Abstract
Sailuotong (SLT) is a standardised three-herb formulation consisting of Panax ginseng, Ginkgo biloba, and Crocus sativus designed for the management of vascular dementia. While the latest clinical trials have demonstrated beneficial effects of SLT in vascular dementia, the underlying cellular mechanisms have not been fully explored. The aim of this study was to assess the ability and mechanisms of SLT to act against hydrogen peroxide (H2O2)-induced oxidative damage in cultured human vascular endothelial cells (EAhy926). SLT (1–50 µg/mL) significantly suppressed the H2O2-induced cell death and abolished the H2O2-induced reactive oxygen species (ROS) generation in a concentration-dependent manner. Similarly, H2O2 (0.5 mM; 24 h) caused a ~2-fold increase in lactate dehydrogenase (LDH) release from the EA.hy926 cells which were significantly suppressed by SLT (1–50 µg/mL) in a concentration-dependent manner. Incubation of SLT (50 µg/mL) increased superoxide dismutase (SOD) activity and suppressed the H2O2-enhanced Bax/Bcl-2 ratio and cleaved caspase-3 expression. In conclusion, our results suggest that SLT protects EA.hy916 cells against H2O2-mediated injury via direct reduction of intracellular ROS generation and an increase in SOD activity. These protective effects are closely associated with the inhibition of the apoptotic death cascade via the suppression of caspase-3 activation and reduction of Bax/Bcl-2 ratio, thereby indicating a potential mechanism of action for the clinical effects observed.
1. Introduction
Cerebrovascular disease (CVD), such as stroke and vascular dementia, is a leading cause of morbidity and mortality, imposing a huge financial burden on the health care system worldwide. Progression of CVD is mediated via a numbers of factors, such as aging, hypertension, inflammation, and atherosclerosis, over a prolonged period [1,2]. It is now well established that many of these factors are closely associated with a chronic increase of oxidative stress, which can lead to vascular endothelial damage [3,4]. Oxidative stress is caused by excessive generation of reactive oxygen species (ROS), including superoxide anions and hydroxyl radicals. Hydrogen peroxide (H2O2) is one of the major ROS and it has been shown to be generated during ischemia-reperfusion injury and in animal models of chronic brain disorders [5,6]. In vitro studies have demonstrated that H2O2 can induce oxidative stress in endothelial cells, causing cellular dysfunction and apoptosis [7].
Interestingly, recent studies have demonstrated a close linkage between neurological dysfunction and vascular disease, highlighting the crucial role of cell–cell signalling between neurons, astrocytes, and endothelial cells within the neurovascular unit (NVU) for normal brain functions [8,9]. It has been suggested that the cerebral endothelium and microvessels system are not just “empty pipes” for blood circulation, but instead act as an intricate paracrine organ that supports neuronal functions and homeostasis [10]. Numerous studies have suggested that endothelial cells play a fundamental role in various physiological and pathological processes. For example, Sorriento et al. demonstrated that endothelial cells are able to synthesise and release catecholamine in response to ischemia, highlighting their vital roles in the control of vascular homeostasis [11]. A recent report also suggested that microRNA contributes significantly to various therapeutic approaches to preserve endothelial integrity and vascular health [12], such as vascularization, a process that is closely associated with ROS-dependent signalling [13,14]. Moreover, salidroside, a glycoside from Rhodiola rosea, has been shown to reduce H2O2-induced ROS generation via the upregulation of miR-103 [15]. Hence, interventions that can suppress ROS-induced endothelial cell damage would be beneficial for CVDs as well as neurological disorders.
Herbal medicine has been widely used in CVD management in many Asian countries for centuries. A large body of evidence has shown that many herbs have remarkable anti-oxidative properties [16]. Sailuotong (SLT) is a standardised three-herb formula combining specific dosages of Panax ginseng (ginseng), Ginkgo biloba (ginkgo), and Crocus sativus (saffron) for the management of vascular dementia (VaD) [17,18]. The chemical profile and optimal ratio of the three herbal extracts have been determined and studied in detail previously [19]. In a chronic cerebral hypoperfusion model induced by bilateral common carotid artery ligation in rats, an eight week treatment of SLT (ig) significantly shortened the persistent time for finding the platform in a Morris Water Maze task. This beneficial effect was found to be associated with an increased acetylcholine level and superoxide dismutase (SOD) activity in the brain [20]. SLT (8.25, 16.5, and 33 mg/kg over 24 h) has been shown to significantly decrease the areas of focal cerebral ischemia/reperfusion injury by increasing cerebral blood flow in anesthetized dogs [21]. Moreover, SLT treatment (16 mg/kg and 8 mg/kg for seven days) also significantly decreased the platelet aggregation rate and whole blood viscosity in rabbits [21].
Cerebral and vascular protective effects of the individual components of SLT have been demonstrated previously. For instance, crocin, the principal active component of Crocus sativus, has been shown to act against cerebral ischemia/reperfusion (I/R) damage via an increase of SOD and glutathione peroxidase (GSH-px) activities and a reduction of metalloproteinase-9 (MMP-9) expression in a global cerebral ischemia mouse model [22]. Similarly, ginsenoside Rb1, a natural steroid glycoside from Panax ginseng, has been shown to reduce brain damage caused by middle cerebral artery occlusion (MCAO) via reduction of cerebral oxidative stress in aged mice [23]. EGb761, a standardised extract of Ginkgo biloba, has been shown to protect bovine vascular endothelial cells against oxidative stress-induced damage via reduced intracellular ROS generation and the regulation of apoptosis-related genes expression, including Bax and Bcl-2 [24,25].
It is worth noting that, despite a recent clinical study demonstrating that SLT improved cognitive function in people with VaD [18], and numerous pre-clinical studies demonstrating the cerebrovascular protective effects of its individual components [23,26,27], the cellular mechanisms of SLT in protecting endothelial cells from oxidative stress have not been previously studied. Thus, the aims of this study were to investigate the effects of SLT on H2O2-induced endothelial cell injury using cultured human umbilical vein cell line, EA.hy926, and to explore the mechanisms of action underlying these effects.
