cAMP/Protein Kinase A Signaling Inhibits Dlx5 Expression via Activation of CREB and Subsequent C/EBPβ Induction in 3T3-L1 Preadipocytes

 , ,
, , 
Abstract
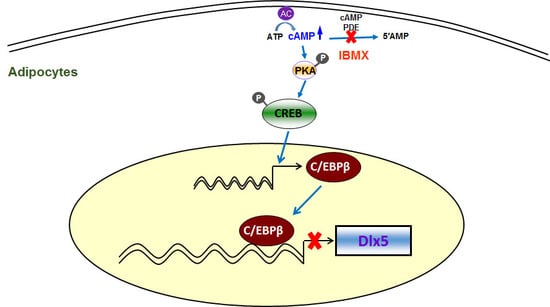
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
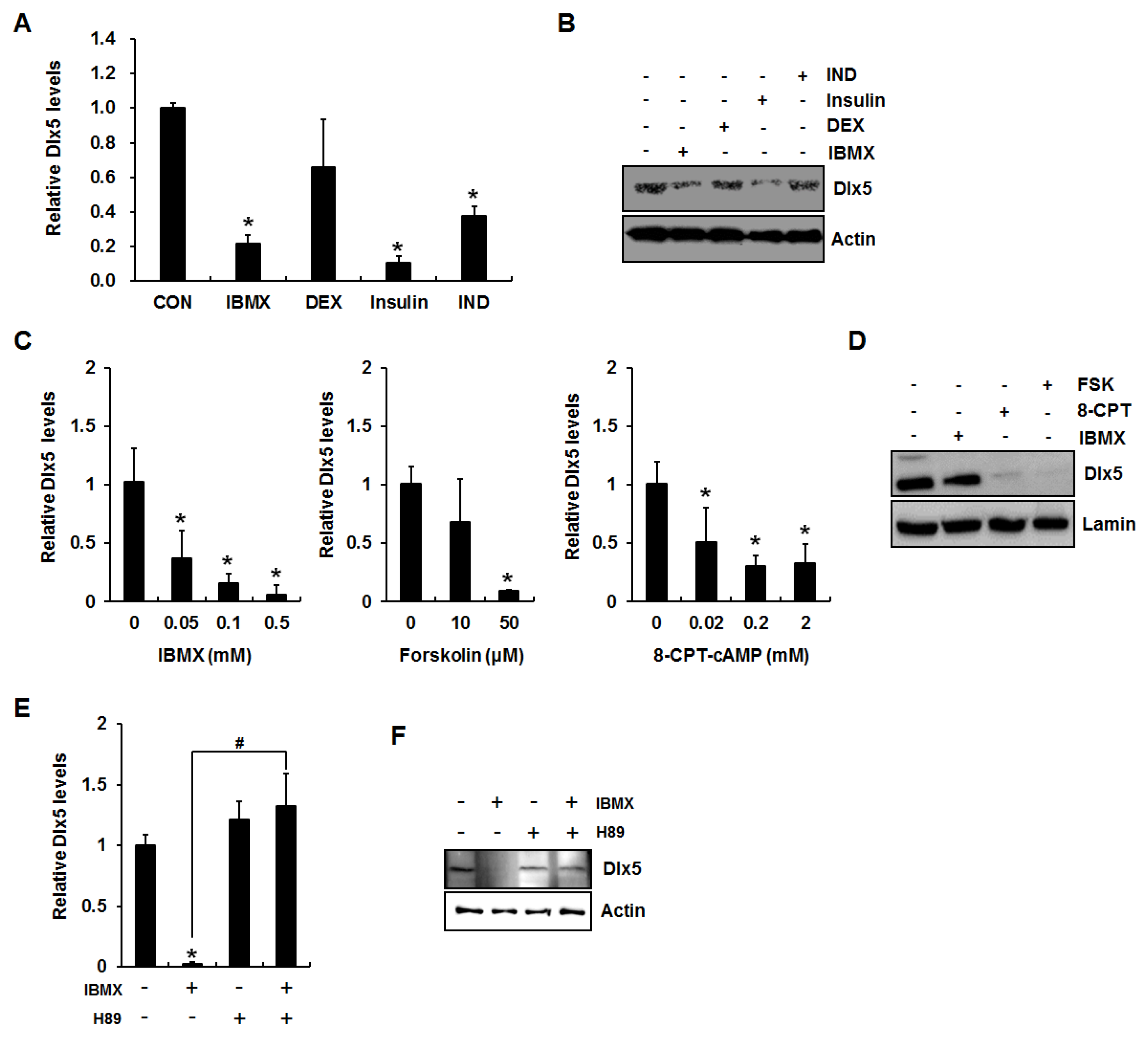
2.1. Activation of cAMP/PKA Signaling Inhibits Distal-Less Homeobox 5 (Dlx5) Expression in 3T3-L1 Preadipocytes
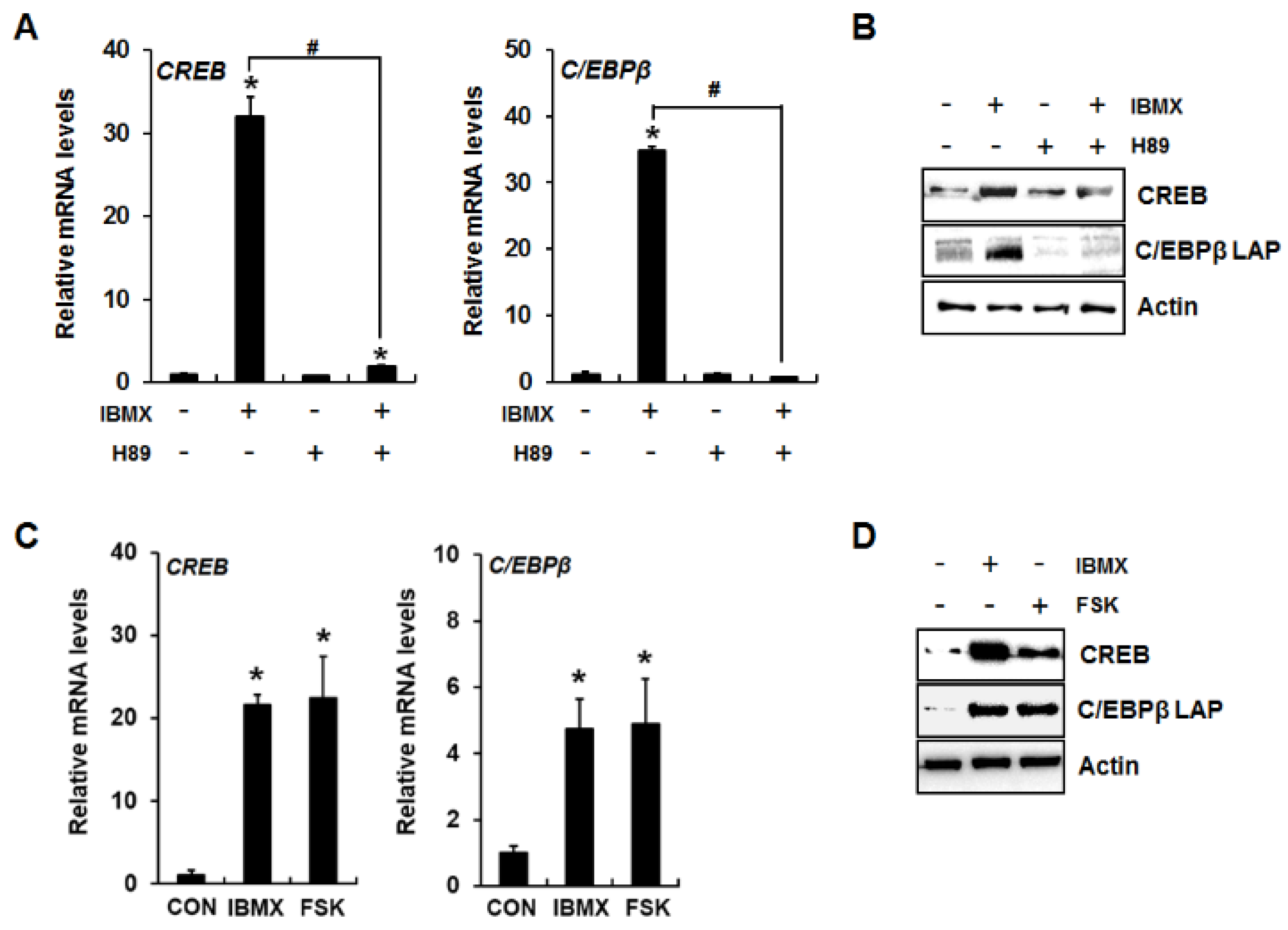
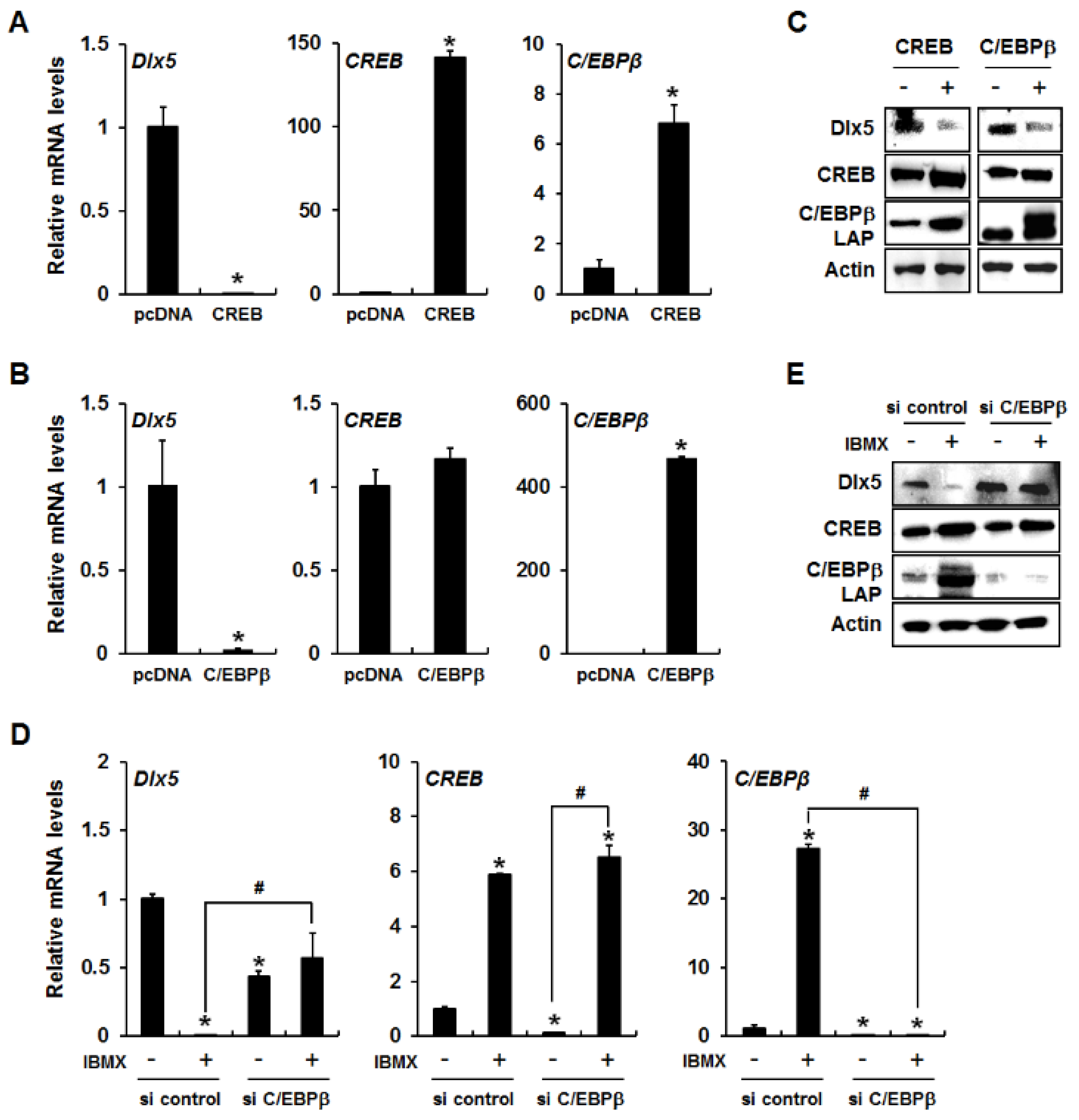
2.2. CREB Mediates cAMP/PKA Signaling-Induced Dlx5 Suppression via Upregulation of C/EBPβ
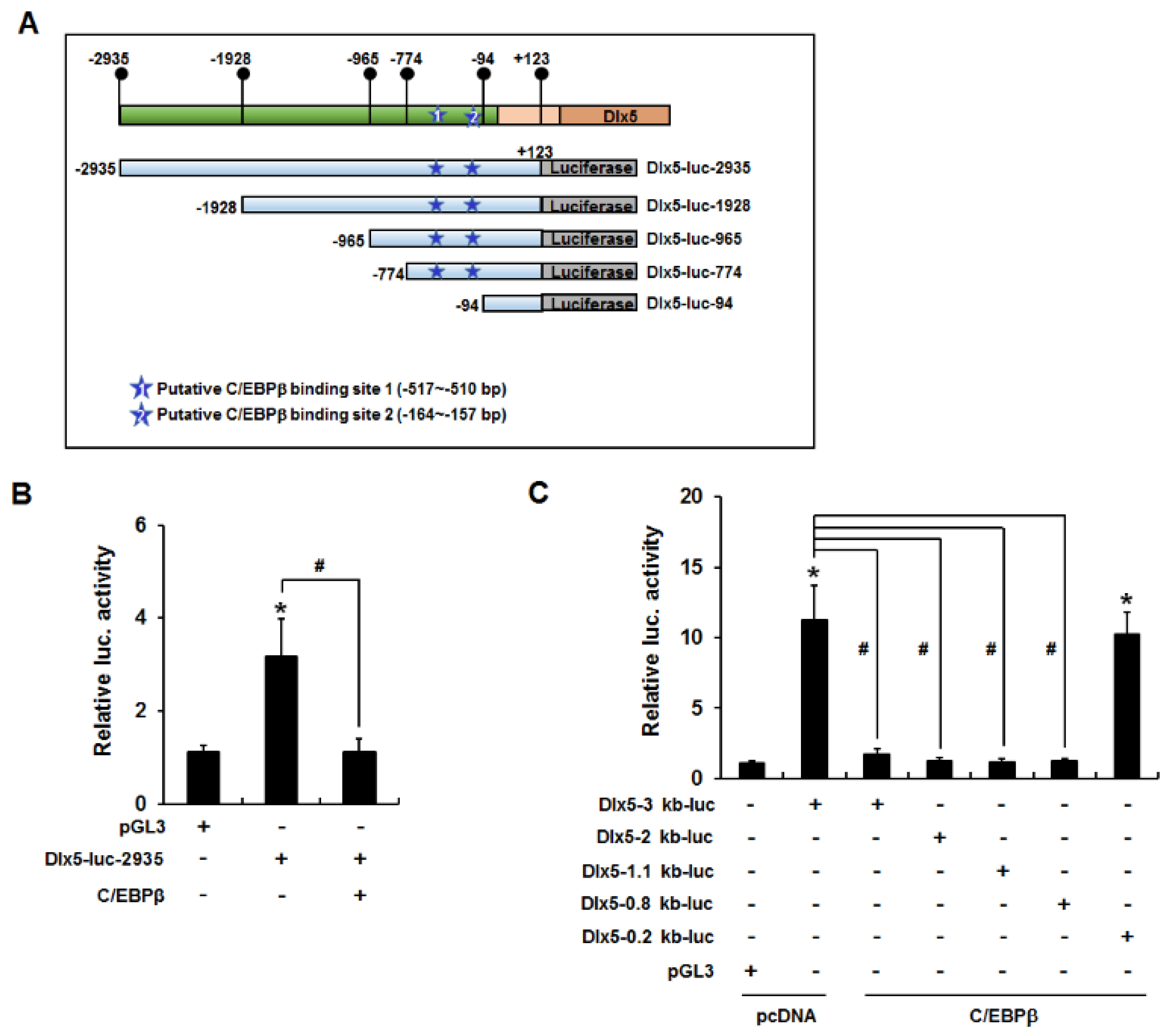
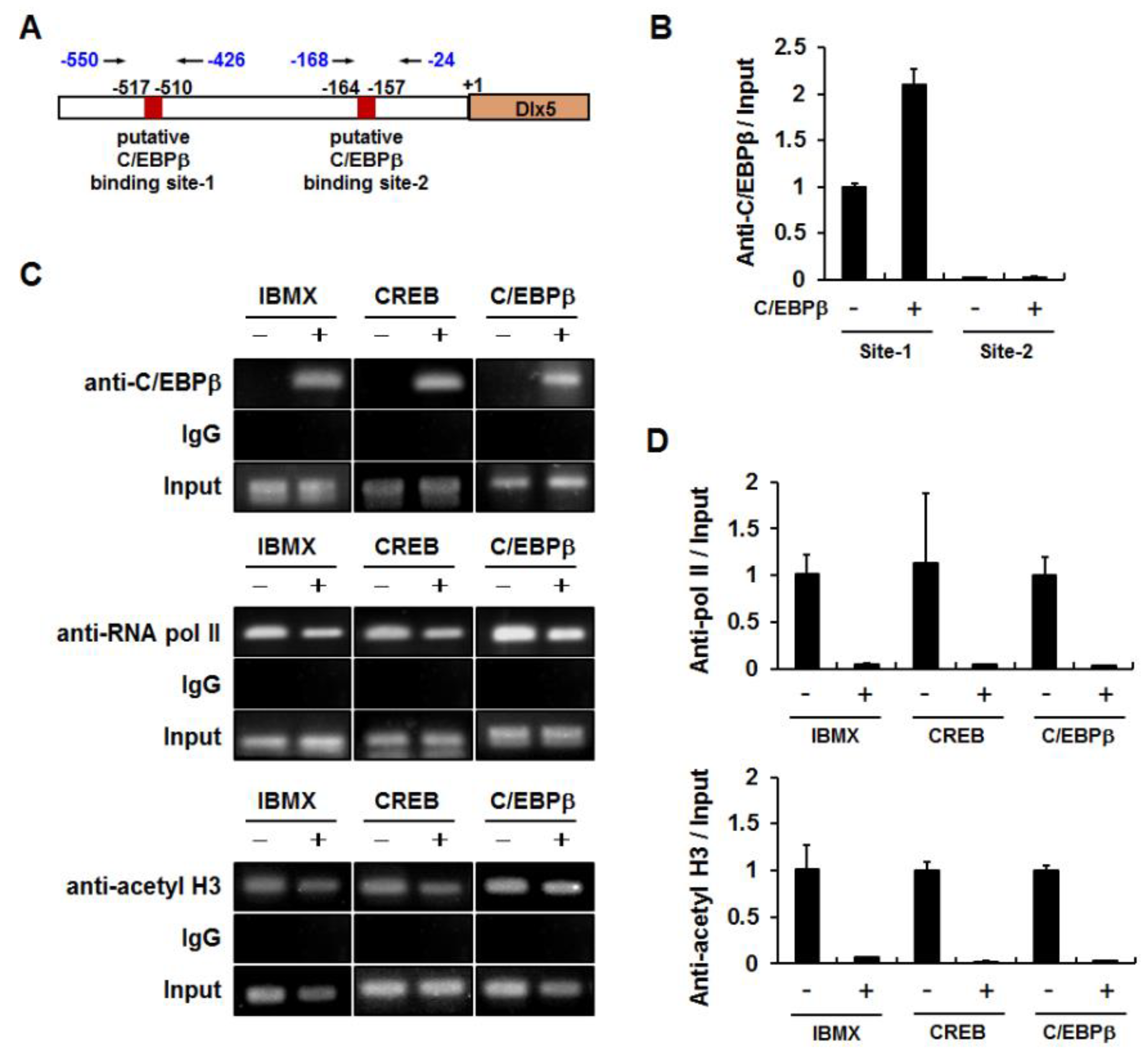
2.3. C/EBPβ Binds to the Dlx5 Promoter and Suppresses Dlx5 Transcription
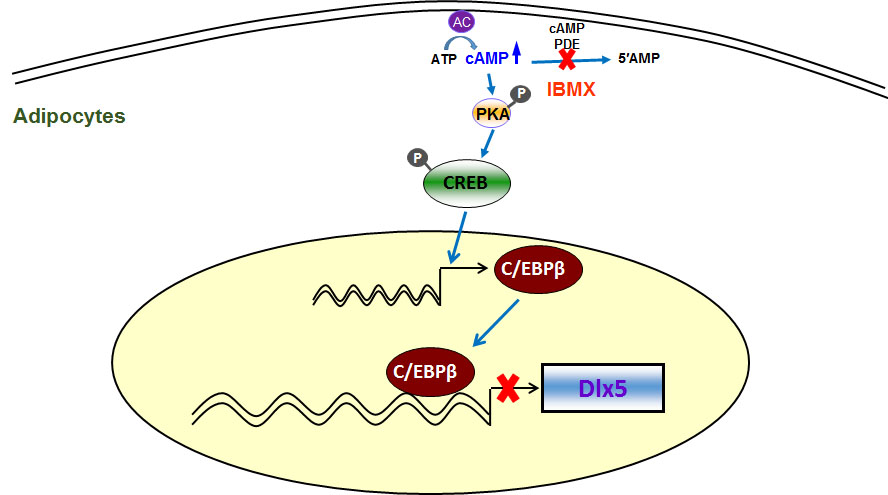
3. Discussion
4. Materials and Methods
4.1. Materials and Cell Culture
4.2. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
4.3. Western Blot Analysis
4.4. C/EBPβ Knockdown by Small Interfering RNA (siRNA)
4.5. Luciferase Reporter Assays
4.6. Chromatin Immunoprecipitation (ChIP) Assays
