
A Holistic Approach to Managing Microalgae for Biofuel Applications

 ,
, 


Abstract
:
1. Introduction
2. Microalgae as the Third Generation Feedstock
2.1. Microalgal Cultivation for Biofuel Production
2.1.1. Effect of Light Intensity on Microalgal Cultivation
2.1.2. Effect of Temperature on Microalgal Cultivation
2.1.3. Effect of Nutrients on Microalgal Cultivation
2.1.4. Effect of CO2 Source on Microalgal Cultivation
2.2. Microalgal Cultivation Methods: Open Systems and PBRs
2.2.1. Open Pond System
2.2.2. PBRs
Stirred-Tank PBRs
Vertical Column PBRs
Horizontal Tubular PBRs
Flat Panel PBRs
2.2.3. Hybrid System
2.2.4. Heterotrophic (Fermenter) System
2.2.5. Mixotrophic System
2.3. Potential Applications of Microalgae
2.3.1. CO2 Sequestration
2.3.2. Wastewater Treatment
2.3.3. Other Potential Applications of Microalgae—Food, Animal Feed, Cosmetics and Fertilizer
3. Microalgae Harvesting
3.1. Bulk Harvesting
3.1.1. Flocculation
3.1.2. Flotation
3.1.3. Gravity Sedimentation
3.2. Concentration
3.2.1. Centrifugation
3.2.2. Ultrasonic Aggregation
3.2.3. Filtration
3.2.4. Electrophoresis
4. Conversion of Microalgae to Biofuel
4.1. Thermochemical Processes
4.1.1. Gasification
4.1.2. Thermochemical Liquefaction
4.1.3. Pyrolysis
4.1.4. Direct Combustion
4.2. Biochemical Conversion
4.2.1. Anaerobic Digestion (AD)
4.2.2. Alcoholic Fermentation
4.2.3. Photobiological H2 Production
5. Converting MAB to Biodiesel
6. Prospects and Challenges
Acknowledgments
Conflicts of Interest
References
- Longwell, J.P.; Rubin, E.S.; Wilson, J. Coal: Energy for the future. Prog. Energy Combust. 1995, 21, 269–360. [Google Scholar] [CrossRef]
- Petroleum, B. Statistical Review of World Energy 2007; BP: London, UK, 2009. [Google Scholar]
- Sayre, R. Microalgae: The potential for carbon capture. Bioscience 2010, 60, 722–727. [Google Scholar] [CrossRef]
- De La Torre Ugarte, G.D. The Economic Impacts of Bioenergy Crop Production on US Agriculture; United States Department of Agriculture: Washington, DC, USA, 2000.
- Judkins, R.R.; Fulkerson, W.; Sanghvi, M.K. The dilemma of fossil-fuel use and global climate change. Energy Fuel 1993, 7, 14–22. [Google Scholar] [CrossRef]
- Ma, C.H.; You, K.; Ji, D.C.; Ma, W.W.; Li, F.Q. Primary discussion of a carbon sink in the oceans. J. Ocean Univ. China 2015, 14, 284–292. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Winkelmann, R.; Levermann, A.; Ridgwell, A.; Caldeira, K. Combustion of available fossil fuel resources sufficient to eliminate the Antarctic Ice Sheet. Sci. Adv. 2015, 1, e1500589. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.; Masek, J.G. The vegetation greenness trend in Canada and US Alaska from 1984–2012 Landsat data. Remote Sens. Environ. 2016, 176, 1–16. [Google Scholar] [CrossRef]
- Bilanovic, D.; Andargatchew, A.; Kroeger, T.; Shelef, G. Freshwater and marine microalgae sequestering of CO2 at different C and N concentrations-response surface methodology analysis. Energy Convers. Manag. 2009, 50, 262–267. [Google Scholar] [CrossRef]
- Outlook, I.E. World Energy Outlook 2007: China and India Insights, International Energy Agency; OECD/IEA: Paris, France, 2007. [Google Scholar]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO2 bio-mitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Ghayal, M.S.; Pandya, M.T. Microalgae biomass: A renewable source of energy. Energy Procedia 2013, 32, 242–250. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wang, B.; Wu, N.; Lan, C.Q. Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl. Microbiol. Biotechnol. 2008, 81, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E. Phycology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Vasudevan, P.T.; Briggs, M. Biodiesel production—Current state of the art and challenges. J. Ind. Microbiol. Biotechnol. 2008, 35, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Bolton, J.R.; Hall, D.O. The maximum efficiency of photosynthesis. Photochem. Photobiol. 1991, 53, 545–548. [Google Scholar] [CrossRef]
- Doucha, J.; Lívanský, K. Outdoor open thin-layer microalgal photobioreactor: Potential productivity. J. Appl. Phycol. 2009, 21, 111–117. [Google Scholar] [CrossRef]
- Hase, R.; Oikawa, H.; Sasao, C.; Morita, M.; Watanabe, Y. Photosynthetic production of microalgal biomass in a raceway system under greenhouse conditions in Sendai city. J. Biosci. Bioeng. 2000, 89, 157–163. [Google Scholar] [CrossRef]
- Morita, M.; Watanabe, Y.; Saiki, H. Photosynthetic productivity of conical helical tubular photobioreactor incorporating Chlorella sorokiniana under field conditions. Biotechnol. Bioeng. 2002, 77, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.O.; Fernández, F.A.; Guerrero, E.C.; Rao, K.K.; Grima, E.M. Outdoor helical tubular photobioreactors for microalgal production: Modeling of fluid-dynamics and mass transfer and assessment of biomass productivity. Biotechnol. Bioeng. 2003, 82, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, F.; Camacho, F.G.; Perez, J.; Sevilla, J.; Grima, E.M. Modeling of biomass productivity in tubular photobioreactors for microalgal cultures: Effects of dilution rate, tube diameter, and solar irradiance. Biotechnol. Bioeng. 1998, 58, 605–616. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, J.; Dunahay, T.; Benemann, J.; Roessler, P. A Look Back at the US Department of Energy’s Aquatic Species Program: Biodiesel from Algae; National Renewable Energy Laboratory: Golden, CO, USA, 1998. [Google Scholar]
- PSchenk, M.; Thomas-Hall, S.R.; Stephens, E.; Marx, U.C.; Mussgnug, J.H.; Posten, C.; Kruse, O.; Hankamer, B. Second generation biofuels: High-efficiency microalgae for biodiesel production. Bioenergy Res. 2008, 1, 20–43. [Google Scholar] [CrossRef]
- Dismukes, G.C.; Carrieri, D.; Bennette, N.; Ananyev, G.M.; Posewitz, M.C. Aquatic phototrophs: Efficient alternatives to land-based crops for biofuels. Curr. Opin. Biotechnol. 2008, 19, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Change, L.-U. Use of US croplands for biofuels increases. Science 2008, 1151861, 319. [Google Scholar]
- De Morais, M.G.; Costa, J.A.V. Isolation and selection of microalgae from coal fired thermoelectric power plant for biofixation of carbon dioxide. Energy Convers. Manag. 2007, 48, 2169–2173. [Google Scholar] [CrossRef]
