Combination Therapy of PEG-HM-3 and Methotrexate Retards Adjuvant-Induced Arthritis

Abstract
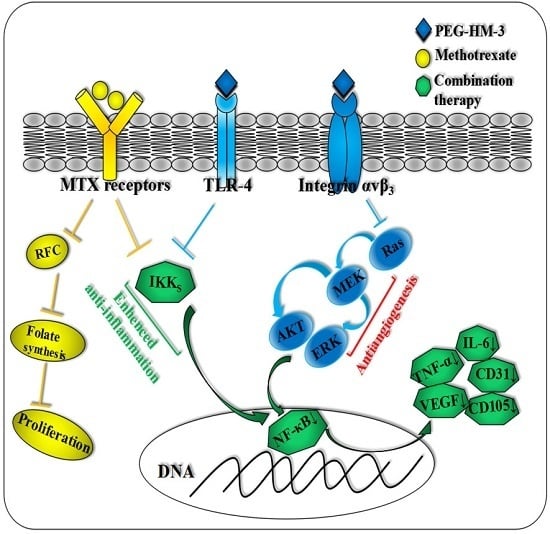
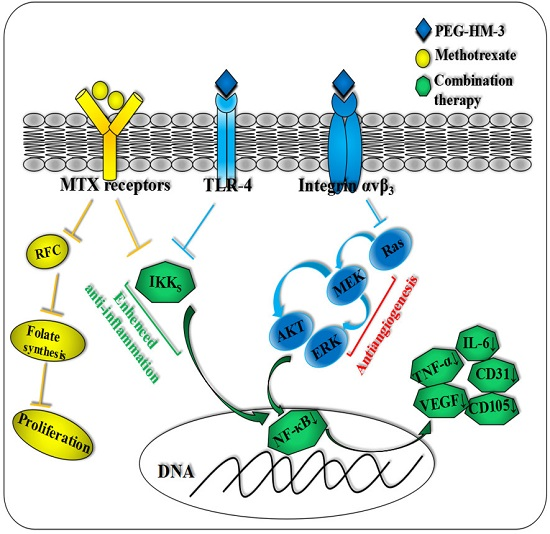
1. Introduction
2. Results
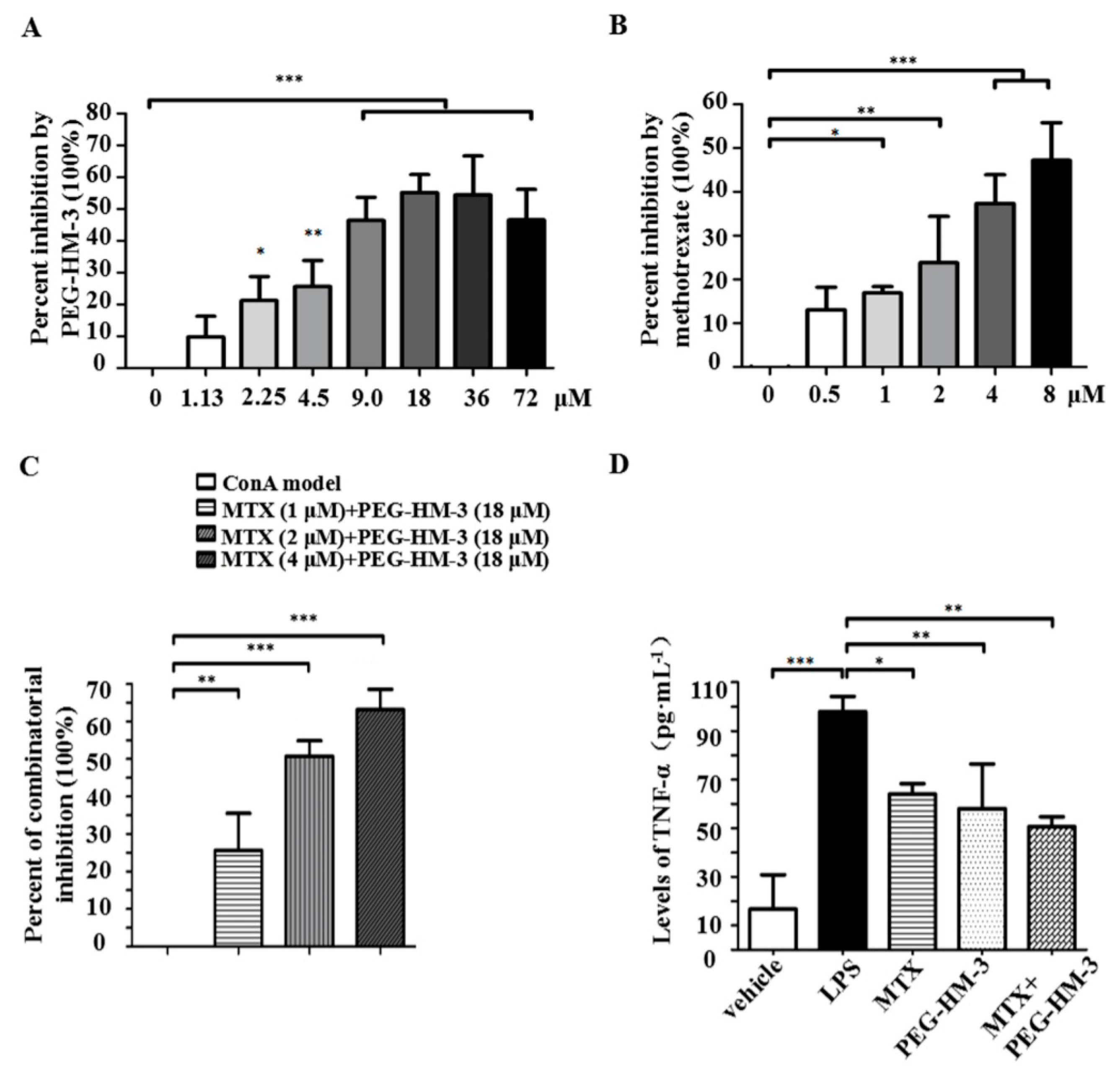
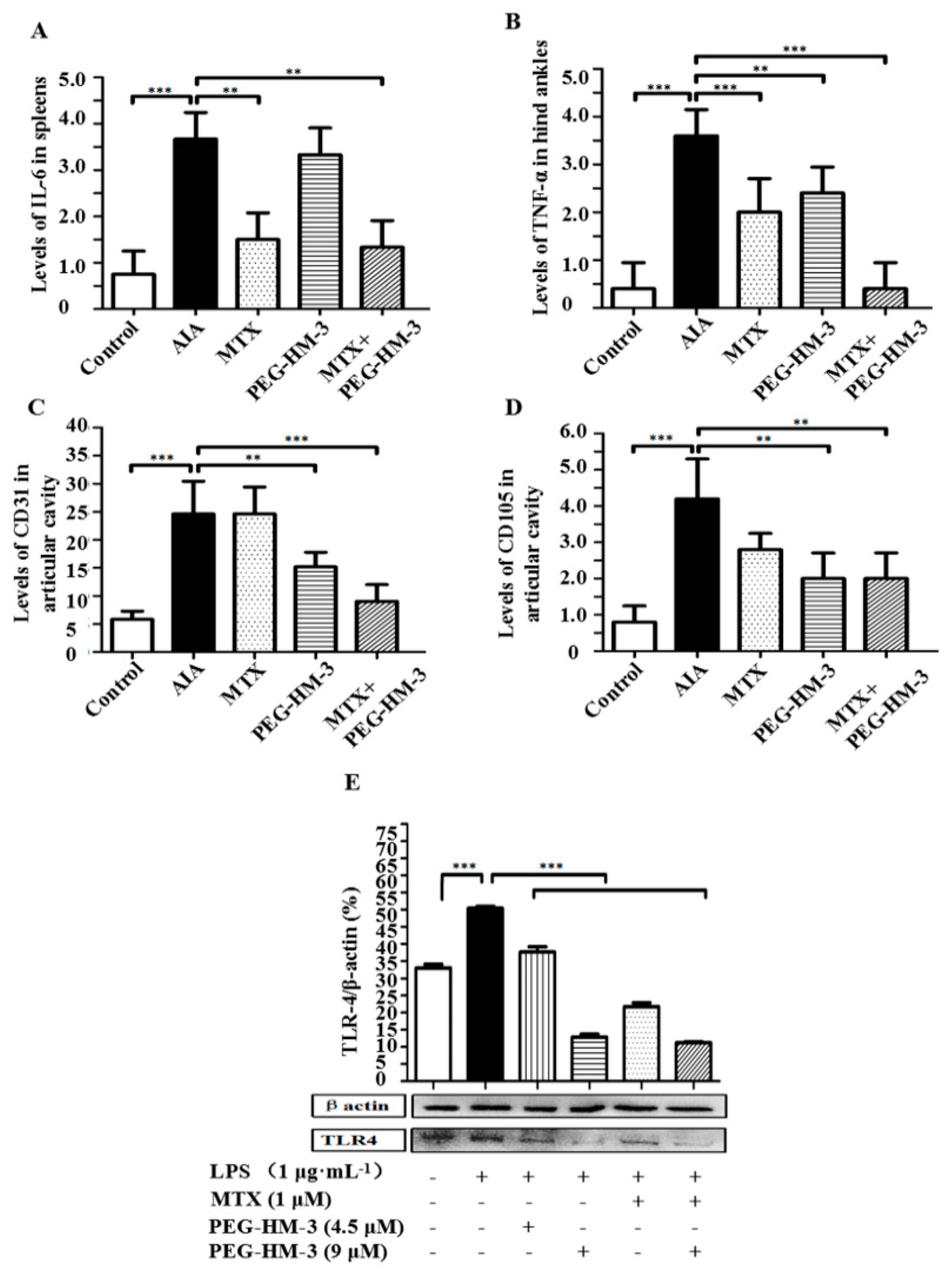
2.1. Lymphoproliferative Responses and Anti-Inflammation Activity
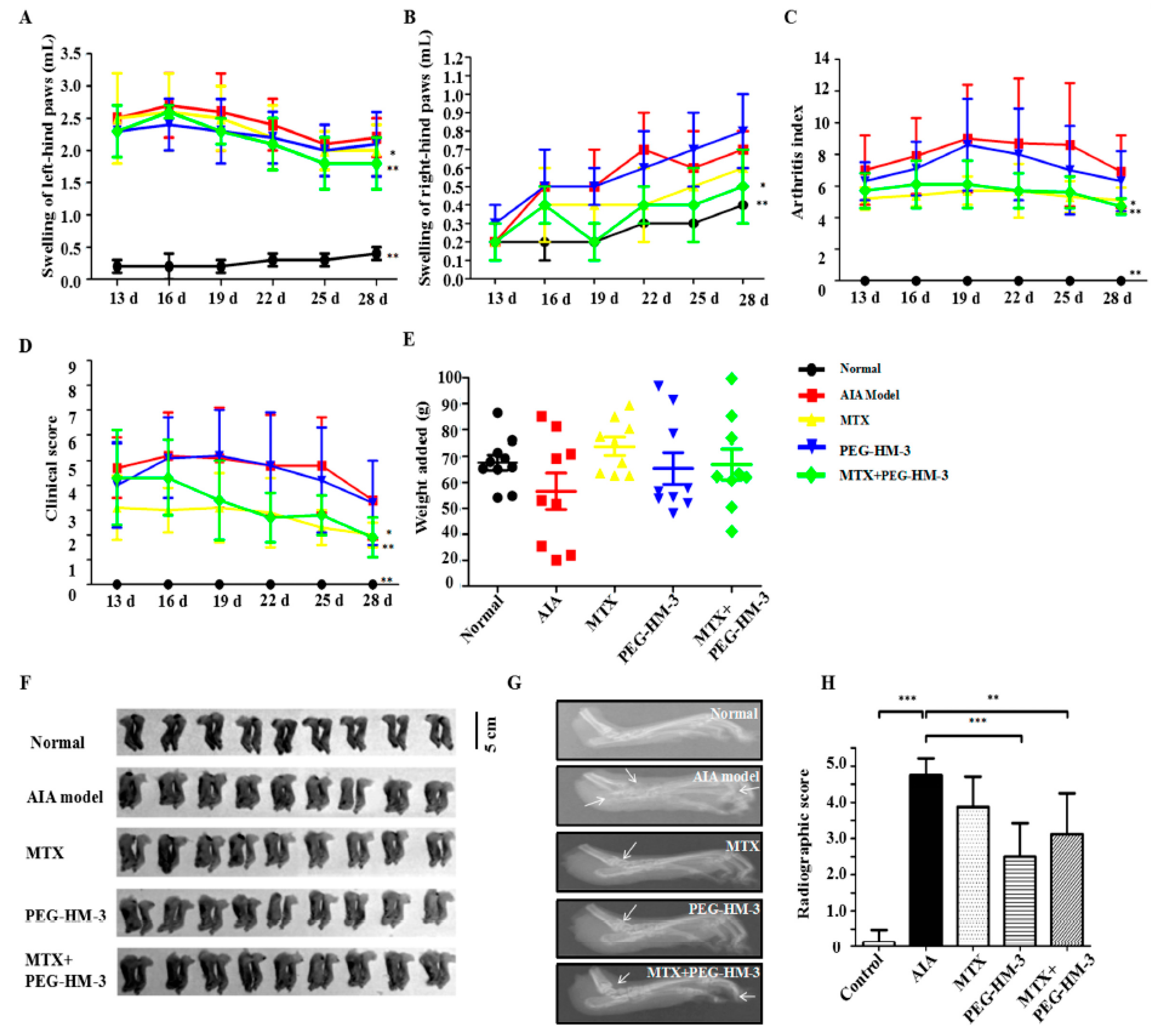
2.2. Efficacy in Adjuvant-Induced Arthritic Animals
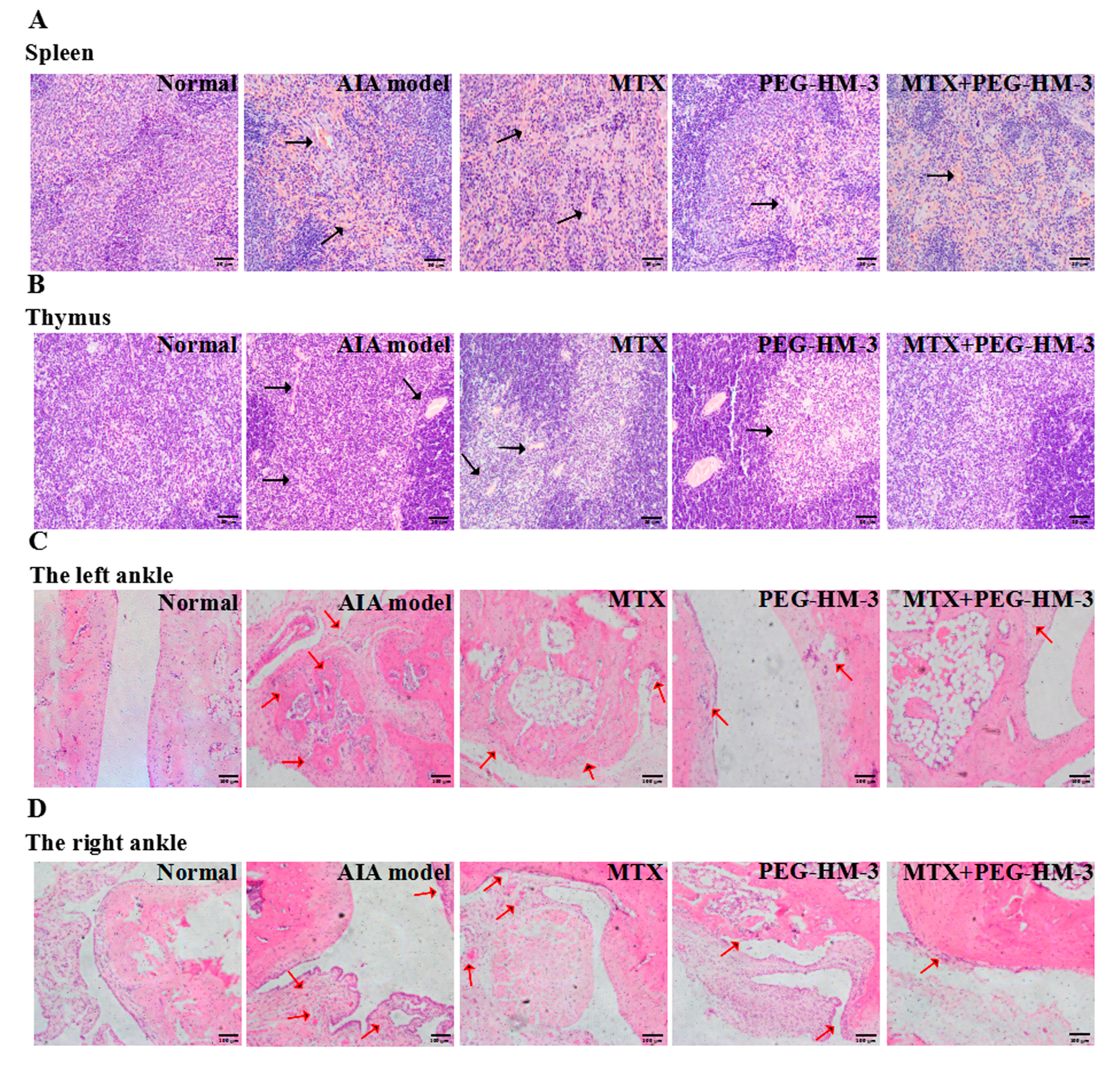
2.3. Histological Evaluation
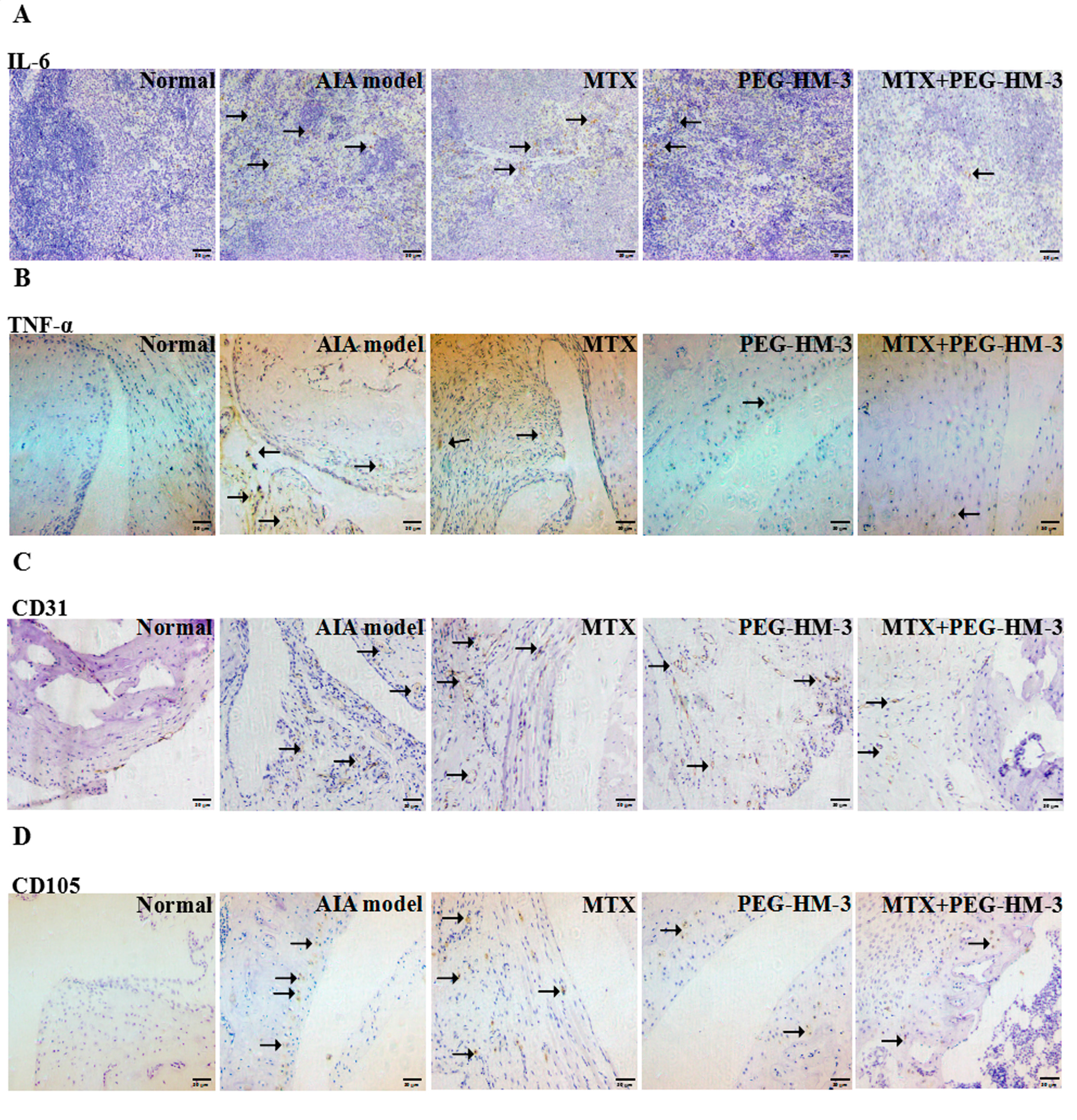
2.4. Immunohistochemical Analysis
2.5. Efficacy in the Expression of TLR-4 Protein
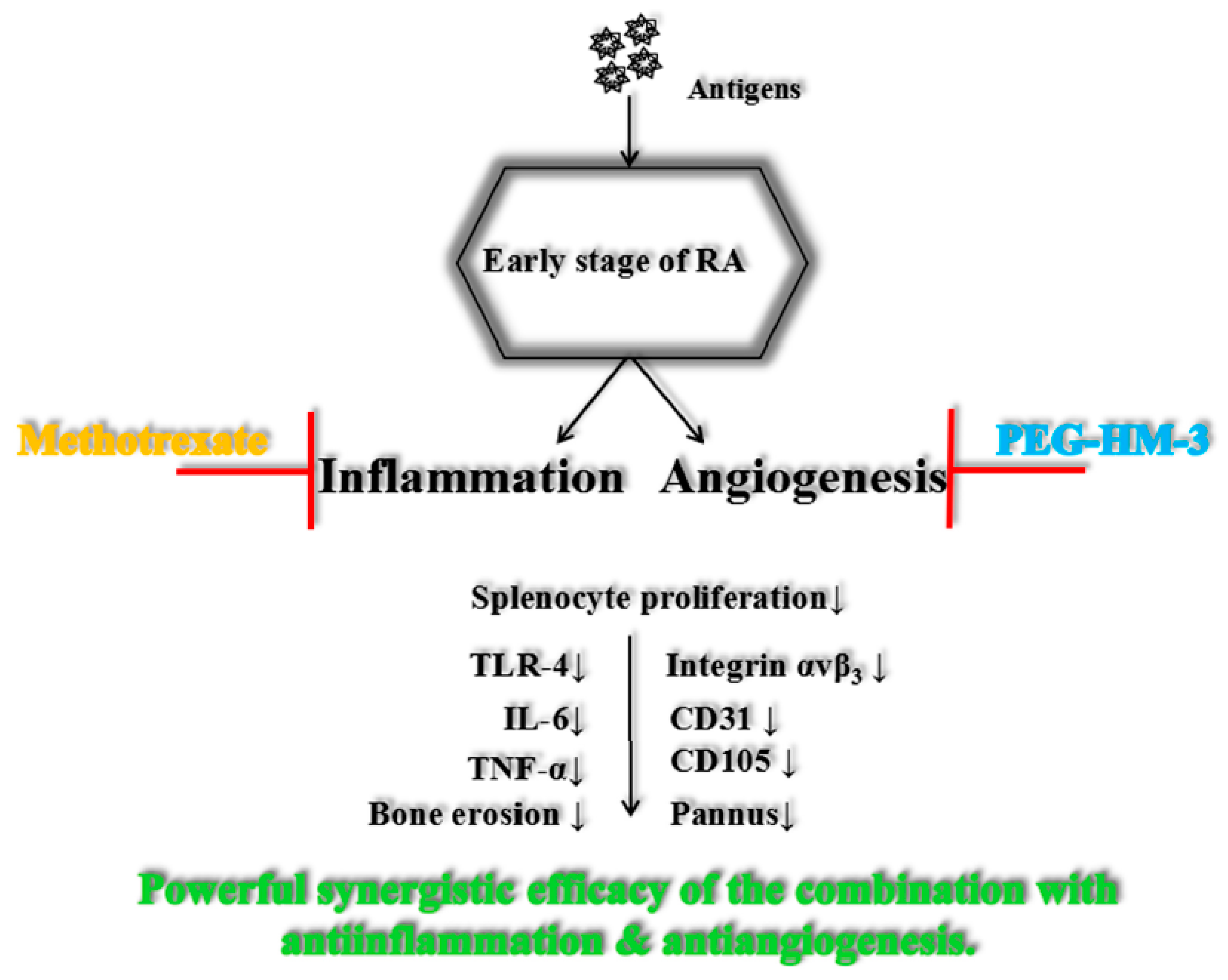
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Materials
4.3. Cell Culture and Stimulation Conditions
4.4. MTT and ELISA Analysis
4.5. Animals and Experiment Design
4.6. Assessment of Arthritis Symptom
4.7. Histopathological Analysis
4.8. Western Blot Assay of Protein Expressions
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AIA | antigen-induced arthritis |
| CD | Crohn’s disease |
| CD | cluster of differentiation |
| CFA | complete Freund’s adjuvant |
| CIA | type II collagen-induced arthritis |
| ConA | concanavalin |
| DMARDs | disease-modifying anti-rheumatic drugs |
| EDTA | ethylene diamine tetraacetic Acid |
| ELISA | enzyme linked immunosorbent assay |
| HPLC | high-performance liquid chromatography |
| IL | interleukin |
| LPS | lipopolysaccharide |
| MTT | methylthiazolyldiphenyl-tetrazolium bromide |
| MTX | methotrexate |
| mPEG-SC20k- | methoxy-polyethylene glycol-succinimidyl carbonate |
| PEG | polyethylene glycol |
| PEG-HM-3 | mPEG-SC20k-HM-3 |
| RA | rheumatoid arthritis |
| RGD | Arg-Gly-Asp |
| SD | Sprague Dawley |
| TLRs | toll-like receptors |
| TNF | tumor necrosis factor |
| VEGF | vascular endothelial cell growth factor |
References
- Smolen, J.S.; Aletaha, D. Rheumatoid arthritis therapy reappraisal: Strategies, opportunities and challenges. Nat. Rev. Rheumatol. 2015, 11, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Zoltan, S.; Timea, B.; Gyorgy, P.; Alisa, E.K. Angiogenesis in rheumatoid arthritis. Autoimmunity 2009, 42, 563–573. [Google Scholar]
