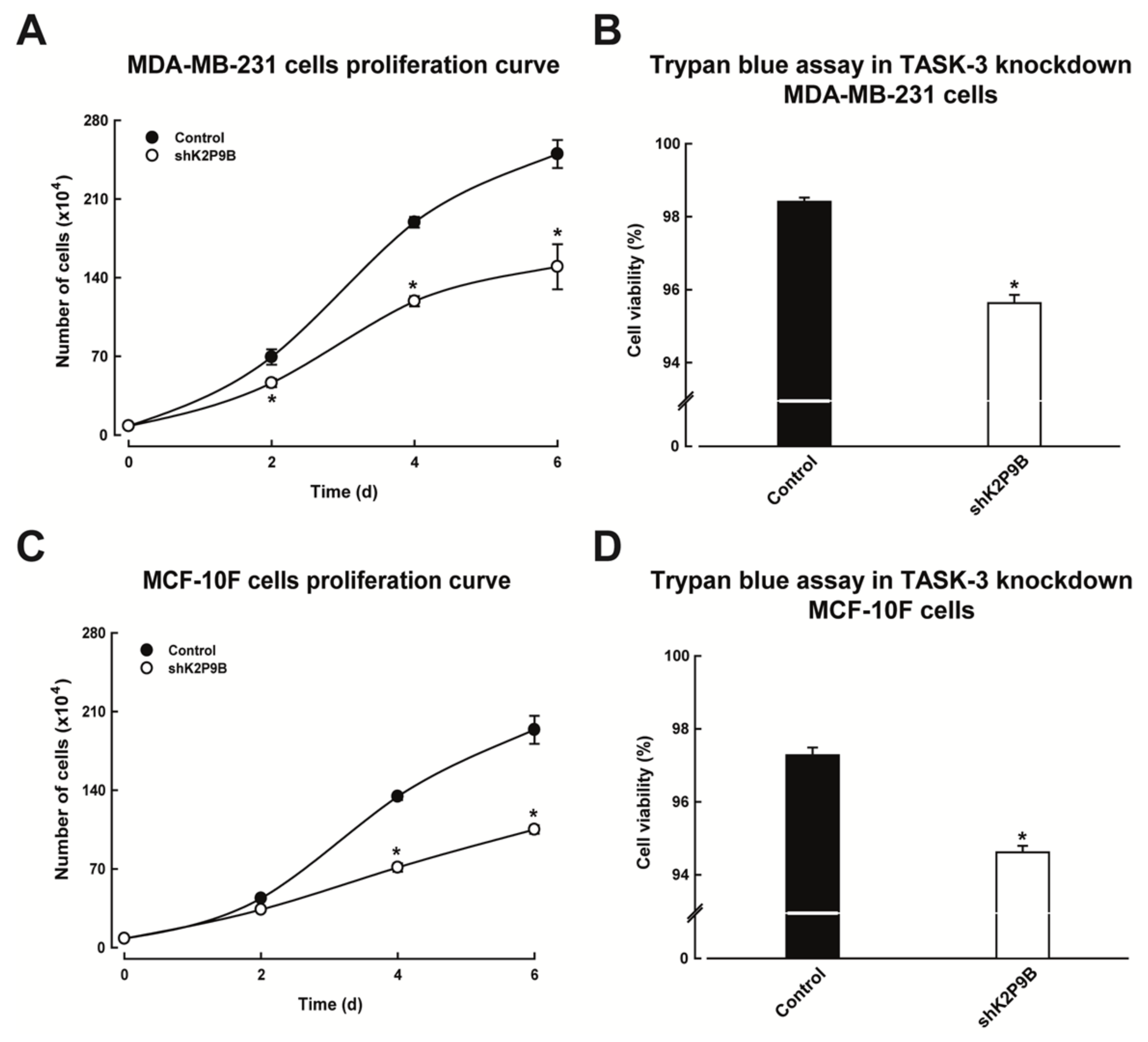
2.4. Reducing the Expression of TASK-3 Inhibits Proliferation of MDA-MB-231 and MCF-10F Cells
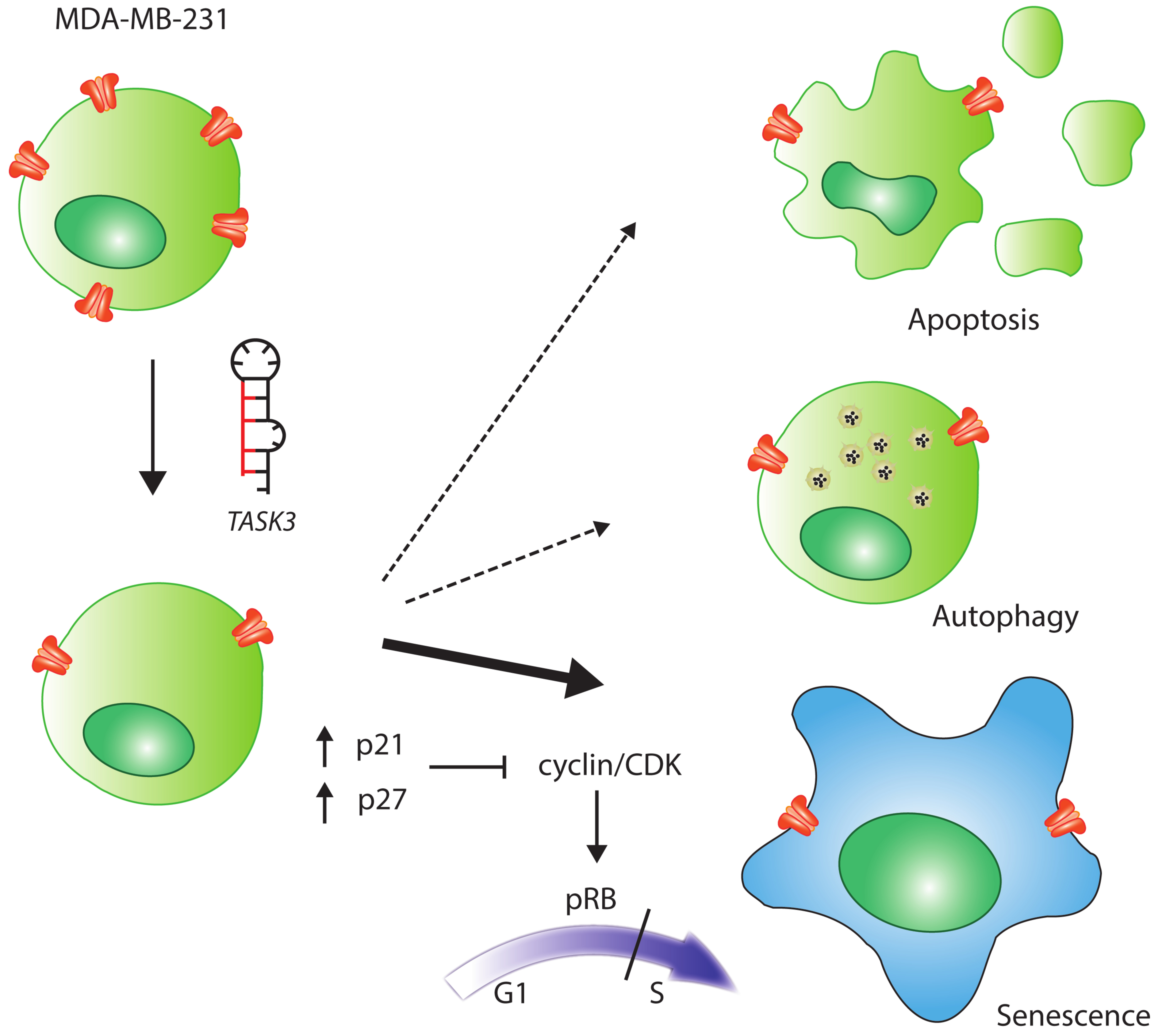
2.5. Cellular Senescence and Autophagy in TASK-3-Depleted MDA-MB-231 and MCF-10F Cells
2.6. TASK-3 Knockdown in MDA-MB-231 and MCF-10F Cells Is Not Accompanied by an Increased Rate of Apoptosis
2.7. Analysis of the Cell Cycle Regulators
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. Immunocytochemistry
4.3. TASK-3 Silencing with Short Hairpin RNA (shRNA)
4.4. Patch-Clamp Recording
4.5. Trypan Blue Exclusion
4.6. Extraction and Quantification of mRNA by Real Time PCR
4.7. Protein Extraction and Western Blotting
4.8. Senescence-Associated β-Galactosidase Staining Assay
4.9. Assessment of Apoptosis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kamangar, F.; Dores, G.M.; Anderson, W.F. Patterns of cancer incidence, mortality, and prevalence across five continents: Defining priorities to reduce cancer disparities in different geographic regions of the world. J. Clin. Oncol. 2006, 24, 2137–2150. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.T.; Goodin, S. Overcoming drug resistance in patients with metastatic breast cancer. Pharmacotherapy 2009, 29, 954–965. [Google Scholar] [CrossRef] [PubMed]
- Ouadid-Ahidouch, H.; Ahidouch, A. K+ channel expression in human breast cancer cells: Involvement in cell cycle regulation and carcinogenesis. J. Membr. Biol. 2008, 221, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bittner, S.; Budde, T.; Wiendl, H.; Meuth, S.G. From the background to the spotlight: Task channels in pathological conditions. Brain Pathol. 2010, 20, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Stühmer, W.; Alves, F.; Hartung, F.; Zientkowska, M.; Pardo, L.A. Potassium channels as tumour markers. FEBS Lett. 2006, 580, 2850–2852. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, M.; Gasparoli, L.; Arcangeli, A. Potassium channels: Novel emerging biomarkers and targets for therapy in cancer. Recent Pat. Anti-Cancer Drug Discov. 2013, 8, 53–65. [Google Scholar] [CrossRef]
- Sontheimer, H. An unexpected role for ion channels in brain tumor metastasis. Exp. Biol. Med. 2008, 233, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion channels and the hallmarks of cancer. Trends Mol. Med. 2010, 16, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.F.; Stock, C. Ion channels and transporters in cancer: Pathophysiology, regulation, and clinical potential. Cancer Res. 2013, 73, 1658–1661. [Google Scholar] [CrossRef] [PubMed]