2. Results
2.1. Effects of SLT on the Viability of EA.hy926 Cells Injured by H2O2
Cytotoxicity of SLT (0.1–200 µg/mL; n = 3) on EA.hy926 cells was examined using MTT (3-(4,5-di-methylthiazol-2-yl) 2,5-diphenyltetrazolium bromide) assay. SLT did not show any significant cytotoxic effects up to 50 µg/mL [28]. Therefore, all the subsequent experiments were conducted at doses no higher than 50 µg/mL SLT.
To evaluate whether SLT could act against H2O2-induced cell damage, cells were pre-incubated with SLT for 60 min, then challenged by H2O2 (0.5 mM) for 24 h; cell viability was measured by MTT assay. EA.hy926 cell viability was markedly reduced by H2O2 (0.5 mM; 24 h) (p < 0.001, n = 3). Pre-incubation of SLT (0.1–50 µg/mL) protected cells from H2O2-induced cell damage (p < 0.01 at 50 µg/mL; n = 3) (Figure 1A,B). These results indicate that SLT could protect EA.hy926 cells from ROS-related cellular damage.
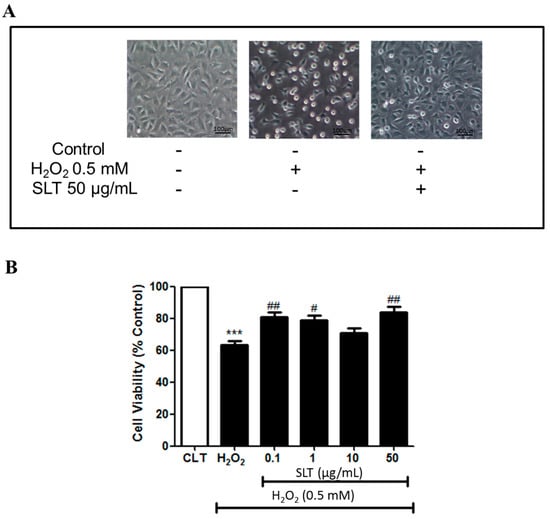
Figure 1.
(A) Representative images of the effect of Sailuotong (SLT) (50 µg/mL) on EA.hy926 cell morphology injured by H2O2 observed under an inverted/phase contract microscope; (B) Effect of Sailuotong (SLT) (0.1–50 µg/mL) on EA.hy926 cells viability injured by H2O2 (n = 3) measured by MTT assay. Data are presented as means ± S.D. *** p < 0.001 vs. control group; # p < 0.05 vs. H2O2 group; ## p < 0.01 vs. H2O2 group.
2.2. Effects of SLT on LDH Leakage and SOD Activity in H2O2 Treated EA.hy926 Cells
Lactate dehydrogenase (LDH) is one of the major representative indicators of cell injury. Therefore, the protective effect of SLT on H2O2-treated EA.hy926 cells was confirmed using LDH assay. As shown in Figure 2A, H2O2 (0.5 mM; 24 h) markedly increased LDH leakage from the EA.hy926 cells (p < 0.05, n = 3), while SLT reduced this H2O2-mediated LDH leakage in a concentration-dependent manner (p < 0.05 at 50 µg/mL compared to H2O2 alone; n = 3).
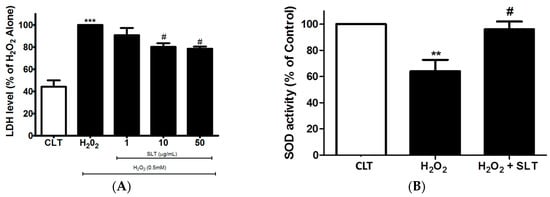
Figure 2.
(A) Effects of SLT (1–50 µg/mL) on H2O2-induced lactate dehydrogenase (LDH) leakage in EA.hy926 cells (n = 3). Data are presented as means ± S.D. *** p < 0.001 vs. control (CLT) group; # p < 0.05 vs. H2O2 group; (B) Effects of SLT (50 µg/mL) on H2O2-inhibited superoxide dismutase (SOD) activity in EA.hy926 cells (n = 3). Data are presented as means ± S.D. ** p < 0.01 vs. control (CLT) group; # p < 0.05 vs. H2O2 group.
To further examine the protective effects of SLT, we measured SOD activity in H2O2-treated EA.hy926 cells. SOD activity was significantly reduced by H2O2 (p < 0.05 compared to control, n = 3). This significant reduction of SOD activity was partly reversed by SLT at 50 µg/mL (Figure 2B).
2.3. Effect of SLT on the Intracellular ROS Generation in H2O2 Treated EA.hy926 Cells
In order to elucidate whether the protective effect of SLT is mediated by a reduction of intracellular oxidative stress, intracellular ROS generation was determined by 2′,7′-Dichlorofluorescin diacetate (DCFH-DA), ROS specific dye. As shown in Figure 3, H2O2 markedly increased (~2-fold) intracellular ROS generation in EA.hy926 cells (p < 0.001 compared to control, n = 3) and SLT (1–50 µg/mL) suppressed this H2O2-induced ROS generation in a concentration-dependent manner (p < 0.001 at 50 µg/mL compared to H2O2 alone; n = 3). Interestingly, the effect of SLT at 50 µg/mL in suppressing H2O2-induced ROS generation is comparable to gallic acid (10 µg/mL), a known potent anti-oxidant [29].