- Colman, R.J.; Rubin, D.T. Optimal Doses of Methotrexate Combined with Anti-TNF Therapy to Maintain Clinical Remission in Inflammatory Bowel Disease. J. Crohn’s Colitis 2015, 9, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Meier, F.M.; Frerix, M.; Hermann, W.; Müllerladner, U. Current immunotherapy in rheumatoid arthritis. Immunotherapy 2013, 5, 955–974. [Google Scholar] [CrossRef] [PubMed]
- Gabay, C.; Hasler, P.; Kyburz, D.; So, A.; Villiger, P.; von Kempis, J.; Walker, U. Biological agents in monotherapy for the treatment of rheumatoid arthritis. Swiss Med. Wkly. 2014, 144. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ren, Y.; Li, P.; Xu, H.M. In vivo anti-tumor activity of polypeptide HM-3 modified by different polyethylene glycols (PEG). Int. J. Mol. Sci. 2011, 12, 2650–2663. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.M.; Pratt, A.G.; Isaacs, J.D. Mechanism of action of methotrexate in rheumatoid arthritis, and the search for biomarkers. Nat. Rev. Rheumatol. 2016, 12, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Szekanecz, Z.; Besenyei, T.; Szentpétery, A.; Koch, A.E. Angiogenesis and vasculogenesis in rheumatoid arthritis. Curr. Opin. Rheumatol. 2010, 22, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Bolon, B.; Campagnuolo, G.; Feige, U. Duration of bone protection by a single osteoprotegerin injection in rats with adjuvant-induced arthritis. Cell. Mol. Life Sci. 2002, 59, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Avraamides, C.J.; Garmysusini, B.; Varner, J.A. Integrins in angiogenesis and lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.M.; Pritchard, J.M.; Macdonald, S.J.; Jamieson, C.; Watson, A.J. The emergence of small molecule non-RGD-mimetic inhibitors for RGD integrins. J. Med. Chem. 2017, 60, 3241–3251. [Google Scholar] [CrossRef] [PubMed]
