
Advances and Future Applications of Augmented Peripheral Nerve Regeneration



Abstract
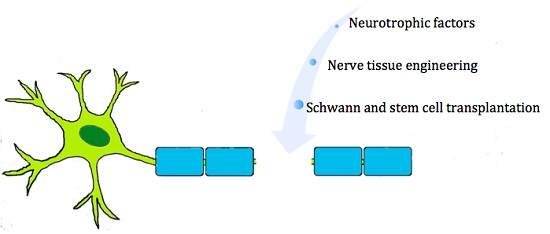
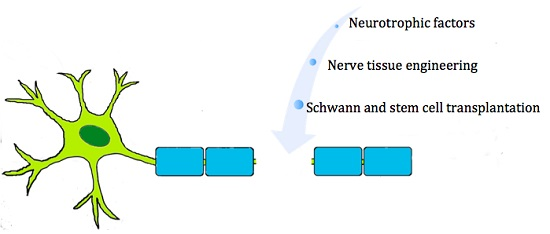
:
1. Introduction
2. Pathogenesis of Nerve Injury and Repair
3. Surgical Methods of Nerve Repair
4. Regulating Peripheral Nerve Regeneration
5. Exogenous Agents Improving Peripheral Nerve Regeneration
6. Schwann Cell Transplantation in Nerve Regeneration
7. Stem Cells in Nerve Regeneration
7.1. Embryonic Stem Cells versus Induced Pluripotent Stem Cells
7.2. Embryonic Stem Cells
7.3. Induced Pluripotent Stem Cells
7.4. Adipose Stem Cells
7.5. Neural Crest Stem Cells
7.6. Bone-Marrow Mesenchymal Stem Cells
8. Nerve Tissue Engineering
9. Future Directions
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ciaramitaro, P.; Mondelli, M.; Logullo, F.; Grimaldi, S.; Battiston, B.; Sard, A.; Scarinzi, C.; Migliaretti, G.; Faccani, G.; Cocito, D. Traumatic peripheral nerve injuries: Epidemiological findings, neuropathic pain and quality of life in 158 patients. J. Peripher. Nerv. Syst. 2010, 15, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Noble, J.; Munro, C.A.; Prasad, V.S.; Midha, R. Analysis of upper and lower extremity peripheral nerve injuries in a population of patients with multiple injuries. J. Trauma 1998, 45, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Braza, D.; Rice, J.B.; Dillingham, T. The incidence of peripheral nerve injury in extremity trauma. Am. J. Phys. Med. Rehabil. 2008, 87, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Bekelis, K.; Missios, S.; Spinner, R.J. Falls and peripheral nerve injuries: An age-dependent relationship. J. Neurosurg. 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Missios, S.; Bekelis, K.; Spinner, R.J. Traumatic peripheral nerve injuries in children: Epidemiology and socioeconomics. J. Neurosurg. Pediatr. 2014, 14, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Fontes, F.; Sonin, T.; Dias, T.; Fragoso, M.; Castro-Lopes, J.M.; Lunet, N. Chemotherapy-induced peripheral neuropathy after neoadjuvant or adjuvant treatment of breast cancer: A prospective cohort study. Support. Care Cancer 2015, 24, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Brull, R.; Hadzic, A.; Reina, M.A.; Barrington, M.J. Pathophysiology and etiology of nerve injury following peripheral nerve blockade. Reg. Anesth. Pain. Med. 2015, 40, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.G.; Huang, S.J.; Zhou, J.; Sun, J.Y.; Guo, H.; Li, F.Y.; Lin, Q.; Lin, H.X.; He, Z.Y. Dosimetric analysis of the brachial plexus among patients with breast cancer treated with post-mastectomy radiotherapy to the ipsilateral supraclavicular area: Report of 3 cases of radiation-induced brachial plexus neuropathy. Radiat. Oncol. 2014, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Coroneos, C.J.; Voineskos, S.H.; Coroneos, M.K.; Alolabi, N.; Goekjian, S.R.; Willoughby, L.I.; Farrokhyar, F.; Thoma, A.; Bain, J.R.; Brouwers, M.C. Obstetrical brachial plexus injury: Burden in a publicly funded, universal healthcare system. J. Neurosurg. Pediatr. 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Brininger, T.L.; Antczak, A.; Breland, H.L. Upper extremity injuries in the u.S. Military during peacetime years and wartime years. J. Hand. Ther. 2008, 21, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, A.L.; Mohrle, C.R.; Galarneau, M.R.; Woodruff, S.I.; Dye, J.L.; Quinn, K.H. Battlefield extremity injuries in operation iraqi freedom. Injury 2009, 40, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, P.N. A conditioning lesion induces changes in gene expression and axonal transport that enhance regeneration by increasing the intrinsic growth state of axons. Exp. Neurol. 2010, 223, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Hanz, S.; Fainzilber, M. Retrograde signaling in injured nerve—The axon reaction revisited. J. Neurochem. 2006, 99, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Michaelevski, I.; Segal-Ruder, Y.; Rozenbaum, M.; Medzihradszky, K.F.; Shalem, O.; Coppola, G.; Horn-Saban, S.; Ben-Yaakov, K.; Dagan, S.Y.; Rishal, I. Signaling to transcription networks in the neuronal retrograde injury response. Sci. Signal. 2010, 3. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.Y.; Gordon, T. The cellular and molecular basis of peripheral nerve regeneration. Mol. Neurobiol. 1997, 14, 67–116. [Google Scholar] [CrossRef] [PubMed]
- Maggi, S.P.; Lowe, J.B., 3rd; Mackinnon, S.E. Pathophysiology of nerve injury. Clin. Plast. Surg. 2003, 30, 109–126. [Google Scholar] [CrossRef]
