
Effect of Thyrotropin on Osteopontin, Integrin αvβ3, and VCAM-1 in the Endothelium via Activation of Akt

Abstract
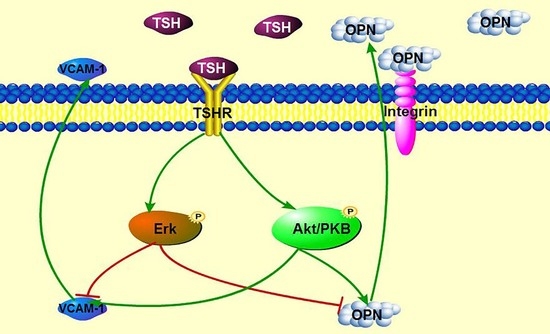
:
1. Introduction
2. Results
2.1. Hormone Levels in Rats of Three Groups of Rats
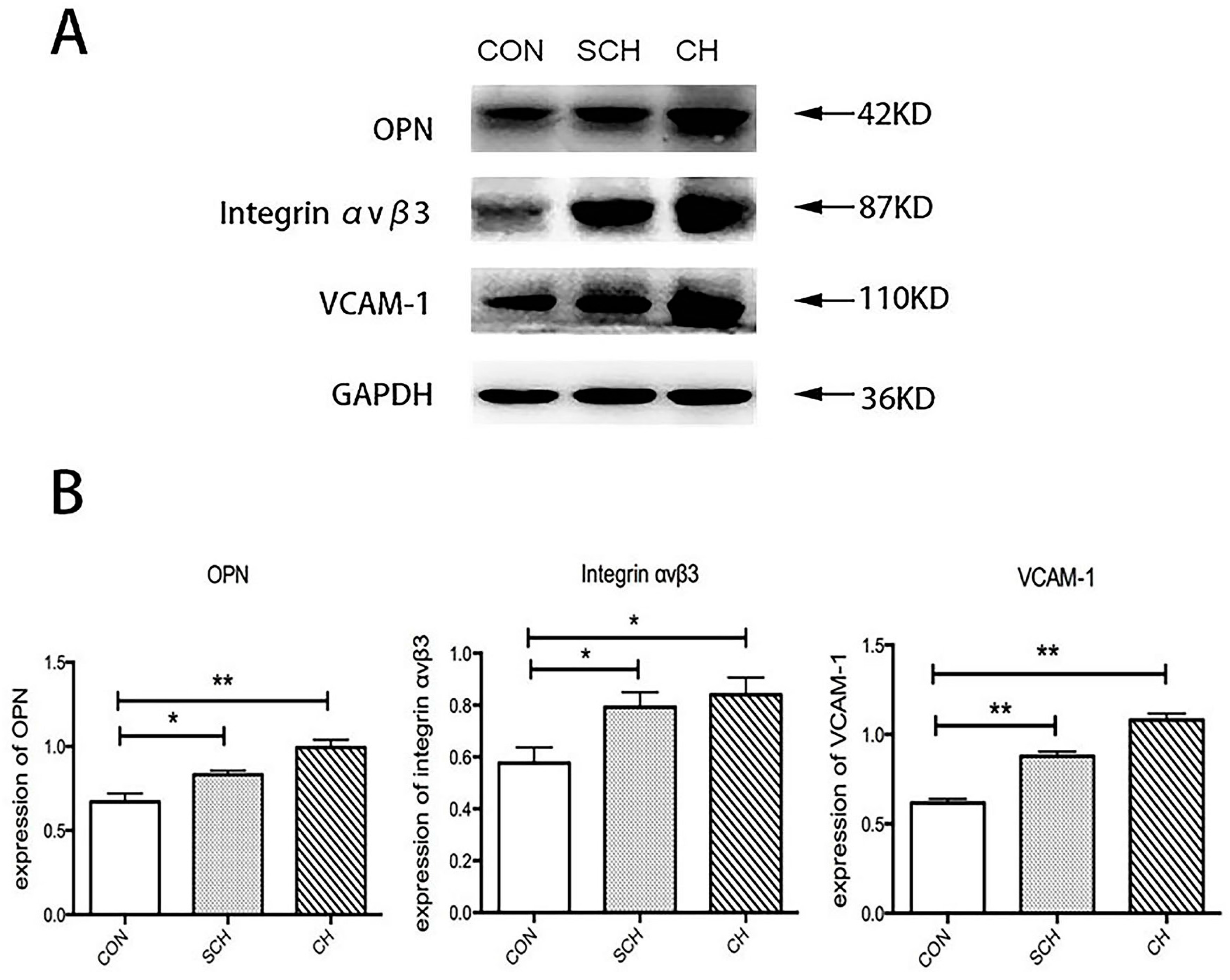
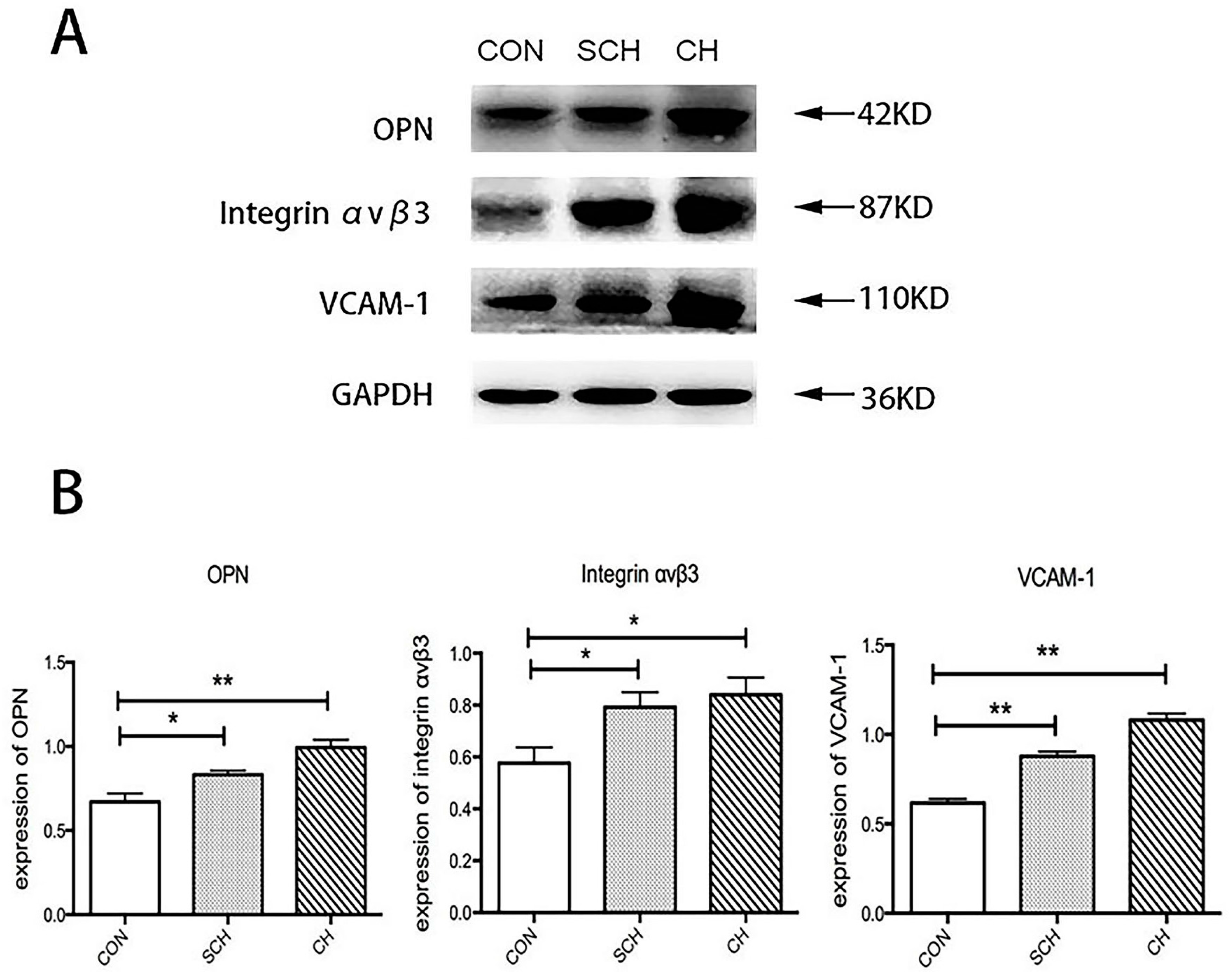
2.2. Western Blot Analysis of Osteopontin (OPN), Integrin αvβ3, and Vascular Cell Adhesion Molecule (VCAM-1) Expression of Aorta Tissues from CH, SCH, and CON Groups
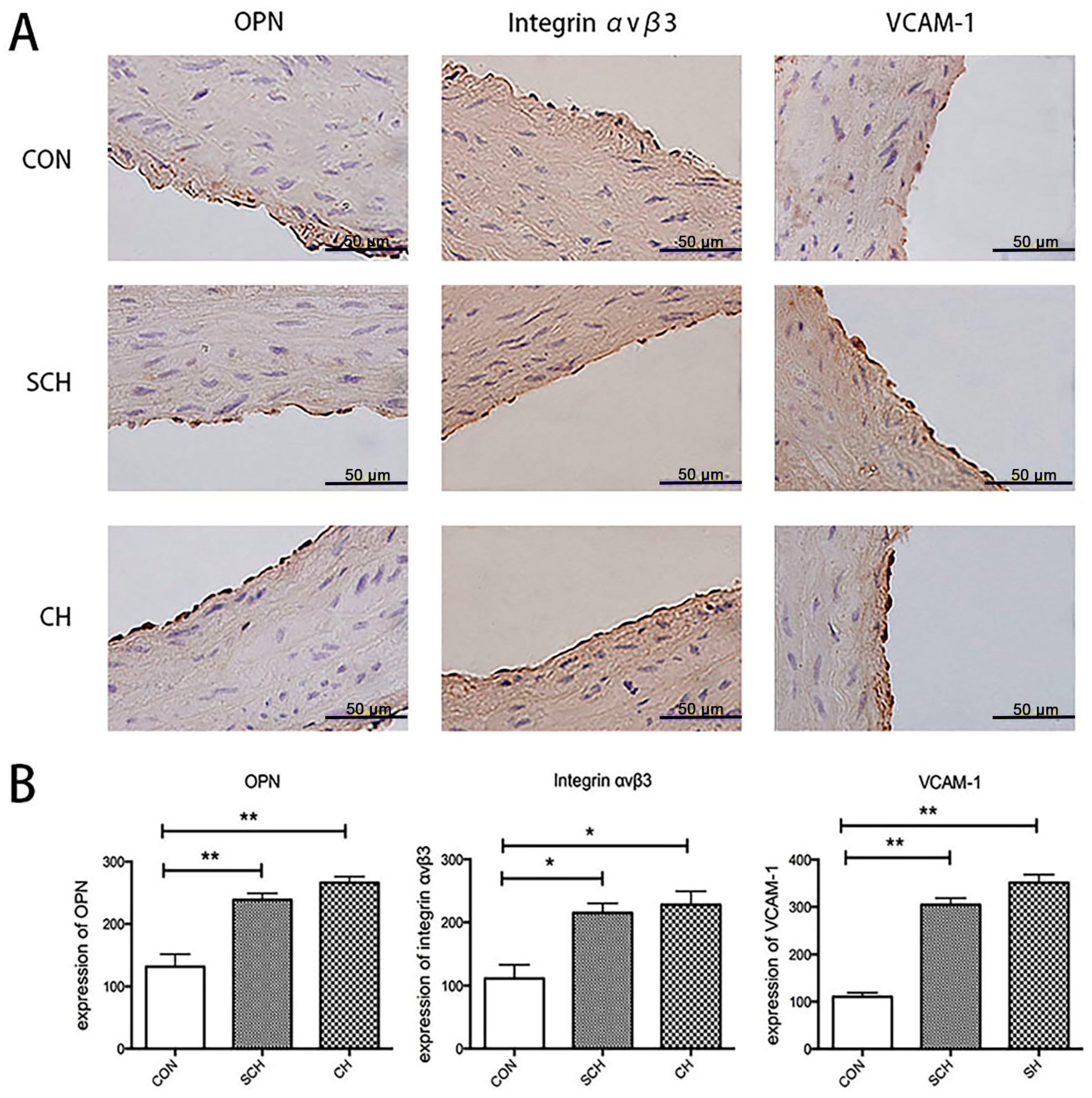
2.3. Immunohistochemical Analysis of OPN, Integrin αvβ3, and VCAM-1 Expression of Aorta Endothelium from CH, SCH, and CON Groups
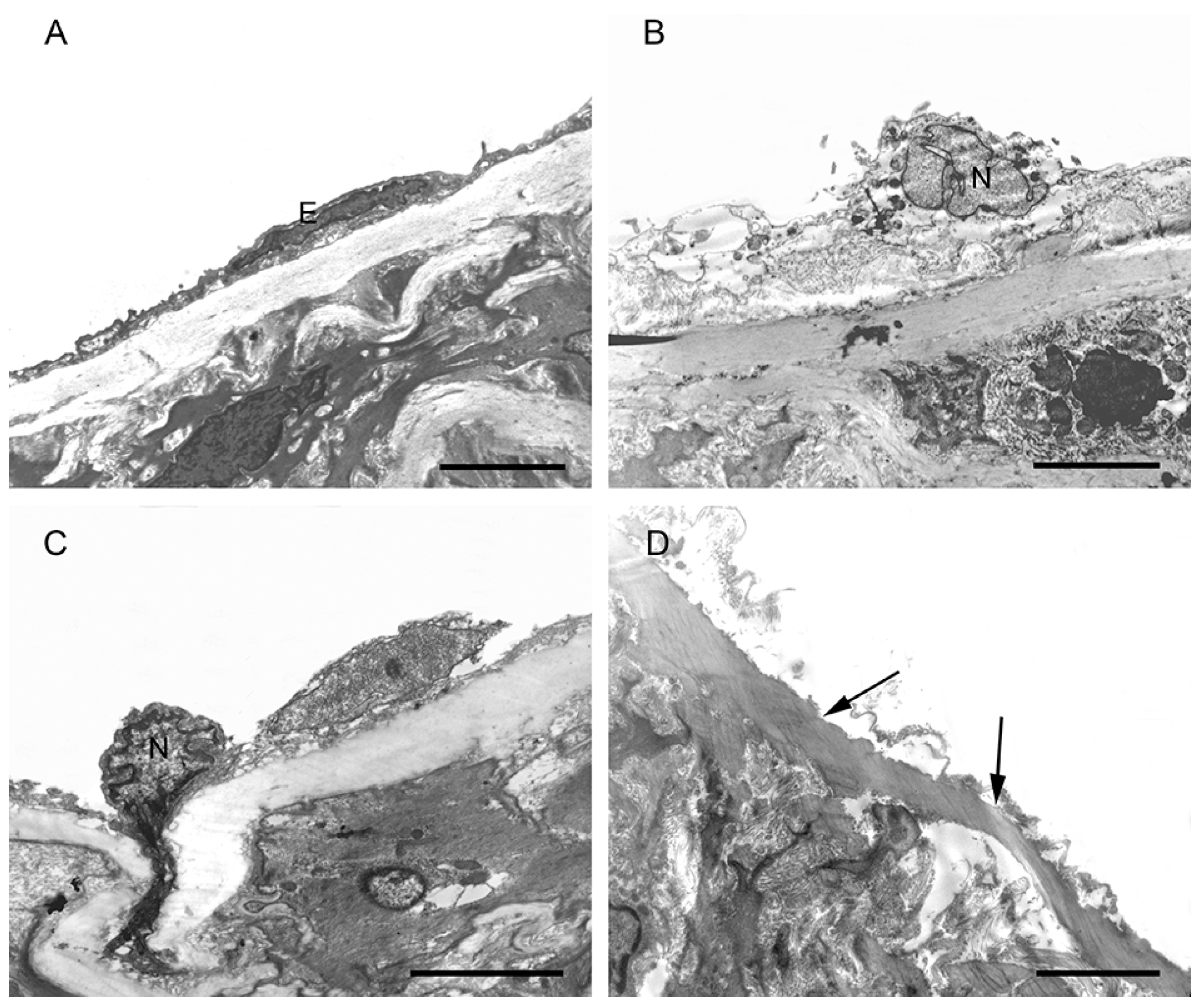
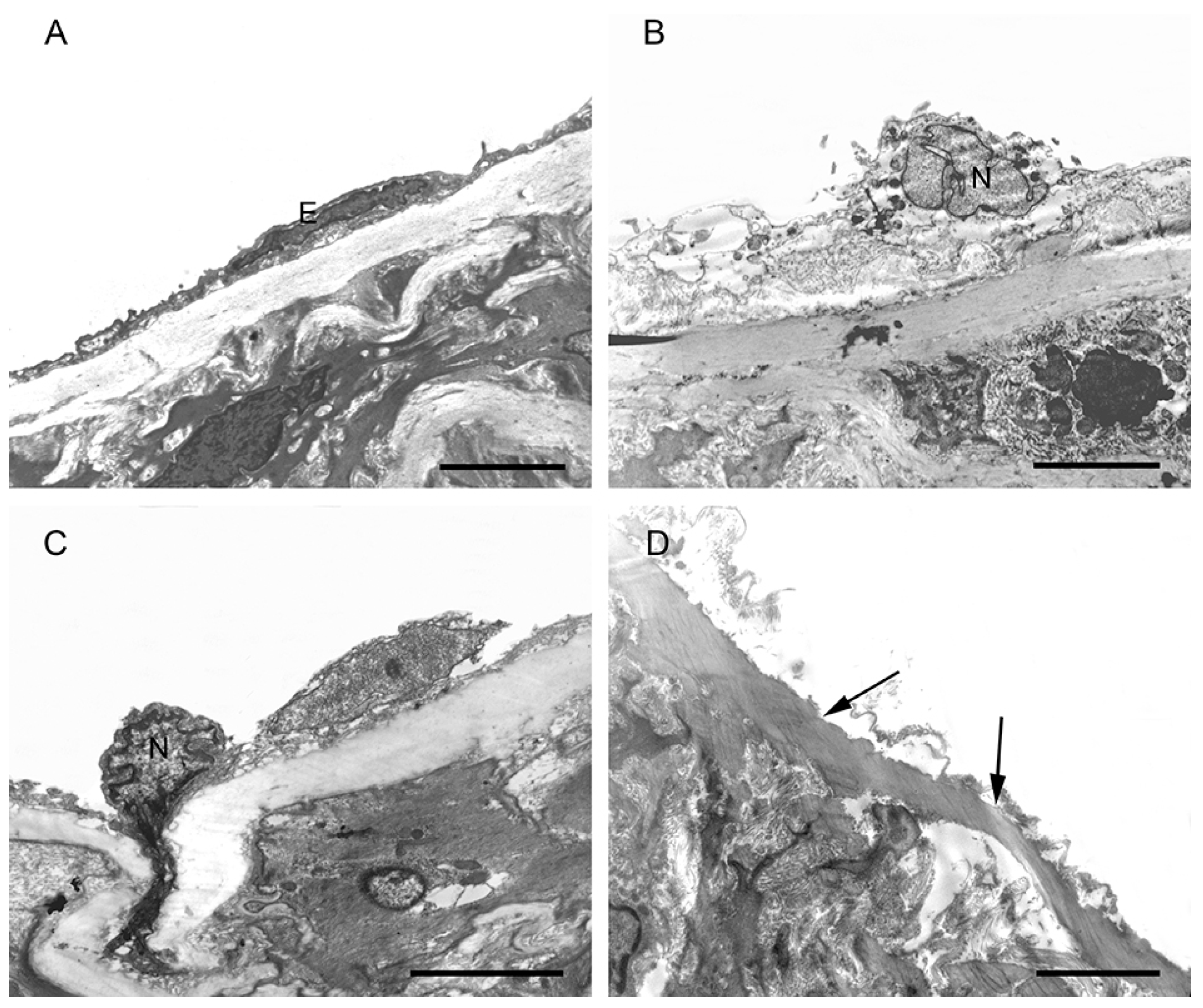
2.4. Morphological Changes in the Aortic Endothelium in CH, SCH, and CON Groups by TEM
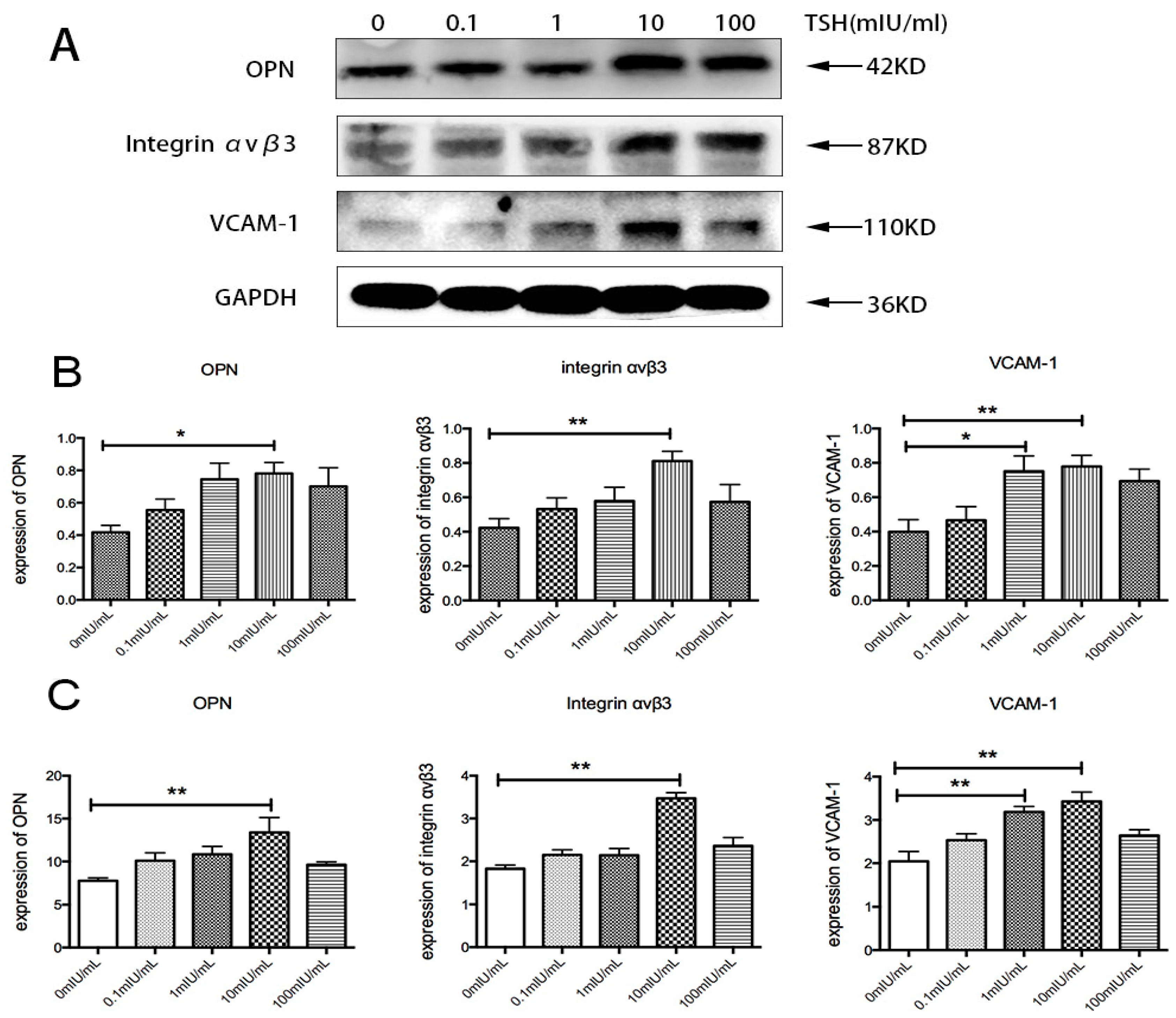
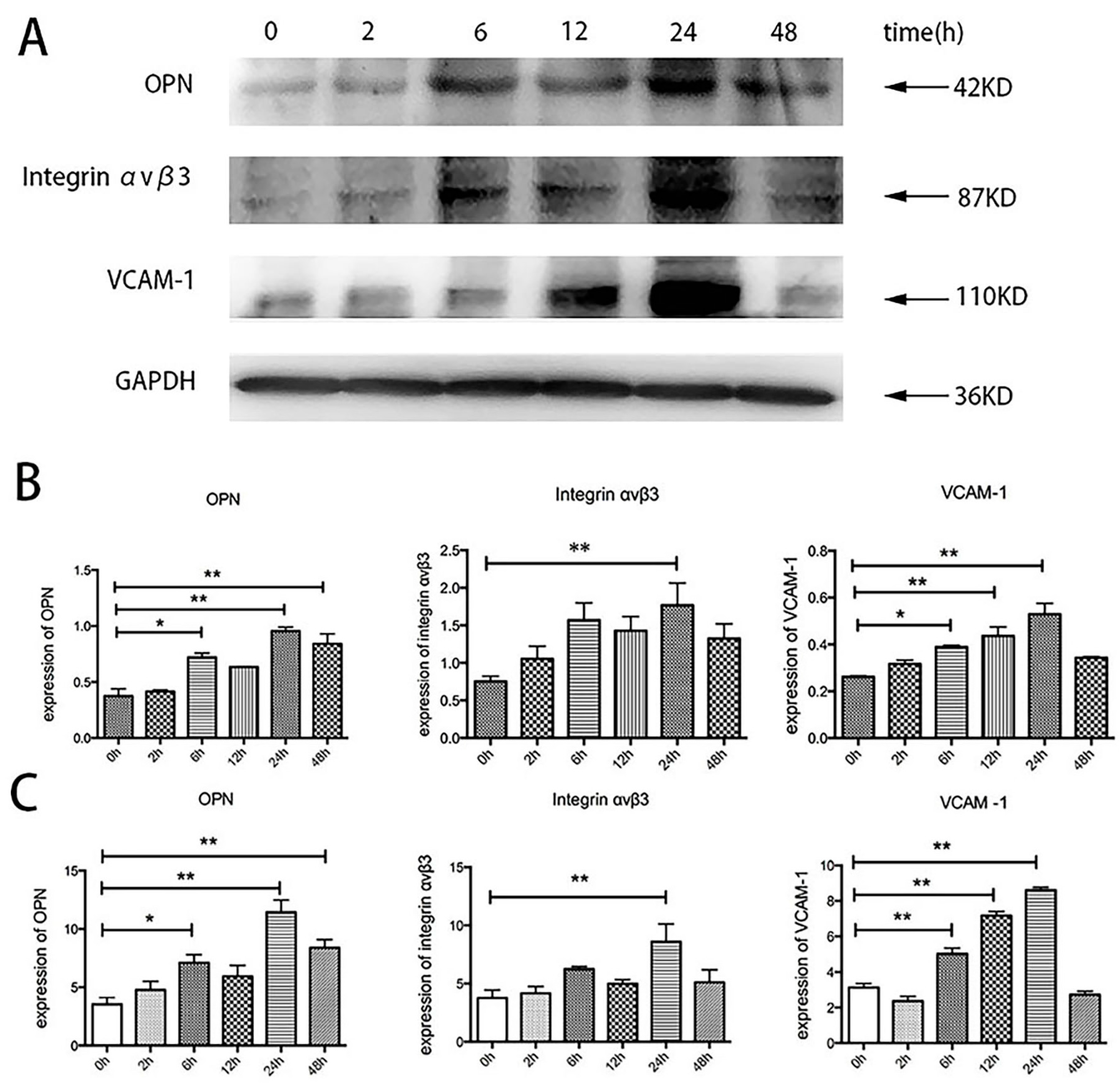
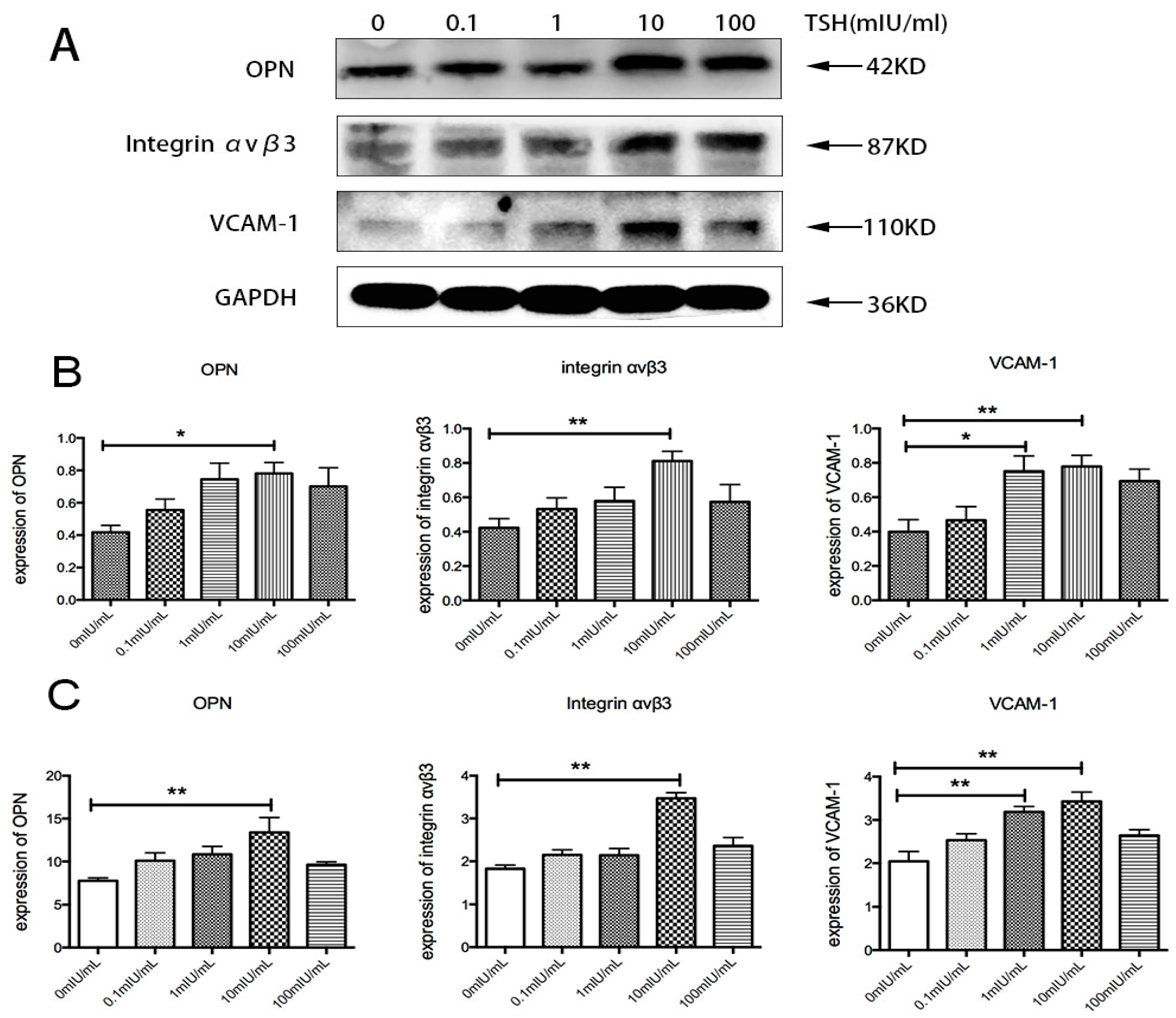
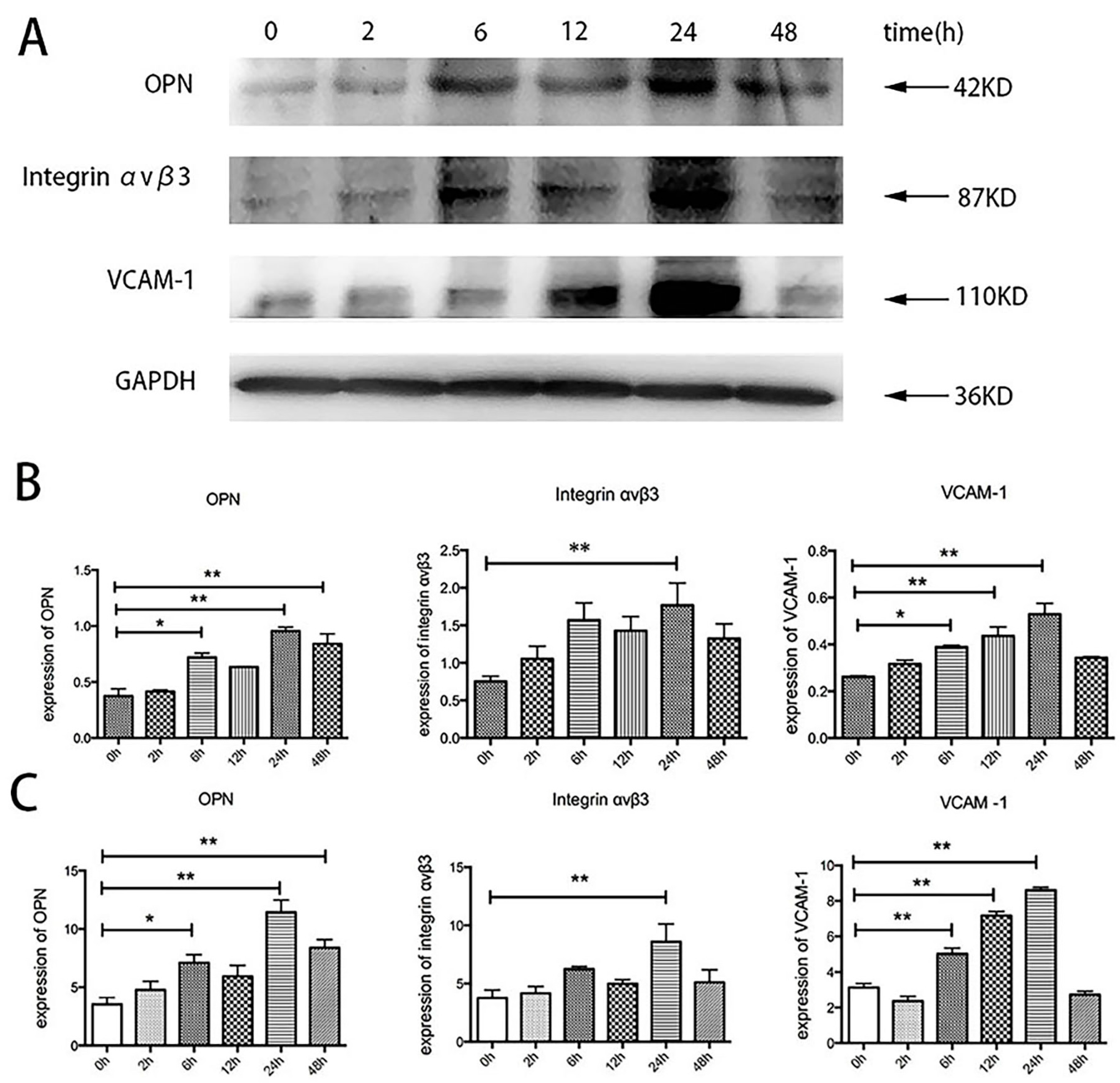
2.5. Effects of TSH on OPN, Integrin αvβ3, and VCAM-1 in Human Umbilical Vein Endothelial Cells (HUVECs)
2.6. Effects of Specific Inhibitors on TSH-Induced OPN, Integrin αvβ3, and VCAM-1 Upregulation in HUVECs
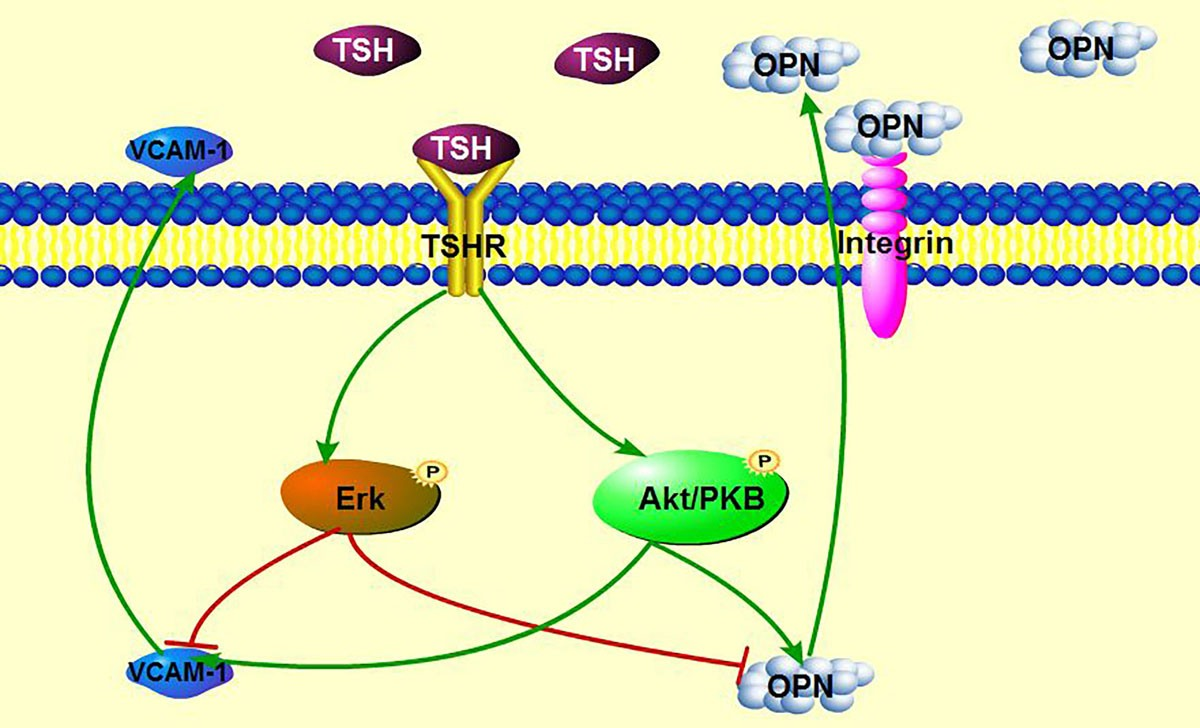
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Hormone Measurements
4.4. Cell Culture
4.5. Western Blot
4.6. Quantitative Real-Time PCR
4.7. Immunohistochemistry (IHC)
4.8. Transmission Electron Microscope (TEM)
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cooper, D.S.; Biondi, B. Subclinical thyroid disease. Lancet 2012, 379, 1142–1154. [Google Scholar] [CrossRef]
