Fortifying Horticultural Crops with Essential Amino Acids: A Review

Abstract
:
1. Introduction
2. Metabolism of Essential Amino Acids in Plants
3. Fortifying Horticultural Crops with the Essential Amino Acids Lys, Met, Thr, and Trp to Improve Their Nutritional Quality
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| T | Transgene |
| WT | Wild type |
| SS | Seed-specific promoter |
| Sup | Suppression the gene by antisense or RNAi |
| M | Mutant |
| NR | Not reported |
| NS | Not significant |
| SSP | Seed storage protein |
| AtCGS | ARABIDOPSIS thaliana cystathionine γ-synthase |
| AtD-CGS | Feedback insensitive form of AtCGS |
| EcSAT | E. coli serine acetyl transferase |
| SAMS | S-adenosyl Met synthase |
| MGL | Met γ-lyase |
| TS | Thr synthase |
| 2S SSA | 2S sunflower seed albumin |
| 2S BN | 2S brazil nut albumin |
| bAK | Bacterial feedback-insensitive Asp kinase |
| bDHDPS | Bacterial feedback-insensitive dihydrodipicolinate synthase |
| LKR | Lys-ketoglutarate reductase |
| Bip | Lys-rich binding protein |
| sb401 | Lys-rich pollen-specific protein |
| RLRH | Rice Lys-rich histone proteins |
| OASA1D | Rice mutated feedback-resistant a subunit of rice anthranilate synthase |
| NtASA2 | Tobacco feedback-insensitive anthranilate synthase 2 |
References
- Grunert, K.G.; Jeppesen, L.F.; Jespersen, K.R.; Sonne, A.M.; Hansen, K.; Trondsen, T.; Young, J.A. Market orientation of value chains: A conceptual framework based on four case studies from the food industry. Eur. J. Mark. 2005, 39, 428–455. [Google Scholar] [CrossRef]
- Galili, G.; Höfgen, R. Metabolic engineering of amino acids and storage proteins in plants. Metab. Eng. 2002, 4, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Amir, R. Fortifying plants with the essential amino acids lysine and methionine to improve nutritional quality. Plant Biotechnol. J. 2013, 11, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Karchi, H.; Shaul, O.; Perl, A.; Cahana, A.; Tzchori, I.B.T.; Galili, S. Production of transgenic plants containing elevated levels of lysine and threonine. Biochem. Soc. Trans. 1994, 22, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Mertz, E.T.; Bates, L.S.; Nelson, O.E. Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science 1964, 145, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Mourad, G.; King, J. LO-methylthreonine-resistant mutant of Arabidopsis defective in isoleucine feedback regulation. Plant Physiol. 1995, 107, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Halgand, F.; Wessel, P.M.; Laprévote, O.; Dumas, R. Biochemical and mass spectrometric evidence for quaternary structure modifications of plant threonine deaminase induced by isoleucine. Biochemistry 2002, 41, 13767–13773. [Google Scholar] [CrossRef] [PubMed]
