
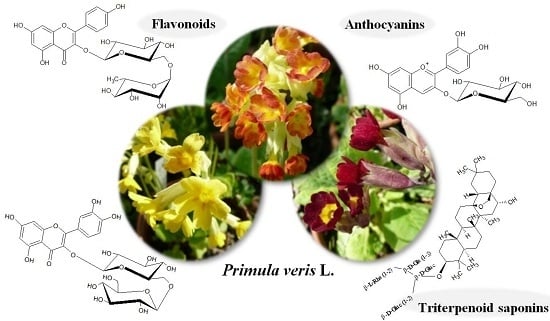
Comparative Metabolite Profiling of Triterpenoid Saponins and Flavonoids in Flower Color Mutations of Primula veris L.


Abstract
:
1. Introduction
2. Results
2.1. Color Variants of Primula veris
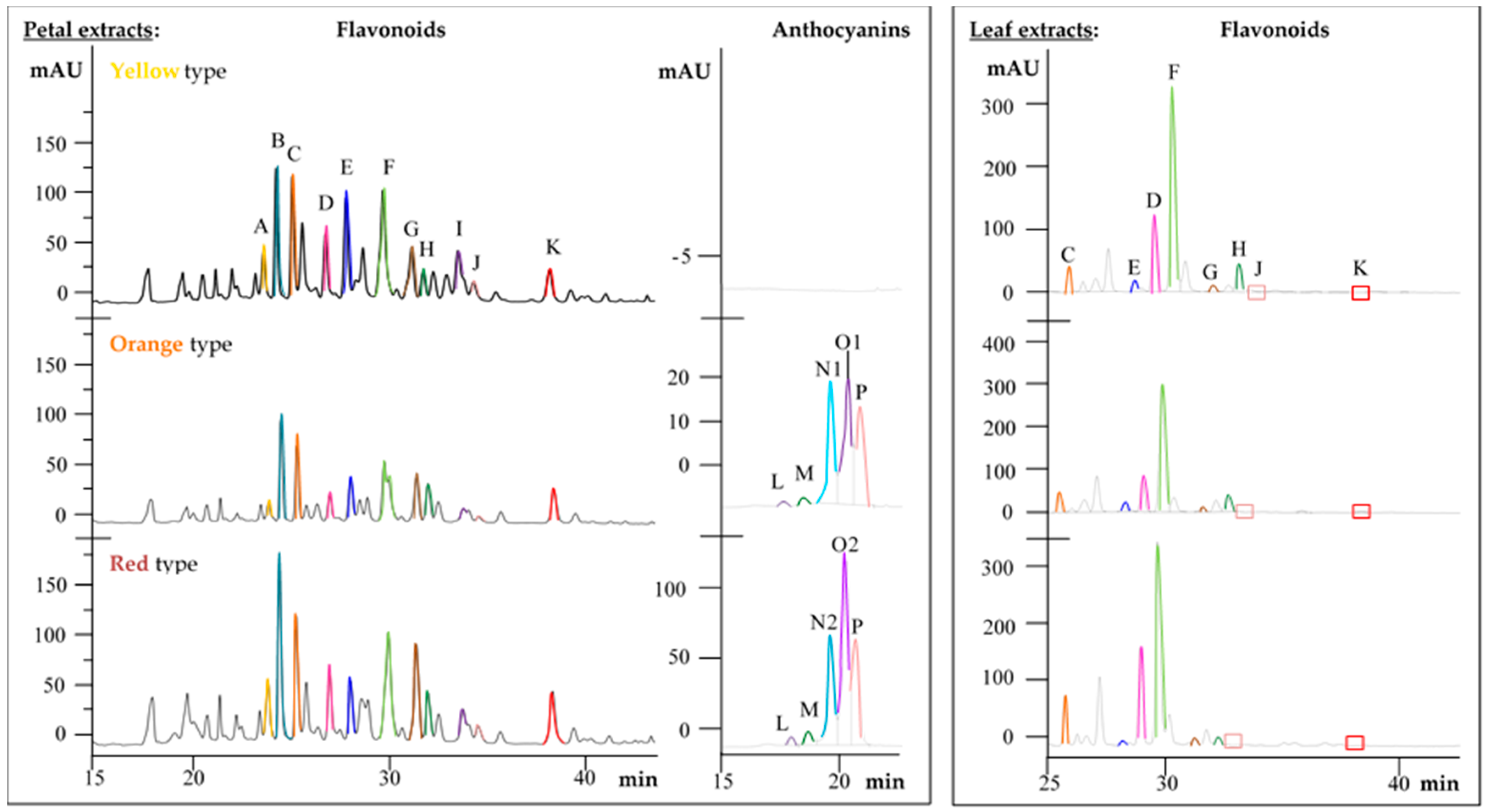
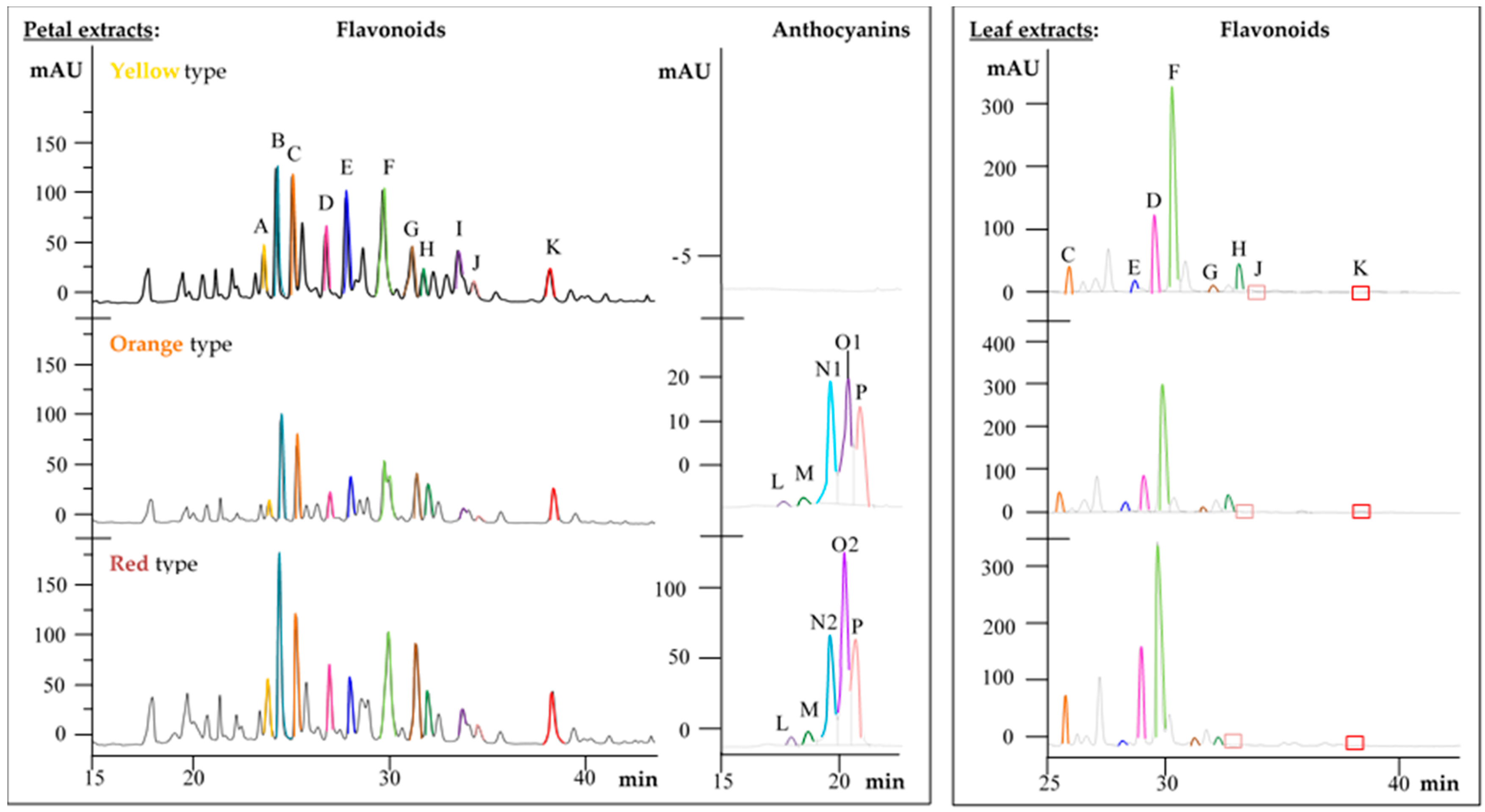
2.2. Metabolite Profiles of the Primula Variants
2.3. Comparison of Metabolite Profiles between Plant Parts
2.4. Identification of Novel Compounds in the Primula Variants
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chemicals
4.3. Sample Preparation
4.4. Microscopic Investigations
4.5. HPLC-DAD Analysis
4.6. LC-MSn Analyses
4.7. Characterization of Phenolic Compounds and Triterpenoid Saponins
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morozowska, M.; Wesołowska, M. In vitro clonal propagation of Primula veris L. and preliminary phytochemical analysis. Acta Biol. Crac. Bot. 2004, 46, 169–175. [Google Scholar]
- Laenger, R.; Sankel, J. Systematics of Primula veris (Primulaceae). Plant Syst. Evol. 1993, 188, 31–55. [Google Scholar]
- Köhlein, F. Primeln und die Verwandten Gattungen Mannsschild, Heilglöckchen, Götterblume, Troddelblume, Goldprimel; Verlag Eugen Ulmer: Stuttgart, Germany, 1984; pp. 104–108. [Google Scholar]
- HagerRom. Hagers Enzyklopädie der Arzneistoffe und Drogen; Springer: Heidelberg, Germany, 2015. [Google Scholar]
- Karrer, P.; Widmer, R. Über Primelfarbstoffe. Pflanzenfarbstoffe V. Helv. Chim. Acta 1927, 10, 758–763. [Google Scholar] [CrossRef]
- Paris, R. On the flavonoids of native species of Primula. Presence of a heteroside of kaempferol in the flowers of Primula officinalis Jacq. Ann. Pharm. Franc. 1959, 17, 331–335. [Google Scholar] [PubMed]
- Harborne, J.B. Comparative biochemistry of the flavonoids—VII, Correlations between flavonoid pigmentation and systematics in the family Primulaceae. Phytochemistry 1968, 7, 1215–1230. [Google Scholar] [CrossRef]
- Bat-Smith, E.C. The phenolic constituents of plants and their taxonomic significance. I. Dicotyledons. J. Linn. Soc. Bot. 1962, 58, 95–173. [Google Scholar] [CrossRef]