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Giri, S.; Rattan, R.; Haq, E.; Khan, M.; Yasmin, R.; Won, J.S.; Key, L.; Singh, A.K.; Singh, I. Aicar inhibits adipocyte differentiation in 3t3l1 and restores metabolic alterations in diet-induced obesity mice model. Nutr. Metab. 2006, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Kim, Y.J.; Yoon, W.J.; Kim, J.I.; Kim, B.G.; Hwang, Y.S.; Wozney, J.M.; Chi, X.Z.; Bae, S.C.; Choi, K.Y.; et al. Dlx5 specifically regulates runx2 type ii expression by binding to homeodomain-response elements in the runx2 distal promoter. J. Biol. Chem. 2005, 280, 35579–35587. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Kwon, T.G.; Park, H.S.; Wozney, J.M.; Ryoo, H.M. Bmp-2-induced osterix expression is mediated by Dlx5 but is independent of runx2. Biochem. Biophys. Res. Commun. 2003, 309, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.L.; Woo, K.M.; Ryoo, H.M.; Baek, J.H. Distal-less homeobox 5 inhibits adipogenic differentiation through the down-regulation of peroxisome proliferator-activated receptor gamma expression. J. Cell. Physiol. 2013, 228, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Qadir, A.S.; Woo, K.M.; Ryoo, H.M.; Baek, J.H. Insulin suppresses distal-less homeobox 5 expression through the up-regulation of microrna-124 in 3t3-l1 cells. Exp. Cell Res. 2013, 319, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Mackall, J.C.; Student, A.K.; Polakis, S.E.; Lane, M.D. Induction of lipogenesis during differentiation in a “preadipocyte” cell line. J. Biol. Chem. 1976, 251, 6462–6464. [Google Scholar] [PubMed]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Ito, H.; Murakami, M.; Yoshida, K.; Tagawa, Y.; Hagiwara, K.; Takagi, A.; Kojima, T.; Suzuki, M.; Banno, Y.; et al. Mechanism of increased pld1 gene expression during early adipocyte differentiation process of mouse cell line 3t3-l1. J. Cell. Biochem. 2010, 109, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Klemm, D.J.; Vinson, C.; Lane, M.D. Role of creb in transcriptional regulation of ccaat/enhancer-binding protein beta gene during adipogenesis. J. Biol. Chem. 2004, 279, 4471–4478. [Google Scholar] [CrossRef] [PubMed]