- Binder, S. Branched-chain amino acid metabolism in Arabidopsis thaliana. Arabidopsis Book 2010, 8, e0137. [Google Scholar] [CrossRef] [PubMed]
- Razal, R.A.; Ellis, S.; Singh, S.; Lewis, N.G.; Towers, G.N. Nitrogen recycling in phenylpropanoid metabolism. Phytochemistry 1996, 41, 31–35. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Chapple, C. The phenylpropanoid pathway in Arabidopsis. Arabidopsis Book 2011, 9, e0152. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, P.F. Tetrahydropterin-dependent amino acid hydroxylases. Annu. Rev. Biochem. 1999, 68, 355–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Galili, G. Transgenic high-lysine rice—A realistic solution to malnutrition? J. Exp. Bot. 2016, 67, 4009–4011. [Google Scholar] [CrossRef] [PubMed]
- Shaul, O.; Galili, G. Concerted regulation of lysine and threonine synthesis in tobacco plants expressing bacterial feedback-insensitive aspartate kinase and dihydrodipicolinate synthase. Plant Mol. Biol. 1993, 23, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Tzchori, I.B.T.; Perl, A.; Galili, G. Lysine and threonine metabolism are subject to complex patterns of regulation in Arabidopsis. Plant Mol. Boil. 1996, 32, 727–734. [Google Scholar] [CrossRef]
- Zhu, X.; Galili, G. Increased lysine synthesis coupled with a knockout of its catabolism synergistically boosts lysine content and also transregulates the metabolism of other amino acids in Arabidopsis seeds. Plant Cell 2003, 15, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Galili, G. Lysine metabolism is concurrently regulated by synthesis and catabolism in both reproductive and vegetative tissues. Plant Physiol. 2004, 135, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Angelovici, R.; Fait, A.; Fernie, A.R.; Galili, G. A seed high-lysine trait is negatively associated with the TCA cycle and slows down Arabidopsis seed germination. New Phytol. 2011, 189, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Perl, A.; Shaul, O.; Galili, G. Regulation of lysine synthesis in transgenic potato plants expressing a bacterial dihydrodipicolinate synthase in their chloroplasts. Plant Mol. Biol. 1992, 19, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Falco, S.C.; Guida, T.; Locke, M.; Mauvais, J.; Sanders, C.; Ward, R.T.; Webber, P. Transgenic canola and soybean seeds with increased lysine. Nat. Biotechnol. 1995, 13, 577–582. [Google Scholar] [CrossRef]