- Surks, M.I.; Ortiz, E.; Daniels, G.H.; Sawin, C.T.; Col, N.F.; Cobin, R.H.; Franklyn, J.A.; Hershman, J.M.; Burman, K.D.; Denke, M.A.; et al. Subclinical thyroid disease: Scientific review and guidelines for diagnosis and management. J. Am. Med. Assoc. 2004, 291, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Hak, A.E.; Pols, H.A.; Visser, T.J.; Drexhage, H.A.; Hofman, A.; Witteman, J.C. Subclinical hypothyroidism is an independent risk factor for atherosclerosis and myocardial infarction in elderly women: The Rotterdam Study. Ann. Intern. Med. 2000, 132, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Zhang, W.; Zhang, Y.Z.; Yang, Q.; Chen, S.H. Carotid intima-media thickness in patients with subclinical hypothyroidism: A meta-analysis. Atherosclerosis 2013, 227, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Duntas, L.H. Thyroid disease and lipids. Thyroid Off. J. Am. Thyroid Assoc. 2002, 12, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tan, Y.; Wang, C.; Zhang, X.; Zhao, Y.; Song, X.; Zhang, B.; Guan, Q.; Xu, J.; Zhang, J.; et al. Thyroid-stimulating hormone levels within the reference range are associated with serum lipid profiles independent of thyroid hormones. J. Clin. Endocrinol. Metab. 2012, 97, 2724–2731. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Wang, J.; Jiang, F.; Wang, B.; Chen, Y.; Li, M.; Liu, H.; Li, C.; Xue, H.; Li, N.; et al. The relationship between serum thyrotropin and components of metabolic syndrome. Endocr. J. 2011, 58, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Heinegard, D.; Hultenby, K.; Oldberg, A.; Reinholt, F.; Wendel, M. Macromolecules in bone matrix. Connect. Tissue Res. 1989, 21, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Tousoulis, D.; Siasos, G.; Maniatis, K.; Oikonomou, E.; Kioufis, S.; Zaromitidou, M.; Paraskevopoulos, T.; Michalea, S.; Kollia, C.; Miliou, A.; et al. Serum osteoprotegerin and osteopontin levels are associated with arterial stiffness and the presence and severity of coronary artery disease. Int. J. Cardiol. 2013, 167, 1924–1928. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, R.; Momiyama, Y.; Taniguchi, H.; Takahashi, R.; Kusuhara, M.; Nakamura, H.; Ohsuzu, F. Plasma osteopontin levels are associated with the presence and extent of coronary artery disease. Atherosclerosis 2003, 170, 333–337. [Google Scholar] [CrossRef]
- Dhore, C.R.; Cleutjens, J.P.; Lutgens, E.; Cleutjens, K.B.; Geusens, P.P.; Kitslaar, P.J.; Tordoir, J.H.; Spronk, H.M.; Vermeer, C.; Daemen, M.J. Differential expression of bone matrix regulatory proteins in human atherosclerotic plaques. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1998–2003. [Google Scholar] [CrossRef] [PubMed]
- Hoshiga, M.; Alpers, C.E.; Smith, L.L.; Giachelli, C.M.; Schwartz, S.M. αvβ3 integrin expression in normal and atherosclerotic artery. Circ. Res. 1995, 77, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Donnini, D.; Ambesi-Impiombato, F.S.; Curcio, F. Thyrotropin stimulates production of procoagulant and vasodilative factors in human aortic endothelial cells. Thyroid Off. J. Am. Thyroid Assoc. 2003, 13, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Zhang, L.; Liu, J.; Guo, T.; Gao, C.; Ni, J. Effects of thyroid-stimulating hormone on the function of human umbilical vein endothelial cells. J. Mol. Endocrinol. 2014, 52, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Napoli, R.; Biondi, B.; Guardasole, V.; D’Anna, C.; de Sena, A.; Pirozzi, C.; Terracciano, D.; Mazzarella, C.; Matarazzo, M.; Sacca, L. Enhancement of vascular endothelial function by recombinant human thyrotropin. J. Clin. Endocrinol. Metab. 2008, 93, 1959–1963. [Google Scholar] [CrossRef] [PubMed]