- Hegnauer, R. Chemotaxonomie der Pflanzen, 5th ed.; Birkhäuser Verlag: Basel, Switzerland, 1969; pp. 387–454. [Google Scholar]
- El Morchid, E.M.; Londono, P.T.; Papagiannopoulos, M.; Gobbo-Neto, L.; Mueller, C. Variation in flavonoid pattern in leaves and flowers of Primula veris of different origin and impact of UV-B. Biochem. Syst. Ecol. 2014, 53, 81–88. [Google Scholar] [CrossRef]
- Kraft, K. Symptomatische Phytotherapie bei Husten. Stellenwert pflanzlicher Antitussiva und Expektorantien. Pharm. Unserer Zeit 2008, 6, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.; Ganzera, M.; Stuppner, H. Analysis of phenolic glycosides and saponins in Primula elatior and Primula veris (primula root) by liquid chromatography, evaporative light scattering detection and mass spectrometry. J. Chromatogr. A 2006, 1112, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Pemp, E.; Krenn, L. Analytik Pharmazeutisch Relevanter Flavonoide Mit DC und LC/DAD/MS, 1st ed.; Südwestdeutscher Verlag für Hochschulschriften AG & Co. KG: Saarbrücken, Germany, 2005. [Google Scholar]
- Kim, J.; Matsuba, Y.; Ning, J.; Schulmiller, A.L.; Hammar, D.; Jones, A.D.; Pickersky, E.; Last, R.L. Analysis of natural and induced variation in tomato glandular trichome flavonoids identifies a gene not present in the reference genome. Plant Cell 2014, 26, 3272–3285. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Casals, I.; Andres-Lacueva, C.; Izquierdo-Pulido, M.; Lamuela-Raventos, R.M. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of the phenolic composition of cocoa (Theobroma cacao). J. Mass Spectrom. 2002, 38, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Fabre, N.; Rustan, I. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. Am. Soc. Mass Spectrom. 2001, 12, 707–715. [Google Scholar] [CrossRef]
- Gonzales, G.B.; Raes, K.; Coleus, S.; Struijs, K.; Smagghe, G.; Camp, J.V. Angiotensin converting enzyme inhibitory effects by plant phenolic compounds: A study of structure activity relationships. J. Chromatogr. A 2013, 61, 11832–11839. [Google Scholar]
