
Cannibalism Affects Core Metabolic Processes in Helicoverpa armigera Larvae—A 2D NMR Metabolomics Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
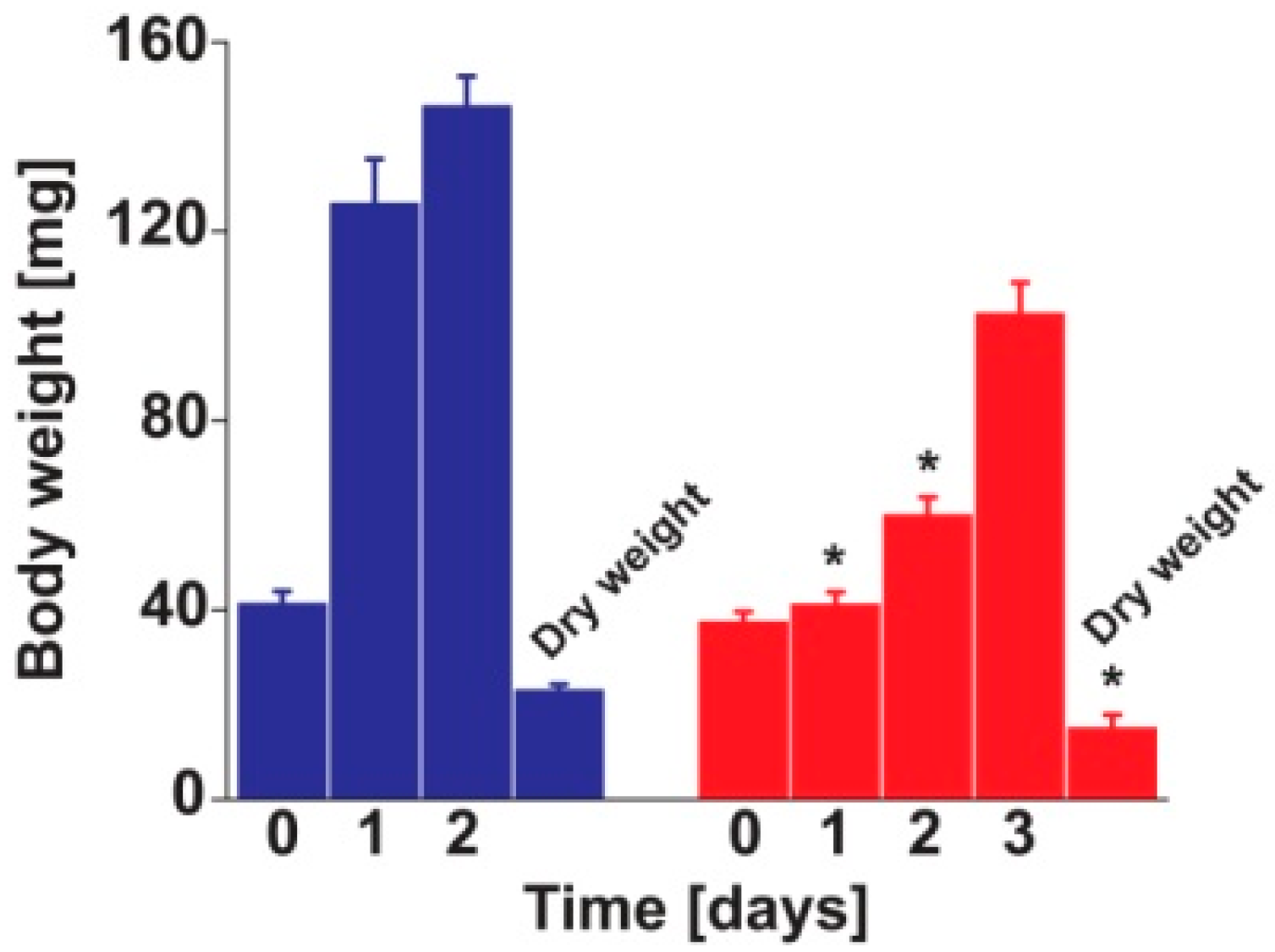
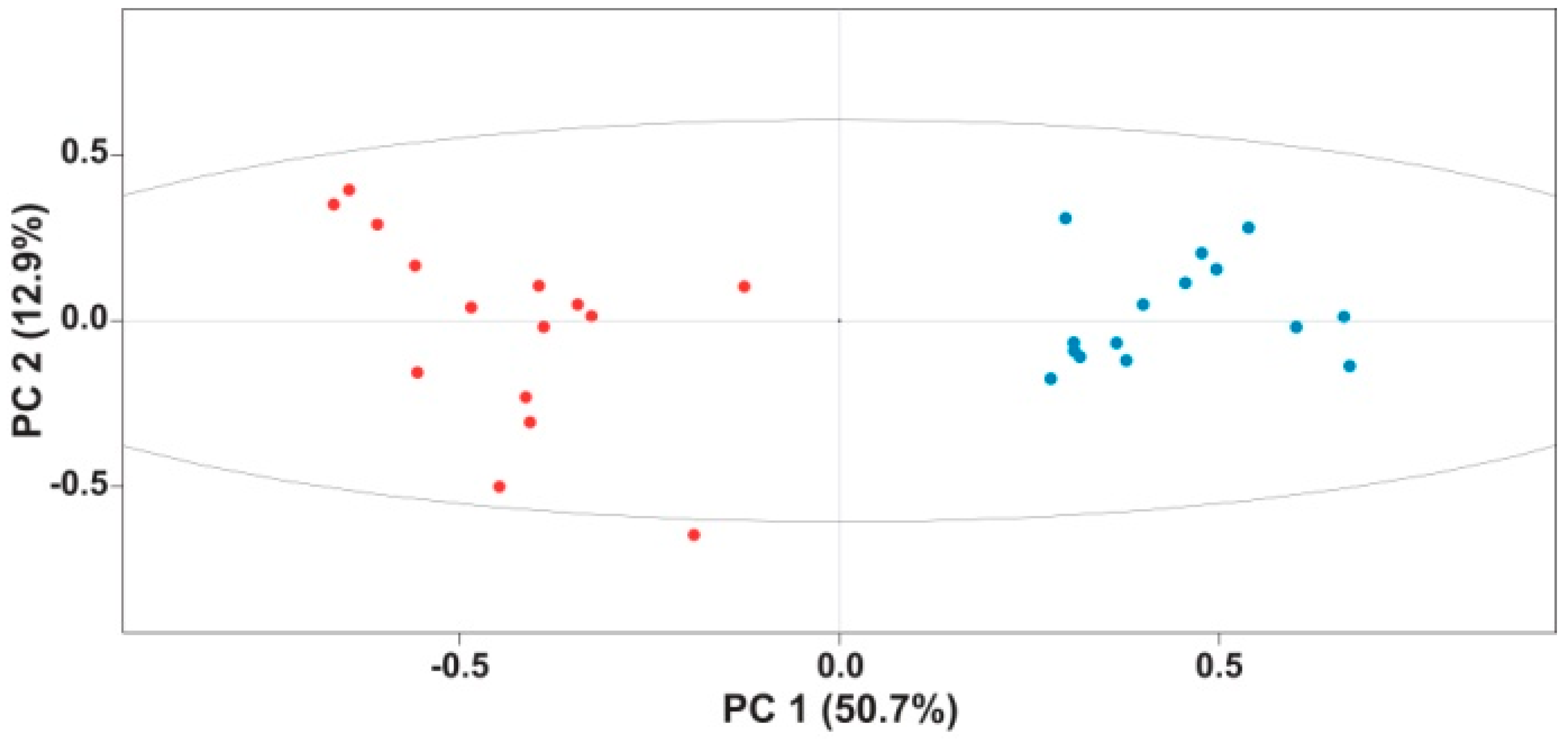
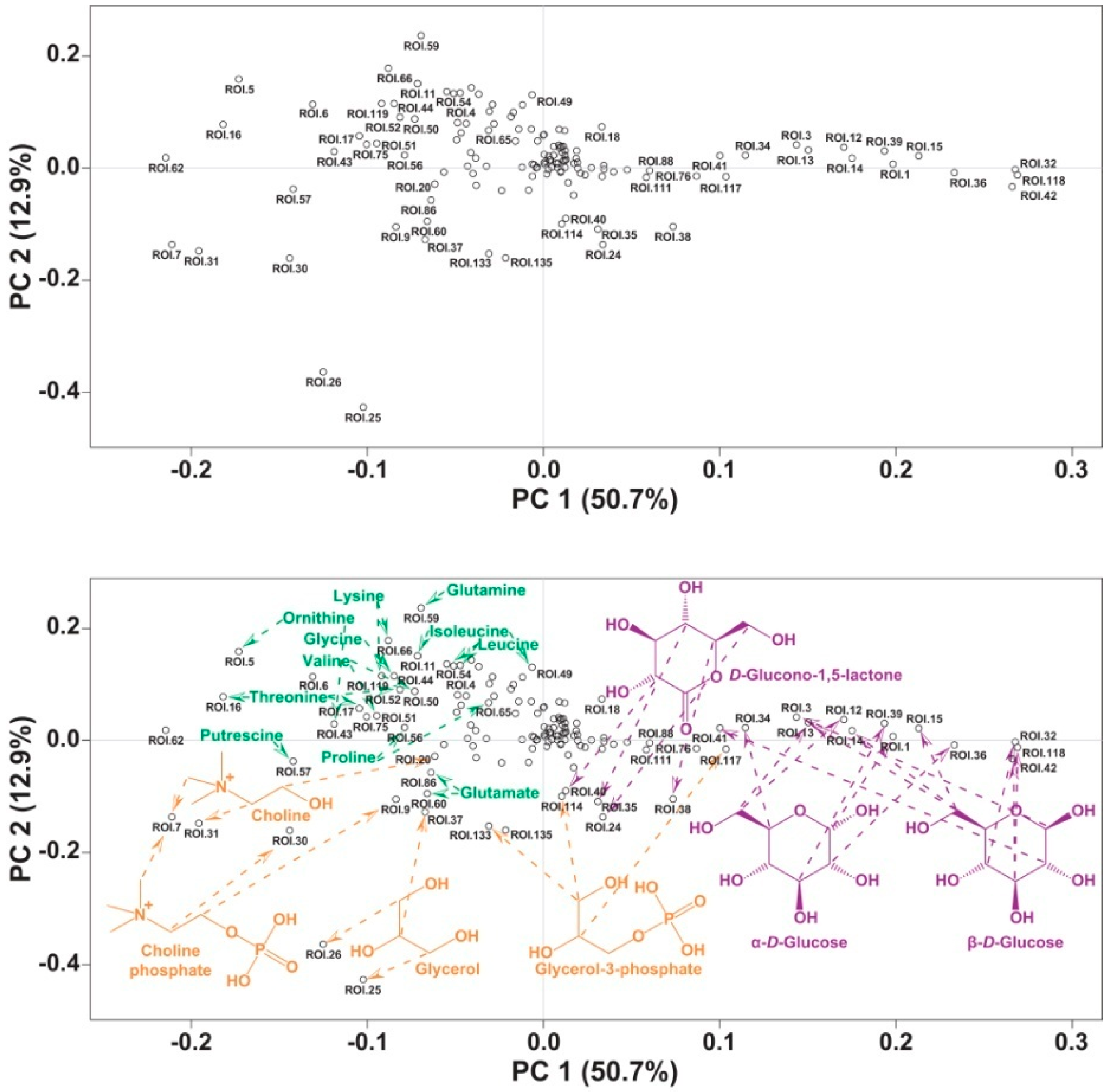
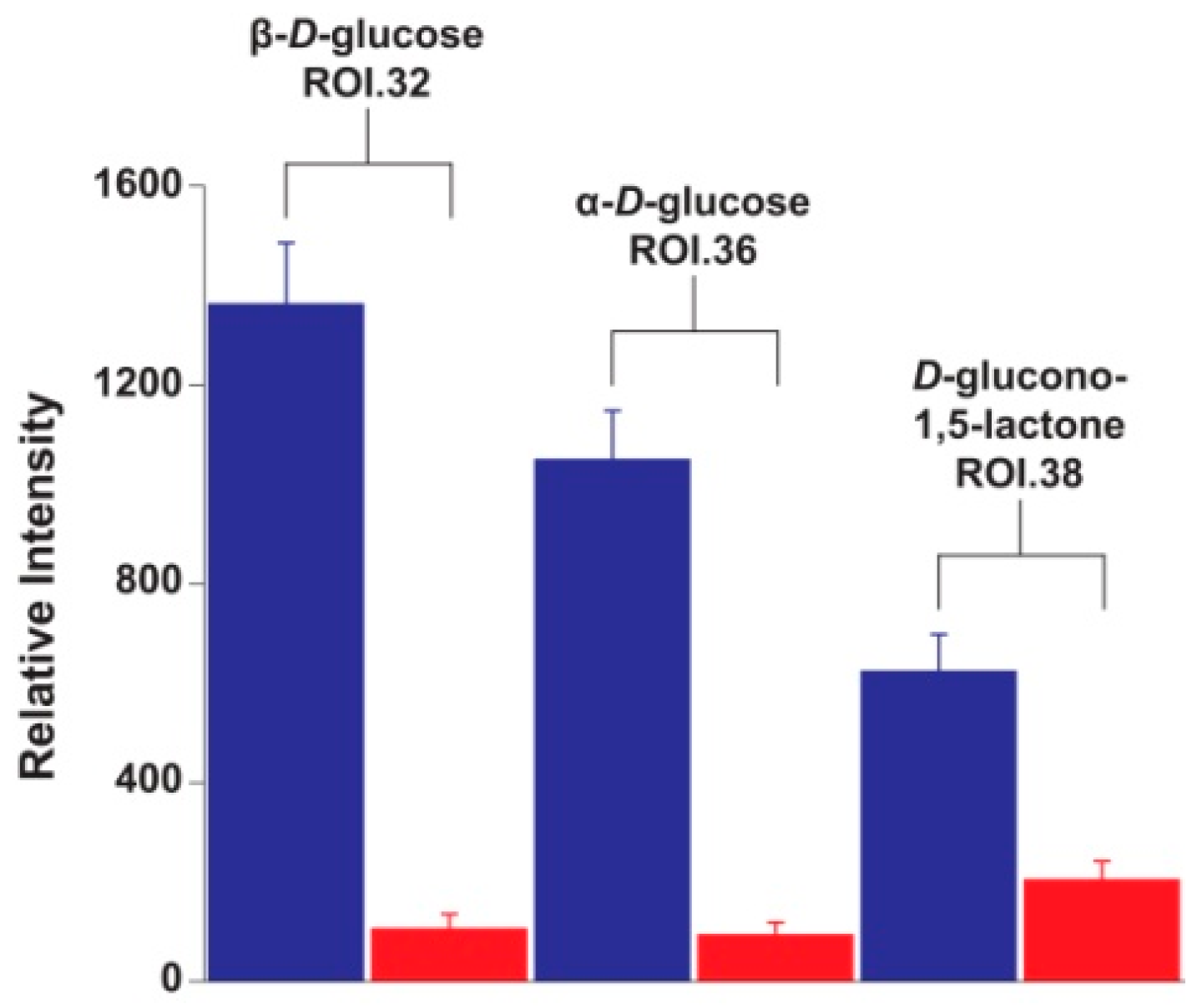
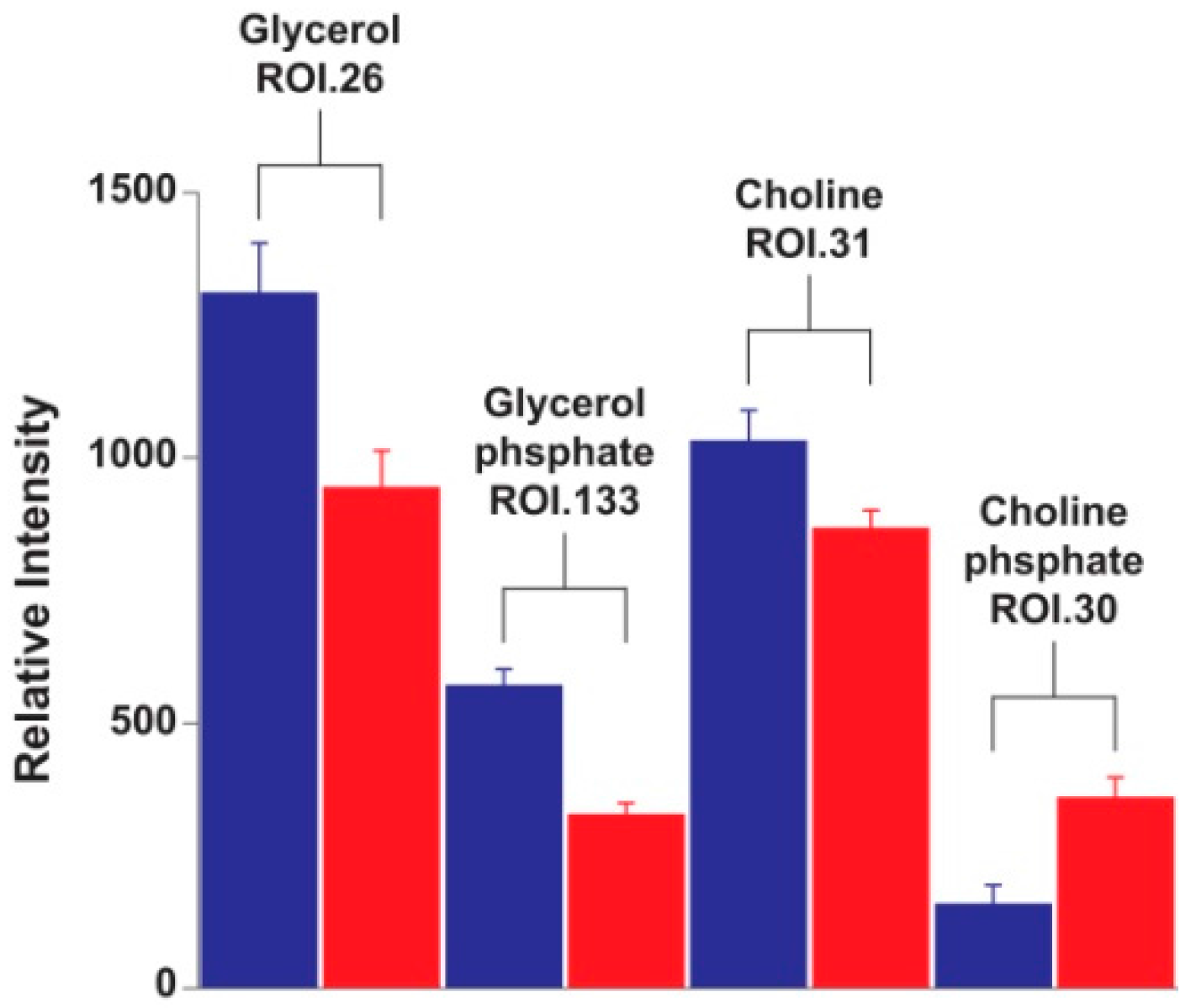
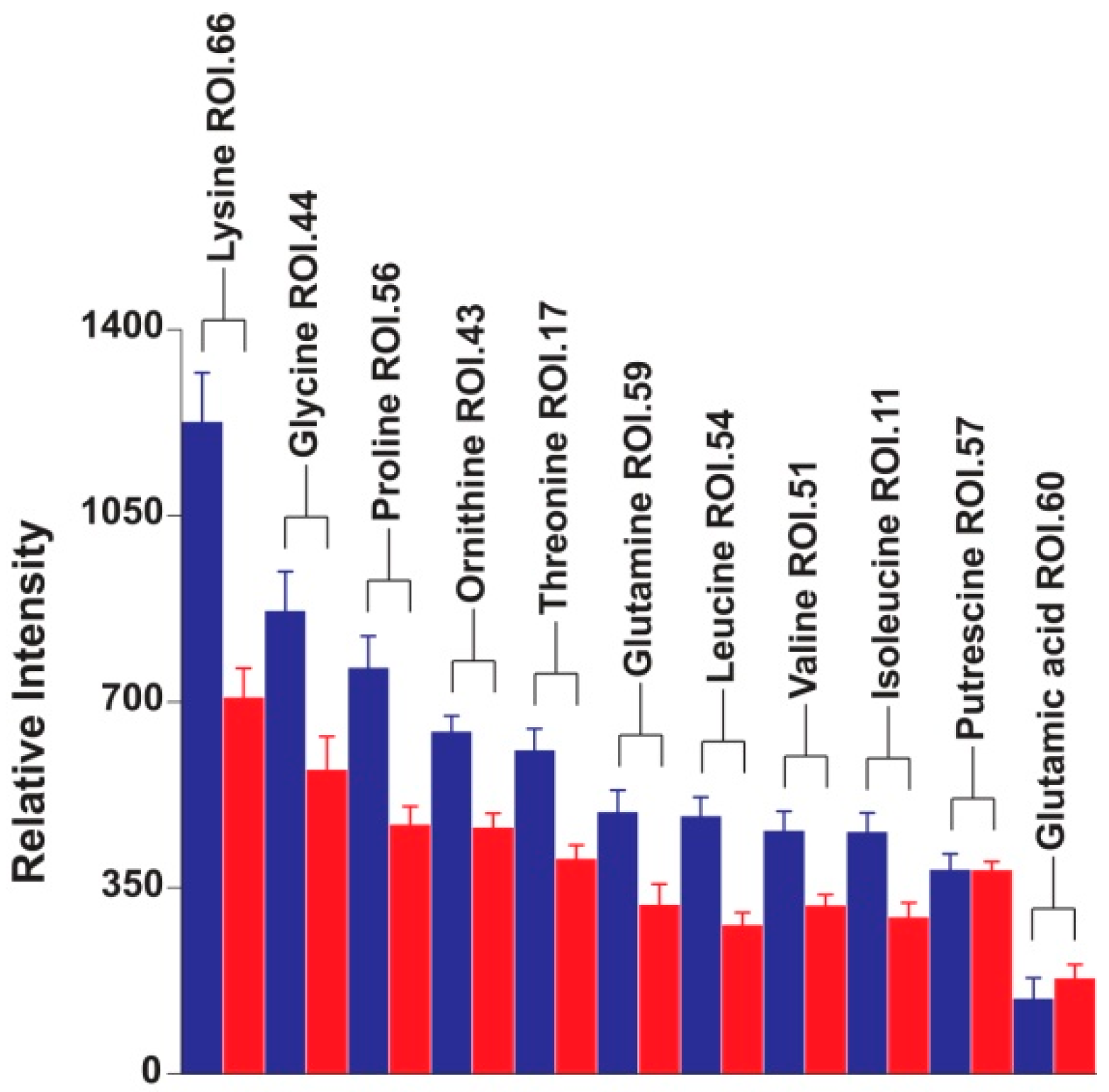