- Waller, A. Experiments on the section of the glossopharyngeal and hypoglossal nerves of the frog, and observations of the alterations produced therby in the structure of their primitive fibres. Philos. Trans. R. Soc. Lond. 1850, 140, 423–429. [Google Scholar] [CrossRef]
- Geuna, S.; Raimondo, S.; Ronchi, G.; di Scipio, F.; Tos, P.; Czaja, K.; Fornaro, M. Chapter 3: Histology of the peripheral nerve and changes occurring during nerve regeneration. Int. Rev. Neurobiol. 2009, 87, 27–46. [Google Scholar] [PubMed]
- Lee, S.K.; Wolfe, S.W. Peripheral nerve injury and repair. J. Am. Acad. Orthop. Surg. 2000, 8, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Madl, C.M.; Heilshorn, S.C. Matrix interactions modulate neurotrophin-mediated neurite outgrowth and pathfinding. Neural Regen. Res. 2015, 10, 514–517. [Google Scholar] [PubMed]
- Hopker, V.H.; Shewan, D.; Tessier-Lavigne, M.; Poo, M.; Holt, C. Growth-cone attraction to netrin-1 is converted to repulsion by laminin-1. Nature 1999, 401, 69–73. [Google Scholar] [PubMed]
- Gaudet, A.D.; Popovich, P.G.; Ramer, M.S. Wallerian degeneration: Gaining perspective on inflammatory events after peripheral nerve injury. J. Neuroinflamm. 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.; Martini, R. Myelin and macrophages in the PNS: An intimate relationship in trauma and disease. Brain Res. 2015, 1641, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Brushart, T.M. Preferential reinnervation of motor nerves by regenerating motor axons. J. Neurosci. 1988, 8, 1026–1031. [Google Scholar] [PubMed]
- Joshi, A.R.; Bobylev, I.; Zhang, G.; Sheikh, K.A.; Lehmann, H.C. Inhibition of rho-kinase differentially affects axon regeneration of peripheral motor and sensory nerves. Exp. Neurol. 2015, 263, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Trojaborg, W. Rate of recovery in motor and sensory fibres of the radial nerve: Clinical and electrophysiological aspects. J. Neurol. Neurosurg. Psychiatry 1970, 33, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Zhen, G.; Puttgen, A.; Zhang, J.; Chen, T. Improved long-term recording of nerve signal by modified intrafascicular electrodes in rabbits. Microsurgery 2008, 28, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Lewitus, D.; Vogelstein, R.; Zhen, G.; Choi, Y.; Kohn, J.; Harshbarger, S.; Jia, X. Designing tyrosine-derived polycarbonate polymers for biodegradable regenerative type neural interface capable of neural recording. IEEE Trans. Neural Syst. Rehabil. Eng. 2011, 19, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Zhang, J.; Chen, T.; Chen, Z.; Zhu, Y.; Han, R.; Qiu, Z. Experimental study on harvesting the electric signal of peripheral nerve at rabbits by intrafascicular microelectrodes. Chin. J. Hand Surg. 2002, 18, 245–247. [Google Scholar]
- Jia, X.; Koenig, M.A.; Zhang, X.; Zhang, J.; Chen, T.; Chen, Z. Residual motor signal in long-term human severed peripheral nerves and feasibility of neural signal-controlled artificial limb. J. Hand Surg. 2007, 32, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Gamble, P.; Stephen, M.; MacEwan, M.; Ray, W.Z. Serial assessment of functional recovery following nerve injury utilizing implantable thin-film wireless nerve stimulators. Muscle Nerve 2016. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.; Tyreman, N.; Raji, M.A. The basis for diminished functional recovery after delayed peripheral nerve repair. J. Neurosci. 2011, 31, 5325–5334. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, S.; Wiberg, R.; McGrath, A.M.; Novikov, L.N.; Wiberg, M.; Novikova, L.N.; Kingham, P.J. Effect of delayed peripheral nerve repair on nerve regeneration, Schwann cell function and target muscle recovery. PLoS ONE 2013, 8, e56484. [Google Scholar] [CrossRef] [PubMed]
- Kemp, S.W.; Walsh, S.K.; Midha, R. Growth factor and stem cell enhanced conduits in peripheral nerve regeneration and repair. Neurol. Res. 2008, 30, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Menorca, R.M.; Fussell, T.S.; Elfar, J.C. Nerve physiology: Mechanisms of injury and recovery. Hand. Clin. 2013, 29, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Pabari, A.; Lloyd-Hughes, H.; Seifalian, A.M.; Mosahebi, A. Nerve conduits for peripheral nerve surgery. Plast. Reconstr. Surg. 2014, 133, 1420–1430. [Google Scholar] [CrossRef] [PubMed]