- Yoo, C.; Jun, S.-Y.; Lee, J.-Y.; Ahn, C.-Y.; Oh, H.-M. Selection of microalgae for lipid production under high levels carbon dioxide. Bioresour. Technol. 2010, 101, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Scheibenbogen, K. Photobioreactors: Design and performance with respect to light energy input. In Bioprocess and Algae Reactor Technology, Apoptosis; Springer: Berlin, Germany, 1998; pp. 123–152. [Google Scholar]
- Chang, J.S.; Show, P.L.; Ling, T.C.; Chen, C.Y.; Ho, S.H.; Tan, C.H.; Nagarajan, D.; Phong, W.N. Photobioreactors. In Current Developments in Biotechnology and Bioengineering; Accepted Proof, in Press; Elsevier: Amsterdam, The Netherlands, 2016; Chapter 11; Volume 6. [Google Scholar]
- Falkowski, P.G.; Chen, Y.-B. Photoacclimation of Light Harvesting Systems in Eukaryotic Algae. In Light harvesting Antennas in Photosynthesis; Green, B.R., Parson, W.W., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 423–447. [Google Scholar]
- Cheirsilp, B.; Torpee, S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed batch cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-H.; Chen, C.-Y.; Chang, J.-S. Effect of light intensity and nitrogen starvation on CO2 fixation and lipid/carbohydrate production of an indigenous microalga Scenedesmus obliquus CNW-N. Bioresour. Technol. 2012, 113, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yuan, C.; Hu, G.; Li, F. Effects of light intensity on the growth and lipid accumulation of microalga Scenedesmus sp. 11-1 under nitrogen limitation. Appl. Biochem. Biotechnol. 2012, 166, 2127–2137. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cao, Y.; Xu, H.; Liu, Y.; Sun, J.; Qiao, D.; Cao, Y. Effect of nitrogen-starvation, light intensity and iron on triacylglyceride/carbohydrate production and fatty acid profile of Neochloris oleoabundans HK-129 by a two-stage process. Bioresour. Technol. 2014, 155, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Mao, X.; Zhou, W.; Guarnieri, M.T. Simultaneous production of triacylglycerol and high-value carotenoids by the astaxanthin-producing oleaginous green microalga Chlorella zofingiensis. Bioresour. Technol. 2016, 214, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Muller-Feuga, A.; le Guédes, R.; Hervé, A.; Durand, P. Comparison of artificial light photobioreactors and other production systems using Porphyridium cruentum. J. Appl. Phycol. 1998, 10, 83–90. [Google Scholar] [CrossRef]
- Ras, M.; Steyer, J.P.; Bernard, O. Temperature effect on microalgae: A crucial factor for outdoor production. Rev. Environ. Sci. Biotechnol. 2013, 12, 153–164. [Google Scholar] [CrossRef]
- Gonçalves, A.L.; Pires, J.C.M.; Simões, M. The effects of light and temperature on microalgal growth and nutrient removal: An experimental and mathematical approach. RSC Adv. 2016, 6, 22896–22907. [Google Scholar] [CrossRef]
- Kessler, E. Upper limits of temperature for growth in Chlorella. Plant Syst. Evol. 1985, 151, 67–71. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Xin, L.; Hu, H.-Y.; Zhang, Y.-P. Growth and lipid accumulation properties of a freshwater microalga Scenedesmus sp. under different cultivation temperature. Bioresour. Technol. 2011, 102, 3098–3102. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, L.; Yamada, H.; Katsuda, T.; Yamaji, H. Effects of temperature on the astaxanthin productivity and light harvesting characteristics of the green alga Haematococcus pluvialis. J. Biosci. Bioeng. 2015, 119, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.; Zhang, J.; Hou, D.; Fan, J.; Li, Y.; Huang, J.; Wang, J. The effect of temperature on cell growth and astaxanthin accumulation of Haematococcus pluvialis during a light-dark cyclic cultivation. Bioresour. Technol. 2014, 167, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A review on the assessment of stress conditions for simultaneous production of microalgal lipids and carotenoids. Front. Microbiol. 2016, 7, 546. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.-M.; Jiang, Y.; Chen, F. High-yield production of lutein by the green microalga Chlorella protothecoides in heterotrophic fed-batch culture. Biotechnol. Prog. 2002, 18, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.F.; Fernández-Sevilla, J.M.; Acién, F.G.; Cerón, M.C.; Pérez-Parra, J.; Molina-Grima, E. Biomass and lutein productivity of Scenedesmus almeriensis: Influence of irradiance, dilution rate and temperature. Appl. Microbiol. Biotechnol. 2008, 79, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Welsh, D.T.; Bartoli, M.; Nizzoli, D.; Castaldelli, G.; Riou, S.A.; Viaroli, P. Denitrification, nitrogen fixation, community primary productivity and inorganic-N and oxygen fluxes in an intertidal Zostera noltii meadow. Mar. Ecol. Prog. Ser. 2000, 208, 65–77. [Google Scholar] [CrossRef]
- Moreno, J.; Vargas, M.Á.; Guerrero, M.G. Outdoor cultivation of a nitrogen-fixing marine cyanobacterium, Anabaena sp. ATCC 33047. Biomol. Eng. 2003, 20, 191–197. [Google Scholar] [CrossRef]
- Hsieh, C.-H.; Wu, W.-T. Cultivation of microalgae for oil production with a cultivation strategy of urea limitation. Bioresour. Technol. 2009, 100, 3921–3926. [Google Scholar] [CrossRef] [PubMed]
- Widjaja, A.; Chien, C.-C.; Ju, Y.-H. Study of increasing lipid production from fresh water microalgae Chlorella vulgaris. J. Taiwan Inst. Chem. Eng. 2009, 40, 13–20. [Google Scholar] [CrossRef]
- Breuer, G.; Packo, P.L.; Dirk, E.M.; Draaisma, R.B.; Wijffels, R.H. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains. Bioresour. Technol. 2012, 124, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wang, Y.; Huang, W.; Xu, J.; Wang, Z.; Xu, J.; Yuan, Z. Enhanced accumulation of carbohydrate and starch in Chlorella zofingiensis induced by nitrogen starvation. Appl. Biochem. Biotechnol. 2014, 174, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.; Zhang, Q.; Gu, M.; Cong, W. Effect of phosphorus on lipid accumulation in freshwater microalga Chlorella sp. J. Appl. Phycol. 2013, 25, 311–318. [Google Scholar] [CrossRef]