- Hacham, Y.; Song, L.; Schuster, G.; Amir, R. Lysine enhances methionine content by modulating the expression of S-adenosylmethionine synthase. Plant J. 2007, 51, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Karchi, H.; Shaul, O.; Galili, G. Lysine synthesis and catabolism are coordinately regulated during tobacco seed development. Proc. Nat. Acad. Sci. USA 1994, 91, 2577–2581. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Pearson, K.W.; Meeker, G.; Staraci, L.C.; Sun, S.S. Enhancement of the methionine content of seed proteins by the expression of a chimeric gene encoding a methionine-rich protein in transgenic plants. Plant. Mol. Biol. 1989, 13, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Kuo, C.C.; Staraci, L.C.; Pearson, K.W.; Wainwright, C.; Georgescu, A.; Townsend, J. Accumulation of a Brazil nut albumin in seeds of transgenic canola results in enhanced levels of seed protein methionine. Plant. Mol. Biol. 1992, 18, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Molvig, L.; Tabe, L.M.; Eggum, B.O.; Moore, A.E.; Craig, S.; Spencer, D.; Higgins, T.J. Enhanced methionine levels and increased nutritive value of seeds of transgenic lupins (Lupinus angustifolius L.) expressing a sunflower seed albumin gene. Proc. Nat. Acad. Sci. USA 1997, 94, 8393–8398. [Google Scholar] [CrossRef] [PubMed]
- Tiger, T.T.; Miki, M.C.; Rolis, C.W.; Chen, L.J.; Su, R.C.; Wang, C.S.; Jason, T.C. Enhanced methionine and cysteine levels in transgenic rice seeds by the accumulation of sesame 2S albumin. Biosci. Biotechnol. Biochem. 2003, 67, 1699–1705. [Google Scholar]
- Frankard, V.; Ghislain, M.; Negrutiu, I.; Jacobs, M. High threonine producer mutant of Nicotiana sylvestris (Spegg. and Comes). Theor. Appl. Genet. 1991, 82, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Karchi, H.; Shaul, O.; Galili, G. Seed-specific expression of a bacterial desensitized aspartate kinase increases the production of seed threonine and methionine in transgenic tobacco. Plant J. 1993, 3, 721–727. [Google Scholar] [CrossRef]