- Nichols, C.M.; Brenner, M.J.; Fox, I.K.; Tung, T.H.; Hunter, D.A.; Rickman, S.R.; Mackinnon, S.E. Effects of motor versus sensory nerve grafts on peripheral nerve regeneration. Exp. Neurol. 2004, 190, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Sabongi, R.G.; Fernandes, M.; Dos Santos, J.B. Peripheral nerve regeneration with conduits: Use of vein tubes. Neural Regen. Res. 2015, 10, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Battiston, B.; Geuna, S.; Ferrero, M.; Tos, P. Nerve repair by means of tubulization: Literature review and personal clinical experience comparing biological and synthetic conduits for sensory nerve repair. Microsurgery 2005, 25, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Siemionow, M.; Bozkurt, M.; Zor, F. Regeneration and repair of peripheral nerves with different biomaterials: Review. Microsurgery 2010, 30, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Zheng, R.; Wei, C.; Yan, J.; Ding, Y.; Wang, G.; Li, Z.; Zhang, Z. Tissue-engineered conduit promotes sciatic nerve regeneration following radiation-induced injury as monitored by magnetic resonance imaging. Magn. Reson. Imaging 2016, 34, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Manoli, T.; Schulz, L.; Stahl, S.; Jaminet, P.; Schaller, H.E. Evaluation of sensory recovery after reconstruction of digital nerves of the hand using muscle-in-vein conduits in comparison to nerve suture or nerve autografting. Microsurgery 2014, 34, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Jeon, W.J.; Kim, D.H.; Rhyu, I.J.; Kim, Y.H.; Youn, I.; Park, J.W. An inside-out vein graft filled with platelet-rich plasma for repair of a short sciatic nerve defect in rats. Neural Regen. Res. 2014, 9, 1351–1357. [Google Scholar] [PubMed]
- Sosa, I.; Reyes, O.; Kuffler, D.P. Immunosuppressants: Neuroprotection and promoting neurological recovery following peripheral nerve and spinal cord lesions. Exp. Neurol. 2005, 195, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.M.; MacEwan, M.; Santosa, K.B.; Chenard, K.E.; Ray, W.Z.; Hunter, D.A.; Mackinnon, S.E.; Johnson, P.J. Acellular nerve allografts in peripheral nerve regeneration: A comparative study. Muscle Nerve 2011, 44, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, E.L.; Tuffaha, S.H.; Luciano, J.P.; Yan, Y.; Hunter, D.A.; Magill, C.K.; Moore, A.M.; Tong, A.Y.; Mackinnon, S.E.; Borschel, G.H. Processed allografts and type I collagen conduits for repair of peripheral nerve gaps. Muscle Nerve 2009, 39, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Taras, J.S.; Jacoby, S.M.; Lincoski, C.J. Reconstruction of digital nerves with collagen conduits. J. Hand. Surg. Am. 2011, 36, 1441–1446. [Google Scholar] [CrossRef] [PubMed]
- Wangensteen, K.J.; Kalliainen, L.K. Collagen tube conduits in peripheral nerve repair: A retrospective analysis. Hand 2010, 5, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Ashley, W.W., Jr.; Weatherly, T.; Park, T.S. Collagen nerve guides for surgical repair of brachial plexus birth injury. J. Neurosurg. 2006, 105, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Boeckstyns, M.E.; Sorensen, A.I.; Vineta, J.F.; Rosen, B.; Navarro, X.; Archibald, S.J.; Valss-Sole, J.; Moldovan, M.; Krarup, C. Collagen conduit versus microsurgical neurorrhaphy: 2-year follow-up of a prospective, blinded clinical and electrophysiological multicenter randomized, controlled trial. J. Hand Surg. Am. 2013, 38, 2405–2411. [Google Scholar] [CrossRef] [PubMed]
- Rosson, G.D.; Williams, E.H.; Dellon, A.L. Motor nerve regeneration across a conduit. Microsurgery 2009, 29, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.F.; Kakinoki, R.; Rizzo, M.; Kang, W. Extrusion of a neurotube: A case report. Ochsner J. 2015, 15, 191–192. [Google Scholar] [PubMed]
- Schlosshauer, B.; Dreesmann, L.; Schaller, H.E.; Sinis, N. Synthetic nerve guide implants in humans: A comprehensive survey. Neurosurgery 2006, 59, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Bertleff, M.J.; Meek, M.F.; Nicolai, J.P. A prospective clinical evaluation of biodegradable neurolac nerve guides for sensory nerve repair in the hand. J. Hand Surg. Am. 2005, 30, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Meek, M.F.; Den Dunnen, W.F. Porosity of the wall of a neurolac nerve conduit hampers nerve regeneration. Microsurgery 2009, 29, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Cortes, P.; Garrido, J.; Camara, M.; Ravassa, F.O. Failed digital nerve reconstruction by foreign body reaction to neurolac nerve conduit. Microsurgery 2010, 30, 414–416. [Google Scholar] [PubMed]