- Martin-Jézéquel, V.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolism in diatoms: Implications for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Van den Hende, S.; Vervaeren, H.; Saveyn, H.; Maes, G.; Boon, N. Microalgal bacterial floc properties are improved by a balanced inorganic/organic carbon ratio. Biotechnol. Bioeng. 2011, 108, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Rösch, C.; Skarka, J.; Wegerer, N. Materials flow modeling of nutrient recycling in biodiesel production from microalgae. Bioresour. Technol. 2012, 107, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Morweiser, M.; Kruse, O.; Hankamer, B.; Posten, C. Developments and perspectives of photobioreactors for biofuel production. Appl. Microbiol. Biotechnol. 2010, 87, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Spalding, M.H. Microalgal carbon-dioxide-concentrating mechanisms: Chlamydomonas inorganic carbon transporters. J. Exp. Bot. 2008, 59, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Kunjapur, A.M.; Eldridge, R.B. Photobioreactor design for commercial biofuel production from microalgae. Ind. Eng. Chem. Res. 2010, 49, 3516–3526. [Google Scholar] [CrossRef]
- Fan, J.; Xu, H.; Luo, Y.; Wan, M.; Huang, J.; Wang, W.; Li, Y. Impacts of CO2 concentration on growth, lipid accumulation, and carbon-concentrating-mechanism-related gene expression in oleaginous Chlorella. Appl. Microbiol. Biotechnol. 2015, 99, 2451–2462. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Han, W.; Li, P.; Miao, X.; Zhong, J. CO2 biofixation and fatty acid composition of Scenedesmus obliquus and Chlorella pyrenoidosa in response to different CO2 levels. Bioresour. Technol. 2011, 102, 3071–3076. [Google Scholar] [CrossRef] [PubMed]
- Varshney, P.; Sohoni, S.; Wangikar, P.P.; Bradall, J. Effect of high CO2 concentrations on the growth and macromolecular composition of a heat- and high-light-tolerant microalga. J. Appl. Phycol. 2016, 28, 2631–2640. [Google Scholar] [CrossRef]
- Chiu, S.-Y.; Kao, C.-Y.; Tsai, M.-T.; Ong, S.-C.; Chen, C.-H.; Lin, C.-S. Lipid accumulation and CO2 utilization of Nannochloropsis oculata in response to CO2 aeration. Bioresour. Technol. 2009, 100, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M. Uptake of carbon dioxide from flue gas by microalgae. Energy Convers. Manag. 1996, 37, 1363–1367. [Google Scholar] [CrossRef]
- Hsueh, H.; Chu, H.; Yu, S.T. A batch study on the bio-fixation of carbon dioxide in the absorbed solution from a chemical wet scrubber by hot spring and marine algae. Chemosphere 2007, 66, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Vunjak-Novakovic, G.; Kim, Y.; Wu, X.; Berzin, I.; Merchuk, J.C. Air-lift bioreactors for algal growth on flue gas: Mathematical modeling and pilot-plant studies. Ind. Eng. Chem. Res. 2005, 44, 6154–6163. [Google Scholar] [CrossRef]
- Yeh, K.L.; Chang, J.S.; Chen, W.M. Effect of light supply and carbon source on cell growth and cellular composition of a newly isolated microalga Chlorella vulgaris ESP-31. Eng. Life Sci. 2010, 10, 201–208. [Google Scholar] [CrossRef]
- Chi, Z.; Xie, Y.; Elloy, F.; Zheng, Y.; Hu, Y.; Chen, S. Bicarbonate-based integrated carbon capture and algae production system with alkalihalophilic cyanobacterium. Bioresour. Technol. 2013, 133, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, H.; Wu, Q. Large-scale biodiesel production from microalga Chlorella protothecoides through heterotrophic cultivation in bioreactors. Biotechnol. Bioeng. 2007, 98, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Miao, X.; Wu, Q. High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J. Biotechnol. 2006, 126, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Commercial production of microalgae: Ponds, tanks, tubes and fermenters. J. Biotechnol. 1999, 70, 313–321. [Google Scholar] [CrossRef]
- Setlik, I.; Veladimir, S.; Malek, I. Dual purpose open circulation units for large scale culture of algae in temperate zones. I, Basic design considerations and scheme of a pilot plant. Algol. Stud. 1970, 1, 111–164. [Google Scholar]
- Weissman, J.; Tillett, D. Design and operation of an outdoor microalgae test facility: Large-scale system results. In Aquatic Species Project Report, FY; National Renewable Energy Laboratory: Golden, CO, USA, 1989; Volume 90, pp. 32–56. [Google Scholar]
- Chisti, Y. Biodiesel from microalgae beats bioethanol. Trends Biotechnol. 2008, 26, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Ugwu, C.; Aoyagi, H.; Uchiyama, H. Photobioreactors for mass cultivation of algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-K. Microalgal mass culture systems and methods: Their limitation and potential. J. Appl. Phycol. 2001, 13, 307–315. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Fernández, F.A.; Sevilla, J.F.; Grima, E.M. Photobioreactors for the production of microalgae. Rev. Environ. Sci. Biotechnol. 2013, 12, 131–151. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Meireles, L.A.; Malcata, F.X. Microalgal reactors: A review of enclosed system designs and performances. Biotechnol. Prog. 2006, 22, 1490–1506. [Google Scholar] [CrossRef] [PubMed]
- Suh, I.S.; Lee, C.G. Photobioreactor engineering: Design and performance. Biotechnol. Bioprocess Eng. 2003, 8, 313–321. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, S. Development of suitable photobioreactor for algae production—A review. Renew. Sustain. Energy Rev. 2012, 16, 2347–2353. [Google Scholar] [CrossRef]
- Kumar, K.; Dasgupta, C.N.; Nayak, B.; Lindblad, P.; Das, D. Development of suitable photobioreactors for CO2 sequestration addressing global warming using green algae and cyanobacteria. Bioresour. Technol. 2011, 102, 4945–4953. [Google Scholar] [CrossRef] [PubMed]
- Ogbonna, J.; Yada, H.; Masui, H.; Tanaka, H. A novel internally illuminated stirred tank photobioreactor for large-scale cultivation of photosynthetic cells. J. Ferment. Bioeng. 1996, 82, 61–67. [Google Scholar] [CrossRef]
- Miron, A.S.; Gomez, A.C.; Camacho, F.G.; Grima, E.M.; Chisti, Y. Comparative evaluation of compact photobioreactors for large-scale monoculture of microalgae. J. Biotechnol. 1999, 70, 249–270. [Google Scholar] [CrossRef]
- Gupta, P.L.; Lee, S.M.; Choi, H.J. A mini review: Photobioreactors for large scale algal cultivation. World J. Microbiol. Biotechnol. 2015, 31, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Loubière, K.; Olivo, E.; Bougaran, G.; Pruvost, J.; Robert, R.; Legrand, J. A new photobioreactor for continuous microalgal production in hatcheries based on external-loop airlift and swirling flow. Biotechnol. Bioeng. 2009, 102, 132–147. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.J.; Janssen, M.; Ham, N.; Tramper, J.; Wijffels, R.H. Microalgae cultivation in air-lift reactors: Modeling biomass yield and growth rate as a function of mixing frequency. Biotechnol. Bioeng. 2003, 82, 179. [Google Scholar] [CrossRef] [PubMed]