- Polman, C.H.; O’Connor, P.W.; Havrdova, E.; Hutchinson, M.; Kappos, L. A randomized, placebo-controlled trial of natalizumab for relapsing multiple sclerosis. N. Engl. J. Med. 2006, 354, 899. [Google Scholar] [CrossRef] [PubMed]
- Targan, S.R.; Feagan, B.G.; Fedorak, R.N.; Lashner, B.A.; Panaccione, R. Natalizumab for the treatment of active Crohn’s disease: Results of the ENCORE trial. Gastroenterology 2007, 132, 1672–1683. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, B.; Kappos, L. Natalizumab: Targeting α4-integrins in multiple sclerosis. Neurodegener. Dis. 2008, 5, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Sandborn, W.J.; Yednock, T.A. Novel approaches to treating inflammatory bowel disease: Targeting α-4 integrin. Am. J. Gastroenterol. 2003, 98, 2372–2382. [Google Scholar] [CrossRef] [PubMed]
- Müllerladner, U.; Elices, M.J.; Kriegsmann, J.B.; Strahl, D.; Gay, R.E.; Firestein, G.S.; Gay, S. Alternatively spliced CS-1 fibronectin isoform and its receptor VLA-4 in rheumatoid arthritis synovium. J. Rheumatol. 1997, 24, 1873–1880. [Google Scholar]
- Beniwal-Patel, P.; Saha, S. The role of integrin antagonists in the treatment of inflammatory bowel disease. Expert Opin. Biol. Ther. 2014, 14, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.Y. Alpha 4 integrin antagonists. J. Curr. Pharm. Design 2002, 8, 1229. [Google Scholar] [CrossRef]
- Xu, Y.Z.; Smith, J.L.; Semko, C.M.; Rossiter, K.I.; Fukuda, J.Y.; Dappen, M.S.; Quincy, D.A.; Konradi, A.W.; Mao, W.; Welch, B.; et al. Orally available and efficacious α4β1/α4β7 integrin inhibitors. Bioorg. Med. Chem. Lett. 2013, 23, 4370–4373. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Le, T.D.; Rodan, S.B.; Rodan, G.A. Involvement of αvβ3, integrins in osteoclast function. J. Bone Miner. Metab. 2007, 25, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.M.; Yin, R.; Chen, L.; Siraj, S.; Huang, X.; Wang, M.; Fang, H.; Wang, Y. An RGD-Modified Endostatin-Derived Synthetic Peptide Shows Antitumor Activity in Vivo. Bioconjug. Chem. 2008, 19, 1980–1986. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, J.; Wang, Y.; Xu, H.M. An integrin αvβ3 antagonistic modified peptide inhibits tumor growth through inhibition of the ERK and AKT signaling pathways. Oncol. Rep. 2016, 36, 1953. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Han, H.B.; Hu, J.L.; Zhang, X.J.; Wang, J.J.; Wang, W.J.; Nie, C.H.; Xu, H.M. PEGylated HM-3 presents anti-rheumatic bioactivity by inhibiting angiogenesis and inflammation. J. Mater. Chem. B 2014, 2, 800–813. [Google Scholar]
- Pincus, T.; Cronstein, B.; Braun, J. Methotrexate—The anchor drug—An introduction. Clin. Exp. Rheumatol. 2010, 28, S1–S2. [Google Scholar] [PubMed]
- Salliot, C. Long-term safety of methotrexate monotherapy in patients with rheumatoid arthritis: A systematic literature research. Ann. Rheum. Dis. 2009, 68, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Jacques, P.; Van den, B.F. Emerging therapies for rheumatoid arthritis. Expert Opin. Emerg. Drugs 2013, 18, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Goff, B.L.; Soltner, E.; Charrier, C. A combination of methotrexate and zoledronic acid prevents bone erosions and systemic bone mass loss in collagen induced arthritis. Arthritis Res. Ther. 2009, 11, R185. [Google Scholar] [CrossRef] [PubMed]
- López Mantecón, A.M.; Garrido, G. Combination of Mangifera indica L. Extract Supplementation Plus Methotrexate in Rheumatoid Arthritis Patients: A Pilot Study. Phytother. Res. 2014, 28, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, G.; Curran, G.; Jenkins, R. Et-52simultaneous brain delivery of a three anti-cancer drug combination via a peptide carrier-mediated intravenous injection of the three-drug cocktail. J. Neuro Oncol. 2014, 16. [Google Scholar] [CrossRef]
- Conaghan, P.G.; Emery, P.; Østergaard, M.; Keystone, E.C.; Genovese, M.C.; Hsia, E.C.; Xu, W.C.; Rahman, M.U. Assessment by MRI of inflammation and damage in rheumatoid arthritis patients with methotrexate inadequate response receiving golimumab: Results of the GO-FORWARD trial. Ann. Rheum. Dis. 2011, 70, 1968–1974. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Z.; Wang, W.; Ma, J.; Wang, S.; Su, Z.; Xu, H. Notch Signaling Mediates TNF-α-Induced IL-6 Production in Cultured Fibroblast-Like Synoviocytes from Rheumatoid Arthritis. Clin. Dev. Immunol. 2012, 2012, 350209. [Google Scholar] [CrossRef] [PubMed]