- Pardo, L.A.; Stühmer, W. The roles of K+ channels in cancer. Nat. Rev. Cancer 2014, 14, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xiong, Z.-G. Ion channels as targets for cancer therapy. Int. J. Phys. Pathophysiol. Pharmacol. 2011, 3, 156–166. [Google Scholar]
- Kunzelmann, K. Ion channels and cancer. J. Membr. Biol. 2005, 205, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Plant, L.D.; Zuniga, L.; Araki, D.; Marks, J.D.; Goldstein, S.A. SUMOylation silences heterodimeric TASK potassium channels containing K2P1 subunits in cerebellar granule neurons. Sci. Signal. 2012, 5, ra84. [Google Scholar] [CrossRef] [PubMed]
- Burgos, P.; Zuniga, R.; Dominguez, P.; Delgado-Lopez, F.; Plant, L.D.; Zuniga, L. Differential expression of two-pore domain potassium channels in rat cerebellar granule neurons. Biochem. Biophys. Res. Commun. 2014, 453, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. Roles of K+ channels in regulating tumour cell proliferation and apoptosis. Pflugers Arch. Eur. J. Phys. 2004, 448, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Jan, L.Y. Targeting potassium channels in cancer. J. Cell Biol. 2014, 206, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Schwab, A.; Fabian, A.; Hanley, P.J.; Stock, C. Role of ion channels and transporters in cell migration. Physiol. Rev. 2012, 92, 1865–1913. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Föller, M.; Lang, K.S.; Lang, P.A.; Ritter, M.; Gulbins, E.; Vereninov, A.; Huber, S.M. Ion channels in cell proliferation and apoptotic cell death. J. Membr. Biol. 2005, 205, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.A.; Bockenhauer, D.; O'Kelly, I.; Zilberberg, N. Potassium leak channels and the KCNK family of two-P-domain subunits. Nat. Rev. Neurosci. 2001, 2, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga, L.; Zúñiga, R. Understanding the Cap structure in K2P channels. Front. Physiol. 2016, 7, 228. [Google Scholar] [CrossRef] [PubMed]
- Lotshaw, D.P. Biophysical, pharmacological, and functional characteristics of cloned and native mammalian two-pore domain K+ channels. Cell Biochem. Biophys. 2007, 47, 209–256. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.; Bateman, A.; O’Kelly, I. Altered expression of two-pore domain potassium (K2P) channels in cancer. PLoS ONE 2013, 8, e74589. [Google Scholar] [CrossRef] [PubMed]