- Posten, C. Design principles of photo-bioreactors for cultivation of microalgae. Eng. Life Sci. 2009, 9, 165–177. [Google Scholar] [CrossRef]
- Huntley, M.E.; Redalje, D.G. CO2 mitigation and renewable oil from photosynthetic microbes: A new appraisal. Mitig. Adapt. Strategy Glob. Chang. 2007, 12, 573–608. [Google Scholar] [CrossRef]
- Eriksen, N.T. Production of phycocyanin—A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q.; Chen, F. Growing phototrophic cells without light. Biotechnol. Lett. 2006, 28, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Wu, Q. Biodiesel production from heterotrophic microalgal oil. Bioresour. Technol. 2006, 97, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Graverholt, O.S.; Eriksen, N.T. Heterotrophic high-cell-density fed-batch and continuous-flow cultures of Galdieria sulphuraria and production of phycocyanin. Appl. Microbiol. Biotechnol. 2007, 77, 69–75. [Google Scholar] [CrossRef] [PubMed]
- De Swaaf, M.E.; Sijtsma, L.; Pronk, J.T. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing marine alga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-W.; Zhang, Y.-M.; Chen, F. Application of mathematical models to the determination optimal glucose concentration and light intensity for mixotrophic culture of Spirulina platensis. Process Biochem. 1999, 34, 477–481. [Google Scholar] [CrossRef]
- Graham, L.E.; Graham, J.M.; Wilcox, L.W. Algae, 2nd ed.; Pearson/Benjamin Cummings: San Francisco, CA, USA, 2009. [Google Scholar]
- Andrade, M.R.; Costa, J.A. Mixotrophic cultivation of microalga Spirulina platensis using molasses as organic substrate. Aquaculture 2007, 264, 130–134. [Google Scholar] [CrossRef]
- Chojnacka, K.; Noworyta, A. Evaluation of Spirulina sp. growth in photoautotrophic, heterotrophic and mixotrophic cultures. Enzyme Microb. Technol. 2004, 34, 461–465. [Google Scholar] [CrossRef]
- Kadam, K.L. Power plant flue gas as a source of CO2 for microalgae cultivation: Economic impact of different process options. Energy Convers. Manag. 1997, 38, S505–S510. [Google Scholar] [CrossRef]
- Kadam, K.L. Microalgae Production from Power Plant Flue Gas: Environmental Implications on a Life Cycle Basis; National Renewable Energy Laboratory: Golden, CO, USA, 2001. [Google Scholar]
- Maeda, K.; Owada, M.; Kimura, N.; Omata, K.; Karube, I. CO2 fixation from the flue gas on coal-fired thermal power plant by microalgae. Energy Convers. Manag. 1995, 36, 717–720. [Google Scholar] [CrossRef]
- Stepan, D.J.; Shockey, R.E.; Moe, T.A.; Dorn, R. Carbon Dioxide Sequestering Using Microalgal Systems; University of North Dakota: Grand Forks, ND, USA, 2002. [Google Scholar]
- Cheah, W.Y.; Show, P.L.; Chang, J.S.; Ling, T.C.; Juan, J.C. Biosequestration of atmospheric CO2 and flue gas-containing CO2 by microalgae. Bioresour. Technol. 2015, 184, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Cheah, W.Y.; Ling, T.C.; Juan, J.C.; Lee, D.J.; Chang, J.S.; Show, P.L. Biorefineries of carbon dioxide: From carbon capture and storage (CCS) to bioenergies production. Bioresour. Technol. 2016, 215, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Huang, Y.; Feng, J.; Sun, J.; Zhou, J.; Cen, K. Improving CO2 fixation efficiency by optimizing Chlorella PY-ZU1 culture conditions in sequential bioreactors. Bioresour. Technol. 2013, 144, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Su, Y.; Zhang, Y.; Cui, G. Carbon dioxide fixation and biomass production from combustion flue gas using energy microalgae. Energy 2015, 89, 347–357. [Google Scholar] [CrossRef]
- Kao, C.-Y.; Chen, T.-Y.; Chang, Y.-B.; Chiu, T.-W.; Lin, H.-Y.; Chen, C.-D.; Chang, J.-S.; Lin, C.-S. Utilization of carbon dioxide in industrial flue gases for the cultivation of microalga Chlorella sp. Bioresour. Technol. 2014, 166, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Li, F.F.; Yang, Z.H.; Zeng, R.; Yang, G.; Chang, X.; Yan, J.B.; Hou, Y.L. Microalgae capture of CO2 from actual flue gas discharged from a combustion chamber. Ind. Eng. Chem. Res. 2011, 50, 6496–6502. [Google Scholar] [CrossRef]
- Rao, P.H.; Kumar, R.R.; Raghavan, B.; Subramanian, V.; Sivasubramanian, V. Application of phycoremediation technology in the treatment of wastewater from a leather-processing chemical manufacturing facility. Water SA 2011, 37, 7–14. [Google Scholar] [CrossRef]
- Christenson, L.; Sims, R. Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol. Adv. 2011, 29, 686–702. [Google Scholar] [CrossRef] [PubMed]
- Oswald, W.J. My sixty years in applied algology. J. Appl. Phycol. 2003, 15, 99–106. [Google Scholar] [CrossRef]
- Munoz, R.; Guieysse, B. Algal-bacterial processes for the treatment of hazardous contaminants: A review. Water Res. 2006, 40, 2799–2815. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y.; Wu, Y.H.; Zhu, S.F.; Li, F.M.; Hu, H.Y. Isolation and heterotrophic cultivation of mixotrophic microalgae strains for domestic wastewater treatment and lipid production under dark condition. Bioresour. Technol. 2013, 149, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Renuka, N.; Sood, A.; Prasanna, R.; Ahluwalia, A. phycoremediation of wastewaters: A synergistic approach using microalgae for bioremediation and biomass generation. Int. J. Environ. Sci. Technol. 2015, 12, 1443–1460. [Google Scholar] [CrossRef]
- Chinnasamy, S.; Bhatnagar, A.; Hunt, R.W.; Das, K.C. Microalgae cultivation in a wastewater dominated by carpet mill effluents for biofuel applications. Bioresour. Technol. 2010, 101, 3097–3105. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Barrow, C.; Shahidi, F. Marine Nutraceuticals and Functional Foods; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Da Silva Vaz, B.; Moreira, J.B.; de Morais, M.G.; Costa, J.A.V. Microalgae as a new source of bioactive compounds in food supplements. Curr. Opin. Food Sci. 2016, 7, 73–77. [Google Scholar]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Schenk, P.M. Towards sustainable sources for omega-3 fatty acids production. Curr. Opin. Biotechnol. 2014, 26, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Chen, C.C.; Huynh, P.; Chang, J.S. Exploring the potential of using algae in cosmetics. Bioresour. Technol. 2015, 184, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Jeffrey, S.; Volkman, J.; Dunstan, G. Nutritional properties of microalgae for mariculture. Aquaculture 1997, 151, 315–331. [Google Scholar] [CrossRef]