- Nishina, N.; Kaneko, Y.; Kameda, H.; Kuwana, M.; Takeuchi, T. Reduction of plasma IL-6 but not TNF-α by methotrexate in patients with early rheumatoid arthritis: A potential biomarker for radiographic progression. Clin. Rheumatol. 2013, 32, 1661–1666. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.W.; Lee, N.R.; Pi, R.H.; Lim, Y.S.; Lee, Y.M.; Lee, J.M.; Jeong, H.S.; Chung, S.H. IL-6 inhibitors for treatment of rheumatoid arthritis: Past, present, and future. Arch. Pharm. Res. 2015, 38, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, K.; Hirano, T. IL-6 in autoimmune disease and chronic inflammatory proliferative disease. Cytokine Growth Factor Rev. 2002, 13, 357–368. [Google Scholar] [CrossRef]
- Hashizume, M.; Hayakawa, N.; Suzuki, M.; Mihara, M. IL-6/sIL-6R trans-signalling, but not TNF-α induced angiogenesis in a HUVEC and synovial cell co-culture system. Rheumatol. Int. 2009, 29, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Catar, R.; Witowski, J.; Zhu, N.; Lücht, C.; Derrac, S.A.; Uceda, F.J.; Chen, L.; Jones, S.A.; Fielding, C.A.; Rudolf, A.; et al. IL-6 trans-signaling links inflammation with angiogenesis in the peritoneal membrane. J. Am. Soc. Nephrol. 2016, 28, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Straub, R.H. Integrins and their ligands in rheumatoid arthritis. Arthritis Res. Ther. 2011, 13, 244. [Google Scholar] [CrossRef] [PubMed]
- Konisti, S.; Kiriakidis, S.; Paleolog, E.M. Angiogenesis in Rheumatoid Arthritis. Angiogenesis and Vascularisation; Springer: Vienna, Austria, 2013; pp. 339–365. [Google Scholar]