- Hacham, Y.; Matityahu, I.; Schuster, G.; Amir, R. Overexpression of mutated forms of aspartate kinase and cystathionine γ-synthase in tobacco leaves resulted in the high accumulation of methionine and threonine. Plant J. 2008, 54, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Zeh, M.; Casazza, A.P.; Kreft, O.; Roessner, U.; Bieberich, K.; Willmitzer, L.; Hesse, H. Antisense inhibition of threonine synthase leads to high methionine content in transgenic potato plants. Plant Physiol. 2001, 127, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Kreps, J.A.; Ponappa, T.; Dong, W.; Town, C.D. Molecular basis of α-methyltryptophan resistance in amt-1, a mutant of Arabidopsis thaliana with altered tryptophan metabolism. Plant Physiol. 1996, 110, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Last, R.L. The Arabidopsis thaliana Trp5 mutant has a feedback-resistant anthranilate synthase and elevated soluble tryptophan. Plant Physiol. 1996, 110, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wakasa, K.; Hasegawa, H.; Nemoto, H.; Matsuda, F.; Miyazawa, H.; Tozawa, Y.; Miyagawa, H. High-level tryptophan accumulation in seeds of transgenic rice and its limited effects on agronomic traits and seed metabolite profile. J. Exp. Bot. 2006, 57, 3069–3078. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Asada, Y.; Takahashi, Y.; Yabe, N.; Komeda, Y.; Nishioka, T.; Wakasa, K. Metabolic changes in Arabidopsis thaliana expressing the feedback-resistant anthranilate synthase α subunit gene OASA1D. Phytochemistry 2006, 67, 2349–2362. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Tozawa, Y.; Hasegawa, H.; Terakawa, T.; Ohkawa, Y.; Wakasa, K. Use of a feedback-insensitive α subunit of anthranilate synthase as a selectable marker for transformation of rice and potato. Mol. Breed. 2004, 14, 363–373. [Google Scholar] [CrossRef]
- Araújo, W.L.; Ishizaki, K.; Nunes-Nesi, A.; Larson, T.R.; Tohge, T.; Krahnert, I.; Leaver, C.J. Identification of the 2-hydroxyglutarate and isovaleryl-CoA dehydrogenases as alternative electron donors linking lysine catabolism to the electron transport chain of Arabidopsis mitochondria. Plant Cell 2010, 22, 1549–1563. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Amir, R.; Fernie, A.R. The regulation of essential amino acid synthesis and accumulation in plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Shaul, O.; Galili, G. Threonine overproduction in transgenic tobacco plants expressing a mutant desensitized aspartate kinase of Escherichia coli. Plant Physiol. 1992, 100, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Guenoune, D.; Amir, R.; Badani, H.; Wolf, S.; Galili, S. Combined expression of S-VSPα in two different organelles enhances its accumulation and total lysine production in leaves of transgenic tobacco plants. J. Exp. Bot. 2002, 53, 1867–1870. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, W.; Bauw, G.; van Montagu, M.; Inzé, D. Distinct phenotypes generated by overexpression and suppression of S-adenosyl-l-methionine synthetase reveal developmental patterns of gene silencing in tobacco. Plant Cell 1994, 6, 1401–1414. [Google Scholar] [CrossRef] [PubMed]