- Marris, E. Putting the carbon back: Black is the new green. Nature 2006, 442, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Gudin, C.; Thepenier, C. Bioconversion of Solar Energy into Organic Chemicals by Microalgae. Adv. Biotechnol. Processes 1986, 6, 73–110. [Google Scholar]
- Grima, E.M.; Belarbi, E.-H.; Fernández, F.A.; Medina, A.R.; Chisti, Y. Recovery of microalgal biomass and metabolites: Process options and economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef]
- Divakaran, R.; Pillai, V.S. Flocculation of algae using chitosan. J. Appl. Phycol. 2002, 14, 419–422. [Google Scholar] [CrossRef]
- Sukenik, A.; Shelef, G. Algal autoflocculation—Verification and proposed mechanism. Biotechnol. Bioeng. 1984, 26, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, S.; Kim, J.; Kwon, G.; Yoon, B.; Oh, H. Effects of harvesting method and growth stage on the flocculation of the green alga Botryococcus braunii. Lett. Appl. Microbiol. 1998, 27, 14–18. [Google Scholar] [CrossRef]
- Uduman, N.; Qi, Y.; Danquah, M.K.; Forde, G.M.; Hoadley, A. Dewatering of microalgal cultures: A major bottleneck to algae-based fuels. J. Renew. Sustain. Energy 2010, 2, 12701. [Google Scholar] [CrossRef]
- Bilanovic, D.; Shelef, G.; Sukenik, A. Flocculation of microalgae with cationic polymers—Effects of medium salinity. Biomass 1988, 17, 65–76. [Google Scholar] [CrossRef]
- Sukenik, A.; Bilanovic, D.; Shelef, G. Flocculation of microalgae in brackish and sea waters. Biomass 1988, 15, 187–199. [Google Scholar] [CrossRef]
- Oh, H.-M.; Lee, S.J.; Park, M.-H.; Kim, H.-S.; Kim, H.-C.; Yoon, J.-H.; Kwon, G.-S.; Yoon, B.-D. Harvesting of Chlorella vulgaris using a bioflocculant from Paenibacillus sp. AM49. Biotechnol. Lett. 2001, 23, 1229–1234. [Google Scholar] [CrossRef]
- Wiley, P.E.; Brenneman, K.J.; Jacobson, A.E. Improved algal harvesting using suspended air flotation. Water Environ. Res. 2009, 81, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Edzwald, J. Algae, bubbles, coagulants, and dissolved air flotation. Water Sci. Technol. 1993, 27, 67–81. [Google Scholar]
- Pushparaj, B.; Pelosi, E.; Torzillo, G.; Materassi, R. Microbial biomass recovery using a synthetic cationic polymer. Bioresour. Technol. 1993, 43, 59–62. [Google Scholar] [CrossRef]
- Muylaert, K.; Vandamme, D.; Meesschaert, B.; Foubert, I. Flocculation of microalgae using cationic starch. In Phycologia; The Physiological Society: London, UK, 2009; p. 63. [Google Scholar]
- Phoochinda, W.; White, D.A. Removal of algae using froth flotation. Environ. Technol. 2003, 24, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Ndikubwimana, T.; Chang, J.; Xiao, Z.; Shao, W.; Zeng, X.; Ng, I.S.; Lu, Y. Flotation: A promising microalgae harvesting and dewatering technology for biofuels production. Biotechnol. J. 2016, 11, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Yoon, R.; Luttrell, G. The effect of bubble size on fine particle flotation. Miner. Process. Extr. Metall. Review. 1989, 5, 101–122. [Google Scholar] [CrossRef]
- Nurdogan, Y.; Oswald, W.J. Tube settling of high-rate pond algae. Water Sci. Technol. 1996, 33, 229–241. [Google Scholar] [CrossRef]
- Zimmerman, W.B.; Tesar, V.; Butler, S.; Bandulasena, H.C. Microbubble generation. Recent Pat. Eng. 2008, 2, 1–8. [Google Scholar] [CrossRef]
- Ghernaout, D.; Benblidia, C.; Khemici, F. Microalgae removal from Ghrib Dam (Ain Defla, Algeria) water by electroflotation using stainless steel electrodes. Desalination Water Treat. 2015, 54, 3328–3337. [Google Scholar] [CrossRef]
- Santander, M.; Rodrigues, R.T.; Rubio, J. Modified jet flotation in oil (petroleum) emulsion/water separations. Colloids Surf. A Physicochem. Eng. Asp. 2011, 375, 237–244. [Google Scholar] [CrossRef]
- Cheng, Y.L.; Juang, Y.C.; Liao, G.Y.; Tsai, P.W.; Ho, S.H.; Yeh, K.L.; Chen, C.Y.; Chang, J.S.; Liu, J.C.; Chen, W.M.; et al. Harvesting of Scenedesmus obliquus FSP-3 using dispersed ozone flotation. Bioresour. Technol. 2011, 102, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Jameson, G.J. Application of the Jameson cell technology for algae and phosphorus removal from maturation ponds. Int. J. Miner. Process. 2004, 73, 23–28. [Google Scholar]
- Chen, Y.; Liu, J.; Ju, Y.-H. Flotation removal of algae from water. Colloids Surf. B Biointerfaces 1998, 12, 49–55. [Google Scholar] [CrossRef]
- Heasman, M.; Diemar, J.; O’connor, W.; Sushames, T.; Foulkes, L. Development of extended shelf-life microalgae concentrate diets harvested by centrifugation for bivalve molluscs—A summary. Aquac. Res. 2000, 31, 637–659. [Google Scholar] [CrossRef]
- Bosma, R.; van Spronsen, W.A.; Tramper, J.; Wijffels, R.H. Ultrasound, a new separation technique to harvest microalgae. J. Appl. Phycol. 2003, 15, 143–153. [Google Scholar] [CrossRef]
- Knuckey, R.M.; Brown, M.R.; Robert, R.; Frampton, D.M. Production of microalgal concentrates by flocculation and their assessment as aquaculture feeds. Aquac. Eng. 2006, 35, 300–313. [Google Scholar] [CrossRef]
- Otto, C.; Baumann, M.; Schreiner, T.; Bartsch, G.; Borberg, H.; Schwandt, P.; Schmid-Schönbein, H. Standardized ultrasound as a new method to induce platelet aggregation: Evaluation, influence of lipoproteins and of glycoprotein IIb/IIIa antagonist tirofiban. Eur. J. Ultrasound 2001, 14, 157–166. [Google Scholar] [CrossRef]
- Mohn, F. Experiences and Strategies in the Recovery of Biomass from Mass Cultures of Microalgae, Algae Biomass: Production and Use/Sponsored by the National Council for Research and Development, Israel and the Gesellschaft fur Strahlen-und Umweltforschung (GSF), Munich, Germany; Shelef, G., Soeder, C.J., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 1980. [Google Scholar]
- Petrusevski, B.; Bolier, G.; van Breemen, A.; Alaerts, G. Tangential flow filtration: A method to concentrate freshwater algae. Water Res. 1995, 29, 1419–1424. [Google Scholar] [CrossRef]
- MacKay, D.; Salusbury, T. Choosing between centrifugation and crossflow microfiltration. Chem. Eng. 1988, 447, 45–50. [Google Scholar]