- Tsiklauri, L.; Guohua, A.; Ruszaj, D.M.; Alaniyaa, M.; Kemertelidzea, E.; Morris, M.E. Simultaneous determination of the flavonoids robinin and kaempferol in human breast cancer cells by liquid chromatography-tandem mass spectrometry. J. Pharm. Biomed. 2011, 55, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Prior, R.L. Systematic identification and characterization of anthocyanins by HPLC-ESI-MS/MS in common foods in the United States: Fruits and berries. J. Agric. Food Chem. 2005, 53, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Karl, C.; Mueller, G.; Pedersen, P.A. Die Flavonoide in den Blüten von Primula officinalis. J. Med. Plants Res. 1981, 41, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.; Mabry, T.J. The Flavonoids: Advances in Research; Chapman and Hall: New York, NY, USA, 1982. [Google Scholar]
- McClure, J.W. Biochemical, Pharmacological and Structure—Activity Relationships. In Plant Flavonoids in Biology and Medicine; Alan R. Liss. Inc.: New York, NY, USA, 1986; pp. 77–135. [Google Scholar]
- Huck, C.W.; Huber, C.G.; Ongania, K.H.; Bonn, G.K. Isolation and characterization of methoxylated flavones in the flowers of Primula veris by liquid chromatography and mass spectrometry. J. Chromatogr. A 2000, 870, 453–462. [Google Scholar] [CrossRef]
- Huck, C.W.; Bonn, G.K. Evaluation of detection methods for the reversed-phase HPLC determination of 3′,4′,5′-trimethoxyflavone in different phytopharmaceutical products and in human serum. Phytochem. Anal. 2001, 12, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Sastry, C.V.; Rukmini, C.; Ramachandra, L. Chemistry of saponins. III. Isolation of new flavonol glycoside, 4′-O-methylquercetin-3-rutinoside, from Albizzia amara. Indian J. Chem. 1967, 5, 613–615. [Google Scholar]
- Ibrahim, L.F.; Kawashty, S.A.; El-Eraky, W.I.; Shabana, M.M.; El-Negoumy, S.I. A comparative study of flavonoids, pharmacological and antimicrobial effects of Echium longifolium and Heliotropium digynum. Egypt. J. Physiol. Sci. 2002, 24, 63–82. [Google Scholar]
