
Melatonin Attenuates Pulmonary Hypertension in Chronically Hypoxic Rats

 ,
,
Abstract
:1. Introduction
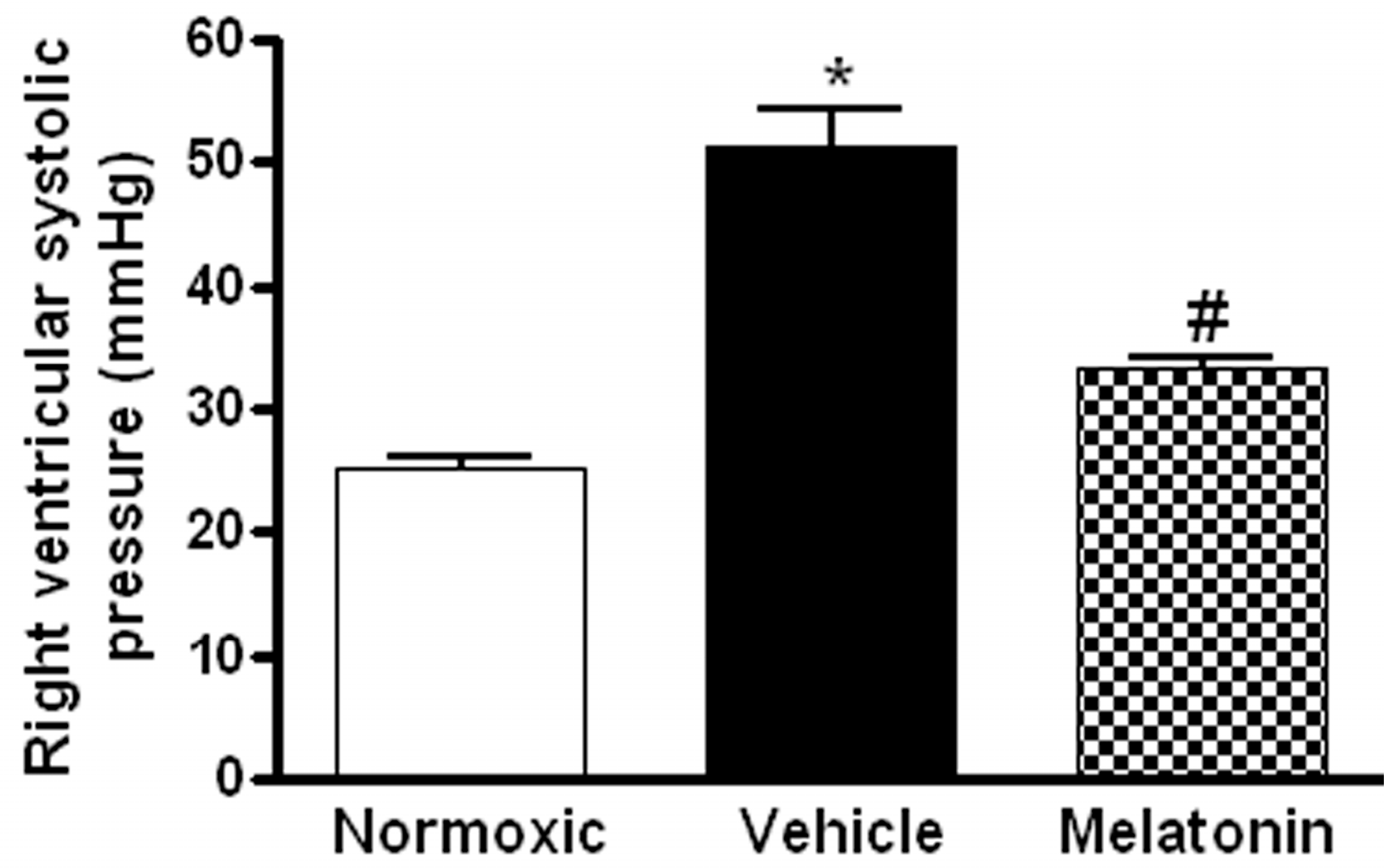
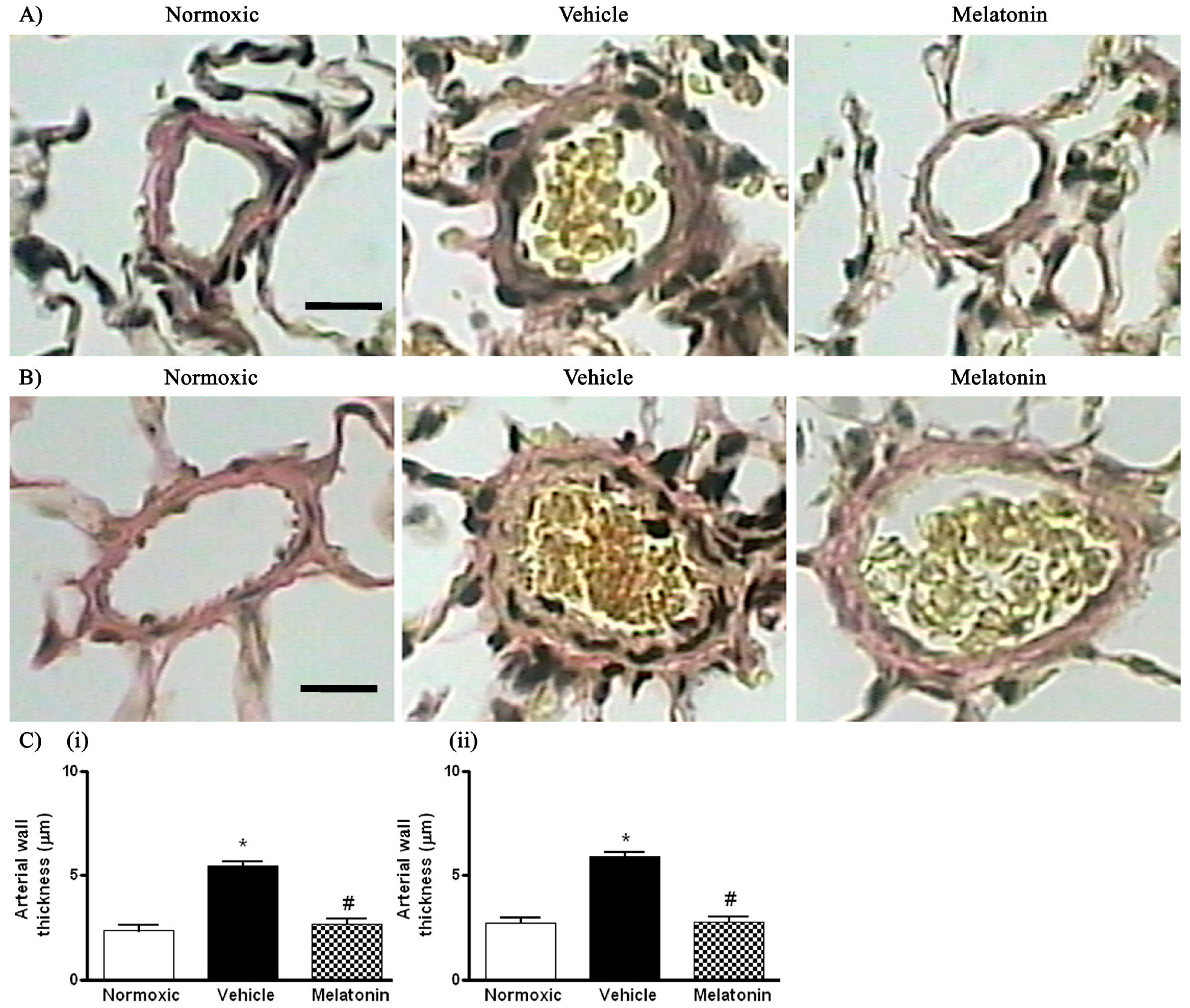
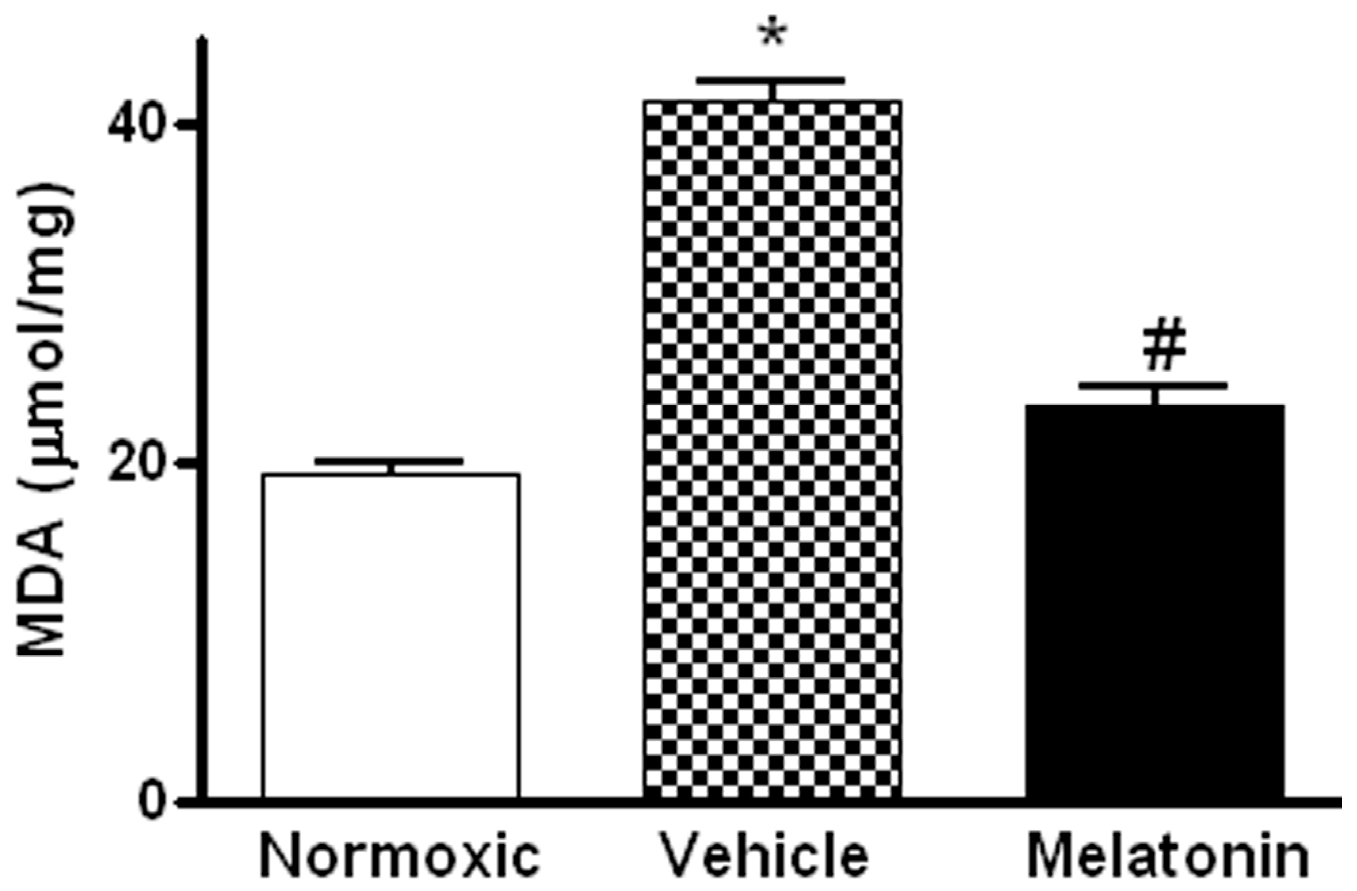
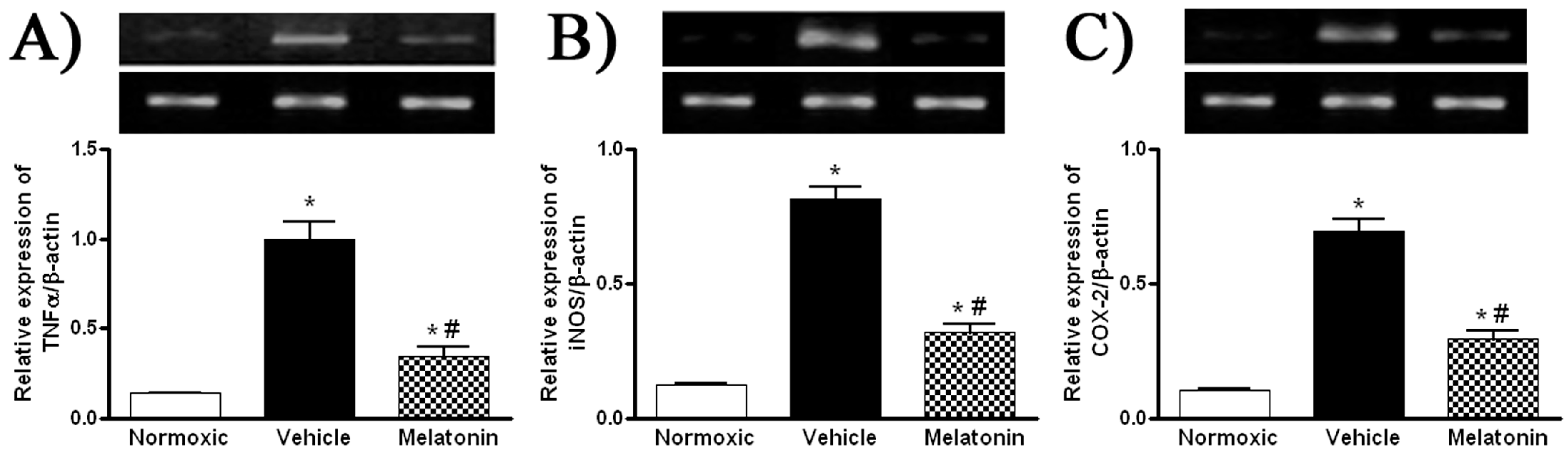
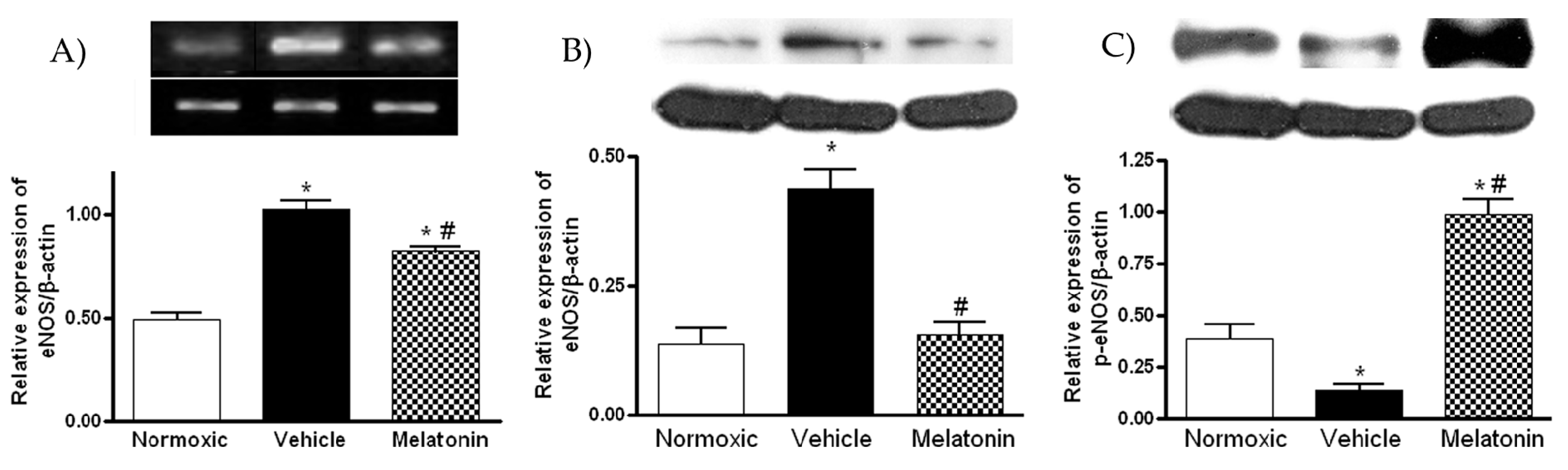
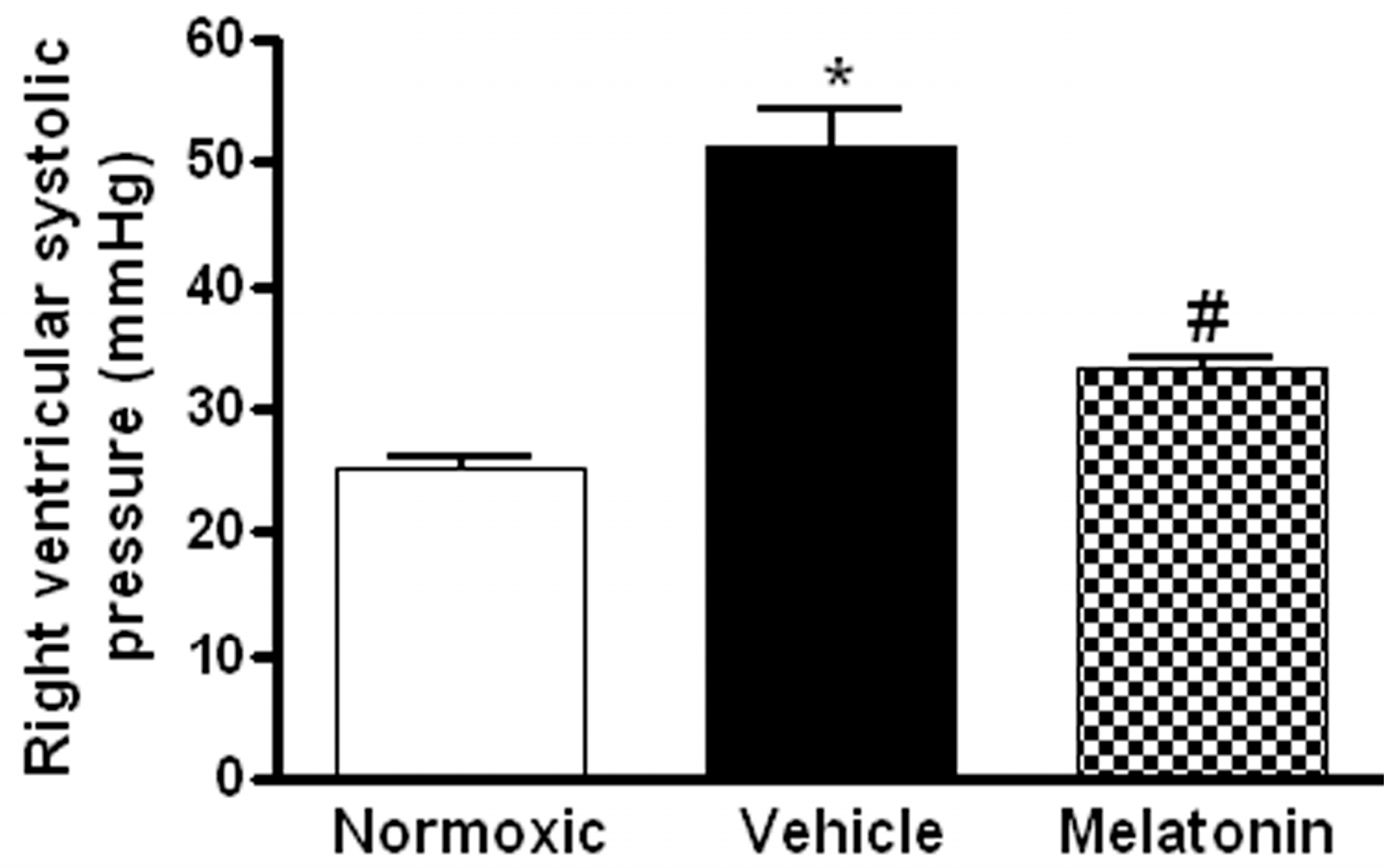
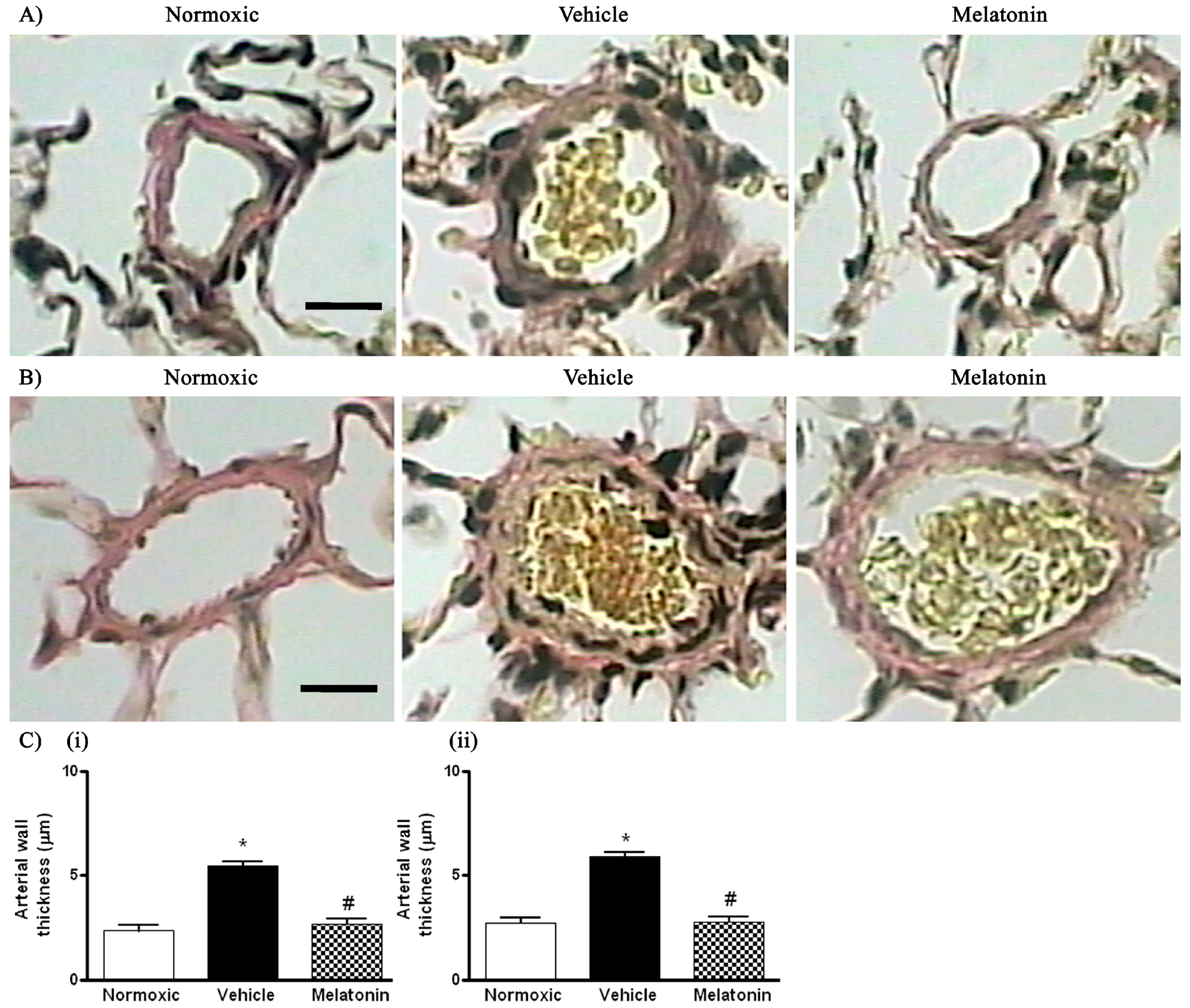
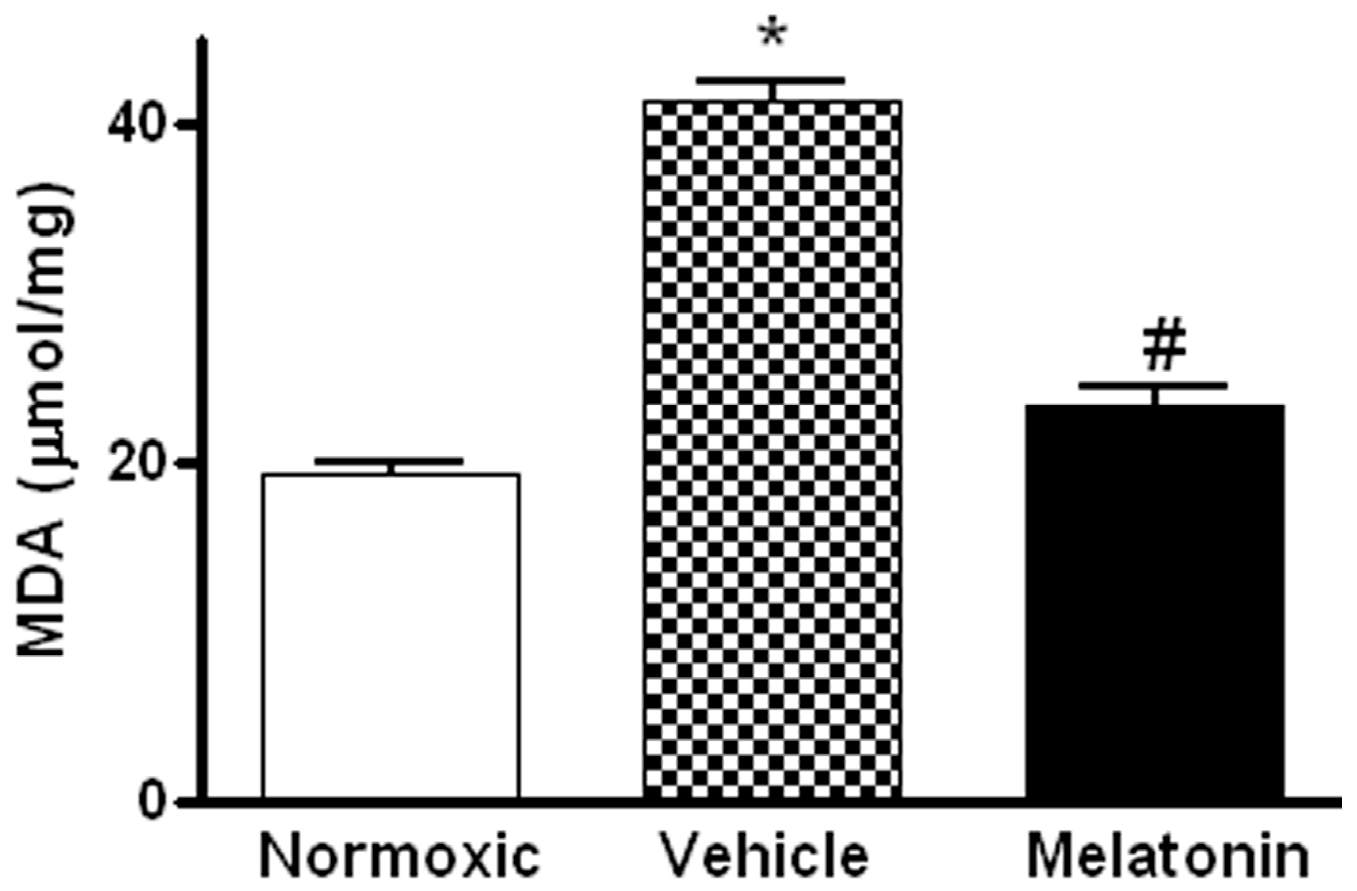
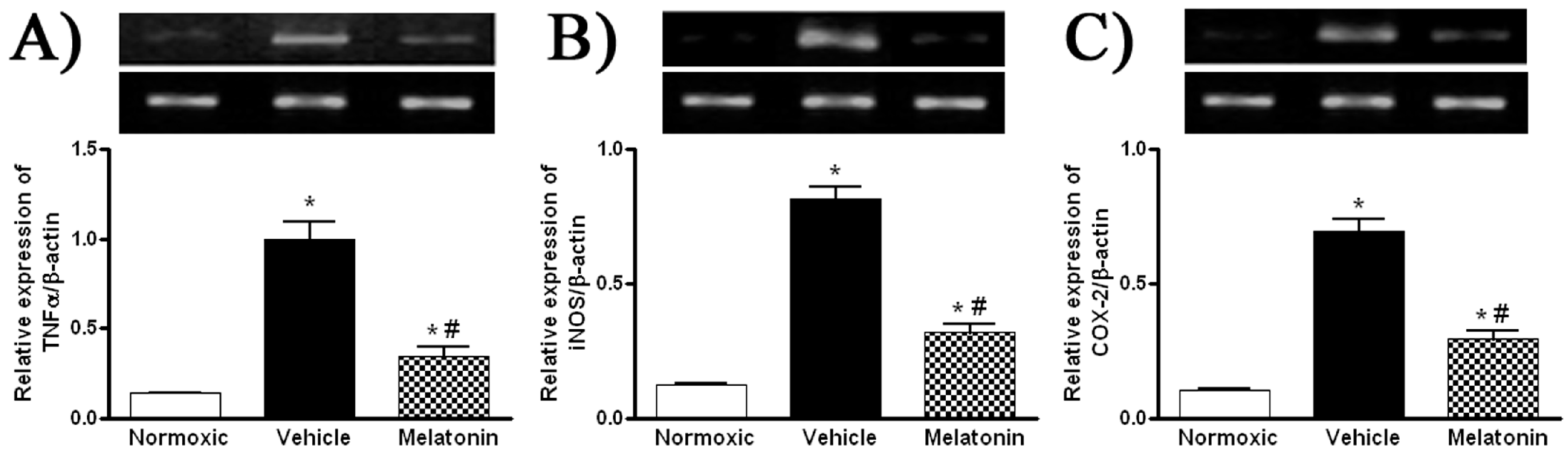
2. Results
3. Discussion
4. Material and Methods
4.1. Hypoxic Treatment on Rats
4.2. Drug Preparation
4.3. Measurements of the Right Ventricular Systolic Pressure and Heart Weights
4.4. Histological Analysis
4.5. Measurement of Malondialdehyde (MDA) Level
4.6. Western Blotting
4.7. Semi-Quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rabinovitch, M.; Gamble, W.; Nadas, A.S.; Miettinen, O.S.; Reid, L. Rat pulmonary circulation after chronic hypoxia: Hemodynamic and structural features. Am. J. Physiol. 1979, 236, H818–H827. [Google Scholar] [PubMed]
- Stanbrook, H.S.; Morris, K.G.; Mcmurtry, I.F. Prevention and reversal of hypoxic pulmonary hypertension by calcium antagonists. Am. Rev. Respir. Dis. 1984, 130, 81–85. [Google Scholar] [PubMed]
- Jefferson, J.A.; Simoni, J.; Escudero, E.; Hurtado, M.E.; Swenson, E.R.; Wesson, D.E.; Schreiner, G.F.; Schoene, R.B.; Johnson, R.J.; Hurtado, A. Increased oxidative stress following acute and chronic high altitude exposure. High Alt. Med. Biol. 2004, 5, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Hasleton, P.S.; Heath, D.; Brewer, D.B. Hypertensive pulmonary vascular disease in states of chronic hypoxia. J. Pathol. 