- Chiriac, S.; Facca, S.; Diaconu, M.; Gouzou, S.; Liverneaux, P. Experience of using the bioresorbable copolyester poly(dl-lactide-epsilon-caprolactone) nerve conduit guide Neurolac™ for nerve repair in peripheral nerve defects: Report on a series of 28 lesions. J. Hand. Surg. Eur. Vol. 2012, 37, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Weber, R.A.; Breidenbach, W.C.; Brown, R.E.; Jabaley, M.E.; Mass, D.P. A randomized prospective study of polyglycolic acid conduits for digital nerve reconstruction in humans. Plast. Reconstr. Surg. 2000, 106, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Shin, R.H.; Friedrich, P.F.; Crum, B.A.; Bishop, A.T.; Shin, A.Y. Treatment of a segmental nerve defect in the rat with use of bioabsorbable synthetic nerve conduits: A comparison of commercially available conduits. J. Bone Jt. Surg. Am. 2009, 91, 2194–2204. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, S.E.; Dellon, A.L. A study of nerve regeneration across synthetic (maxon) and biologic (collagen) nerve conduits for nerve gaps up to 5 cm in the primate. J. Reconstr. Microsurg. 1990, 6, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Krarup, C.; Archibald, S.J.; Madison, R.D. Factors that influence peripheral nerve regeneration: An electrophysiological study of the monkey median nerve. Ann. Neurol. 2002, 51, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, J.; Browne, T. Overcoming short gaps in peripheral nerve repair: Conduits and human acellular nerve allograft. Hand 2014, 9, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Kuffler, D.P. Guidance of regenerating motor axons in vivo by gradients of diffusible peripheral nerve-derived factors. J. Neurobiol. 2000, 42, 212–219. [Google Scholar] [CrossRef]
- Boyd, J.G.; Gordon, T. Neurotrophic factors and their receptors in axonal regeneration and functional recovery after peripheral nerve injury. Mol. Neurobiol. 2003, 27, 277–324. [Google Scholar] [CrossRef]
- Griesbeck, O.; Parsadanian, A.S.; Sendtner, M.; Thoenen, H. Expression of neurotrophins in skeletal muscle: Quantitative comparison and significance for motoneuron survival and maintenance of function. J. Neurosci. Res. 1995, 42, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Barker, P.A. Neurotrophin signaling through the p75 neurotrophin receptor. Prog. Neurobiol. 2002, 67, 203–233. [Google Scholar] [CrossRef]
- English, A.W.; Wilhelm, J.C.; Ward, P.J. Exercise, neurotrophins, and axon regeneration in the PNS. Physiology 2014, 29, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.N.; Lancaster, K.Z.; Zhen, G.; He, J.; Gupta, M.K.; Kong, Y.L.; Engel, E.A.; Krick, K.D.; Ju, A.; Meng, F.; et al. 3D printed anatomical nerve regeneration pathways. Adv. Funct. Mater. 2015, 25, 6205–6217. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Shoichet, M.S. Defining the concentration gradient of nerve growth factor for guided neurite outgrowth. Neuroscience 2001, 103, 831–840. [Google Scholar] [CrossRef]
- Cornejo, M.; Nambi, D.; Walheim, C.; Somerville, M.; Walker, J.; Kim, L.; Ollison, L.; Diamante, G.; Vyawahare, S.; de Bellard, M.E. Effect of NRG1, GDNF, EGF and NGF in the migration of a Schwann cell precursor line. Neurochem. Res. 2010, 35, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, M.P.; Cornuet, P.K.; McIlwrath, S.; Koerber, H.R.; Albers, K.M. Sry-box containing gene 11 (sox11) transcription factor is required for neuron survival and neurite growth. Neuroscience 2006, 143, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, M.P.; McIlwrath, S.L.; Jing, X.; Cornuet, P.K.; Salerno, K.M.; Koerber, H.R.; Albers, K.M. Sox11 transcription factor modulates peripheral nerve regeneration in adult mice. Brain Res. 2009, 1256, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Wang, T.; Huang, S.; Glorioso, J.C.; Albers, K.M. The transcription factor Sox11 promotes nerve regeneration through activation of the regeneration-associated gene Sprr1a. Exp. Neurol. 2012, 233, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, I.E.; Tanabe, K.; Strittmatter, S.M. Small proline-rich repeat protein 1A is expressed by axotomized neurons and promotes axonal outgrowth. J. Neurosci. 2002, 22, 1303–1315. [Google Scholar] [PubMed]