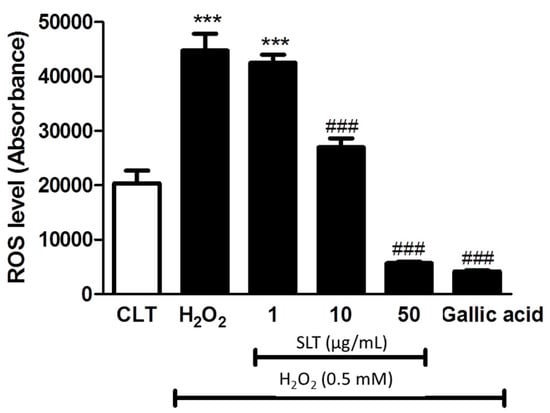
Figure 3.
Effects of SLT (1–50 µg/mL) on H2O2-induced intracellular reactive oxygen species (ROS) generation in EA.hy926 cells. Gallic acid (10 µg/mL), a known potent anti-oxidant, was used as a positive control (n = 3). Data are presented as means ± S.D. *** p < 0.05 vs. control group; ### p < 0.001 vs. H2O2 group.
2.4. Effect of SLT on Protein Expression Level of Bax, Bcl-2, and Cleaved Caspase-3 in H2O2 Treated EA.hy926 Cells
The protein expressions of Bax (pro-apoptotic factor) and Bcl-2 (anti-apoptotic factor) in H2O2 treated EA.hy926 cells were evaluated using Western blotting with the result expressed as Bax/Bcl-2 ratio. As shown in Figure 4A, H2O2 treatment markedly increased the Bax/Bcl-2 ratio (p < 0.05 compared to control, n = 3), while SLT at 50 µg/mL reduced this H2O2-mediated effect significantly (p < 0.05 compared to H2O2 alone; n = 3). The effect of SLT on cleaved caspase-3 protein expression was also determined using Western blotting. It was found that H2O2 caused a significant increase of cleaved capsase-3 expression (n = 3) in EA.hy926 cells, and this effect was completely reversed by SLT at 50 µg/mL (n = 3) (Figure 4B).
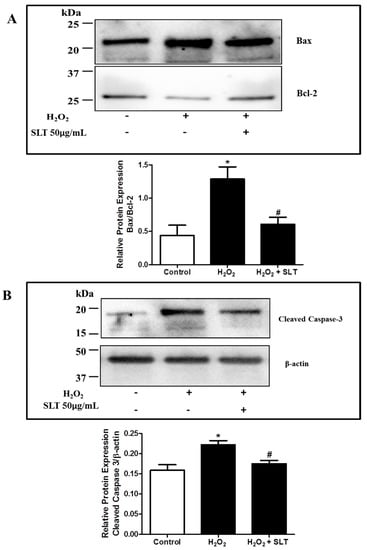
Figure 4.
(A) Effects of SLT (50 µg/mL) on H2O2-upregulated Bax/Bcl-2 ratio in EA.hy926 cells (n = 3). Data are presented as means ± S.D. * p < 0.05 vs. control group; # p < 0.05 vs. H2O2 group. Images are representative of three independent experiments. The present or absent of H2O2 and SLT in the culture is indicated by + or − respectively; (B) Effects of SLT (50 µg/mL) on H2O2-upregulated cleaved caspase-3 expression in EA.hy926 cells (n = 3). Result was expressed as expression of cleaved caspase-3 protein relative to β-actin. Data are presented as means ± S.D. * p < 0.05 vs. control group; # p < 0.05 vs. H2O2 group. Images are representative of three independent experiments. The present or absent of H2O2 and SLT in the culture is indicated by + or − respectively.
3. Discussion
It is well established that endothelial dysfunction caused by elevated cerebrovascular oxidative stress is one of the major mechanisms of CVDs [3,6,30]. Therefore, interventions that can protect endothelial cells from ROS-induced damage would be beneficial for CVDs. In the present study, we demonstrated that SLT could protect EA.hy926 cells from oxidative stress and cell damage caused by H2O2. Furthermore, our results suggest that this effect is possibly mediated by a reduction of the Bax/Bcl-2 ratio and an increase of SOD activity in the EA.hy926 cells.
ROS, such as superoxide anions and hydroxyl radicals, are generated during normal cellular metabolism [31]. Under normal physiological conditions, vascular endothelial cells are in continuous contact with steady-state levels of oxidative metabolites. This constant level of ROS is tightly regulated by a number of anti-oxidative enzymes, such as SOD and glutathione peroxidase [32,33]. However, when this anti-oxidative mechanism is disrupted, excessive ROS will be generated, leading to endothelium dysfunction, which contributes to the development and progression of a number of cerebral and vascular diseases, such as atherosclerosis, stroke, and hypertension [6,34,35]. H2O2 is one of the ROS that have been shown to play a major role in vascular and endothelial dysfunction [3,10,36]. Numerous in vitro studies have demonstrated that high levels of H2O2 can cause significant injury and reduce endothelial cell viability [37,38,39]. In line with this, our results showed that H2O2 (0.5 mM, 24 h) significantly reduced viability and increased LDH leakage in the EA.hy926 cells. Interestingly, pre-incubation of the cells with SLT one hour prior to the addition of H2O2 significantly suppressed the H2O2-mediated cell death and LDH leakage, demonstrating the anti-apoptotic property of SLT.
A recent clinical study demonstrated that SLT can improve cognitive function and memory in people with vascular dementia [18]. Animal studies have suggested these clinically beneficial effects are possibly associated with increases in cerebral blood flow and reductions in platelet aggregation rate and whole blood viscosity [21]. However, the underlying cellular mechanisms of SLT in protecting endothelial cell from ROS-mediated injury had not been explored previously. H2O2 has been shown to induce oxidative stress via an increase in the generation of intracellular ROS in endothelial cells [40]. Our results showed that SLT suppressed the H2O2-induced intracellular ROS generation in a concentration-dependent manner. More importantly, SLT at 50 µg/mL produced a similar effect to our positive control, gallic acid (a known potent anti-oxidant) [29]. In addition, SOD is a major enzyme that protects against oxidative stress damage in endothelial cells [33,41]. In the present study, SLT at 50 µg/mL partly reversed the H2O2-suppressed SOD activity in the EA.hy926 cells. These results highlight the potent anti-oxidant properties of SLT through reduction in intracellular ROS generation and modulation of anti-oxidative enzyme activity.