- Asano, T.; Iwasaki, N.; Kon, S.; Kanayama, M.; Morimoto, J.; Minami, A.; Uede, T. α9β1 integrin acts as a critical intrinsic regulator of human rheumatoid arthritis. Rheumatology 2014, 53, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Zeltz, C.; Gullberg, D. Post-translational modifications of integrin ligands as pathogenic mechanisms in disease. Matrix Biol. 2014, 40, 5. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.A.; Wendholt, D.; Strietholt, S.; Frank, S.; Pundt, N.; Korb-Pap, A.; Joosten, L.A.; van den Berg, W.B.; Kollias, G.; Eckes, B.; et al. The loss of α2β1 integrin suppresses joint inflammation and cartilage destruction in mouse models of rheumatoid arthritis. Arthritis Rheum. 2012, 64, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.L.; Moots, R.J.; Bucknall, R.C.; Edwards, S.W. Neutrophil function in inflammation and inflammatory diseases. Rheumatology 2010, 49, 1618–1631. [Google Scholar] [CrossRef] [PubMed]
- Mackay, C.R. Moving targets: Cell migration inhibitors as new anti-inflammatory therapies. Nat. Immunol. 2008, 9, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Olivera, A.; Moore, T.W.; Hu, F.; Brown, A.P.; Sun, A.; Liotta, D.C.; Snyder, J.P.; Yoon, Y.; Shim, H.; Marcus, A.I. Inhibition of the NF-κB signaling pathway by the curcumin analog, 3,5-Bis(2-pyridinylmethylidene)-4-piperidone (EF31): Anti-inflammatory and anti-cancer properties. Int. Immunopharmacol. 2012, 12, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Criswell, L.A. Gene discovery in rheumatoid arthritis highlights the CD40/NF-κB signaling pathway in disease pathogenesis. Immunol. Rev. 2010, 233, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, S.; Akashi, S.; Yamada, T.; Tanimura, N.; Kobayashi, M.; Konno, K.; Matsumoto, F.; Fukase, K.; Kusumoto, S.; Nagai, Y. Lipid A antagonist, lipid IVa, is distinct from lipid A in interaction with Toll-like receptor 4 (TLR4)-MD-2 and ligand-induced TLR4 oligomerization. Int. Immunol. 2004, 16, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Mueller, B.U.; Jacobsen, F.; Butler, K.M.; Husson, R.N.; Lewis, L.L.; Pizzo, P.A. Combination treatment with azidothymidine and granulocyte colony-stimulating factor in children with human immunodeficiency virus infection. J. Pediatr. 1992, 121, 797–802. [Google Scholar] [CrossRef]