- Reusch, J.E.; Colton, L.A.; Klemm, D.J. Creb activation induces adipogenesis in 3t3-l1 cells. Mol. Cell. Biol. 2000, 20, 1008–1020. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.B.; Kim, W.H.; Han, K.L.; Park, J.H.; Lee, J.; Yeo, J.; Jung, M.H. Camp-response element binding protein (creb) positively regulates mouse adiponectin gene expression in 3t3-l1 adipocytes. Biochem. Biophys. Res. Commun. 2010, 391, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Klemm, D.J.; Roesler, W.J.; Boras, T.; Colton, L.A.; Felder, K.; Reusch, J.E. Insulin stimulates camp-response element binding protein activity in hepg2 and 3t3-l1 cell lines. J. Biol. Chem. 1998, 273, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Yeh, W.C.; Cao, Z.; Classon, M.; McKnight, S.L. Cascade regulation of terminal adipocyte differentiation by three members of the c/ebp family of leucine zipper proteins. Genes Dev. 1995, 9, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Chen, Z.; Friedman, J. Transcriptional regulation of adipogenesis by klf4. Cell Metab. 2008, 7, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.K.; Madsen, L.; Pedersen, L.M.; Hallenborg, P.; Hagland, H.; Viste, K.; Doskeland, S.O.; Kristiansen, K. Cyclic amp (camp)-mediated stimulation of adipocyte differentiation requires the synergistic action of epac- and camp-dependent protein kinase-dependent processes. Mol. Cell. Biol. 2008, 28, 3804–3816. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.P.; Ha, J.M.; Yun, S.J.; Kim, E.K.; Chung, S.W.; Hong, K.W.; Kim, C.D.; Bae, S.S. Transcriptional activation of peroxisome proliferator-activated receptor-gamma requires activation of both protein kinase A and akt during adipocyte differentiation. Biochem. Biophys. Res. Commun. 2010, 399, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Martini, C.N.; Plaza, M.V.; Vila Mdel, C. Pka-dependent and independent camp signaling in 3t3-l1 fibroblasts differentiation. Mol. Cell. Endocrinol. 2009, 298, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Acampora, D.; Merlo, G.R.; Paleari, L.; Zerega, B.; Postiglione, M.P.; Mantero, S.; Bober, E.; Barbieri, O.; Simeone, A.; Levi, G. Craniofacial, vestibular and bone defects in mice lacking the distal-less-related gene dlx5. Development 1999, 126, 3795–3809. [Google Scholar] [PubMed]