- Salerno, K.M.; Jing, X.; Diges, C.M.; Davis, B.M.; Albers, K.M. TRAF family member-associated NF-kappa B activator (TANK) expression increases in injured sensory neurons and is transcriptionally regulated by Sox11. Neuroscience 2013, 231, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.J.; McCord, A.E.; Greenberg, M.E. A biological function for the neuronal activity-dependent component of bdnf transcription in the development of cortical inhibition. Neuron 2008, 60, 610–624. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, J.C.; Xu, M.; Cucoranu, D.; Chmielewski, S.; Holmes, T.; Lau, K.S.; Bassell, G.J.; English, A.W. Cooperative roles of BDNF expression in neurons and Schwann cells are modulated by exercise to facilitate nerve regeneration. J. Neurosci. 2012, 32, 5002–5009. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Lee, V.M.; Wang, Y.; Lin, J.S.; Sock, E.; Wegner, M.; Lei, L. Sox11 regulates survival and axonal growth of embryonic sensory neurons. Dev. Dyn. 2011, 240, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Fricker, F.R.; Bennett, D.L. The role of neuregulin-1 in the response to nerve injury. Future Neurol. 2011, 6, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Birchmeier, C.; Nave, K.A. Neuregulin-1, a key axonal signal that drives Schwann cell growth and differentiation. Glia 2008, 56, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Stassart, R.M.; Fledrich, R.; Velanac, V.; Brinkmann, B.G.; Schwab, M.H.; Meijer, D.; Sereda, M.W.; Nave, K.A. A role for Schwann cell-derived neuregulin-1 in remyelination. Nat. Neurosci. 2013, 16, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Faroni, A.; Mobasseri, S.A.; Kingham, P.J.; Reid, A.J. Peripheral nerve regeneration: Experimental strategies and future perspectives. Adv. Drug. Deliv. Rev. 2015, 82, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, C.K.; Grecco, M.A.; Leite, V.M.; Faloppa, F. Fibroblasts growth factor and nerve fragments effect on tibial nerve regeneration in rats: A comparative study. Acta Ortop. Bras. 2007, 15, 114–117. [Google Scholar] [CrossRef]
- Marquardt, L.M.; Ee, X.; Iyer, N.; Hunter, D.; Mackinnon, S.E.; Wood, M.D.; Sakiyama-Elbert, S.E. Finely tuned temporal and spatial delivery of GDNF promotes enhanced nerve regeneration in a long nerve defect model. Tissue Eng. Part A 2015, 21, 2852–2864. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, R.; Amini, K.; Eskafian, H. Betamethasone-enhanced vein graft conduit accelerates functional recovery in the rat sciatic nerve gap. J. Oral Maxillofac. Surg. 2013, 71, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Azizi, A.; Azizi, S.; Heshmatian, B.; Amini, K. Improvement of functional recovery of transected peripheral nerve by means of chitosan grafts filled with vitamin E, pyrroloquinoline quinone and their combination. Int. J. Surg. 2014, 12, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, R.; Amini, K.; Yousefi, A.; Abdollahi-Pirbazari, M.; Belbasi, A.; Abedi, F. Functional effects of local administration of thyroid hormone combined with chitosan conduit after sciatic nerve transection in rats. J. Oral Maxillofac. Surg. 2013, 71, 1763–1776. [Google Scholar] [CrossRef] [PubMed]
- Elfar, J.C.; Jacobson, J.A.; Puzas, J.E.; Rosier, R.N.; Zuscik, M.J. Erythropoietin accelerates functional recovery after peripheral nerve injury. J. Bone Jt. Surg. Am. 2008, 90, 1644–1653. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Zhu, C.; Yin, J.B.; Zhang, T.; Lu, Y.C.; Ren, J.; Li, Y.Q. Slow-releasing rapamycin-coated bionic peripheral nerve scaffold promotes the regeneration of rat sciatic nerve after injury. Life Sci. 2015, 122, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Gold, B.G.; Katoh, K.; Storm-Dickerson, T. The immunosuppressant FK506 increases the rate of axonal regeneration in rat sciatic nerve. J. Neurosci. 1995, 15, 7509–7516. [Google Scholar] [PubMed]
- Shahraki, M.; Mohammadi, R.; Najafpour, A. Influence of tacrolimus (FK506) on nerve regeneration using allografts: A rat sciatic nerve model. J. Oral Maxillofac. Surg. 2015, 73, 1438.e1–1438.e9. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.S.; Zeleny-Pooley, M.; Gold, B.G. Comparative dose-dependence study of FK506 and cyclosporin a on the rate of axonal regeneration in the rat sciatic nerve. J. Pharmacol. Exp. Ther. 1997, 282, 1084–1093. [Google Scholar] [PubMed]
- Price, R.D.; Yamaji, T.; Yamamoto, H.; Higashi, Y.; Hanaoka, K.; Yamazaki, S.; Ishiye, M.; Aramori, I.; Matsuoka, N.; Mutoh, S.; et al. Fk1706, a novel non-immunosuppressive immunophilin: Neurotrophic activity and mechanism of action. Eur. J. Pharmacol. 2005, 509, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Gold, B.G.; Zeleny-Pooley, M.; Wang, M.S.; Chaturvedi, P.; Armistead, D.M. A nonimmunosuppressant FKBP-12 ligand increases nerve regeneration. Exp. Neurol. 1997, 147, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Gold, B.G.; Densmore, V.; Shou, W.; Matzuk, M.M.; Gordon, H.S. Immunophilin FK506-binding protein 52 (not FK506-binding protein 12) mediates the neurotrophic action of FK506. J. Pharmacol. Exp. Ther. 1999, 289, 1202–1210. [Google Scholar] [PubMed]