- Napoli, R.; Apuzzi, V.; Bosso, G.; D’Anna, C.; de Sena, A.; Pirozzi, C.; Marano, A.; Lupoli, G.A.; Cudemo, G.; Oliviero, U.; et al. Recombinant human thyrotropin enhances endothelial-mediated vasodilation of conduit arteries. J. Clin. Endocrinol. Metab. 2009, 94, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Dardano, A.; Ghiadoni, L.; Plantinga, Y.; Caraccio, N.; Bemi, A.; Duranti, E.; Taddei, S.; Ferrannini, E.; Salvetti, A.; Monzani, F. Recombinant human thyrotropin reduces endothelium-dependent vasodilation in patients monitored for differentiated thyroid carcinoma. J. Clin. Endocrinol. Metab. 2006, 91, 4175–4178. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R. The Croonian Lecture, 1993. The endothelium: Maestro of the blood circulation. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1994, 343, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Poredos, P. Endothelial dysfunction and cardiovascular disease. Pathophysiol. Haemost. Thromb. 2002, 32, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Ruschitzka, F.T.; Noll, G.; Luscher, T.F. The endothelium in coronary artery disease. Cardiology 1997, 88, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Valentina, V.N.; Marijan, B.; Chedo, D.; Branka, K. Subclinical hypothyroidism and risk to carotid atherosclerosis. Arq. Bras. Endocrinol. Metabol. 2011, 55, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Kim, S.H.; Park, K.S.; Park, S.W.; Cho, Y.W. Regression of the increased common carotid artery-intima media thickness in subclinical hypothyroidism after thyroid hormone replacement. Endocr. J. 2009, 56, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Sara, J.D.; Zhang, M.; Gharib, H.; Lerman, L.O.; Lerman, A. Hypothyroidism is associated with coronary endothelial dysfunction in women. J. Am. Heart Assoc. 2015. [Google Scholar] [CrossRef]