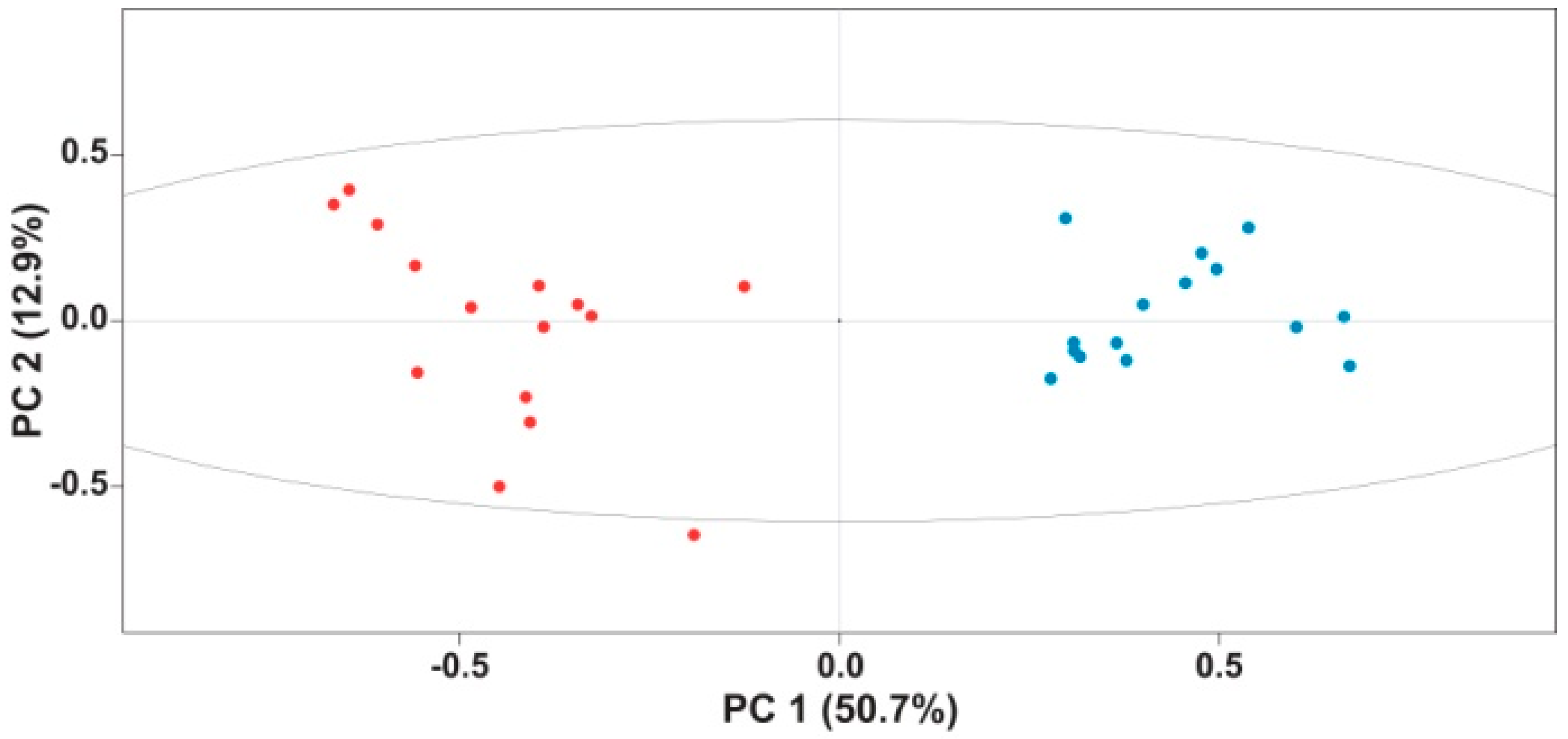
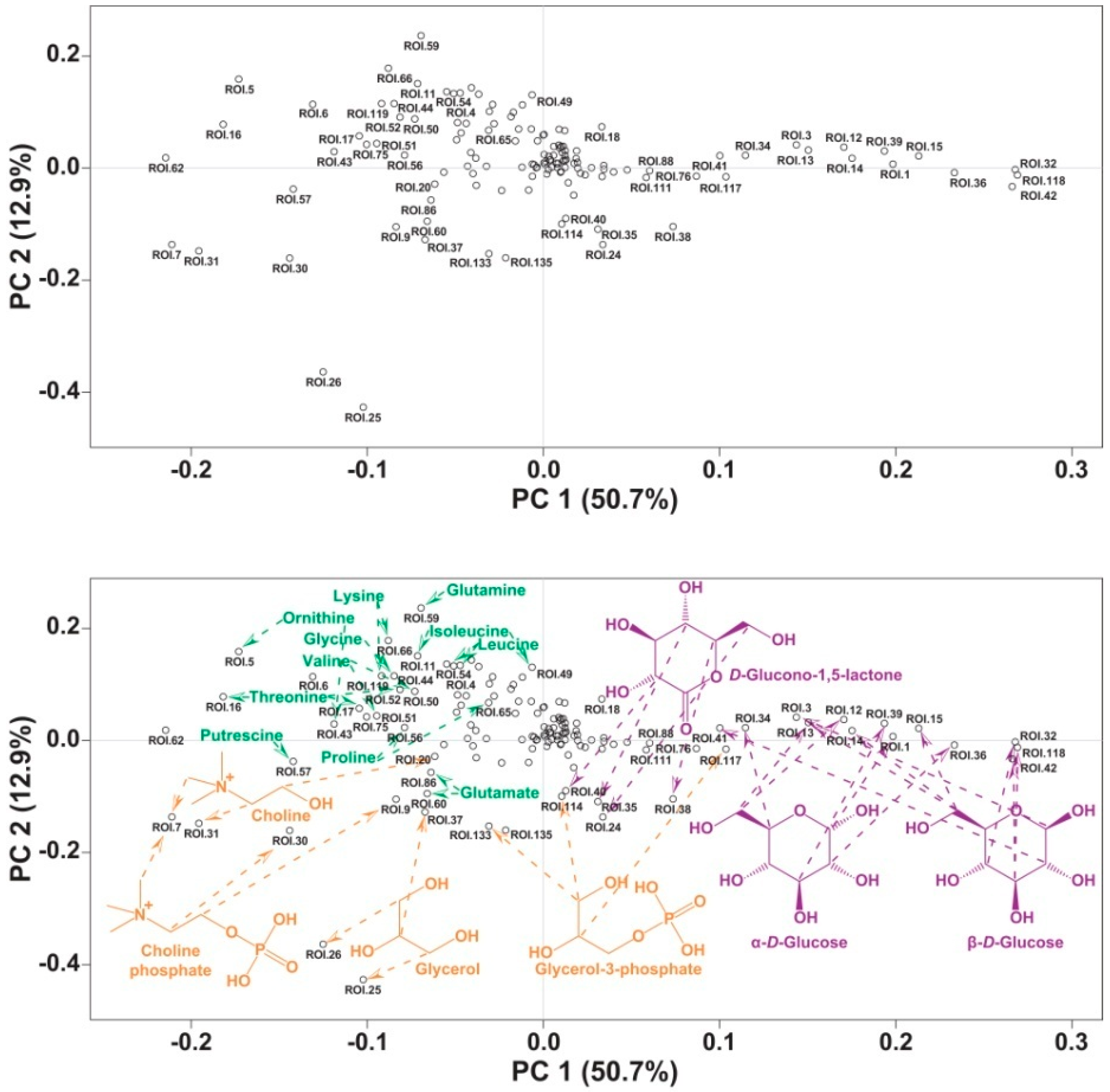
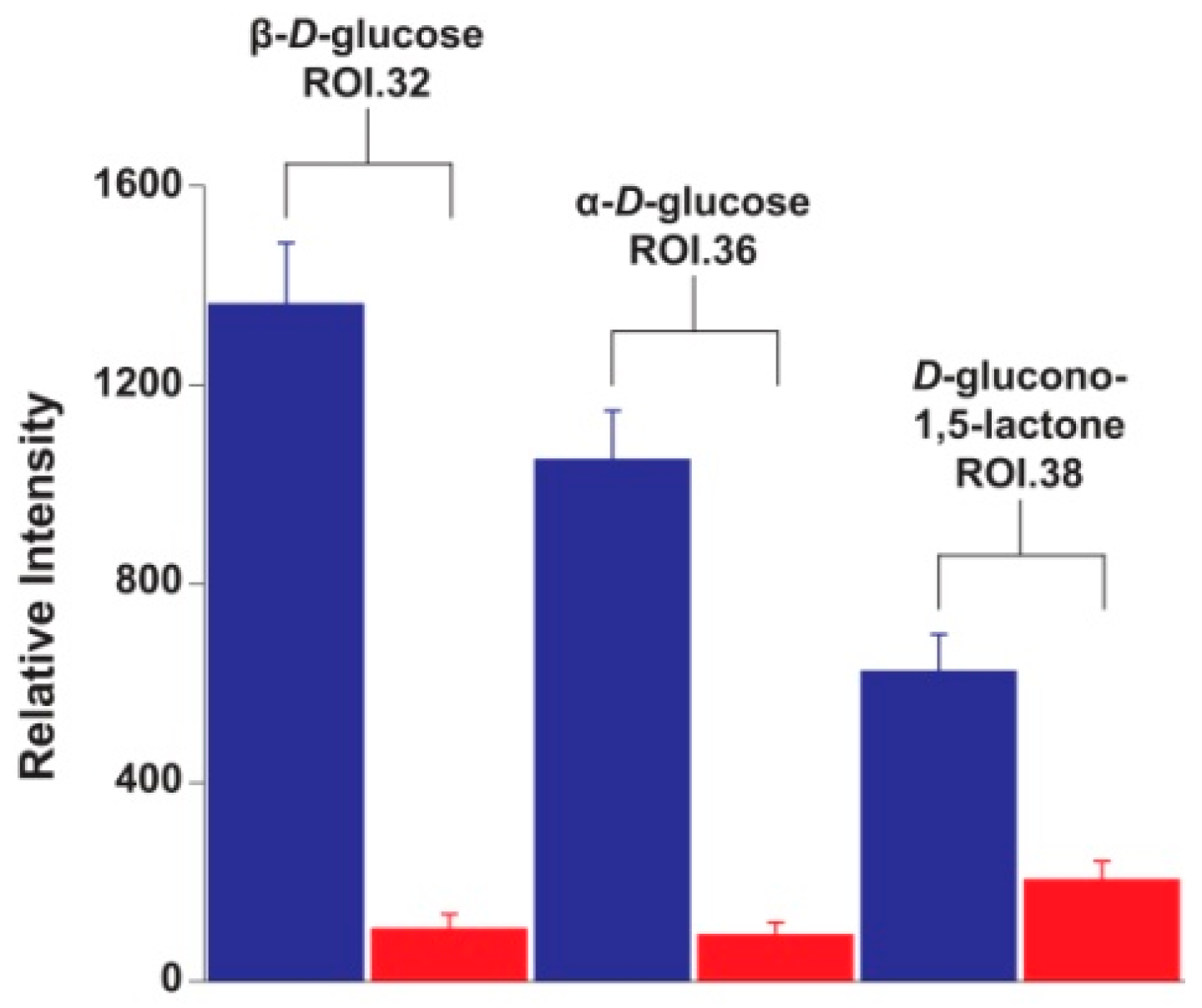
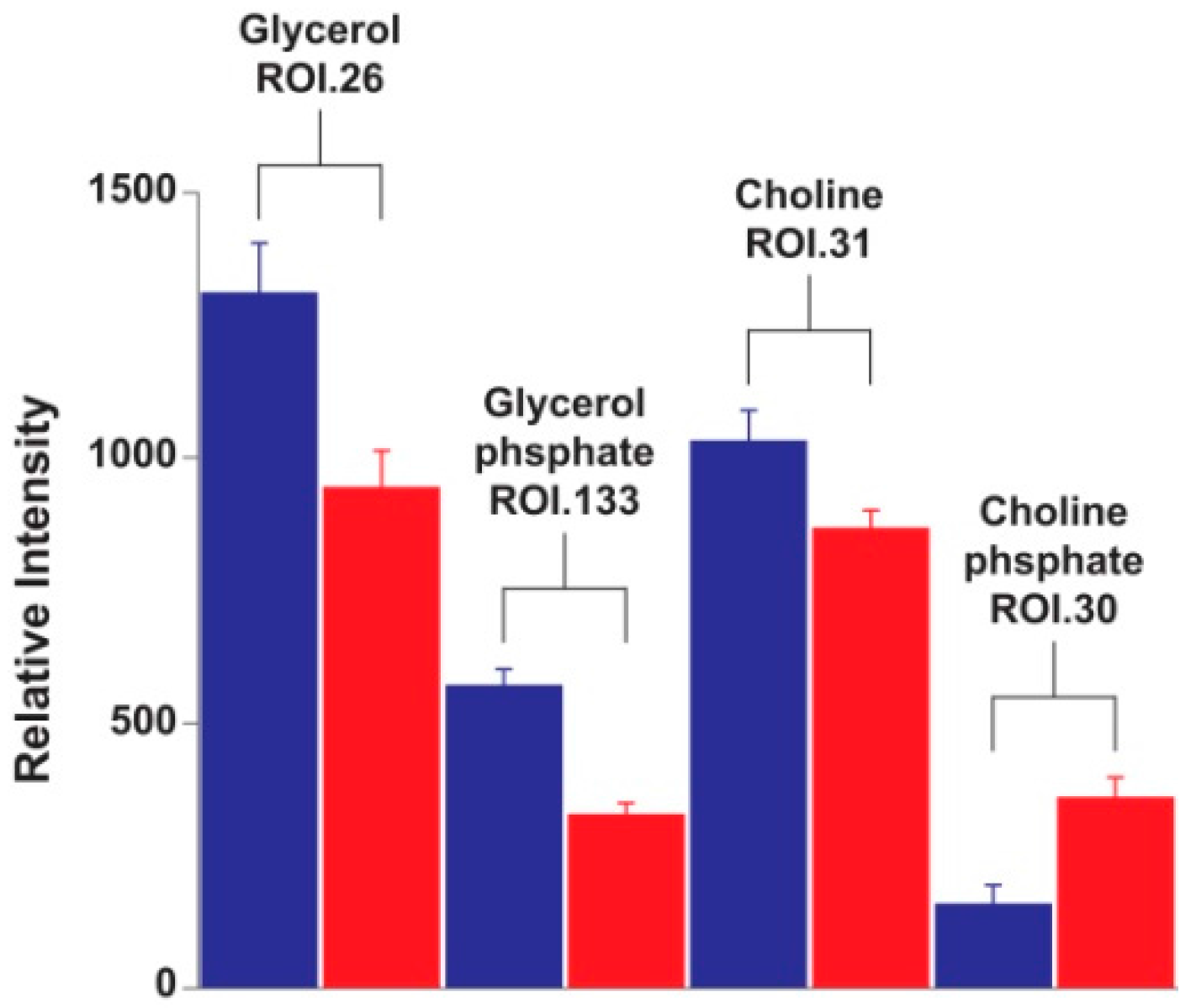
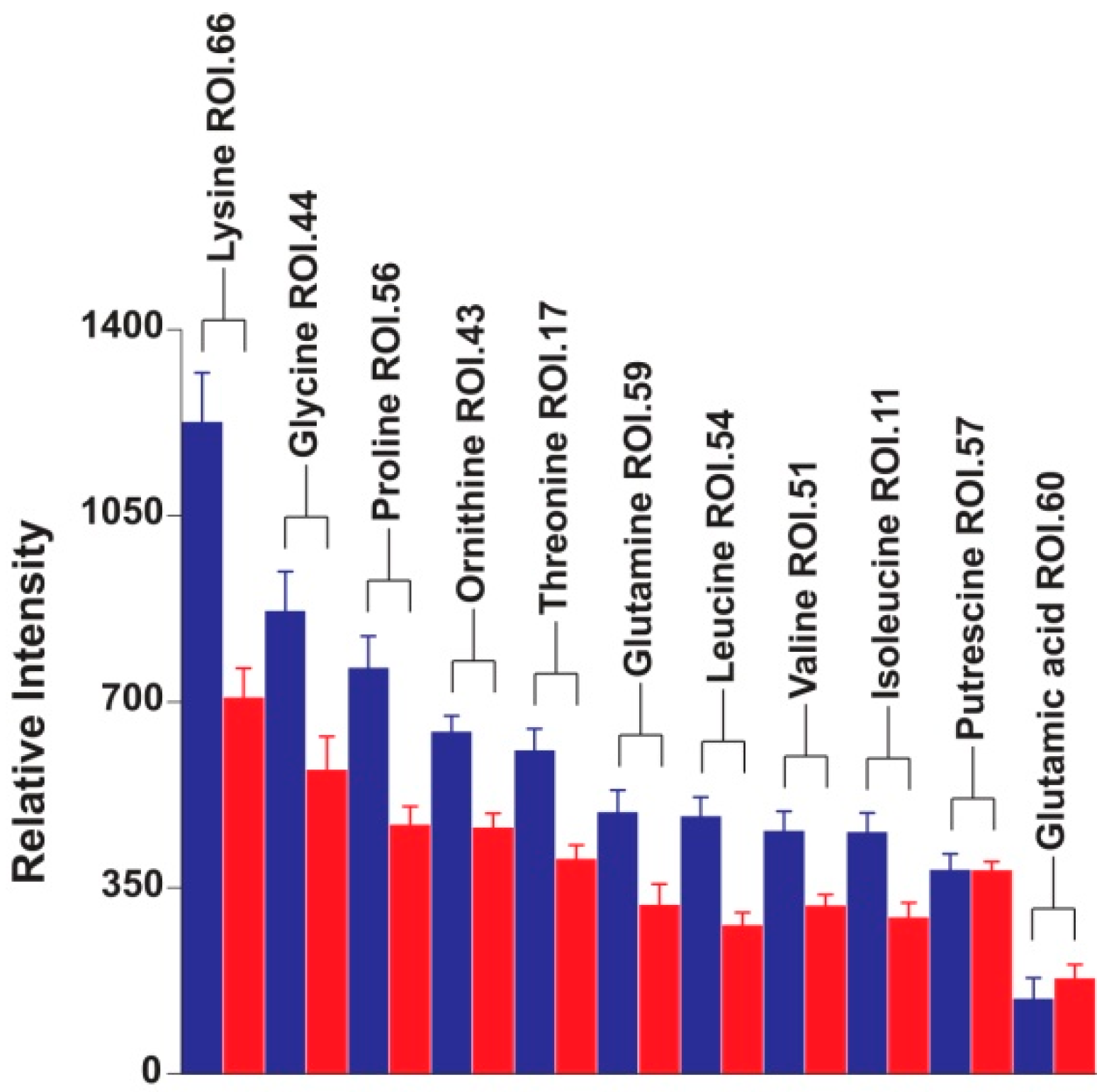
2. Results and Discussion
3. Materials and Methods
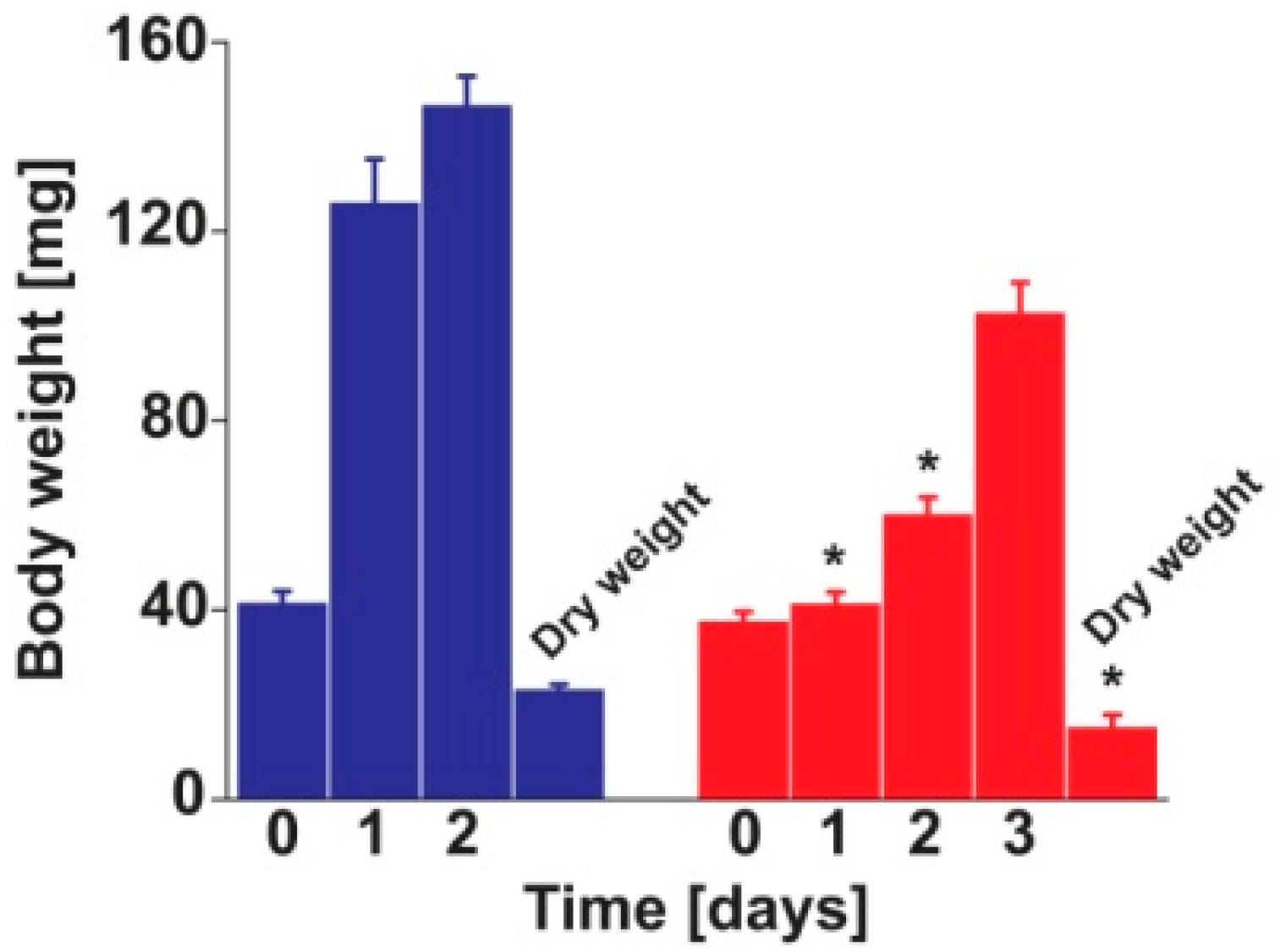
3.1. Insect Bioassay
3.2. Sample Preparation for NMR
3.3. NMR Spectra Acquisition and Processing Parameters