A number of apoptosis-related proteins, including Bax, Bcl-2, and caspase-3, are required for cellular survival regulation [42]. In this study, we examined if SLT can suppress the apoptotic signalling pathway transduction triggered by H2O2. Our results showed that H2O2 increased the Bax/Bcl-2 ratio; this H2O2-induced effect was inhibited by pre-treatment of SLT. It has been demonstrated that intracellular ROS can increase cytosolic caspases activity via activation of Bax and dissociation of cytochrome C from the inner mitochondrial membrane [43]. Several studies have suggested that Bax/Bcl-2 plays a role in determining cell apoptosis process [44,45]. H2O2 has been shown to downregulate Bcl-2 and upregulate Bax (i.e., increased Bax/Bcl-2 ratio), leading to caspase-3 cleavage, and eventually apoptosis [46]. Caspases-3 is one of the most important enzymes responsible for the cleaving of many cellular substrates during apoptosis [47]. In line with this, we demonstrated that the anti-apoptotic effect of SLT appears to be associated with the inhibition of the apoptotic death cascade via suppression of caspase-3 activation and a reduction in the Bax/Bcl-2 ratio in EA.hy926 cells.
Endothelial dysfunctions are closely associated with vascular dementia and other neurological disorders [9,30]. For instance, changes in brain vascular endothelial cell morphology can reduce the blood-brain barrier permeability, leading to cognitive decline and dementia [48]. Moreover, an increase in ROS generation and oxidative stress in vasculature has been suggested as one of the central pathologies of both vascular dementia and Alzheimer’s disease [49]. Given that current therapies for these diseases are limited [50,51], the development of new therapies/interventions is urgently needed. In this regard, SLT has recently been demonstrated to improve cognitive function and memory in people with vascular dementia. Despite the relatively small sample size in this study, the findings have highlighted the therapeutic potential of SLT in improving cognitive function in people with dementia [18]. It is important to point out that, although some preclinical pharmacokinetic, toxicity, and pharmacodynamics studies of SLT and its individual components have been conducted in several animal models of cerebral disorders [20,21,22,27,52], the cellular and molecular effects of SLT in endothelial cells have not been fully explored. The results of this study demonstrate the effects and underlying signalling mechanisms of SLT against oxidative stress in EA.hy926 cells, providing new insights and molecular evidence to previous in vivo and clinical observations.
The present study has several limitations. Multiple sources of ROS have been suggested to contribute to endothelial cell damage [53,54]. For example, Xie et al. demonstrated the significant role of mitochondrial-derived ROS in age-related cardiovascular diseases [55]. Our current experiments have only examined the global cellular ROS generation in response to exogenous H2O2; more detailed studies, such as direct measurement of mitochondrial ROS, are required to clarify the cellular target and anti-oxidative property of SLT in endothelial cell. Although our results show that the anti-apoptotic property of SLT was associated with reduction in caspase-3 activation and Bax/Bcl-2 ratio, the effects of SLT on other cellular signalling pathways should also be explored using additional experimental models (e.g., heavy metal-induced [56] and hypoxia-induced [57] oxidative stress models). Additionally, contributions of individual components of SLT to the observed effects were not determined.
4. Materials and Methods
4.1. Reagents and Antibodies
SLT extracts were provided in-kind by Shineway Pharmaceutical Group (Shijiazhuang, China). Hydrogen peroxide (H2O2), dimethylsulfoxide (DMSO), and trypan blue were purchased from Sigma-Aldrich (St. Louis, MO, USA). Dulbecco’s Modified Eagle’s Medium Ham’s F-12 (DMEM/F12) (1:1 Mix) with L-Glutamine was purchased from Lonza (Basel, Switzerland). Fetal bovine serum (FBS) and penicillin and streptomycin (PS) were purchased from Gibco Life Technologies (Waltham, MA, USA). 3-(4,5-di-methylthiazol-2-yl) 2,5-diphenyltetrazolium bromide (MTT) was purchased from Astral Scientific (Caringbah, Australia). The superoxide dismutase (SOD) activity assay kit was purchased from Cayman Chemical (Ann Arbor, MI, USA). CytoTox non-radioactive cytotoxicity assay was obtained from Promega Corporation (Madison, WI, USA). The cellular ROS/Superoxide detection assay kit was purchased from Abcam (Cambridge, UK). Anti-Bax, anti-Bcl-2, anti-cleaved caspase-3, and anti-beta actin antibodies were obtained from Santa Cruz Biotechnology (Dallas, TX, USA). All other reagents and chemicals were of chemical analytical grade.
4.2. EA.hy 926 Cell Culture
The permanent human endothelial cell line EA.hy926 was originally derived from a human umbilical vein obtained from ATCC (Manassas, VA, USA). In this study, cells were grown in DMEM/F12 (1:1 Mix) supplemented with 10% fetal bovine serum (FBS), 1% l-glutamine, and 1% penicillin-streptomycin in a humidified atmosphere of 5% CO2 at 37 °C. During cell culture, the medium was changed every three days until the cells reached 90% confluence. To assess the effects of SLT on EA.hy926 cells, the cells were treated with increasing concentrations of SLT (0.1, 1, 10, 50 µg/mL) for 1 h followed by H2O2 (0.5 mM) or vehicle for 24 h unless stated otherwise.