- Del Busto-Mesa, A.; Cabrera-Rego, J.O.; Carrero-Fernandez, L.; Hernandez-Roca, C.V.; Gonzalez-Valdes, J.L.; de la Rosa-Pazos, J.E. Changes in arterial stiffness, carotid intima-media thickness, and epicardial fat after l-thyroxine replacement therapy in hypothyroidism. Endocrinol. Nutr. 2015, 62, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Denhardt, D.T.; Noda, M.; O’Regan, A.W.; Pavlin, D.; Berman, J.S. Osteopontin as a means to cope with environmental insults: Regulation of inflammation, tissue remodeling, and cell survival. J. Clin. Investig. 2001, 107, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ma, X.; Wang, Y.; Li, X.; Qi, Y.; Cui, B.; Li, X.; Ning, G.; Wang, S. The expression and pathophysiological role of osteopontin in Graves’ disease. J. Clin. Endocrinol. Metab. 2011, 96, E1866–E1870. [Google Scholar] [CrossRef] [PubMed]
- Reza, S.; Shaukat, A.; Arain, T.M.; Riaz, Q.S.; Mahmud, M. Expression of osteopontin in patients with thyroid dysfunction. PLoS ONE 2013, 8, e56533. [Google Scholar] [CrossRef] [PubMed]
- Liou, Y.M.; Chang, L.Y.; Liaw, J.M.; Jiang, M.J. Osteopontin gene expression in the aorta and the heart of propylthiouracil-induced hypothyroid mice. J. Biomed. Sci. 2005, 12, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Sajid, M.; Stouffer, G.A. The role of αvβ3 integrins in vascular healing. Thromb. Haemost. 2002, 87, 187–193. [Google Scholar] [PubMed]
- Eliceiri, B.P.; Cheresh, D.A. Adhesion events in angiogenesis. Curr. Opin. Cell Biol. 2001, 13, 563–568. [Google Scholar] [CrossRef]
- Sadeghi, M.M.; Krassilnikova, S.; Zhang, J.; Gharaei, A.A.; Fassaei, H.R.; Esmailzadeh, L.; Kooshkabadi, A.; Edwards, S.; Yalamanchili, P.; Harris, T.D.; et al. Detection of injury-induced vascular remodeling by targeting activated αvβ3 integrin in vivo. Circulation 2004, 110, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Spiecker, M.; Peng, H.-B.; Liao, J.K. Inhibition of endothelial vascular cell adhesion molecule-1 expression by nitric oxide involves the induction and nuclear translocation of IκBα. J. Biol. Chem. 1997, 272, 30969–30974. [Google Scholar] [CrossRef] [PubMed]