- Han, J.; Ye, M.; Qiao, X.; Xu, M.; Wang, B.-R.; Guo, D.-A. Characterization of phenolic compounds in the Chinese herbal drug Artemisia annua by liquid chromatography coupled to electrospray ionization mass spectrometry. J. Pharam. Biomed. Anal. 2008, 47, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Kulevanova, S.; Stefova, M.; Panovks, T.K.; Stafilov, T. HPLC identification and determination of myricetin, quercetin, kaempferol and total flavonoids in herbal drugs. Maced. Pharm. Bull. 2003, 48, 25–30. [Google Scholar]
- Teshesche, R.; Toja, B.T.; Wulff, G. Über die Sapogenine aus den Wurzeln von Primula veris L. Liebigs Ann. Chem. 1966, 696, 160–179. [Google Scholar] [CrossRef]
- Hashimoto, N.; Ohsawa, R.; Kitajima, J.; Iwashina, T. New flavonol glycosides from leaves and flowers of Primula sieboldii. Nat. Prod. Commun. 2015, 10, 421–423. [Google Scholar] [PubMed]
- Shipunov, A.; Kosenko, Y.; Volkova, P. Floral polymorphism in common primrose (Primula vulgaris Huds., Primulaceae) of the Northeastern Black Sea coast. Plant Syst. Evol. 2011, 296, 167–178. [Google Scholar] [CrossRef]
- Volkova, P.A.; Schanzer, I.A. Colour polymorphism in common primrose (Primula vulgaris Huds.): Many colours—Many species? Plant Syst. Evol. 2013, 299, 1057–1087. [Google Scholar] [CrossRef]
- Hoshino, A.; Abe, Y.; Saito, N.; Inagaki, Y.; Iida, S. The gene encoding flavanone 3-hydroxylase is expressed normally in the pale yellow flowers of the Japanese morning glory carrying the speckled mutation which produce neither flavonol nor anthocyanin but accumulate chalcone, aurone and flavanone. Plant Cell Physiol. 1997, 38, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Clegg, M.; Durbin, M.L. Flower color variation: A model for experimental study of evolution. Proc. Natl. Acad. Sci. USA 2000, 97, 7016–7023. [Google Scholar] [CrossRef] [PubMed]
- McCubbin, A. Heteromorphic Self-Incompatibility in Primula: Twenty-First Century Tools Promise to Unravel a Classic Nineteeneth Centruy Model System; Springer: Berlin/Heidelberg, Germany, 2008; pp. 289–308. [Google Scholar]
- Nowak, M.D.; Russo, G.; Schlapbach, R.; Huu, C.N.; Lenhard, M.; Conti, E. The draft genome of Primula veris yields insights into the molecular basis of heterostyly. Genome Biol. 2015, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kofler, L. Über das Saponin der Primulawurzel. Arch. Pharm. 1924, 262, 318–328. [Google Scholar] [CrossRef]