- Voloshyna, I.; Besana, A.; Castillo, M.; Matos, T.; Weinstein, I.B.; Mansukhani, M.; Robinson, R.B.; Cordon-Cardo, C.; Feinmark, S.J. TREK-1 is a novel molecular target in prostate cancer. Cancer Res. 2008, 68, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.; Wiser, O.; Slavin, A.; Mu, D.; Powers, S.; Jan, L.Y.; Hoey, T. Oncogenic potential of TASK3 (KCNK9) depends on K+ channel function. Proc. Natl. Acad. Sci. USA 2003, 100, 7803–7807. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.P.; Talley, E.M.; Manger, J.P.; Bayliss, D.A. Motoneurons express heteromeric TWIK-related acid-sensitive K+ (TASK) channels containing TASK-1 (KCNK3) and TASK-3 (KCNK9) subunits. J. Neurosci. 2004, 24, 6693–6702. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Han, J.; Talley, E.M.; Bayliss, D.A.; Kim, D. Functional expression of TASK-1/TASK-3 heteromers in cerebellar granule cells. J. Physiol. 2004, 554, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Enyedi, P.; Czirják, G. Molecular background of leak K+ currents: Two-pore domain potassium channels. Physiol. Rev. 2010, 90, 559–605. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Chen, L.; Zhang, X.; See, L.-H.; Koch, C.M.; Yen, C.; Tong, J.J.; Spiegel, L.; Nguyen, K.C.; Servoss, A.; et al. Genomic amplification and oncogenic properties of the KCNK9 potassium channel gene. Cancer Cell 2003, 3, 297–302. [Google Scholar] [CrossRef]
- Innamaa, A.; Jackson, L.; Asher, V.; Van Shalkwyk, G.; Warren, A.; Hay, D.; Bali, A.; Sowter, H.; Khan, R. Expression and prognostic significance of the oncogenic K2P potassium channel KCNK9 (TASK-3) in ovarian carcinoma. Anticancer Res. 2013, 33, 1401–1408. [Google Scholar] [PubMed]
- Kim, C.J.; Cho, Y.G.; Jeong, S.W.; Kim, Y.S.; Kim, S.Y.; Nam, S.W.; Lee, S.H.; Yoo, N.J.; Lee, J.Y.; Park, W.S. Altered expression of KCNK9 in colorectal cancers. APMIS 2004, 112, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Luo, L.; Lal, B.; Ma, X.; Chen, L.; Hann, C.L.; Fulton, A.M.; Leahy, D.J.; Laterra, J.; Li, M. A monoclonal antibody against KCNK9 K+ channel extracellular domain inhibits tumour growth and metastasis. Nat. Commun. 2016, 7, 10339. [Google Scholar] [CrossRef] [PubMed]
- Pardo, L.A.; Stühmer, W. Eag1: An emerging oncological target. Cancer Res. 2008, 68, 1611–1613. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Zheng, Y.; Yan, Y.; Bargonetti, J.; Foster, D.A. Mutant p53 in MDA-MB-231 breast cancer cells is stabilized by elevated phospholipase D activity and contributes to survival signals generated by phospholipase D. Oncogene 2006, 25, 7305–7310. [Google Scholar] [CrossRef] [PubMed]