- Saalbach, I.; Waddell, D.; Pickardt, T.; Schieder, O.; Müntz, K. Stable expression of the sulphur-rich 2S albumin gene in transgenic Vicia narbonensis increases the methionine content of seeds. J. Plant Physiol. 1995, 145, 674–681. [Google Scholar] [CrossRef]
- Di, R.; Kim, J.; Martin, M.N.; Leustek, T.; Jhoo, J.; Ho, C.T.; Tumer, N.E. Enhancement of the primary flavor compound methional in potato by increasing the level of soluble methionine. J. Agric. Food Chem. 2003, 51, 5695–5702. [Google Scholar] [CrossRef] [PubMed]
- Demidov, D.; Horstmann, C.; Meixner, M.; Pickardt, T.; Saalbach, I.; Galili, G.; Müntz, K. Additive effects of the feed-back insensitive bacterial aspartate kinase and the Brazil nut 2S albumin on the methionine content of transgenic narbon bean (Vicia narbonensis L.). Mol. Breed. 2003, 11, 187–201. [Google Scholar] [CrossRef]
- Chiaiese, P.; Ohkama-Ohtsu, N.; Molvig, L.; Godfree, R.; Dove, H.; Hocart, C.; Tabe, L.M. Sulphur and nitrogen nutrition influence the response of chickpea seeds to an added, transgenic sink for organic sulphur. J. Exp. Bot. 2004, 55, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Amira, G.; Ifat, M.; Tal, A.; Hana, B.; Shmuel, G.; Rachel, A. Soluble methionine enhances accumulation of a 15 kDa zein, a methionine-rich storage protein, in transgenic alfalfa but not in transgenic tobacco plants. J. Exp. Bot. 2005, 56, 2443–2452. [Google Scholar] [CrossRef] [PubMed]
- Dancs, G.; Kondrák, M.; Bánfalvi, Z. The effects of enhanced methionine synthesis on amino acid and anthocyanin content of potato tubers. BMC Plant Biol. 2008, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, M.S.; Rahman, S.M.; Nakamoto, Y.; Fujiwara, T.; Naito, S.; Wakasa, K.; Ishimoto, M. Differential response of methionine metabolism in two grain legumes, soybean and azuki bean, expressing a mutated form of Arabidopsis cystathionine γ-synthase. J. Plant Physiol. 2013, 170, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Brotherton, J.E.; Song, H.S.; Widholm, J.M. Increasing tryptophan synthesis in a forage legume Astragalus sinicus by expressing the tobacco feedback-insensitive anthranilate synthase (ASA2) gene. Plant Physiol. 