- Mollah, M.Y.; Morkovsky, P.; Gomes, J.A.; Kesmez, M.; Parga, J.; Cocke, D.L. Fundamentals, present and future perspectives of electrocoagulation. J. Hazard. Mater. 2004, 114, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Alfafara, C.G.; Nakano, K.; Nomura, N.; Igarashi, T.; Matsumura, M. Operating and scale-up factors for the electrolytic removal of algae from eutrophied lakewater. J. Chem. Technol. Biotechnol. 2002, 77, 871–876. [Google Scholar] [CrossRef]
- Prakash, J.; Pushparaj, B.; Carlozzi, P.; Torzillo, G.; Montaini, E.; Materassi, R. Microalgal biomass drying by a simple solar device. Int. J. Sol. Energy 1997, 18, 303–311. [Google Scholar] [CrossRef]
- Desmorieux, H.; Decaen, N. Convective drying of spirulina in thin layer. J. Food Eng. 2005, 66, 497–503. [Google Scholar] [CrossRef]
- Hejazi, M.A.; Wijffels, R.H. Milking of microalgae. Trends Biotechnol. 2004, 22, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, K.; Sawayama, S. Liquid fuel production using microalgae. J. Jpn. Pet. Inst. 2005, 48, 251. [Google Scholar] [CrossRef]
- Biomass R&D, Technical Advisory Committee. Roadmap for Biomass Technologies in the United States, Washington, DC, USA. Available online: www.bioproducts-bioenergy.gov/pdfs/FinalBiomassRoadmap.pdf (accessed on 22 December 2002).
- Demirbaş, A. Biomass resource facilities and biomass conversion processing for fuels and chemicals. Energy Convers Manag. 2001, 42, 1357–1378. [Google Scholar] [CrossRef]
- Hirano, A.; Hon-Nami, K.; Kunito, S.; Hada, M.; Ogushi, Y. Temperature effect on continuous gasification of microalgal biomass: Theoretical yield of methanol production and its energy balance. Catal. Today 1998, 45, 399–404. [Google Scholar] [CrossRef]
- Raheem, A.; Wan Azlina, K.G.; Taufiq Yap, Y.H.; Danquah, M.K.; Harun, R. Thermochemical conversion of microalgal biomass for biofuel production. Renew. Sustain. Energy Rev. 2015, 49, 990–999. [Google Scholar] [CrossRef]
- Goyal, H.; Seal, D.; Saxena, R. Bio-fuels from thermochemical conversion of renewable resources: A review. Renew. Sustain. Energy Rev. 2008, 12, 504–517. [Google Scholar] [CrossRef]
- McKendry, P. Energy production from biomass (part 2): Conversion technologies. Bioresour. Technol. 2002, 83, 47–54. [Google Scholar] [CrossRef]
- Dote, Y.; Sawayama, S.; Inoue, S.; Minowa, T.; Yokoyama, S.-Y. Recovery of liquid fuel from hydrocarbon-rich microalgae by thermochemical liquefaction. Fuel 1994, 73, 1855–1857. [Google Scholar] [CrossRef]
- Minowa, T.; Yokoyama, S.Y.; Kishimoto, M.; Okakura, T. Oil production from algal cells of Dunaliella tertiolecta by direct thermochemical liquefaction. Fuel 1995, 74, 1735–1738. [Google Scholar] [CrossRef]
- Demirbaş, A. Oily products from mosses and algae via pyrolysis. Energy Sources Part A 2006, 28, 933–940. [Google Scholar] [CrossRef]
- Miao, X.; Wu, Q. High yield bio-oil production from fast pyrolysis by metabolic controlling of Chlorella protothecoides. J. Biotechnol. 2004, 110, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Wu, Q.; Yang, C. Fast pyrolysis of microalgae to produce renewable fuels. J. Anal. Appl. Pyrolysis 2004, 71, 855–863. [Google Scholar] [CrossRef]
- Bridgwater, A. IEA Bioenergy 27th Update. Biomass Pyrolysis, Biomass and Bioenergy; Pergamon-Elsevier Science Ltd.: England, UK, 2007; Volume 31. [Google Scholar]
- Clark, J.H.; Deswarte, F. Introduction to Chemicals from Biomass; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Kadam, K.L. Environmental implications of power generation via coal-microalgae cofiring. Energy 2002, 27, 905–922. [Google Scholar] [CrossRef]
- Zhu, L.D.; Hiltunen, E.; Antila, E.; Zhong, J.J.; Yuan, Z.H.; Wang, Z.M. Microalgal biofuels: Flexible bioenergies for sustainable development. Renew. Sustain. Energy Rev. 2014, 30, 1035–1046. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chang, H.Y.; Chang, J.S. Producing carbohydrate-rich microalgal biomass grown under mixotrophic conditions as feedstock for biohydrogen production. Int. J. Hydrog. Energy 2016, 41, 4413–4420. [Google Scholar] [CrossRef]
- Leite, G.B.; Abdelaziz, A.E.; Hallenbeck, P.C. Algal biofuels: Challenges and opportunities. Bioresour. Technol. 2013, 145, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Lakaniemi, A.M.; Tuovinen, O.H.; Puhakka, J.A. Anaerobic conversion of microalgal biomass to sustainable energy carriers—A review. Bioresour. Technol. 2013, 135, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Appels, L.; Baeyens, J.; Degrève, J.; Dewil, R. Principles and potential of the anaerobic digestion of waste-activated sludge. Prog. Energy Combust. Sci. 2008, 34, 755–781. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, C.; Mandy, A.; Ballesteros, I.; Ballesteros, M. Impact of temperature and photoperiod on anaerobic biodegradability of microalgae grown in urban wastewater. Int. Biodeterior. Biodegrad. 2016, 106, 16–23. [Google Scholar] [CrossRef]
- Hernandez, D.; Riano, B.; Coca, M.; Solana, M.; Bertucco, A.; Garcia-Gonzalez, M.C. Microalgae cultivation in high rate algal ponds using slaughterhouse wastewater for biofuel applications. Chem. Eng. J. 2016, 285, 449–458. [Google Scholar] [CrossRef]
- Mendez, L.; Mahdy, A.; Ballesteros, M.; Gonzalez-Fernandez, C. Methane production of thermally pretreated Chlorella vulgaris and Scenedesmus sp. biomass at increasing biomass loads. Appl. Energy 2014, 129, 238–242. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, W.; Chen, B.Y.; Cheng, C.L.; Lo, Y.C.; Ho, S.H.; Chang, J.S.; Ren, N. Exploring the inhibitory characteristics of acid hydrolysates upon butanol fermentation: A toxicological assessment. Bioresour. Technol. 2015, 198, 571–576. [Google Scholar] [CrossRef] [PubMed]
- John, R.P.; Anisha, G.S.; Nampoothiri, K.M.; Pandey, A. Micro and microalgal biomass: A renewable source for bioethanol. Bioresour. Technol. 2011, 102, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Markou, G.; Angelidaki, I.; Nerantzis, E.; Georgakakis, D. Bioethanol production by carbohydrate-enriched biomass of Arthrospira (Spirulina) platensis. Energies 2013, 6, 3937–3950. [Google Scholar] [CrossRef]
- Ho, S.H.; Huang, S.W.; Chen, C.Y.; Hasunuma, T.; Kondo, A.; Chang, J.S. Bioethanol production using carbohydrate-rich microalgae biomass as feedstock. Bioresour. Technol. 2013, 135, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Castro, Y.A.; Ellis, J.T.; Miller, C.D.; Sims, R.C. Optimization of wastewater microalgae saccharification using dilute acid hydrolysis for acetone, butanol, and ethanol fermentation. Appl. Energy 2015, 140, 14–19. [Google Scholar] [CrossRef]