4.3. Measurement of Cell Viability
Cell viability was determined using MTT assay. In brief, cells were seeded in 96-well plates at a density of 1.0 × 105 cells/well and allowed to attach for 24 h. After incubation with the above-mentioned treatments, the culture supernatant was removed, then the cells were incubated with MTT (5 mg/mL) in DMEM/F12 medium at 37 °C for 4 h. After MMT incubation, the culture medium with dye was replaced with 150 µL DMSO and was agitated in a plate shaker for 5 min. Next, the optical density (O.D.) of each well was measured at 560 nm using a Microplate Reader (BMG Labtech, Ortenberg, Germany). Cell viability was expressed as a percentage relative to control.
4.4. Measurement of Intracellular ROS Level
Intracellular ROS level was evaluated using the cellular ROS/superoxide detection assay kit (Abcam, Cambridge, UK) according to the manufacturer’s instructions. In brief, cells were seeded in 96-well plates at a density of 1.0 × 105 cells/well and allowed to attach for 24 h. After incubation with the above-mentioned treatments, the culture supernatant was removed and the cells were washed with 100 µL/well of 1× assay buffer. The ROS specific stain, DCFH-DA, was added to the cells and allowed to incubate in the dark for 60 min. After the incubation, intracellular ROS level was determined using a fluorescence microplate reader (Ex = 488 nm, Em = 520 nm) (BMG Labtech, Ortenberg, Germany).
4.5. Measurement of Lactate Dehydrogenase (LDH) Leakage
Lactate dehydrogenase (LDH) release was evaluated using the non-radioactive assay kit (Promega Corporation, Madison, WI, USA) according to the manufacturer’s instructions. Briefly, cells were seeded in 96-well plates at a density of 1.0 × 105 cells/well and allowed to attach for 24 h. After incubation with the above-mentioned treatments, 50 µL of supernatant per well was transferred to a 96-well flat clear bottom plate. An equal amount of CytoTox reagent was added to each well and allowed to incubate for 30 min at room temperature. A stop solution was added to terminate the reaction at the end of the incubation period. LDH level was measured at 490 nm using a microplate reader (BMG Labtech, Ortenberg, Germany).
4.6. Measurement of Intracellular SOD Activity
Cells were seeded in six-well plates at a density of 1.0 × 106 cells/well and allowed to attach for 24 h. The cells were treated with vehicle, H2O2 (0.5 mM; 24 h) alone or H2O2 (0.5 mM; 24 h) with SLT (50 µg/mL; 1 h prior addition of H2O2). After the treatment, the cells were lysed by Freeze Thaw method three times. The activity of SOD was determined using a commercially available kit, according to the manufacturer’s instructions.
4.7. Western Blotting
Cells were seeded in six-well plates at a density of 1.0 × 106 cells/well and allowed to attach for 24 h. The cells were treated with vehicle, H2O2 (0.5 mM; 24 h) alone or H2O2 (0.5 mM; 24 h) with SLT (50 µg/mL; 1 h prior addition of H2O2). After the treatment, the cells were homogenized and lysed in a Radioimmunoprecipitation assay RIPA buffer (Thermo Scientific, Waltham, MA, USA) in the presence of protease inhibitors (Roche Applied Science, Penzberg, Germany) to obtain protein extracts. Protein concentrations were determined using the bovine serum albumin (BSA) protein assay kit (Pierce, Waltham, MA, USA). Samples (25 µg of protein per lane) were loaded onto a mini-PROTEAN TGXTM precast electrophoresis gel (BioRad, Hercules, CA, USA). After electrophoresis (110 V, 90 min), the separated proteins were transferred to polyvinylidene difluoride (PVDF) membranes using iBlot 2 gel transfer system (Thermofisher, Waltham, MA, USA). Non-specific sites were blocked with 5% non-fat dry milk in Phosphate Buffered Saline Tween-20 (PBSt) for 60 min, and the blots were then incubated with anti-Bax, 1:1000 (Santa Cruz), anti-Bcl-2, 1:1000 (Santa Cruz), anti-cleaved caspase-3, 1:1000 (Santa Cruz, Dallas, TX, USA), and anti-beta actin, 1:10,000 (Santa Cruz) in PBSt overnight at 4 °C. Anti-rabbit horseradish peroxidase (HRP) conjugated immunoglobulin G (IgG), 1:1000 (DakoCytomation, Glostrup, Denmark) in PBSt (60 min, room temperature) was used to detect the binding of its correspondent antibody. β-actin was used to verify equal loading of protein in each lane. The protein expression was detected with Western Lightning Chemiluminescence Reagent Plus (PerkinElmer Life Sciences, Waltham, MA, USA) and quantified by Quantity One (version 4.6.7) software (BioRad).
4.8. Statistical Analysis
Data were presented as mean ± SEM of n experiments. Statistical comparisons were performed using t-test or one-way analysis of variance (ANOVA), where appropriate. Differences were considered to be statistically significant at p < 0.05. All statistical analyses were performed using GraphPad Prism 5 software (GraphPad Software, Inc., La Jolla, CA, USA).
5. Conclusions
In conclusion, our results revealed that SLT can inhibit H2O2-induced endothelial cell injury via the direct reduction of intracellular ROS generation and increase of SOD activity. These protective effects are closely associated with the inhibition of the apoptotic death cascade through suppression of caspase-3 activation and reduction of Bax/Bcl-2 ratio. Our data suggest that SLT possesses potent anti-oxidative and anti-apoptotic activities, which at least partially contribute to its cognitive enhancing effects observed in the clinical study. Further studies are required to investigate the effects and mechanisms of SLT between different cell types within the neurovascular unit and the interaction/synergistic effects between individual components of SLT
Acknowledgments
The authors have received funding support from Ministry of Science and Technology of China, National Natural Science Foundation of China, Academy of China Medical Sciences of China, Shineway Pharmaceutical Group and Western Sydney University. Sai Wang Seto is supported by a Cardiac Health Institute Research Fellowship. Genevieve Z. Steiner is a recipient of a National Health and Medical Research Council (NHMRC)–Australian Research Council (ARC) Dementia Research Development Fellowship (APP1102532). The funding bodies played no role in generation of the data presented in this publication.