- Wright, A.; Sticher, O. Triterpene saponins from Primula veris subsp. macrocalyx and Primula elatior subsp. meyeri. J. Nat. Prod. 1992, 55, 1299–1306. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Compound | tR (min) | λmax (nm) | m/z | HPLC-ESI-MS/MS Fragments, m/z | Plant Variants | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yellow Variant | Orange Variant | Red Variant | ||||||||||||
| p | l | r | p | l | r | p | l | r | ||||||
| Flavonoids | [M-H]− | |||||||||||||
| (+)-Catechin (A) | 23.7 | 228, 280 | 289 | 245, 203, 186 | + | − | − | + | − | − | + | − | − | st |
| Isorhamnetin | 50.3 | 254, 368 | 315 | 300, 271, 151 | − | − | − | + | − | − | + | − | − | st |
| Flavonoid glycosides | [M-H]− | |||||||||||||
| Quercetin-3-O-rutinoside (H) | 32.1 | 256, 356 | 609 | 301, 179, 151 | + | + | − | + | + | − | + | + | − | st |
| Quercetin-3-gentiobioside (E) | 28.3 | 256, 354 | 625 | 463, 301, 179, 151 | + | + | − | + | + | − | + | + | − | [15,16] |
| Quercetin-trihexoside (C) | 25.5 | 256, 358 | 787 | 625, 463, 301, 179 | + | + | − | + | + | − | + | + | − | [16] |
| Kaempferol-3-O-diglucoside-7-O-glucoside (D) | 29.4 | 270, 350 | 771 | 609, 285, 257, 151 | + | + | − | + | + | − | + | + | − | [17] |
| Kaempferol-3-O-rutinoside (J) | 34.4 | 264, 350 | 593 | 447, 285, 257, 229 | + | + | − | + | + | − | + | + | − | [15] |
| Kaempferol-3-O-galactoside-rhamnoside-7-O-rhamnoside (F) | 30.1 | 254, 356 | 739 | 575, 429, 282, 255 | + | + | − | + | + | − | + | + | − | [18] |
| Methylated flavonoid glycosides | [M-H]− | |||||||||||||
| Quercetin-methylether-rhamnoside-glucoside-rhamnoside (G) | 31.5 | 254, 354 | 769 | 737, 623, 605, 315, 300, 271 | + | + | − | + | + | − | + | + | − | [13] |
| Methyl-quercetin-dihexoside (I) | 33.2 | 254, 356 | 639 | 477, 315, 300, 271 | + | − | − | + | − | − | + | − | − | |
| Methyl-quercetin-rhamnoside-hexoside (K) | 37.6 | 254, 356 | 623 | 477, 315, 300, 271 | + | + | − | + | + | − | + | + | − | |
| Methyl-myricetin-trihexoside (B) | 24.3 | 278, 342 | 817 | 655, 493, 331, 316, 271 | + | − | − | + | − | − | + | − | − | |