1968, 95, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, K.R.; Fagan, K.A.; Frid, M.G. Hypoxia-induced pulmonary vascular remodeling: Cellular and molecular mechanisms. Circ. Res. 2006, 99, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Joppa, P.; Petrasova, D.; Stancak, B.; Dorková, Z.; Tkáčová, R. Oxidative stress in patients with COPD and pulmonary hypertension. Wien. Klin. Wochenschr. 2007, 119, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Pratico, D.; Basili, S.; Vieri, M.; Cordova, C.; Violi, F.; Fitzgerald, G.A. Chronic obstructive pulmonary disease is associated with an increase in urinary levels of isoprostane F2α-III, an index of oxidant stress. Am. J. Respir. Crit. Care Med. 1998, 158, 1709–1714. [Google Scholar] [CrossRef] [PubMed]
- Hoshikawa, Y.; Ono, S.; Suzuki, S.; Tanita, T.; Chida, M.; Song, C.; Noda, M.; Tabata, T.; Voelkel, N.F.; Fujimura, S.; et al. Generation of oxidative stress contributes to the development of pulmonary hypertension induced by hypoxia. J. Appl. Physiol. 2001, 90, 1299–1306. [Google Scholar] [PubMed]
- Frid, M.G.; Brunetti, J.A.; Burke, D.L.; Carpenter, T.C.; Davie, N.J.; Reeves, J.T.; Roedersheimer, M.T.; van Rooijen, N.; Stenmark, K.R. Hypoxia-induced pulmonary vascular remodeling requires recruitment of circulating mesenchymal precursors of a monocyte/macrophage lineage. Am. J. Pathol. 2006, 168, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Takabatake, N.; Nakamura, H.; Abe, S.; Inoue, D.; Hino, T.; Saito, H.; Yuki, H.; Kato, S.; Tomoike, H. The relationship between chronic hypoxemia and activation of the tumor necrosis factor-α system in patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2000, 161, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Shaul, P.W.; North, A.J.; Brannon, T.S.; Ujiie, K.; Wells, L.B.; Nisen, P.A.; Lowenstein, C.J.; Snyder, S.H.; Star, R.A. Prolonged in vivo hypoxia enhances nitric oxide synthase type I and type III gene expression in adult rat lung. Am. J. Respir. Cell Mol. Biol. 1995, 13, 167–174. [Google Scholar] [CrossRef]