- Cheng, H.H.; Whang, L.M.; Chan, K.C.; Chung, M.C.; Wu, S.H.; Liu, C.P.; Tien, S.Y.; Chen, S.Y.; Chang, J.S.; Lee, W.J. Biological butanol production from microalgae-based biodiesel residues by Clostridium acetobutylicum. Bioresour. Technol. 2015, 184, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Bharathiraja, B.; Chakravarthy, M.; Kumar, R.R.; Yogendran, D.; Yuvaraj, D.; Jayamuthunagai, J.; Kumar, R.P.; Palani, S. Aquatic biomass (algae) as a future feed stock for bio-refineries: A review on cultivation, processing and products. Renew. Sustain. Energy Rev. 2015, 47, 634–653. [Google Scholar] [CrossRef]
- Yang, Z.; Guo, R.; Xu, X.; Fan, X.; Luo, S. Fermentative hydrogen production from lipid-extracted microalgal biomass residues. Appl. Energy 2011, 88, 3468–3472. [Google Scholar] [CrossRef]
- Eroglu, E.; Melis, A. Microalgal hydrogen production research. Int. J. Hydrog. Energy 2016, 41, 12772–12798. [Google Scholar] [CrossRef]
- Melis, A.; Zhang, L.; Forestier, M.; Ghirardi, M.L.; Seibert, M. Sustained photobiological hydrogen gas production upon reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii. Plant Physiol. 2000, 122, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Pragya, N.; Pandey, K.K.; Sahoo, P.K. A review on harvesting, oil extraction and biofuels production technologies from microalgae. Renew. Sustain. Energy Rev. 2013, 24, 159–171. [Google Scholar] [CrossRef]
- Barnwal, B.; Sharma, M. Prospects of biodiesel production from vegetable oils in India. Renew. Sustain. Energy Rev. 2005, 9, 363–378. [Google Scholar] [CrossRef]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Kondo, A.; Noda, H. Biodiesel fuel production by transesterification of oils. J. Biosci. Bioeng. 2001, 92, 405–416. [Google Scholar] [CrossRef]
- Zhu, L. Microalgal culture strategies for biofuel production: A review. Biofuels Bioprod. Biorefining 2015, 9, 801–814. [Google Scholar] [CrossRef]
- Yen, H.W.; Hu, I.C.; Chen, C.Y.; Ho, S.H.; Lee, D.J.; Chang, J.S. Microalgae-based biorefinery—From biofuels to natural products. Bioresour. Technol. 2013, 135, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ho, S.H.; Cheng, C.L.; Guo, W.Q.; Nagarajan, D.; Ren, N.Q.; Lee, D.J.; Chang, J.S. Perspectives on the feasibility of using microalgae for industrial wastewater treatment. Bioresour. Technol. 2016, 222, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Lauersen, K.J.; Huber, I.; Wichmann, J.; Baier, T.; Leiter, A.; Gaukel, V.; Kartushin, V.; Rattenholl, A.; Steinweg, C.; von Riesen, L.; et al. Investigating the dynamics of recombinant protein secretion from a microalgal host. J. Biotechnol. 2015, 215, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Raeesossadati, M.J.; Ahmadzadeh, H.; McHenry, M.P.; Moheimani, N.R. CO2 bioremediation by microalgae in photobioreactors: Impacts of biomass and CO2 concentrations, light, and temperature. Algal Res. 2014, 6, 78–85. [Google Scholar] [CrossRef]
- Sharma, Y.; Singh, B. Development of biodiesel: Current scenario. Renew. Sustain. Energy Rev. 2009, 13, 1646–1651. [Google Scholar] [CrossRef]
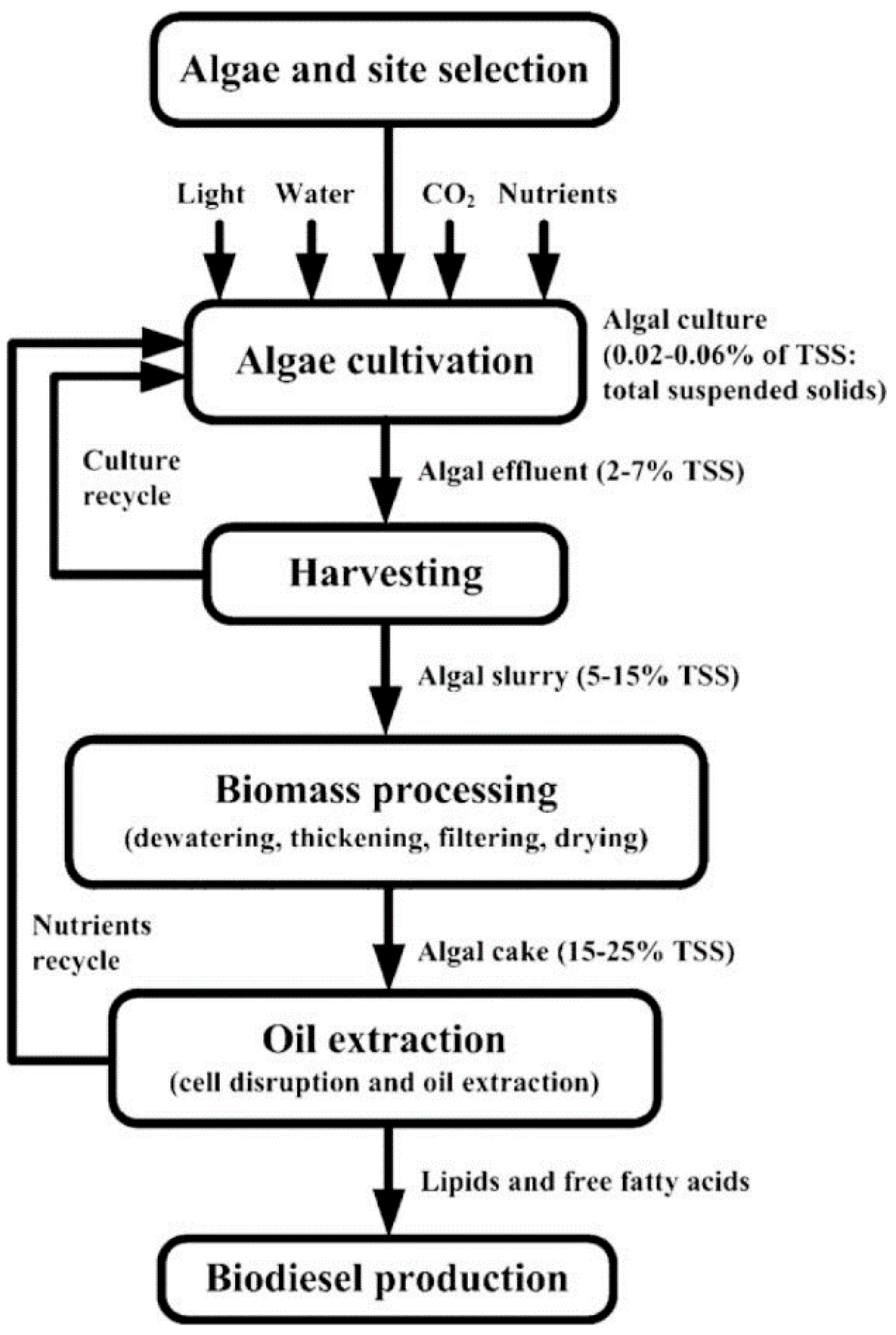

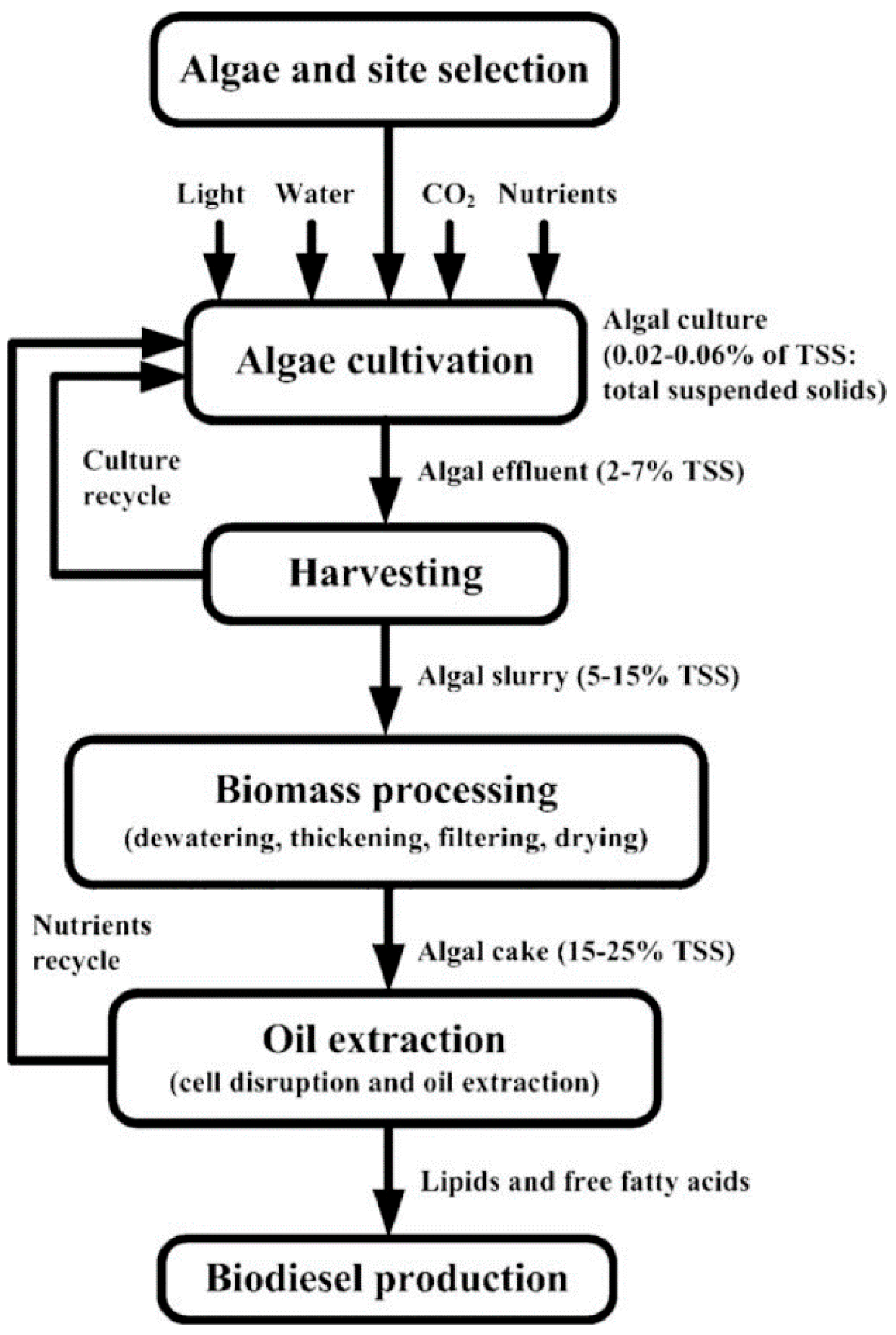
{kind=link}
{kind=link}
| Crops | Gallon acre−1·year−1 |
|---|---|
| Corn | 15 |
| Soybean | 48 |
| Sunflower | 102 |
| Rapeseed | 127 |
| Oil palm | 635 |
| Microalgae—actual biomass yield | 1850 |
| Microalgae—theoretical laboratory yield | 5000–15,000 |
| Microalgae Species | Lipid Content (% Dry Weight Biomass) | Lipid Productivity (mg/L/day) |
|---|---|---|
| Ankistrodesmus sp. | 24.0–31.0 | - |
| Botryococcus braunii | 25.0–75.0 | - |
| Chaetoceros muelleri | 33.6 | 21.8 |
| Chaetoceros calcitrans | 14.6–16.4/39.8 | 17.6 |
| Chlorella emersonii | 25.0–63.0 | 10.3–50.0 |
| Chlorella protothecoides | 14.6–57.8 | 1214 |
| Chlorella sorokiniana | 19.0–22.0 | 44.7 |
| Chlorella vulgaris | 5.0–58.0 | 11.2–40.0 |
| Chlorella sp. | 10.0–48.0 | 42.1 |
| Chlorella pyrenoidosa | 2.0 | - |
| Chlorella | 18.0–57.0 | 18.7 |
| Chlorella zofingiensis | 45.5 | 473.0 |
| Chlorococcum sp. | 19.3 | 53.7 |
| Crypthecodinium cohnii | 20.0–51.1 | - |
| Dunaliella salina | 6.0–25.0 | 116.0 |
| Dunaliella primolecta | 23.1 | - |
| Dunaliella tertiolecta | 16.7–71.0 | - |
| Dunaliella sp. | 17.5–67.0 | 33.5 |
| Ellipsoidion sp. | 27.4 | 47.3 |
| Euglena gracilis | 14.0–20.0 | - |
| Haematococcus pluvialis | 25.0 | - |
| Isochrysis galbana | 7.0–40.0 | - |
| Isochrysis sp. | 7.1–33 | 37.8 |
| Monodus subterraneus | 16.0 | 30.4 |
| Monallanthus salina | 20.0–22.0 | - |
| Nannochloris sp. | 20.0–56.0 | 60.9–76.5 |
| Nannochloropsis oculata | 22.7–29.7 | 84.0–142.0 |
| Nannochloropsis sp. | 12.0–53.0 | 37.6–90.0 |
| Neochloris oleoabundans | 29.0–65.0 | 90.0–134.0 |
| Nitzschia sp. | 16.0–47.0 | - |
| Oocystis pusilla | 10.5 | - |
| Pavlova salina | 30.9 | 49.4 |
| Pavlova lutheri | 35.5 | 40.2 |
| Phaeodactylum tricornutum | 18.0–57.0 | 44.8 |
| Porphyridium cruentum | 9.0–18.8/60.7 | 34.8 |
| Scenedesmus obliquus | 11.0–55.0 | - |
| Scenedesmus quadricauda | 1.9–18.4 | 35.1 |
| Scenedesmus sp. | 19.6–21.1 | 40.8–53.9 |
| Skeletonema sp. | 13.3–31.8 | 27.3 |
| Skeletonema costatum | 13.5–51.3 | 17.4 |
| Spirulina platensis | 4.0–16.6 | - |
| Spirulina maxima | 4.0–9.0 | - |
| Thalassiosira pseudonana | 20.6 | 17.4 |
| Tetraselmis suecica | 8.5–23.0 | 27.0–36.4 |
| Tetraselmis sp. | 12.6–14.7 | 43.4 |
| Growth Condition | Maximum Productivities | ||
|---|---|---|---|
| Biomass (g·L−1·day−1) | Triacylglycerol (mg·L−1·day−1) | Astaxanthin (mg·L−1·day−1) | |
| Low Light | 0.83 ± 0.05 | 11.3 ± 0.7 | 0.05 ± 0.01 |
| Nitrogen Deprivation | 0.41 ± 0.02 | 91.5 ± 5.5 | 1.08 ± 0.06 |
| High Light | 1.40 ± 0.09 | 173.6 ± 11.2 | 2.01 ± 0.14 |
| Nitrogen Deprivation + High Light | 0.53 ± 0.03 | 145.8 ± 9.7 | 1.79 ± 0.17 |
| Algae Species | Xmax (g·L−1) | Paerial (g·m−2·day−1) | Pvolume (g·L−1·day−1) | PE (%) |
|---|---|---|---|---|
| Chlorella sp. | 10 | 25 | - | - |
| N/A | 0.14 | 35 | 0.117 | - |
| Spirulina platensis | - | - | 0.18 | - |
| Spirulina platensis | 0.47 | 14 | 0.05 | - |
| Haematococcus pluvialis | 0.202 | 15.1 | - | - |
| Spirulina | 1.24 | 69.16 | - | - |
| Spirulina platensis | 0.9 | 12.2 | 0.15 | - |
| Spirulina platensis | 1.6 | 19.4 | 0.32 | - |
| Anabaena sp. | 0.23 | 23.5 | 0.24 | >2 |
| Chlorella sp. | 40 | 23.5 | - | 6.48 |