- Masseau, I.; Bowles, D.K. Carotid endothelial VCAM-1 is an early marker of carotid atherosclerosis and predicts coronary artery disease in swine. J. Biomed. Sci. Eng. 2015, 8, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Douglas, R.S.; Afifiyan, N.F.; Hwang, C.J.; Chong, K.; Haider, U.; Richards, P.; Gianoukakis, A.G.; Smith, T.J. Increased generation of fibrocytes in thyroid-associated ophthalmopathy. J. Clin. Endocrinol. Metab. 2010, 95, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, E.F.; Papageorgiou, K.I.; Fernando, R.; Raychaudhuri, N.; Cockerham, K.P.; Charara, L.K.; Goncalves, A.C.; Zhao, S.X.; Ginter, A.; Lu, Y.; et al. Increased expression of TSH receptor by fibrocytes in thyroid-associated ophthalmopathy leads to chemokine production. J. Clin. Endocrinol. Metab. 2012, 97, E740–E746. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ren, J.; Jing, Q.; Lu, S.; Zhang, Y.; Liu, Y.; Yu, C.; Gao, P.; Zong, C.; Li, X.; et al. TSH/TSHR signaling suppresses fatty acid synthase (FASN) expression in adipocytes. J. Cell. Physiol. 2015, 230, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Tsygankova, O.M.; Saavedra, A.; Rebhun, J.F.; Quilliam, L.A.; Meinkoth, J.L. Coordinated regulation of Rap1 and thyroid differentiation by cyclic AMP and protein kinase A. Mol. Cell. Biol. 2001, 21, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Tsygankova, O.M.; Feshchenko, E.; Klein, P.S.; Meinkoth, J.L. Thyroid-stimulating hormone/cAMP and glycogen synthase kinase 3β elicit opposing effects on Rap1GAP stability. J. Biol. Chem. 2004, 279, 5501–5507. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.M.; Song, J.H.; Kim, D.W.; Kim, H.; Chung, H.K.; Hwang, J.H.; Kim, J.M.; Hwang, E.S.; Chung, J.; Han, J.H.; et al. Regulation of the phosphatidylinositol 3-kinase, Akt/protein kinase B, FRAP/mammalian target of rapamycin, and ribosomal S6 kinase 1 signaling pathways by thyroid-stimulating hormone (TSH) and stimulating type TSH receptor antibodies in the thyroid gland. J. Biol. Chem. 2003, 278, 21960–21971. [Google Scholar] [CrossRef] [PubMed]
- Balzan, S.; del Carratore, R.; Nicolini, G.; Beffy, P.; Lubrano, V.; Forini, F.; Iervasi, G. Proangiogenic effect of TSH in human microvascular endothelial cells through its membrane receptor. J. Clin. Endocrinol. Metab. 2012, 97, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Puri, K.D.; Doggett, T.A.; Huang, C.Y.; Douangpanya, J.; Hayflick, J.S.; Turner, M.; Penninger, J.; Diacovo, T.G. The role of endothelial PI3Kγ activity in neutrophil trafficking. Blood 2005, 106, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.A.; Huang, J.H.; Liao, W.S. Phosphatidylinositol 3-kinase as a mediator of TNF-induced NFκB activation. J. Immunol. 2000, 164, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Ozes, O.N.; Mayo, L.D.; Gustin, J.A.; Pfeffer, S.R.; Pfeffer, L.M.; Donner, D.B. NF-κB activation by tumour necrosis factor requires the Akt serine-threonine kinase. Nature 1999, 401, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, Y.; Yu, X.; Wang, X.; Bao, S.; Li, J.; Fan, C.; Shan, Z.; Teng, W. Maternal subclinical hypothyroidism impairs neurodevelopment in rat offspring by inhibiting the CREB signaling pathway. Mol. Neurobiol. 2015, 52, 432–441. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON | SCH | CH | ||||
|---|---|---|---|---|---|---|
| TSH (mIU/L) | TT4 (μg/dL) | TSH (mIU/L) | TT4 (μg/dL) | TSH (mIU/L) | TT4 (μg/dL) | |
| 4 Weeks after surgery | 0.19 ± 0.10 | 4.24 ± 0.82 | 21.65 ± 8.54 * | 1.52 ± 0.93 * | 21.8 ± 3.65 * | 1.34 ± 0.35 * |