- Le Cras, T.D.; Xue, C.; Rengasamy, A.; Johns, R.A. Chronic hypoxia upregulates endothelial and inducible NO synthase gene and protein expression in rat lung. Am. J. Physiol. 1996, 270, L164–L170. [Google Scholar] [PubMed]
- Resta, T.C.; Gonzales, R.J.; Dail, W.G.; Sanders, T.C.; Walker, S.B.R. Selective upregulation of arterial endothelial nitric oxide synthase in pulmonary hypertension. Am. J. Physiol. 1997, 272, H806–H813. [Google Scholar] [PubMed]
- Resta, T.C.; Walker, B.R. Chronic hypoxia selectively augments endothelium-dependent pulmonary arterial vasodilation. Am. J. Physiol. 1996, 270, H888–H896. [Google Scholar] [PubMed]
- Drummond, G.R.; Cai, H.; Davis, M.E.; Ramasamy, S.; Harrison, D.G. Transcriptional and posttranscriptional regulation of endothelial nitric oxide synthase expression by hydrogen peroxide. Circ. Res. 2000, 86, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Davis, M.E.; Drummond, G.R.; Harrison, D.G. Induction of endothelial NO synthase by hydrogen peroxide via a Ca2+/calmodulin-dependent protein kinase II/janus kinase 2-dependent pathway. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1571–1576. [Google Scholar] [CrossRef] [PubMed]
- Cella, G.; Sbarai, A.; Mazzaro, G.; Vanzo, B.; Romano, S.; Hoppensteadt, D.; Fareed, J. Plasma markers of endothelial dysfunction in chronic obstructive pulmonary disease. Clin. Appl. Thromb. Hemost. 2001, 7, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Clini, E.; Bianchi, L.; Pagani, M.; Ambrosino, N. Endogenous nitric oxide in patients with stable COPD: Correlates with severity of disease. Thorax 1998, 53, 881–883. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.J.; Lopez-Pingarron, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, Afmk and Amk. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Girouard, H.; Chulak, C.; Lejossec, M.; Lamontagne, D.; de Champlain, J. Vasorelaxant effects of the chronic treatment with melatonin on mesenteric artery and aorta of spontaneously hypertensive rats. J. Hypertens. 2001, 19, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.M.; Meki, A.R.; Rahma, H.H. Inhibitory effects of melatonin on vascular reactivity: Possible role of vasoactive mediators. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 357–367. [Google Scholar] [CrossRef]
- Torres, F.; González-Candia, A.; Montt, C.; Ebensperger, G.; Chubretovic, M.; Serón-Ferré, M.; Reyes, R.V.; Llanos, A.J.; Herrera, E.A. Melatonin reduces oxidative stress and improves vascular function in pulmonary hypertensive newborn sheep. J. Pineal Res. 2015, 58, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Maarman, G.; Blackhurst, D.; Thienemann, F.; Blauwet, L.; Butrous, G.; Davies, N.; Sliwa, K.; Lecour, S. Melatonin as a preventive and curative therapy against pulmonary hypertension. J. Pineal Res. 2015, 59, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Wang, Y.; Zhou, L.; Liu, L.; Zhang, P.; Deng, W.; Yuan, Y. Melatonin attenuates hypoxic pulmonary hypertension by inhibiting the inflammation and the proliferation of pulmonary arterial smooth muscle cells. J. Pineal Res. 2014, 57, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Bhat, G.B.; Block, E.R. Effect of hypoxia on phospholipid metabolism in porcine pulmonary artery endothelial cells. Am. J. Physiol. 1992, 262, L606–L613. [Google Scholar] [PubMed]