- Duprat, F.; Lesage, F.; Fink, M.; Reyes, R.; Heurteaux, C.; Lazdunski, M. TASK, a human background K+ channel to sense external pH variations near physiological pH. EMBO J. 1997, 16, 5464–5471. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Bang, H.; Kim, D. TASK-3, a new member of the tandem pore K+ channel family. J. Biol. Chem. 2000, 275, 9340–9347. [Google Scholar] [CrossRef] [PubMed]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 conjugation system in mammalian autophagy. Int. J. Biochem. Cell Biol. 2004, 36, 2503–2518. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast APG8P, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, I.; Zanzouri, M.; Honoré, E.; Duprat, F.; Ehrengruber, M.U.; Lazdunski, M.; Patel, A.J. K+-dependent cerebellar granule neuron apoptosis: Role of task leak K+ channels. J. Biol. Chem. 2003, 278, 32068–32076. [Google Scholar] [CrossRef] [PubMed]
- Meuth, S.G.; Herrmann, A.M.; Ip, C.W.; Kanyshkova, T.; Bittner, S.; Weishaupt, A.; Budde, T.; Wiendl, H. The two-pore domain potassium channel TASK3 functionally impacts glioma cell death. J. Neurooncol. 2008, 87, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Nagy, D.; Gönczi, M.; Dienes, B.; Szöor, Á.; Fodor, J.; Nagy, Z.; Tóth, A.; Fodor, T.; Bai, P.; Szücs, G.; et al. Silencing the KCNK9 potassium channel (TASK-3) gene disturbs mitochondrial function, causes mitochondrial depolarization, and induces apoptosis of human melanoma cells. Arch. Dermatol. Res. 2014, 306, 885–902. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.-W.; Park, H.S.; Kim, E.-J.; Cho, Y.-W.; Kim, G.-T.; Mun, Y.-J.; Choi, E.-J.; Lee, J.-S.; Han, J.; Kang, D. Reduction of breast cancer cell migration via up-regulation of TASK-3 two-pore domain k+ channel. Acta Physiol. 2012, 204, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Pocsai, K.; Kosztka, L.; Bakondi, G.; Gönczi, M.; Fodor, J.; Dienes, B.; Szentesi, P.; Kovács, I.; Feniger-Barish, R.; Kopf, E.; et al. Melanoma cells exhibit strong intracellular TASK-3-specific immunopositivity in both tissue sections and cell culture. Cell. Mol. Life Sci. 2006, 63, 2364–2376. [Google Scholar] [CrossRef] [PubMed]