| 4 Weeks after l-T4 injection | 0.36 ± 0.19 | 4.01 ± 0.97 | 4.01 ± 0.97 | 3.16 ± 1.09 | 13.0 ± 6.09 * | 1.61 ± 0.55 * |
| 14 Weeks after l-T4 injection | 0.37 ± 0.26 | 3.51 ± 0.97 | 6.79 ± 3.70 * | 3.86 ± 0.78 | 16.14 ± 4.58 * | 1.77 ± 0.73 * |
| Gene | Primer (5′ to 3′) |
|---|---|
| VCAM1 | Forward: GCGGAGACAGGAGACACAGTACTAA |
| Reverse: GAGCACGAGAAGCTCAGGAGAA | |
| SPP1 | Forward: ACAGCCACAAGCAGTCCAGATTA |
| Reverse: TCCTGACTATCAATCACATCGGAAT | |
| ITGB3 | Forward: GAGGTCATCCCTGGCCTCAA |
| Reverse: CTGGCAGGCACAGTCACAATC | |
| GAPDH | Forward: CAATGACCCCTTCATTGACC |
| Reverse: GACAAGCTTCCCGTTCTCAG |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.; Jiang, F.; Lai, Y.; Wang, H.; Liu, A.; Wang, C.; Zhang, Y.; Teng, W.; Shan, Z. Effect of Thyrotropin on Osteopontin, Integrin αvβ3, and VCAM-1 in the Endothelium via Activation of Akt. Int. J. Mol. Sci. 2016, 17, 1484. https://doi.org/10.3390/ijms17091484
Yan Y, Jiang F, Lai Y, Wang H, Liu A, Wang C, Zhang Y, Teng W, Shan Z. Effect of Thyrotropin on Osteopontin, Integrin αvβ3, and VCAM-1 in the Endothelium via Activation of Akt. International Journal of Molecular Sciences. 2016; 17(9):1484. https://doi.org/10.3390/ijms17091484
Chicago/Turabian StyleYan, Yumeng, Fengwei Jiang, Yaxin Lai, Haoyu Wang, Aihua Liu, Chuyuan Wang, Yuanyuan Zhang, Weiping Teng, and Zhongyan Shan. 2016. "Effect of Thyrotropin on Osteopontin, Integrin αvβ3, and VCAM-1 in the Endothelium via Activation of Akt" International Journal of Molecular Sciences 17, no. 9: 1484. https://doi.org/10.3390/ijms17091484
APA StyleYan, Y., Jiang, F., Lai, Y., Wang, H., Liu, A., Wang, C., Zhang, Y., Teng, W., & Shan, Z. (2016). Effect of Thyrotropin on Osteopontin, Integrin αvβ3, and VCAM-1 in the Endothelium via Activation of Akt. International Journal of Molecular Sciences, 17(9), 1484. https://doi.org/10.3390/ijms17091484