- Robledo, R.F.; Rajan, L.; Li, X.; Lufkin, T. The D1x5 and D1x6 homeobox genes are essential for craniofacial, axial, and appendicular skeletal development. Genes Dev. 2002, 16, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.C.; Dusting, G.J.; Guo, N.; Peshavariya, H.M.; Taylor, C.J.; Dilley, R.; Narumiya, S.; Jiang, F. Prostacyclin receptor suppresses cardiac fibrosis: Role of creb phosphorylation. J. Mol. Cell. Cardiol. 2010, 49, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ozaki, N.; Yamada, T.; Miura, Y.; Oiso, Y. H-89 potentiates adipogenesis in 3t3-l1 cells by activating insulin signaling independently of protein kinase a. Life Sci. 2007, 80, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, D.; Zhou, Y.; Zhou, B.; Yang, Y.; Chen, H.; Song, J. Protein kinase a suppresses the differentiation of 3t3-l1 preadipocytes. Cell Res. 2008, 18, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Lane, M.D. Activation and centromeric localization of ccaat/enhancer-binding proteins during the mitotic clonal expansion of adipocyte differentiation. Genes Dev. 1999, 13, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Dentesano, G.; Straccia, M.; Ejarque-Ortiz, A.; Tusell, J.M.; Serratosa, J.; Saura, J.; Sola, C. Inhibition of cd200r1 expression by c/ebp beta in reactive microglial cells. J. Neuroinflam. 2012, 9, 165. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.Y.; Sun, G.; Jiang, L.; Shao, L.; Hu, Y.; Jiang, Y.; Wang, Y.; An, W. Epidermal growth factor down-regulates the expression of human hepatic stimulator substance via ccaat/enhancer-binding protein beta in hepg2 cells. Biochem. J. 2010, 431, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Hoeffler, J.P.; Meyer, T.E.; Yun, Y.; Jameson, J.L.; Habener, J.F. Cyclic amp-responsive DNA-binding protein: Structure based on a cloned placental cdna. Science 1988, 242, 1430–1433. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-L.; Qadir, A.S.; Park, H.-J.; Chung, E.; Lee, Y.-S.; Woo, K.M.; Ryoo, H.-M.; Kim, H.J.; Baek, J.-H. cAMP/Protein Kinase A Signaling Inhibits Dlx5 Expression via Activation of CREB and Subsequent C/EBPβ Induction in 3T3-L1 Preadipocytes. Int. J. Mol. Sci. 2018, 19, 3161. https://doi.org/10.3390/ijms19103161