| Chlorella sp. | 40 | 11.1 | - | 5.98 |
| Chlorella sp. | 40 | 32.2 | - | 5.42 |
| Chlorella sp. | 40 | 18.1 | - | 6.07 |
| Microalgae | T (°C) | CO2 (%) | Pvolume (g·L−1·day−1) | (g·L−1·day−1) | Carbon Usage Efficiency (%) |
|---|---|---|---|---|---|
| Chlorella sp. | 26 | Air | 0.682 a | - | - |
| Chlorella sp. | 26 | 2 | 1.445 a | - | 58 |
| Chlorella sp. | 26 | 5 | 0.899 a | - | 27 |
| Chlorella sp. | 26 | 10 | 0.106 a | - | 20 |
| Chlorella sp. | 26 | 15 | 0.099 a | - | 16 |
| Chlorella kessleri | 30 | 18 | 0.087 | - | - |
| Scenedesmus sp. | 25 | 10 | 0.218 | - | - |
| Chlorella vulgaris | 25 | 10 | 0.105 | - | - |
| Botryococcus braunii | 25 | 10 | 0.027 | - | - |
| Scenedesmus sp. | 25 | Flue gas | 0.203 | - | - |
| Botryococcus braunii | 25 | Flue gas | 0.077 | - | - |
| Chlorella vulgaris | 25 | Air | 0.040 | - | - |
| Chlorella vulgaris | 25 | Air | 0.024 | - | - |
| Haematococcus pluvialis | 20 | 16–34 | 0.076 | 0.143 | - |
| Scenedesmus obliquus | - | Air | 0.009 | 0.016 | - |
| Scenedesmus obliquus | - | Air | 0.016 | 0.031 | - |
| Chlorella vulgaris | 27 | 15 | - | 0.624 | - |
| Scenedesmus obliquus | 30 | 18 | 0.14 | 0.260 | - |
| Spirulina sp. | 30 | 12 | 0.22 | 0.413 | - |
| Properties | Butanol | Ethanol |
|---|---|---|
| Melting point (°C) | −89.3 | −114.0 |
| Specific gravity | 0.810–0.812 | 0.79 |
| Ignition temperature (°C) | 35–37 | 276–456 |
| Auto-ignition temperature (°C) | 343–345 | 422 |
| Flash point (°C) | 25–29 | 12.77 |
| Relative density | 0.81 | 0.805–0.812 |
| Critical temperature (°C) | 287 | 239.85 |
| Explosive limits (vol % in air) | 1.4–11.3 | 3.3–19.0 |
| Vapor pressure (kPa at 20 °C) | 0.5 | 5.95 |
| Boiling point (°C) | 117–118 | 78 |
| Density at 20 °C (g/mL) | 0.8098 | 0.7851 |
| Energy density (MJ/L−1) | 27.0–29.2 | 19.6 |
| Energy content (BTU/gal) | 110,000 | 84,000 |
| Liquid heat capacity at STP (kJ/kmol·°K) | 178 | 112.3 |
| Research octane number | 96 | 129 |
| Motor octane number | 78 | 102 |
| Viscosity (10−3 Pa·s) | 2.593 | 1.078 |
| Biomass | Methane Yield (m3·kg−1) |
|---|---|
| Laminaria sp. | 0.26–0.28 |
| Gracilaria sp. | 0.28–0.40 |
| Sargassum sp. | 0.12–0.19 |
| Macrocystis | 0.39–0.41 |
| L. digitata | 0.50 |
| Ulva sp. | 0.20 |
| Water hyacinth | 0.13–0.21 |
| Sorghum | 0.26–0.39 |
| Poplar | 0.23–0.32 |
| Food waste | 0.54 |
| Microalgae—ACAD model | 0.54 |
| Inhibitor | Moderate Inhibitory Concentration (mg·L−1) | Strongly Inhibitory Concentration (mg·L−1) |
|---|---|---|
| Na+ | 3500–5500 | 8000 |
| NH4+ | 1500–3500 | 3000 |
| K+ | 2500–4500 | 12,000 |
| Ca2+ | 2500–4000 | 8000 |
| Mg2+ | 1000–1500 | 3000 |
| S2− | 200 | 200 |
| Cu2+ | ns (1) | 0.5 (2) |
| Cr3+ | ns (1) | 200–250 (3) |
| Cr6+ | 10 | 3.0 (2) |
| Zn2+ | ns (1) | 1.0 (2) |
| Ni2+ | ns (1) | 30 (3) |
| VFAs | ns (1) | 6.7–9.0 (4) |
| 18-C LCFA | ns (1) | 1000 |
| Plant Source | Seed Oil Content (% Oil by wt in Biomass) | Oil Yield (L·Oil/ha-Year) | Land Use (m2·Year/kg Biodiesel) | Biodiesel Productivity (kg·Biodiesel/ha-Year) |
|---|---|---|---|---|
| Corn (Zea mays L.) | 44 | 172 | 66 | 152 |
| Hemp (Cannabis sativa L.) | 33 | 363 | 31 | 321 |
| Soybean (Glycine max L.) | 18 | 636 | 18 | 562 |
| Jatropha (Jatropha curcas L.) | 28 | 741 | 15 | 656 |
| Camelina (Camelina sativa L.) | 42 | 915 | 12 | 809 |
| Rapeseed (Brassica napus L.) | 41 | 974 | 12 | 862 |
| Sunflower (Helianthus annuus L.) | 40 | 1070 | 11 | 946 |
| Castor (Ricinus communis) | 48 | 1307 | 9 | 1156 |
| Palm oil (Elaeis guineensis) | 36 | 5366 | 2 | 4747 |
| Microalgae (low oil content) | 30 | 58,700 | 0.2 | 51,927 |
| Microalgae (medium oil content) | 50 | 97,800 | 0.1 | 86,515 |
| Microalgae (high oil content) | 70 | 136,900 | 0.1 | 121,104 |
| Parameter | Open Pond | PBR | Fermenter |
|---|---|---|---|
| Land requirement | High | Varied | Low |
| Water loss | Very high | Low | Low |
| Hydrodynamic stress on algae | Very low | Low-high | Unknown |
| Gas transfer control | Low | High | High |
| CO2 loss | High | Low | No CO2 required |
| O2 inhibition | Usually low enough due to continuous spontaneous outgassing | High | O2 supply should be sufficient |
| Temperature | Highly varied | Cooling required | Needs to be maintained |
| Startup period | 6–8 weeks | 2–4 weeks | 2–4 weeks |
| Construction costs | USD $100,000 per hectare | USD $1 million per hectare | Low |
| Operation costs | Low | Very high | Very high |
| Limiting factor for growth | Light | Light | O2 |
| Control over parameters | Low | Medium | Very high |
| Technology | Readily available | Under development | Readily available |
| Pollution risk | High | Medium | Low |
| Pollution control | Difficult | Easy | Easy |
| Species control | Difficult | Easy | Easy |
| Weather dependence | High: light intensity, temperature, rainfall | Medium | Low |
| Maintenance | Easy | Difficult | Difficult |
| Cleaning | Easy | Difficult | Difficult |
| Overheating risk | Low | High | Unknown |
| Excessive O2 levels risk | Low | High | Unknown |
| Cell density in culture | 0.1–0.5 g·L−1 | 2–8 g·L−1 | 15.5 or even 80.0–110.0 g·L−1 |
| Light-induced products (pigments, chlorophyll, etc.) | No impact | No impact | Reduced |
| Surface area-to-volume ratio | High | Very high | Not applicable |
| Applicability to different species | Low | High | Low |
| Ease of scale-up | High | Varied | High |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Show, P.L.; Tang, M.S.Y.; Nagarajan, D.; Ling, T.C.; Ooi, C.-W.; Chang, J.-S. A Holistic Approach to Managing Microalgae for Biofuel Applications. Int. J. Mol. Sci. 2017, 18, 215. https://doi.org/10.3390/ijms18010215
Show PL, Tang MSY, Nagarajan D, Ling TC, Ooi C-W, Chang J-S. A Holistic Approach to Managing Microalgae for Biofuel Applications. International Journal of Molecular Sciences. 2017; 18(1):215. https://doi.org/10.3390/ijms18010215
Chicago/Turabian StyleShow, Pau Loke, Malcolm S. Y. Tang, Dillirani Nagarajan, Tau Chuan Ling, Chien-Wei Ooi, and Jo-Shu Chang. 2017. "A Holistic Approach to Managing Microalgae for Biofuel Applications" International Journal of Molecular Sciences 18, no. 1: 215. https://doi.org/10.3390/ijms18010215
APA StyleShow, P. L., Tang, M. S. Y., Nagarajan, D., Ling, T. C., Ooi, C.-W., & Chang, J.-S. (2017). A Holistic Approach to Managing Microalgae for Biofuel Applications. International Journal of Molecular Sciences, 18(1), 215. https://doi.org/10.3390/ijms18010215