Author Contributions
Sai Wang Seto, Dennis Chang, Alan Bensoussan, Hosen Kiat and Jianxun Liu conceived the study. Sai Wang Seto and Wai Man Ko designed the experiments. Sai Wang Seto, Wai Man Ko and Xian Zhou performed the experiments. Sai Wang Seto and Dennis Chang wrote the manuscript. Wai Man Ko, Simon M. Y. Lee, Maggie P. M. Hoi and Genevieve Z. Steiner performed data and statistical analysis. Alan Bensoussan, Hosen Kiat, Jianxun Liu, Simon M. Y. Lee, Maggie P. M. Hoi and Genevieve Z. Steiner edited the manuscript significantly. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Capone, C.; Faraco, G.; Anrather, J.; Zhou, P.; Iadecola, C. Cyclooxygenase 1-derived prostaglandin E2 and EP1 receptors are required for the cerebrovascular dysfunction induced by angiotensin II. Hypertension 2010, 55, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Kwan, Y.W.; Ngai, S.M. Modulatory effect of interleukin-1β on rat isolated basilar artery contraction. Eur. J. Pharmacol. 2006, 531, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, A.; Srivastava, S.; Siow, R.C.; Cash, D.; Modo, M.; Duchen, M.R.; Fraser, P.A.; Williams, S.C.; Mann, G.E. Sulforaphane preconditioning of the Nrf2/HO-1 defense pathway protects the cerebral vasculature against blood-brain barrier disruption and neurological deficits in stroke. Free Radic. Biol. Med. 2013, 65, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Chrissobolis, S.; Drummond, G.R.; Faraci, F.M.; Sobey, C.G. Chronic aldosterone administration causes Nox2-mediated increases in reactive oxygen species production and endothelial dysfunction in the cerebral circulation. J. Hypertens. 2014, 32, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Kaneko, Y.; Tsukamoto, E.; Frankowski, H.; Kouyama, K.; Kita, Y.; Niikura, T.; Aiso, S.; Bredesen, D.E.; Matsuoka, M.; et al. Molecular characterization of neurohybrid cell death induced by Alzheimer’s amyloid-β peptides via p75NTR/PLAIDD. J. Neurochem. 2004, 90, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Poon, C.C.; Seto, S.W.; Au, A.L.; Zhang, Q.; Li, R.W.; Lee, W.Y.; Leung, G.P.; Kong, S.K.; Yeung, J.H.; Ngai, S.M.; et al. Mitochondrial monoamine oxidase-A-mediated hydrogen peroxide generation enhances 5-hydroxytryptamine-induced contraction of rat basilar artery. Br. J. Pharmacol. 2010, 161, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.L.; Hou, Y.C.; Chao, P.D.; Weng, C.S.; Ho, F.M. The antioxidant effects of quercetin metabolites on the prevention of high glucose-induced apoptosis of human umbilical vein endothelial cells. Br. J. Nutr. 2009, 101, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Guo, S. Using zebrafish to assess the impact of drugs on neural development and function. Expert Opin. Drug Discov. 2009, 4, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Chang, D.; Jenkins, A.; Bensoussan, A.; Kiat, H. Angiogenesis in Ischemic Stroke and Angiogenic Effects of Chinese Herbal Medicine. J. Clin. Med. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.; Rodriguez-Yanez, M.; Sobrino, T.; Leira, R.; Castillo, J. Platelets, inflammation, and atherothrombotic neurovascular disease: The role of endothelial dysfunction. Cerebrovasc. Dis. 2005, 20, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Sorriento, D.; Santulli, G.; del Giudice, C.; Anastasio, A.; Trimarco, B.; Iaccarino, G. Endothelial cells are able to synthesize and release catecholamines both in vitro and in vivo. Hypertension 2012, 60, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G. MicroRNAs and Endothelial (Dys) Function. J. Cell. Physiol. 2016, 231, 1638–1644. [Google Scholar] [CrossRef] [PubMed]
- Quesada, I.M.; Lucero, A.; Amaya, C.; Meijles, D.N.; Cifuentes, M.E.; Pagano, P.J.; Castro, C. Selective inactivation of NADPH oxidase 2 causes regression of vascularization and the size and stability of atherosclerotic plaques. Atherosclerosis 2015, 242, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fotheringham, L.; Wittchen, E.S.; Hartnett, M.E. Rap1 GTPase Inhibits Tumor Necrosis Factor-alpha-Induced Choroidal Endothelial Migration via NADPH Oxidase- and NF-κB-Dependent Activation of Rac1. Am. J. Pathol. 2015, 185, 3316–3325. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.C.; Gao, X.F.; Ruan, C.; Ge, Z.R.; Lu, J.D.; Zhang, J.J.; Zhang, Y.; Wang, L.; Shi, H.M. miR-103 Regulates Oxidative Stress by Targeting the BCL2/Adenovirus E1B 19 kDa Interacting Protein 3 in HUVECs. Oxidative Med. Cell. Longev. 2015, 2015, 489647. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.; Wong, C.Y.; Wan, C.W.; Kwok, C.Y.; Wu, J.H.; Ng, K.M.; So, C.H.; Au, A.L.; Poon, C.C.; Seto, S.W.; et al. Evaluation of anti-oxidant capacity of root of Scutellaria baicalensis Georgi, in comparison with roots of Polygonum multiflorum Thunb and Panax ginseng CA Meyer. Am. J. Chin. Med. 2010, 38, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, F.; Wei, C.; Song, H.; Wu, L.; Tang, Y.; Jia, J. Rationale and design of a multicenter, phase 2 clinical trial to investigate the efficacy of traditional Chinese medicine SaiLuoTong in vascular dementia. J. Stroke Cerebrovasc. Dis. 2014, 23, 2626–2634. [Google Scholar] [CrossRef] [PubMed]