- Huang, Q.; Ma, Y.; Adebayo, A.; Pope, P.M. Increased macrophage activation mediated through toll-like receptors in rheumatoid arthritis. Arthritis Rheumatol. 2007, 56, 2192–2201. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, M.; Yamamura, M.; Aita, T.; Okamoto, A.; Ueno, A.; Oqawa, N.; Akashi, S.; Miyake, K.; Godowski, P.J.; Makino, H. Expression of toll-like receptor 2 on CD16+ blood monocytes and synovial tissue macrophages in rheumatoid arthritis. Arthritis Rheumatol. 2004, 50, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Guangquan, J.; Renqiong, C.; Jianxian, Z. Atractylenolide I inhibits lipopolysaccharide-induced inflammatory responses via mitogen-activated protein kinase pathways in RAW264.7 cells. Immunopharmacol. Immunotoxicol. 2014, 36, 420–425. [Google Scholar]
- Burnham, D.K.; Lackey, A.; Manering, M.; Jaensson, E.; Pearson, J.; Tyler, D.O.; Melson, D.; Talent, L.G. Effects of 17α-ethinylestradiol on immune parameters in the lizard Sceloporus occidentalis. Environ. Toxicol. 2003, 18, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Vernonroberts, B.; Liyanage, S.P.; Currey, H.L.F. Adjuvant arthritis in the rat. Curr. Protoc. Immunol. 2001, 15, 389–397. [Google Scholar]
- Apparailly, F.; Verwaerde, C.; Jacquet, C.; Auriault, C.; Sany, J.; Jorgensen, C. Adenovirus-mediated transfer of viral IL-10 gene inhibits murine collagen-induced arthritis. J. Immunol. 1998, 160, 5213–5220. [Google Scholar] [PubMed]
- Huang, B.; Wang, Q.T.; Song, S.S.; Wu, Y.J.; Ma, Y.K.; Zhang, L.L.; Chen, J.Y.; Wu, H.X.; Jiang, L.; Wei, W. Combined use of etanercept and MTX restores CD4⁺/CD8⁺ ratio and Tregs in spleen and thymus in collagen-induced arthritis. Inflamm. Res. 2012, 61, 1229. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W.S. Image J, US National Institutes of Health, Bethesda, MD, USA. Available online: http://www.rsb.info.nih.gov/ij/ (accessed on 8 June 2017).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Left Hind Paws | Normal | AIA Model | MTX | PEG-HM-3 | MTX + PEG-HM-3 |