2000, 123, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Brotherton, J.E.; Widholm, J.M.; Portis, A.R. Targeting a nuclear anthranilate synthase α-subunit gene to the tobacco plastid genome results in enhanced tryptophan biosynthesis. Return of a gene to its pre-endosymbiotic origin. Plant Physiol. 2001, 127, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Kita, Y.; Nakamoto, Y.; Takahashi, M.; Kitamura, K.; Wakasa, K.; Ishimoto, M. Manipulation of amino acid composition in soybean seeds by the combination of deregulated tryptophan biosynthesis and storage protein deficiency. Plant Cell Rep. 2010, 29, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Araki, M. Consumer acceptance of food crops developed by genome editing. Plant Cell Rep. 2016, 35, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
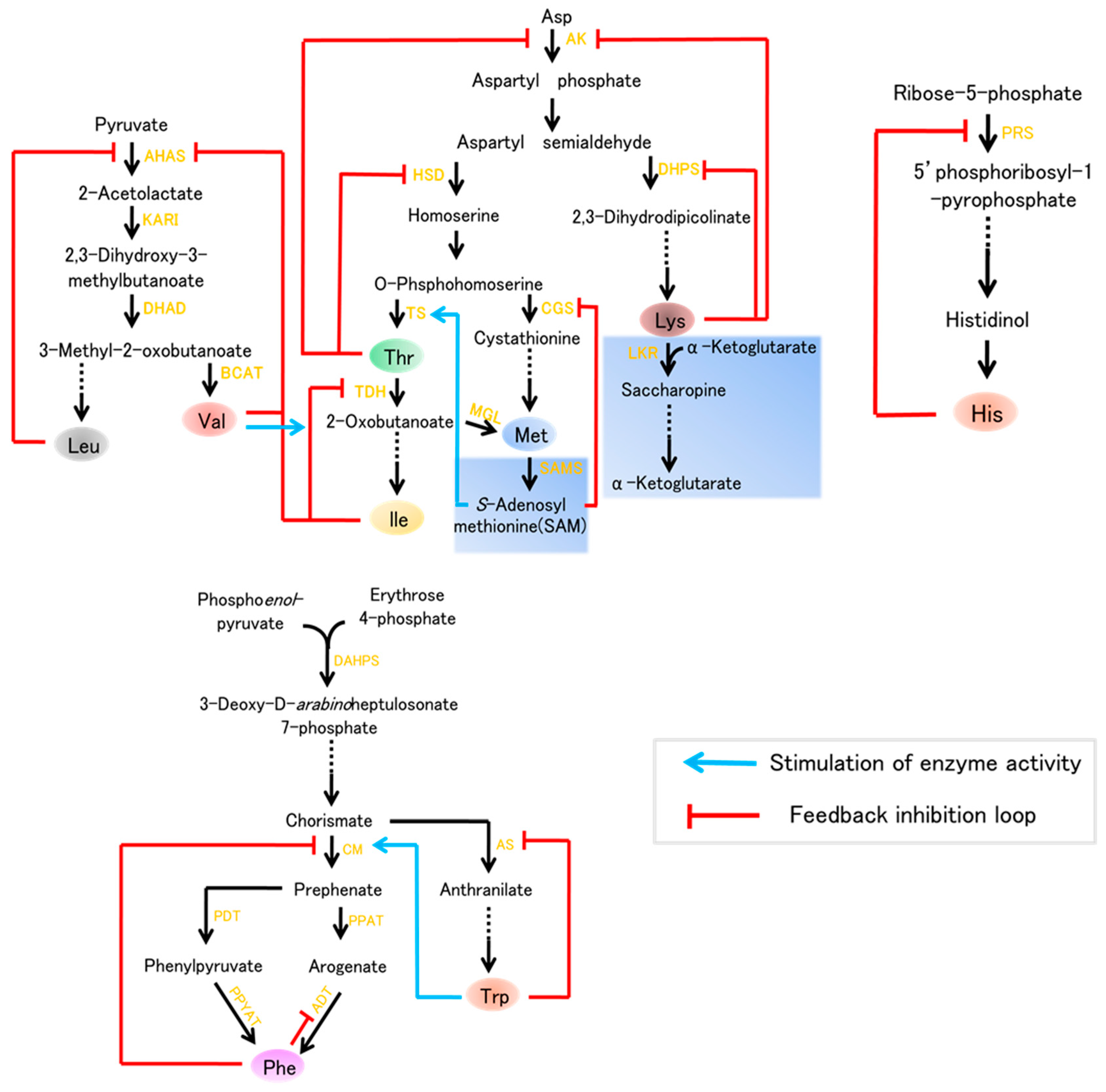

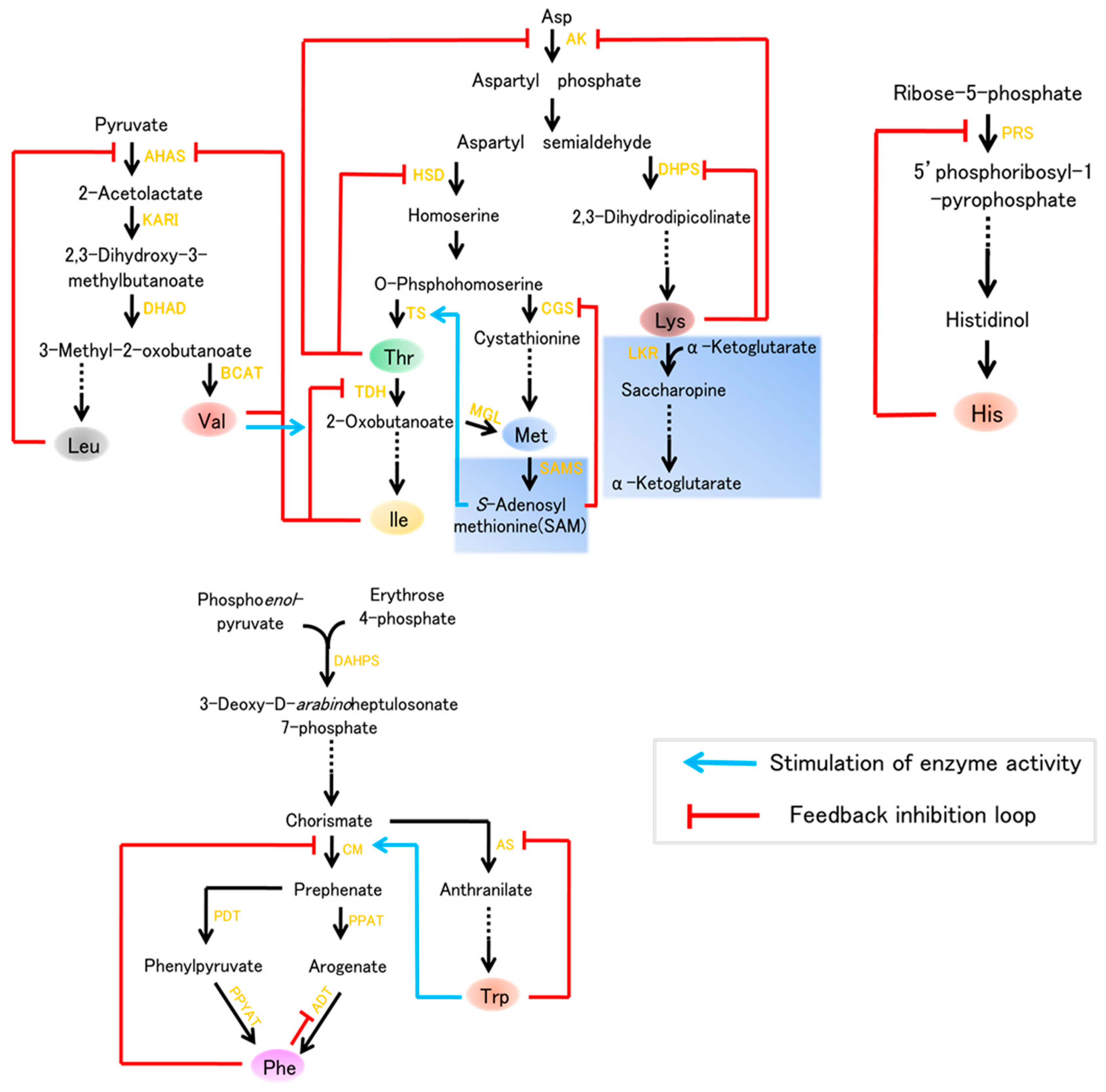
{kind=link}
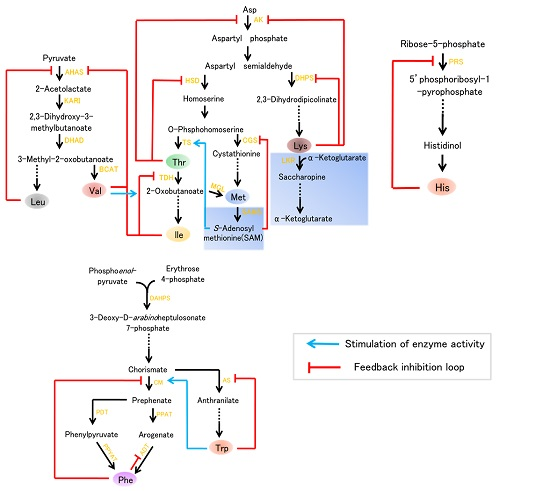
{kind=link}
| Amino Acid | Plant | Transgene | Tissue | Free fold Increase T/WT (Souble) | Phenotype | References |
|---|---|---|---|---|---|---|
| Lys | Potato | Constitutive::DHDPS | Leaves | 2–4 | ND | [20] |
| Potato | Constitutive::DHDPS | Roots | 1.2–5 | ND | [20] | |
| Potato | Constitutive::DHDPS | Tubers | 2–3.5 | ND | [20] | |
| Tobacco | Constitutive::AK | Leaves | 1.1–2 | ND | [39] | |
| Tobacco | Constitutive::DHDPS | Leaves | 9–11 | A partial loss of apical dominance, delayed flowering and senescence, partial sterility, and abnormal leaf morphology | [15] | |
| Tobacco | Seed-specific::DHDPS | Seeds | 1–2 | ND | [23] | |
| Canola | Seed-specific::DHDPS/AK | Seeds | More than 100 | ND | [21] | |
| Soybean | Seed-specific::DHDPS/AK | Seeds | Several hundreds | Wrinkled seed with poor germinaiton | [21] | |
| Soybean | Soybean vegetative storage proteins(S-VSPs) | Leaves | NR | ND | [40] | |
| Tobacco | Constitutive::DHDPS | Leaves | 30 | Mosaic green color in newly developed leaves at the tip of the apex and partial loss of apical dominance | [22] | |
| Tobacco | Constitutive::DHDPS/Constitutive::AtCGS | Leaves | 24 | ND | [22] | |
| Thr | Tobacco | Constitutive::AK | Leaves | 2–8 | ND | [39] |
| Tobacco | Constitutive::DHDPS | Leaves | 6–8 | A partial loss of apical dominance, delayed flowering and senescence, partial sterility,and abnormal leaf morphology | [15] | |
| Tobacco | Seed-specific::DHDPS | Seeds | 1 | ND | [23] | |
| Tobacco | Supress AK | Seeds | About 5.8 | ND | [23] | |
| Tobacco | Seed-specific::DHDPS/AK | Seeds | About 3.9 | ND | [23] | |
| Tobacco | Constitutive::AtCGS | Leaves | 1.8 | ND | [22] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 173 | ND | [30] | |