- Steiner, G.Z.; Yeung, A.; Liu, J.X.; Camfield, D.A.; Blasio, F.M.; Pipingas, A.; Scholey, A.B.; Stough, C.; Chang, D.H. The effect of Sailuotong (SLT) on neurocognitive and cardiovascular function in healthy adults: A randomised, double-blind, placebo controlled crossover pilot trial. BMC Complement. Altern. Med. 2016, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.L.; Cong, W.; Wang, J.; Zhang, Y. SLT for Vascular Dementia; China Academy of Chinese Medical Sciences: Beijing, China, 2008. [Google Scholar]
- Xu, L.; Liu, J.X.; Cong, W.H.; Wei, C.E. Effects of Weinaokang capsule on intracephalic cholinergic system and capability of scavenging free radicas in chronic cerebral hypoperfusion rats. Zhongguo Zhong Yao Za Zhi 2008, 33, 531–534. [Google Scholar] [PubMed]
- Liu, J. Development of an Evidence-Based Chinese Herbal Medicine for the Management of Vascular Dementia. Ph.D. Thesis, The University of Western Sydney, Sydney, Australia, 2008. [Google Scholar]
- Zheng, Y.Q.; Liu, J.X.; Wang, J.N.; Xu, L. Effects of crocin on reperfusion-induced oxidative/nitrative injury to cerebral microvessels after global cerebral ischemia. Brain Res. 2007, 1138, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zheng, L.; Lu, S.; Yang, Y. Neuroprotective effects of pretreatment of ginsenoside Rb1 on severe cerebral ischemia-induced injuries in aged mice: Involvement of anti-oxidant signaling. Geriatr. Gerontol. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Nie, B.; Fu, S.; Hu, J.; Yin, L.; Lin, L.; Wang, X.; Lu, P.; Xu, X.M. EGb761 protects hydrogen peroxide-induced death of spinal cord neurons through inhibition of intracellular ROS production and modulation of apoptotic regulating genes. J. Mol. Neurosci. 2009, 38, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.C.; Du, G.H.; Zhang, J.T. Protective effect of ginkgo biloba extract on endothelial cell against damage induced by oxidative stress. J. Cardiovasc. Pharmacol. 2002, 40, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Sung, J.H.; Cho, E.H.; Won, C.K.; Lee, H.J.; Kim, M.O.; Koh, P.O. Gingko biloba Extract (EGb 761) prevents ischemic brain injury by activation of the Akt signaling pathway. Am. J. Chin. Med. 2009, 37, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Q.; Liu, J.X.; Li, X.Z.; Xu, L. Effects and mechanism of Weinaokang on reperfusion-induced vascular injury to cerebral microvessels after global cerebral ischemia. Chin. J. Integr. Med. 2010, 16, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Chang, D.; Ko, W.M.; Kiat, H.; Bensoussan, A.; Lee, S.M.Y.; Hoi, M.P.M.; Liu, J. Angiogenic effects of Sailuotong (SLT) on EA.hy926 cell in vitro and zebrafish in vivo. Int. J. Mol. Sci. 2017. manuscript under preparation. [Google Scholar]
- Chen, C.H.; Liu, T.Z.; Chen, C.H.; Wong, C.H.; Chen, C.H.; Lu, F.J.; Chen, S.C. The efficacy of protective effects of tannic acid, gallic acid, ellagic acid, and propyl gallate against hydrogen peroxide-induced oxidative stress and DNA damages in IMR-90 cells. Mol. Nutr. Food Res. 2007, 51, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.W.; Yang, G.Y.; Kiat, H.; Bensoussan, A.; Kwan, Y.W.; Chang, D. Diabetes Mellitus, Cognitive Impairment, and Traditional Chinese Medicine. Int. J. Endocrinol. 2015, 2015, 810439. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, E.A.; Mahmoud, A.M.; Khalifa, A.M.; Ali, S.S. Physiological and pathophysiological reactive oxygen species as probed by EPR spectroscopy: The underutilized research window on muscle ageing. J. Physiol. 2016, 594, 4591–4613. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U. Nitric oxide and oxidative stress in vascular disease. Pflugers Arch. 2010, 459, 923–939. [Google Scholar] [CrossRef] [PubMed]
- Spanier, G.; Xu, H.; Xia, N.; Tobias, S.; Deng, S.; Wojnowski, L.; Forstermann, U.; Li, H. Resveratrol reduces endothelial oxidative stress by modulating the gene expression of superoxide dismutase 1 (SOD1), glutathione peroxidase 1 (GPx1) and NADPH oxidase subunit (Nox4). J. Physiol. Pharmacol. 2009, 60, 111–116. [Google Scholar] [PubMed]
- Hermann, C.; Zeiher, A.M.; Dimmeler, S. Shear stress inhibits H2O2-induced apoptosis of human endothelial cells by modulation of the glutathione redox cycle and nitric oxide synthase. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 3588–3592. [Google Scholar] [CrossRef] [PubMed]
- Aliev, G.; Palacios, H.H.; Walrafen, B.; Lipsitt, A.E.; Obrenovich, M.E.; Morales, L. Brain mitochondria as a primary target in the development of treatment strategies for Alzheimer disease. Int. J. Biochem. Cell Biol. 2009, 41, 1989–2004. [Google Scholar] [CrossRef] [PubMed]
- Breton-Romero, R.; Lamas, S. Hydrogen peroxide signaling in vascular endothelial cells. Redox Biol. 2014, 2, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Asahina, T.; Kashiwagi, A.; Nishio, Y.; Ikebuchi, M.; Harada, N.; Tanaka, Y.; Takagi, Y.; Saeki, Y.; Kikkawa, R.; Shigeta, Y. Impaired activation of glucose oxidation and NADPH supply in human endothelial cells exposed to H2O2 in high-glucose medium. Diabetes 1995, 44, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Belkhiri, A.; Richards, C.; Whaley, M.; McQueen, S.A.; Orr, F.W. Increased expression of activated matrix metalloproteinase-2 by human endothelial cells after sublethal H2O2 exposure. Lab. Investig. 1997, 77, 533–539. [Google Scholar] [PubMed]