|---|---|---|---|---|---|
| Synovitis | 0.00 ± 0.00 *** | 3.00 ± 0.82 | 1.33 ± 0.58 ** | 1.33 ± 0.58 ** | 1.00 ± 0.00 *** |
| Pannus | 0.00 ± 0.00 *** | 3.17 ± 0.29 | 2.00 ± 0.82 | 1.67 ± 0.58 * | 1.33 ± 0.58 ** |
| Inflammation | 0.00 ± 0.00 *** | 2.67 ± 0.58 | 1.33 ± 0.58 * | 1.33 ± 0.58 * | 1.00 ± 0.00 ** |
| Bone erosion | 0.00 ± 0.00 *** | 4.33 ± 0.58 | 1.33 ± 0.58 *** | 1.33 ± 0.58 *** | 1.00 ± 0.00 ** |
| Right Hind Paws | Normal | AIA Model | MTX | PEG-HM-3 | MTX + PEG-HM-3 |
| Synovitis | 0.00 ± 0.00 *** | 3.33 ± 0.58 | 1.67 ± 0.58 ** | 1.33 ± 0.58 *** | 1.00 ± 0.00 *** |
| Pannus | 0.00 ± 0.00 *** | 3.67 ± 0.58 | 1.67 ± 0.58 ** | 1.17 ± 0.29 *** | 1.00 ± 0.00 *** |
| Inflammation | 0.00 ± 0.00 *** | 2.67 ± 0.58 | 1.00 ± 0.00 *** | 1.33 ± 0.58 ** | 1.00 ± 0.00 *** |
| Bone erosion | 0.00 ± 0.00 *** | 3.33 ± 0.58 | 1.67 ± 0.58 ** | 1.33 ± 0.58 *** | 1.33 ± 0.58 *** |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, J.; Wu, X.; Setrerrahmane, S.; Qian, K.; Hou, Y.; Yu, L.; Lin, C.; Wu, Q.; Xu, H. Combination Therapy of PEG-HM-3 and Methotrexate Retards Adjuvant-Induced Arthritis. Int. J. Mol. Sci. 2017, 18, 1538. https://doi.org/10.3390/ijms18071538
Hao J, Wu X, Setrerrahmane S, Qian K, Hou Y, Yu L, Lin C, Wu Q, Xu H. Combination Therapy of PEG-HM-3 and Methotrexate Retards Adjuvant-Induced Arthritis. International Journal of Molecular Sciences. 2017; 18(7):1538. https://doi.org/10.3390/ijms18071538
Chicago/Turabian StyleHao, Jingchao, Xiaodong Wu, Sarra Setrerrahmane, Kun Qian, Yueying Hou, Liting Yu, Chenyu Lin, Qianqian Wu, and Hanmei Xu. 2017. "Combination Therapy of PEG-HM-3 and Methotrexate Retards Adjuvant-Induced Arthritis" International Journal of Molecular Sciences 18, no. 7: 1538. https://doi.org/10.3390/ijms18071538
APA StyleHao, J., Wu, X., Setrerrahmane, S., Qian, K., Hou, Y., Yu, L., Lin, C., Wu, Q., & Xu, H. (2017). Combination Therapy of PEG-HM-3 and Methotrexate Retards Adjuvant-Induced Arthritis. International Journal of Molecular Sciences, 18(7), 1538. https://doi.org/10.3390/ijms18071538