- Hoben, G.; Yan, Y.; Iyer, N.; Newton, P.; Hunter, D.A.; Moore, A.M.; Sakiyama-Elbert, S.E.; Wood, M.D.; Mackinnon, S.E. Comparison of acellular nerve allograft modification with Schwann cells or vegf. Hand 2015, 10, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Strauch, B.; Rodriguez, D.M.; Diaz, J.; Yu, H.L.; Kaplan, G.; Weinstein, D.E. Autologous Schwann cells drive regeneration through a 6-cm autogenous venous nerve conduit. J. Reconstr. Microsurg. 2001, 17, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Mosahebi, A.; Fuller, P.; Wiberg, M.; Terenghi, G. Effect of allogeneic Schwann cell transplantation on peripheral nerve regeneration. Exp. Neurol. 2002, 173, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Casella, G.T.; Bunge, R.P.; Wood, P.M. Improved method for harvesting human Schwann cells from mature peripheral nerve and expansion in vitro. Glia 1996, 17, 327–338. [Google Scholar] [CrossRef]
- Dilwali, S.; Patel, P.B.; Roberts, D.S.; Basinsky, G.M.; Harris, G.J.; Emerick, K.S.; Stankovic, K.M. Primary culture of human schwann and Schwannoma cells: Improved and simplified protocol. Hear. Res. 2014, 315, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.L.; Lassner, F.; Becker, M.; Walter, G.F.; Bader, A.; Berger, A. Viability of cultured nerve grafts: An assessment of proliferation of Schwann cells and fibroblasts. Microsurgery 1999, 19, 356–363. [Google Scholar] [CrossRef]
- Levi, A.D.; Burks, S.S.; Anderson, K.D.; Dididze, M.; Khan, A.; Dietrich, W.D. The use of autologous schwann cells to supplement sciatic nerve repair with a large gap—First in human experience. Cell Transplant. 2016, 25, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Amoh, Y.; Kanoh, M.; Niiyama, S.; Hamada, Y.; Kawahara, K.; Sato, Y.; Hoffman, R.M.; Katsuoka, K. Human hair follicle pluripotent stem (hfPS) cells promote regeneration of peripheral-nerve injury: An advantageous alternative to ES and iPS cells. J. Cell. Biochem. 2009, 107, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Ou, Y.C.; Liao, S.L.; Chen, W.Y.; Chen, S.Y.; Wu, C.W.; Wang, C.C.; Wang, W.Y.; Huang, Y.S.; Hsu, S.H. Transplantation of bone marrow stromal cells for peripheral nerve repair. Exp. Neurol. 2007, 204, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.; Lacy, T.; Hearn, J. The origins of human embryonic stem cell research policies in the us states. Sci. Public Policy 2013, 40, 544–558. [Google Scholar] [CrossRef]
- Kington, R. 2009 Guidelines on Stem Cell Research. Available online: http://stemcells.nih.gov/policy/pages/2009guidelines.aspx (accessed on 28 June 2016).
- Salmikangas, P.; Menezes-Ferreira, M.; Reischl, I.; Tsiftsoglou, A.; Kyselovic, J.; Borg, J.J.; Ruiz, S.; Flory, E.; Trouvin, J.H.; Celis, P.; et al. Manufacturing, characterization and control of cell-based medicinal products: Challenging paradigms toward commercial use. Regen. Med. 2015, 10, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.; Sato, D.; Sahara, Y. New governmental regulatory system for stem cell-based therapies in japan. Ther. Innov. Regul. Sci. 2014, 48, 681–688. [Google Scholar] [CrossRef]
- Ziegler, L.; Grigoryan, S.; Yang, I.H.; Thakor, N.V.; Goldstein, R.S. Efficient generation of schwann cells from human embryonic stem cell-derived neurospheres. Stem Cell Rev. 2011, 7, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Jiang, J.; Wei, L.; Zhou, X.; Fraser, J.L.; Snider, B.J.; Yu, S.P. Transplantation of embryonic stem cells improves nerve repair and functional recovery after severe sciatic nerve axotomy in rats. Stem Cells 2008, 26, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Uemura, T.; Takamatsu, K.; Okada, M.; Kazuki, K.; Tabata, Y.; Ikada, Y.; Nakamura, H. Acceleration of peripheral nerve regeneration using nerve conduits in combination with induced pluripotent stem cell technology and a basic fibroblast growth factor drug delivery system. J. Biomed. Mater. Res. A 2014, 102, 1370–1378. [Google Scholar] [CrossRef] [PubMed]
- Faroni, A.; Smith, R.J.; Lu, L.; Reid, A.J. Human Schwann-like cells derived from adipose-derived mesenchymal stem cells rapidly de-differentiate in the absence of stimulating medium. Eur. J. Neurosci. 2016, 43, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Gong, K.; Zheng, Z.; Wang, A.; Ao, Q.; Gong, Y.; Zhang, X. Chitosan/silk fibroin-based tissue-engineered graft seeded with adipose-derived stem cells enhances nerve regeneration in a rat model. J. Mater. Sci. Mater. Med. 2011, 22, 1947–1964. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Ao, Q.; Wei, Y.; Song, J. Transplantation of miRNA-34a overexpressing adipose-derived stem cell enhances rat nerve regeneration. Wound Repair Regen. 