- Jia, L.Q.; Yang, G.L.; Ren, L.; Chen, W.N.; Feng, J.Y.; Cao, Y.; Zhang, L.; Li, X.T.; Lei, P. Tanshinone IIA reduces apoptosis induced by hydrogen peroxide in the human endothelium-derived EA.hy926 cells. J. Ethnopharmacol. 2012, 143, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Jiang, F.; Wang, B.; Yu, Y.; Zhang, X.; Yin, Z.; Liu, C. Ophiopogonin D prevents H2O2-induced injury in primary human umbilical vein endothelial cells. J. Ethnopharmacol. 2010, 128, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Giray, B.; Kan, E.; Bali, M.; Hincal, F.; Basaran, N. The effect of vitamin E supplementation on antioxidant enzyme activities and lipid peroxidation levels in hemodialysis patients. Clin. Chim. Acta 2003, 338, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.L.; Kang, Y.; Li, J.K.; Li, Q.S.; Liu, E.L.; Liu, X.X. Protective effects of hyperoside against H2O2-induced apoptosis in human umbilical vein endothelial cells. Mol. Med. Rep. 2016, 14, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Buccellato, L.J.; Tso, M.; Akinci, O.I.; Chandel, N.S.; Budinger, G.R. Reactive oxygen species are required for hyperoxia-induced Bax activation and cell death in alveolar epithelial cells. J. Biol. Chem. 2004, 279, 6753–6760. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Afaq, F.; Mukhtar, H. Involvement of nuclear factor-κB, Bax and Bcl-2 in induction of cell cycle arrest and apoptosis by apigenin in human prostate carcinoma cells. Oncogene 2002, 21, 3727–3738. [Google Scholar] [CrossRef] [PubMed]
- Vogel, M.W. Cell death, Bcl-2, Bax, and the cerebellum. Cerebellum 2002, 1, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kang, J.S.; Park, J.H.; Lee, Y.J.; Choi, J.S.; Kang, Y.H. Polyphenolic flavonoids differ in their antiapoptotic efficacy in hydrogen peroxide-treated human vascular endothelial cells. J. Nutr. 2003, 133, 985–991. [Google Scholar] [PubMed]
- Fuentes-Prior, P.; Salvesen, G.S. The protein structures that shape caspase activity, specificity, activation and inhibition. Biochem. J. 2004, 384, 201–232. [Google Scholar] [CrossRef] [PubMed]
- Acharya, N.K.; Goldwaser, E.L.; Forsberg, M.M.; Godsey, G.A.; Johnson, C.A.; Sarkar, A.; DeMarshall, C.; Kosciuk, M.C.; Dash, J.M.; Hale, C.P.; et al. Sevoflurane and Isoflurane induce structural changes in brain vascular endothelial cells and increase blood-brain barrier permeability: Possible link to postoperative delirium and cognitive decline. Brain Res. 2015, 1620, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Bennett, S.; Grant, M.M.; Aldred, S. Oxidative stress in vascular dementia and Alzheimer’s disease: A common pathology. J. Alzheimers Dis. 2009, 17, 245–257. [Google Scholar] [PubMed]
- Baskys, A.; Hou, A.C. Vascular dementia: Pharmacological treatment approaches and perspectives. Clin. Interv. Aging 2007, 2, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Kalaria, R.N.; Corbett, A.; Ballard, C. Update on Vascular Dementia. J. Geriatr. Psychiatry Neurol. 2016, 29, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Cong, W.H.; Liu, J.X.; Xu, L. Effects of extracts of Ginseng and Ginkgo biloba on hippocampal acetylcholine and monoamines in PDAP-pV717I transgenic mice. Zhongguo Zhong Xi Yi Jie He Za Zhi 2007, 27, 810–813. [Google Scholar] [PubMed]
- Coelho-Santos, V.; Socodato, R.; Portugal, C.; Leitao, R.A.; Rito, M.; Barbosa, M.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Minshall, R.D.; et al. Methylphenidate-triggered ROS generation promotes caveolae-mediated transcytosis via Rac1 signaling and c-Src-dependent caveolin-1 phosphorylation in human brain endothelial cells. Cell. Mol. Life Sci. 2016, 73, 4701–4716. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Lopez, I.; Diaz-Morales, N.; Rovira-Llopis, S.; de Maranon, A.M.; Orden, S.; Alvarez, A.; Banuls, C.; Rocha, M.; Murphy, M.P.; Hernandez-Mijares, A.; et al. The mitochondria-targeted antioxidant MitoQ modulates oxidative stress, inflammation and leukocyte-endothelium interactions in leukocytes isolated from type 2 diabetic patients. Redox Biol. 2016, 10, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Santulli, G.; Reiken, S.R.; Yuan, Q.; Osborne, B.W.; Chen, B.X.; Marks, A.R. Mitochondrial oxidative stress promotes atrial fibrillation. Sci. Rep. 2015, 5, 11427. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.; Mittal, M.; Mehta, A. Heavy metal induced oxidative stress & its possible reversal by chelation therapy. Indian J. Med. Res. 2008, 128, 501–523. [Google Scholar] [PubMed]
- Liu, B.; Ren, K.D.; Peng, J.J.; Li, T.; Luo, X.J.; Fan, C.; Yang, J.F.; Peng, J. Suppression of NADPH oxidase attenuates hypoxia-induced dysfunctions of endothelial progenitor cells. Biochem. Biophys. Res. Commun. 2016. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).