| Anthocyanins | [M]+ | |||||||||||||
| Cyanidin-3-O-glucoside (M) | 18.1 | 280, 520 | 449 | 287, 231, 213 | − | * | * | + | * | * | + | * | * | st |
| Cyanidin-hexoside (L) | 17.3 | 280, 520 | 449 | 287, 231, 213 | − | * | * | + | * | * | + | * | * | [19] |
| Peonidin-hexoside (O1) | 19.6 | 280, 520 | 463 | 301, 286, 258 | − | * | * | + | * | * | + | * | * | [19] |
| Peonidin-dihexoside (O2) | 19.2 | 280, 520 | 625 | 301, 286, 258 | − | * | * | + | * | * | + | * | * | [19] |
| Malvidin-hexoside (P) | 20.2 | 260, 520 | 493 | 331, 315, 298, 281 | − | * | * | + | * | * | + | * | * | [19] |
| Triterpenoid saponins | [M-H]− | |||||||||||||
| Primula acid I | 53.1 | − | 1104 | 924, 465, 447, 246 | + | + | + | + | + | + | + | + | + | st |
| Primula acid II | 52.4 | − | 1236 | 924, 465 | − | − | + | − | − | + | − | − | + | [12] |
| Priverosaponin B-22-acetate | 52.1 | − | 1162 | 982, 465 | − | − | + | − | − | + | − | − | + | [12] |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apel, L.; Kammerer, D.R.; Stintzing, F.C.; Spring, O. Comparative Metabolite Profiling of Triterpenoid Saponins and Flavonoids in Flower Color Mutations of Primula veris L. Int. J. Mol. Sci. 2017, 18, 153. https://doi.org/10.3390/ijms18010153
Apel L, Kammerer DR, Stintzing FC, Spring O. Comparative Metabolite Profiling of Triterpenoid Saponins and Flavonoids in Flower Color Mutations of Primula veris L. International Journal of Molecular Sciences. 2017; 18(1):153. https://doi.org/10.3390/ijms18010153
Chicago/Turabian StyleApel, Lysanne, Dietmar R. Kammerer, Florian C. Stintzing, and Otmar Spring. 2017. "Comparative Metabolite Profiling of Triterpenoid Saponins and Flavonoids in Flower Color Mutations of Primula veris L." International Journal of Molecular Sciences 18, no. 1: 153. https://doi.org/10.3390/ijms18010153
APA StyleApel, L., Kammerer, D. R., Stintzing, F. C., & Spring, O. (2017). Comparative Metabolite Profiling of Triterpenoid Saponins and Flavonoids in Flower Color Mutations of Primula veris L. International Journal of Molecular Sciences, 18(1), 153. https://doi.org/10.3390/ijms18010153