- Chang, S.W.; Stelzner, T.J.; Weil, J.V.; Voelkel, N.F. Hypoxia increases plasma glutathione disulfide in rats. Lung 1989, 167, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.S.; Whatley, R.E.; Cain, P.; McIntyre, T.M.; Prescott, S.M.; Zimmerman, G.Z. Hydrogen peroxide stimulates the synthesis of platelet-activating factor by endothelium and induces endothelial cell-dependent neutrophil adhesion. J. Clin. Investig. 1988, 82, 2045–2055. [Google Scholar] [CrossRef] [PubMed]
- Hartung, H.P.; Parnham, M.J.; Winkelmann, J.; Englberger, W.; Hadding, U. Platelet activating factor (PAF) induces the oxidative burst in macrophages. Int. J. Immunopharmacol. 1983, 5, 115–121. [Google Scholar] [CrossRef]
- Ali, M.H.; Schlidt, S.A.; Chandel, N.S.; Hynes, K.L.; Schumacker, P.T.; Gewertz, B.L. Endothelial permeability and IL-6 production during hypoxia: Role of ROS in signal transduction. Am. J. Physiol. 1999, 277, L1057–L1065. [Google Scholar] [PubMed]
- Okada, M.; Matsuto, T.; Miida, T.; Inano, K. Differences in the effects of cytokines on the expression of adhesion molecules in endothelial cells. Ann. Med. Interne (Paris) 1997, 148, 125–129. [Google Scholar] [PubMed]
- Wood, J.G.; Johnson, J.S.; Mattioli, L.F.; Gonzalez, N.C. Systemic hypoxia promotes leukocyte-endothelial adherence via reactive oxidant generation. J. Appl. Physiol. 1999, 87, 1734–1740. [Google Scholar] [PubMed]
- Beck-Schimmer, B.; Schimmer, R.C.; Madjdpour, C.; Bonvini, J.M.; Pasch, T.; Ward, P.A. Hypoxia mediates increased neutrophil and macrophage adhesiveness to alveolar epithelial cells. Am. J. Respir. Cell Mol. Biol. 2001, 25, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.J.; Lopes, R.H.; Lamy-Freund, M.T. Permeability of pure lipid bilayers to melatonin. J. Pineal Res. 1995, 19, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Cummins, E.P.; Taylor, C.T. Hypoxia-responsive