Lee H-L, Qadir AS, Park H-J, Chung E, Lee Y-S, Woo KM, Ryoo H-M, Kim HJ, Baek J-H. cAMP/Protein Kinase A Signaling Inhibits Dlx5 Expression via Activation of CREB and Subsequent C/EBPβ Induction in 3T3-L1 Preadipocytes. International Journal of Molecular Sciences. 2018; 19(10):3161. https://doi.org/10.3390/ijms19103161
Chicago/Turabian StyleLee, Hye-Lim, Abdul S. Qadir, Hyun-Jung Park, Eunkyung Chung, Yun-Sil Lee, Kyung Mi Woo, Hyun-Mo Ryoo, Hyun Jeong Kim, and Jeong-Hwa Baek. 2018. "cAMP/Protein Kinase A Signaling Inhibits Dlx5 Expression via Activation of CREB and Subsequent C/EBPβ Induction in 3T3-L1 Preadipocytes" International Journal of Molecular Sciences 19, no. 10: 3161. https://doi.org/10.3390/ijms19103161
APA StyleLee, H.-L., Qadir, A. S., Park, H.-J., Chung, E., Lee, Y.-S., Woo, K. M., Ryoo, H.-M., Kim, H. J., & Baek, J.-H. (2018). cAMP/Protein Kinase A Signaling Inhibits Dlx5 Expression via Activation of CREB and Subsequent C/EBPβ Induction in 3T3-L1 Preadipocytes. International Journal of Molecular Sciences, 19(10), 3161. https://doi.org/10.3390/ijms19103161