- Kosztka, L.; Rusznák, Z.; Nagy, D.; Nagy, Z.; Fodor, J.; Szucs, G.; Telek, A.; Gönczi, M.; Ruzsnavszky, O.; Szentandrássy, N.; et al. Inhibition of TASK-3 (KCNK9) channel biosynthesis changes cell morphology and decreases both DNA content and mitochondrial function of melanoma cells maintained in cell culture. Melanoma Res. 2011, 21, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Shen, Y.; Zheng, Y.; Joseph, T.; Jackson, D.; Foster, D.A. Phospholipase D prevents apoptosis in v-Src-transformed rat fibroblasts and MDA-MB-231 breast cancer cells. Biochem. Biophys. Res. Commun. 2003, 302, 615–619. [Google Scholar] [CrossRef]
- Leanza, L.; O'Reilly, P.; Doyle, A.; Venturini, E.; Zoratti, M.; Szegezdi, E.; Szabo, I. Correlation between potassium channel expression and sensitivity to drug-induced cell death in tumor cell lines. Curr. Pharm. Des. 2014, 20, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Majumder, P.K.; Grisanzio, C.; O'Connell, F.; Barry, M.; Brito, J.M.; Xu, Q.; Guney, I.; Berger, R.; Herman, P.; Bikoff, R.; et al. A prostatic intraepithelial neoplasia-dependent p27kip1 checkpoint induces senescence and inhibits cell proliferation and cancer progression. Cancer Cell 2008, 14, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Young, A.P.; Schlisio, S.; Minamishima, Y.A.; Zhang, Q.; Li, L.; Grisanzio, C.; Signoretti, S.; Kaelin, W.G. VHL loss actuates a HIF-independent senescence programme mediated by Rb and p400. Nat. Cell Biol. 2008, 10, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, K.; Van Bockstaele, D.R.; Berneman, Z.N. The cell cycle: A review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003, 36, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Walworth, N.C. Cell-cycle checkpoint kinases: Checking in on the cell cycle. Curr. Opin. Cell Biol. 2000, 12, 697–704. [Google Scholar] [CrossRef]
- Ukomadu, C.; Dutta, A. P21 dependent inhibition of colon cancer cell growth by mevastatin is independent of inhibition of G1 cyclin-dependent kinases. J. Biol. Chem. 2003, 278, 43586–43594. [Google Scholar] [CrossRef] [PubMed]
- Comes, N.; Serrano-Albarrás, A.; Capera, J.; Serrano-Novillo, C.; Condom, E.; Cajal, S.; Ferreres, J.C.; Felipe, A. Involvement of potassium channels in the progression of cancer to a more malignant phenotype. Biochim. Biophys. Acta 2015, 1848, 2477–2492. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga, L.; Márquez, V.; González-Nilo, F.D.; Chipot, C.; Cid, L.P.; Sepúlveda, F.V.; Niemeyer, M.I. Gating of a pH-sensitive K2P potassium channel by an electrostatic effect of basic sensor residues on the selectivity filter. PLoS ONE 2011, 6, e16141. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Common Name | GenBank Accession | Abbreviation | Primer Pair, Sense (5′–3′) | Product Size (bp) |