transcription factors. Pflug. Arch. 2005, 450, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Mitani, Y.; Maruyama, J.; Jiang, B.A.; Ikeyama, Y.; Dida, F.A.; Yamamoto, H.; Imanaka-Yoshida, K.; Shimpo, H.; Mizoguchi, A.; et al. A NF-κB inhibitor pyrrolidine dithiocarbamate ameliorates pulmonary hypertension in rats. Chest J. 2007, 132, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Strukova, S. Blood coagulation-dependent inflammation. Coagulation-dependent inflammation and inflammation-dependent thrombosis. Front. Biosci. 2006, 11, 59–80. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.N.; Pohlman, T.H.; Vocelka, C.; Farr, A.; Lindley, G.; Chandler, W.; Griscavage-Ennis, J.M.; Verrier, E.D. NF-κB mediates a procoagulant response in monocytes during extracorporeal circulation. J. Thorac. Cardiovasc. Surg. 2003, 125, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: A pleiotropic molecule regulating inflammation. Biochem. Pharmacol. 2010, 80, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.; Collado, P.S.; Gonzalez-Gallego, J. Melatonin inhibits the expression of the inducible isoform of nitric oxide synthase and NF-κB activation in rat skeletal muscle. J. Pineal Res. 2006, 41, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Chetsawang, B.; Putthaprasart, C.; Phansuwan-Pujito, P.; Govitrapong, P. Melatonin protects against hydrogen peroxide-induced cell death signaling in SH-SY5Y cultured cells: Involvement of NF-κB, Bax and Bcl-2. J. Pineal Res. 2006, 41, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, W.; Wang, N.P.; Gui, S.Y.; Wu, L.; Sun, W.Y.; Xu, S.Y. Melatonin ameliorates carbon tetrachloride-induced hepatic fibrogenesis in rats via inhibition of oxidative stress. Life Sci. 2005, 77, 1902–1915. [Google Scholar] [CrossRef] [PubMed]