2016, 24, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Kingham, P.J.; Kolar, M.K.; Novikova, L.N.; Novikov, L.N.; Wiberg, M. Stimulating the neurotrophic and angiogenic properties of human adipose-derived stem cells enhances nerve repair. Stem Cells Dev. 2014, 23, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Sowa, Y.; Kishida, T.; Imura, T.; Numajiri, T.; Nishino, K.; Tabata, Y.; Mazda, O. Adipose-derived stem cells promote peripheral nerve regeneration in vivo without differentiation into schwann-like lineage. Plast. Reconstr. Surg. 2016, 137, 318e–330e. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Madura, T.; Sakai, Y.; Yano, K.; Terenghi, G.; Hosokawa, K. Glial differentiation of human adipose-derived stem cells: Implications for cell-based transplantation therapy. Neuroscience 2013, 236, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Liu, F.; Zhang, C.; Zhang, Z.; Guo, J.; Ren, C.; Kong, Z. Pluripotent hair follicle neural crest stem-cell-derived neurons and schwann cells functionally repair sciatic nerves in rats. Mol. Neurobiol. 2009, 40, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Grimoldi, N.; Colleoni, F.; Tiberio, F.; Vetrano, I.G.; Cappellari, A.; Costa, A.; Belicchi, M.; Razini, P.; Giordano, R.; Spagnoli, D.; et al. Stem cell salvage of injured peripheral nerve. Cell. Transplant. 2015, 24, 213–222. [Google Scholar] [PubMed]
- Cuevas, P.; Carceller, F.; Dujovny, M.; Garcia-Gomez, I.; Cuevas, B.; Gonzalez-Corrochano, R.; Diaz-Gonzalez, D.; Reimers, D. Peripheral nerve regeneration by bone marrow stromal cells. Neurol. Res. 2002, 24, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, P.; Carceller, F.; Garcia-Gomez, I.; Yan, M.; Dujovny, M. Bone marrow stromal cell implantation for peripheral nerve repair. Neurol. Res. 2004, 26, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, A.; Mohammadi, R.; Amini, K. Transplantation of undifferentiated bone-marrow stromal cells into a vein graft accelerates sciatic nerve regeneration in streptozotocin induced diabetic rats. Curr. Neurovasc. Res. 2014, 11, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, R.; Azizi, S.; Delirezh, N.; Hobbenaghi, R.; Amini, K.; Malekkhetabi, P. The use of undifferentiated bone marrow stromal cells for sciatic nerve regeneration in rats. Int. J. Oral Maxillofac. Surg. 2012, 41, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Yu, Z.; Li, J.; Dang, X.; Wang, K. Schwann-like cells seeded in acellular nerve grafts improve nerve regeneration. BMC Musculoskelet. Disord. 2014, 15, 165. [Google Scholar] [CrossRef] [PubMed]
- Narsinh, K.H.; Plews, J.; Wu, J.C. Comparison of human induced pluripotent and embryonic stem cells: Fraternal or identical twins? Mol. Ther. 2011, 19, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Kim, H.; Elkabetz, Y.; Al Shamy, G.; Panagiotakos, G.; Barberi, T.; Tabar, V.; Studer, L. Isolation and directed differentiation of neural crest stem cells derived from human embryonic stem cells. Nat. Biotechnol. 2007, 25, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Chambers, S.M.; Tomishima, M.J.; Studer, L. Derivation of neural crest cells from human pluripotent stem cells. Nat. Protoc. 2010, 5, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Ichisaka, T.; Yamanaka, S. Generation of germline-competent induced pluripotent stem cells. Nature 2007, 448, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Yoneda, Y. The timing of retroviral silencing correlates with the quality of induced pluripotent stem cell lines. Biochim. Biophys. Acta 2011, 1810, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Agostini, M.; Tucci, P.; Steinert, J.R.; Shalom-Feuerstein, R.; Rouleau, M.; Aberdam, D.; Forsythe, I.D.; Young, K.W.; Ventura, A.; Concepcion, C.P. MicroRNA-34a regulates neurite outgrowth, spinal morphology, and function. Proc. Natl. Acad. Sci. USA 2011, 108, 21099–21104. [Google Scholar] [CrossRef] [PubMed]
- Shyamala, K.; Yanduri, S.; Girish, H.C.; Murgod, S. Neural crest: The fourth germ layer. J. Oral Maxillofac. Pathol. 2015, 19, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lim, H.; Li, Z.; Oh, Y.; Kovlyagina, I.; Choi, I.Y.; Dong, X.; Lee, G. Generation of multipotent induced neural crest by direct reprogramming of human postnatal fibroblasts with a single transcription factor. Cell Stem Cell 2014, 15, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Krishnan, U.M.; Sethuraman, S. Development of biomaterial scaffold for nerve tissue engineering: Biomaterial mediated neural regeneration. J. Biomed. Sci. 2009, 16, 108. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Murugan, R.; Ramakrishna, S.; Wang, X.; Ma, Y.X.; Wang, S. Fabrication of nano-structured porous plla scaffold intended for nerve tissue engineering. Biomaterials 2004, 25, 1891–1900. [Google Scholar] [CrossRef] [PubMed]