| Met | Tobacco | antisense SAMS | Leaves | 433 | Vein showed dark gren | [41] |
| Vicia narbonensis | Seed-specific::2S BN/AK | Seeds | 2 | ND | [42] | |
| Lupin | Seed-specific::2S SSA | Seeds | 2 | Statistically significant increases in live weight gain, true protein digestibility, biological value, and net protein utilization | [26] | |
| Potato | Antisense TS | Leaves | 239 | Severe growth retardation, strong chlorosis, and an acute reduction in tuber yield | [31] | |
| Potato | Constitutive::AtCGS | Leaves | 2–7.1 | ND | [43] | |
| Potato | Constitutive::AtCGS | Root | 1.2–4.1 | ND | [43] | |
| Bean | Seed-specific::2S BN | Seeds | 1.25 | ND | [44] | |
| Chickpea | Seed-specific::2S SSA | Seeds | 1.9 | ND | [45] | |
| Alfalfa | Constitutive::AtCGS | Leaves | 37 | ND | [46] | |
| Tobacco | Constitutive::DHDPS | Leaves | 2 | ND | [22] | |
| Tobacco | Constitutive::AtCGS | Leaves | 5 | ND | [22] | |
| Tobacco | Constitutive::DHDPS/Constitutive::AtCGS | Leaves | 10 | ND | [22] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 39 | [22] | ||
| Potato | Constitutive::AtCGS/zein | Tubers | 2–6 | ND | [47] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 39 | ND | [30] | |
| Soybean | Seed -specific::AtCGS | Seeds | 2.1 | ND | [48] | |
| Azuki bean | Seed -specific::AtCGS | Seeds | 2.3 | ND | [48] | |
| Trp | Astragalus | Constitutive::ASA2 | Hariy roots | 1.3–5.5 | ND | [49] |
| Tobacoo | Constitutive::ASA2 | Leaves | 16 | ND | [50] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 39 | ND | [30] | |
| Soybean | Seed -specific::OASA1D | Seeds | 50.9 | ND | [51] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Xu, M.; Wang, W.; Galili, G. Fortifying Horticultural Crops with Essential Amino Acids: A Review. Int. J. Mol. Sci. 2017, 18, 1306. https://doi.org/10.3390/ijms18061306
Wang G, Xu M, Wang W, Galili G. Fortifying Horticultural Crops with Essential Amino Acids: A Review. International Journal of Molecular Sciences. 2017; 18(6):1306. https://doi.org/10.3390/ijms18061306
Chicago/Turabian StyleWang, Guoping, Mengyun Xu, Wenyi Wang, and Gad Galili. 2017. "Fortifying Horticultural Crops with Essential Amino Acids: A Review" International Journal of Molecular Sciences 18, no. 6: 1306. https://doi.org/10.3390/ijms18061306
APA StyleWang, G., Xu, M., Wang, W., & Galili, G. (2017). Fortifying Horticultural Crops with Essential Amino Acids: A Review. International Journal of Molecular Sciences, 18(6), 1306. https://doi.org/10.3390/ijms18061306