- Whorton, A.R.; Simonds, D.B.; Piantadosi, C.A. Regulation of nitric oxide synthesis by oxygen in vascular endothelial cells. Am. J. Physiol. 1997, 272, L1161–L1166. [Google Scholar] [PubMed]
- Lilley, E.; Gibson, A. Antioxidant protection of NO-induced relaxations of the mouse anococcygeus against inhibition by superoxide anions, hydroquinone and carboxy-PTIO. Br. J. Pharmacol. 1996, 119, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Shaul, P.W.; Wells, L.B.; Horning, K.M. Acute and prolonged hypoxia attenuate endothelial nitric oxide production in rat pulmonary arteries by different mechanisms. J. Cardiovasc. Pharmacol. 1993, 22, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Kilic, E.; Kilic, U.; Reiter, R.J.; Bassetti, C.L.; Hermann, D.M. Tissue-plasminogen activator-induced ischemic brain injury is reversed by melatonin: Role of iNOS and Akt. J. Pineal Res. 2005, 39, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Kilic, U.; Kilic, E.; Reiter, R.J.; Bassetti, C.L.; Hermann, D.M. Signal transduction pathways involved in melatonin-induced neuroprotection after focal cerebral ischemia in mice. J. Pineal Res. 2005, 38, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Chun, W.; Kong, P.J.; Han, J.A.; Cho, B.P.; Kwon, O.Y.; Lee, H.J.; Kim, S.S. Sustained activation of Akt by melatonin contributes to the protection against kainic acid-induced neuronal death in hippocampus. J. Pineal Res. 2006, 40, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Pogan, L.; Bissonnette, P.; Parent, L.; Sauvé, R. The effects of melatonin on Ca2+ homeostasis in endothelial cells. J. Pineal Res. 2002, 33, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, A.J. Mechanisms of dysfunction of the nitric oxide pathway in vascular diseases. Nitric Oxide 2002, 6, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Balonan, L.; Ballard, H.J.; Ho, S. Chronic hypoxia inhibits the antihypertensive effect of melatonin on pulmonary artery. Int. J. Cardiol. 