|---|---|---|---|---|---|
| KCNK9 | K2P9, TASK-3 | NM_001282534.1 | qHsKCNK9_F1 | 5′-GTC TCA TTT TCC CCC ACC TTT CCA G-3′ | 148 |
| qHsKCNK9_R1 | 5′-GGG TGG GGT GAG AAA TGT AAG GCA-3′ | ||||
| KCNK3 | K2P3, TASK-1 | NM_002246.2 | qHnP3_F | 5′-GGT GCT CAT CGG CTT CTT CT-3′ | 199 |
| qHnP3_R | 5′-GAA GCT GAA GGC CAC GTA CT-3′ | ||||
| CCNA1 | Cyclin A1 | NM_003914.3 | qHsCCNA1_F | 5′-TGA AAT AAG GCA CAG ACC CAA AGC A-3′ | 89 |
| qHsCCNA1_R | 5′-ACC AGC CAG TCC ACC AGA ATC GT-3′ | ||||
| CCND1 | Cyclin D1 | NM_053056.2 | qHsCCND1_F | 5′-GCT CCT GTG CTG CGA AGT GGA A-3′ | 126 |
| qHsCCND1_R | 5′-TTT GAA GTA GGA CAC CGA GGG CG-3′ | ||||
| CCNE1 | Cyclin E1 | NM_001238.2 | qHsCCNE1_F | 5′-AAG GTT TCA GGG TAT CAG TGG TGC G-3′ | 191 |
| qHsCCNE1_R | 5′-GGC TTT CTT TGC TCG GGC TTT G-3′ | ||||
| CDK4 | CDK4 | NM_000075.3 | qHsCDK4_F | 5′-TCG TGA GGT GGC TTT ACT GAG GCG-3′ | 194 |
| qHsCDK4_R | 5′-TCC TTG ATC GTT TCG GCT GGC A-3′ | ||||
| CDKN1A | p21, Cip1 | NM_001291549.1 | qHsCDKN1A_F | 5′-TGT CCG TCA GAA CCC ATG C-3′ | 139 |
| qHsCDKN1A_R | 5′-AAA GTC GAA GTT CCA TCG CTC-3′ | ||||
| CDKN1B | p27, Kip1 | NM_004064.4 | qHsCDKN1B_F | 5′-GGG TCT GTG TCT TTT GGC TCC GA-3′ | 94 |
| qHsCDKN1B_R | 5′-CCG CCT CTC TCG CAC TCT CAA A-3′ | ||||
| RPL19 | L19 | NM_000981 | qHsRPL19_F | 5′-CAT CCG CAA GCC TGT GAC G-3′ | 132 |
| qHsRPL19_R | 5′-TGT GAC CTT CTC TGG CAT TCG-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zúñiga, R.; Valenzuela, C.; Concha, G.; Brown, N.; Zúñiga, L. TASK-3 Downregulation Triggers Cellular Senescence and Growth Inhibition in Breast Cancer Cell Lines. Int. J. Mol. Sci. 2018, 19, 1033. https://doi.org/10.3390/ijms19041033
Zúñiga R, Valenzuela C, Concha G, Brown N, Zúñiga L. TASK-3 Downregulation Triggers Cellular Senescence and Growth Inhibition in Breast Cancer Cell Lines. International Journal of Molecular Sciences. 2018; 19(4):1033. https://doi.org/10.3390/ijms19041033
Chicago/Turabian StyleZúñiga, Rafael, Claudio Valenzuela, Guierdy Concha, Nelson Brown, and Leandro Zúñiga. 2018. "TASK-3 Downregulation Triggers Cellular Senescence and Growth Inhibition in Breast Cancer Cell Lines" International Journal of Molecular Sciences 19, no. 4: 1033. https://doi.org/10.3390/ijms19041033
APA StyleZúñiga, R., Valenzuela, C., Concha, G., Brown, N., & Zúñiga, L. (2018). TASK-3 Downregulation Triggers Cellular Senescence and Growth Inhibition in Breast Cancer Cell Lines. International Journal of Molecular Sciences, 19(4), 1033. https://doi.org/10.3390/ijms19041033