- Prabhakaran, M.P.; Venugopal, J.R.; Ramakrishna, S. Mesenchymal stem cell differentiation to neuronal cells on electrospun nanofibrous substrates for nerve tissue engineering. Biomaterials 2009, 30, 4996–5003. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Zhu, Q.; Liu, X.; Yang, W.; Jian, Y.; Zhou, X.; He, B.; Gu, L.; Yan, L.; Lin, T.; et al. Three-dimensional reconstruction of the microstructure of human acellular nerve allograft. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Product | Material | Company | Clinical or Preclinical | Comment |
|---|---|---|---|---|
| NeuraGen® | Collagen Type I | Integra LifeSciences Co., Plainsboro, NJ, USA | Taras et al. † [47] Wangensteen et al. † [48] Ashley et al. † [49] Whitlock et al. [46] | Absorbable. Good clinical data for sensory and motor recovery. |
| NeuraWrap™ | Collagen Type I | Integra LifeSciences Co., Plainsboro, NJ, USA | n/a | Protective wrap. |
| NeuroFlex™ | Collagen Type I | Collagen Matrix, Inc., Franklin Lakes, NJ, USA | n/a | Flexible. For gaps 2.5 cm or less. |
| NeuroMatrix™ | Collagen Type I | Collagen Matrix, Inc., Franklin Lakes, NJ, USA | n/a | For gaps 2.5 cm or less. |
| NeuroMend™ | Collagen Type I | Collagen Matrix, Inc., Franklin Lakes, NJ, USA | n/a | Self curling protective wrap. |
| NeuroTube® | Polyglycolic acid | Synovis Micro Companies Alliance, Birmingham, AL, USA | Battiston et al. † [39] Rosson et al. † [51] Duncan et al. † [52] | Absorbable. Acidic degradation limits quantity used [53] Clinical data available for sensory and motor recovery. Risk of extrusion. |
| Neurolac® | Poly(d,l-lactide-co-ε-caprolactone) | Polyganics BV, Groningen, The Netherlands | Bertleff et al. † [54] Meek et al. [55] Hernandez et al † [56] Chiriac et al. † [57] | Absorbable. For gaps less than 20 mm. Risk of foreign body reaction. |
| Salutunnel™ | Polyvinyl alcohol hydrogel | Salumedica LLC, Atlanta, GA, USA | n/a | Non-degradable. |
| AxoGuard™ Nerve Connector | Porcine intestinal submucosa | Cook Biotech, Inc., West Lafayette, IN, USA | n/a | For gaps less than 5 mm. |
| Stem Cell by Source | Advantage | Disadvantages | Clinical or Preclinical Data | Mechanisms of Regenerative Benefit |
|---|---|---|---|---|
| Embryonic | Totipotent. | Regulatory restrictions. Ethical concerns. Risk of teratoma. Limited availability. | Ziegler et al. [110] Cui et al. [111] | Schwann Cells, GDNF, NGF, FGF, BDNF, angiopoietin-1. 35%–60% yield differentiation to Schwann cells [110,111]. |
| iPSC | Inducible from easily obtainable somatic cells. | Genetic manipulation required. Potential risk of teratoma. | Ikeda et al. [112] | Differentiation to Schwann cells. |
| Adipose | Ease of harvest. Widely available. | Variability of results. Require stimulation for neurotrophic effect [113]. | Wei et al. [114] He et al. [115] Kingham et al. [116] Sowa et al. [117] Tomita et al. [118] | Production of neurotrophic and angiogenic factors [116,118]. |
| Neural crest | Ease of harvest from skin and follicle cells. Can be induced from fibroblasts, hESC, iPSC. | - | Lin et al. [119] Amoh et al. [104] Grimoldi et al.† [120] | Differentiate into neurons and SC. |
| Bone marrow | Familiarity with harvesting. | Bone marrow biopsy required. Heterogeneity of cells. | Cuevas et al. [121,122] Chen et al. [105] Mohammadi et al. [123,124] Fan et al. [125] | Fibroblast-like cell differentiation. Increase production of myelin basic protein, NGF, BDNF, GDNF, CNTF. |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, S.; Eisenberg, H.M.; Jia, X. Advances and Future Applications of Augmented Peripheral Nerve Regeneration. Int. J. Mol. Sci. 2016, 17, 1494. https://doi.org/10.3390/ijms17091494
Jones S, Eisenberg HM, Jia X. Advances and Future Applications of Augmented Peripheral Nerve Regeneration. International Journal of Molecular Sciences. 2016; 17(9):1494. https://doi.org/10.3390/ijms17091494
Chicago/Turabian StyleJones, Salazar, Howard M. Eisenberg, and Xiaofeng Jia. 2016. "Advances and Future Applications of Augmented Peripheral Nerve Regeneration" International Journal of Molecular Sciences 17, no. 9: 1494. https://doi.org/10.3390/ijms17091494
APA StyleJones, S., Eisenberg, H. M., & Jia, X. (2016). Advances and Future Applications of Augmented Peripheral Nerve Regeneration. International Journal of Molecular Sciences, 17(9), 1494. https://doi.org/10.3390/ijms17091494