2008, 126, 340–345. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Product Size (bp) | Annealing Temperature (T °C) | No. of Cycles (X) |
|---|---|---|---|---|
| COX-2 | F: 5′-gtgttgacgtccagatcaca-3′ R: 5′-agtatgagcctgctggtttg-3′ | 662 | 58 | 28 |
| iNOS | F: 5′-tagaggaacatctggccagg3′ R: 5′-tggccgacctgatgttgcca-3′ | 255 | 58 | 33 |
| TNFα | F: 5′-atgagcacagaaagcatgatc-3′ R: 5′-tacaggcttgtcactcgaatt-3′ | 276 | 50 | 35 |
| β-actin | F: 5′-agccatgtacgtagccatcc-3′ R: 5′-ctctcagctgtggtggtgaa-3′ | 228 | 56 | 28 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, M.W.; Yeung, H.M.; Lau, C.F.; Poon, A.M.S.; Tipoe, G.L.; Fung, M.L. Melatonin Attenuates Pulmonary Hypertension in Chronically Hypoxic Rats. Int. J. Mol. Sci. 2017, 18, 1125. https://doi.org/10.3390/ijms18061125
Hung MW, Yeung HM, Lau CF, Poon AMS, Tipoe GL, Fung ML. Melatonin Attenuates Pulmonary Hypertension in Chronically Hypoxic Rats. International Journal of Molecular Sciences. 2017; 18(6):1125. https://doi.org/10.3390/ijms18061125
Chicago/Turabian StyleHung, Ming Wai, Hang Mee Yeung, Chi Fai Lau, Angela Ming See Poon, George L. Tipoe, and Man Lung Fung. 2017. "Melatonin Attenuates Pulmonary Hypertension in Chronically Hypoxic Rats" International Journal of Molecular Sciences 18, no. 6: 1125. https://doi.org/10.3390/ijms18061125
APA StyleHung, M. W., Yeung, H. M., Lau, C. F., Poon, A. M. S., Tipoe, G. L., & Fung, M. L. (2017). Melatonin Attenuates Pulmonary Hypertension in Chronically Hypoxic Rats. International Journal of Molecular Sciences, 18(6), 1125. https://doi.org/10.3390/ijms18061125