3.4. Multivariate Statistical Analyses of 1H and 1H-13C HSQC NMR Spectra
3.5. Candidate Compounds
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Polis, G. The evolution and dynamics of intraspecific predation. Annu. Rev. Ecol. Syst. 1981, 12, 225–251. [Google Scholar] [CrossRef]
- Pfennig, D.; Ho, S.; Hoffman, E. Pathogen transmission as a selective force against cannibalism. Anim. Behav. 1998, 55, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Gould, F.; Holtzman, G.; Rabb, R.; Smith, M. Genetic variation in predatory and cannibalistic tendencies of Heliothis virescens strains. Ann. Entomol. Soc. Am. 1980, 73, 243–250. [Google Scholar] [CrossRef]
- Joyner, K.; Gould, F. Developmental consequences of cannibalism in Heliothis zea (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1985, 78, 24–28. [Google Scholar] [CrossRef]
- Richardson, M.; Mitchell, R.; Reagel, P.; Hanks, L. Causes and consequences of cannibalism in noncarnivorous insects. Annu. Rev. Entomol. 2009, 55, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Kakimoto, T.; Fujisaki, K.; Miyatake, T. Egg laying preference, larval dispersion, and cannibalism in Helicoverpa armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2003, 96, 793–798. [Google Scholar] [CrossRef]
- Fitt, G. The ecology of Heliothis species in relation to agroecosystems. Annu. Rev. Entomol. 1989, 34, 17–52. [Google Scholar] [CrossRef]
- Parde, V.; Sharma, H.; Kachole, M. Inhibition of Helicoverpa armigera gut pro-proteinase activation in response to synthetic protease inhibitors. Entomol. Exp. Appl. 2012, 142, 104–113. [Google Scholar] [CrossRef]
- Bodenhausen, G.; Ruben, D. Natural abundance nitrogen-15 NMR by enhanced heteronuclear spectroscopy. Chem. Phys. Lett. 1980, 69, 185–189. [Google Scholar] [CrossRef]
- Le Guennec, A.; Giraudeau, P.; Caldarelli, S. Evaluation of fast 2D NMR for metabolomics. Anal. Chem. 2014, 86, 5946–5954. [Google Scholar] [CrossRef] [PubMed]
- Fardus-Reid, F.; Warrena, J.; Le Gresley, A. Validating heteronuclear 2D quantitative NMR. Anal. Methods 2016, 8, 2013–2019. [Google Scholar] [CrossRef]
- Tang, Q.; Hu, Y.; Kang, L.; Wang, C.Z. Characterization of glucose-induced glucose oxidase gene and protein expression in Helicoverpa armigera larvae. Arch. Insect Biochem. 2012, 79, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Wolf, G. The discovery of a vitamin role for carnitine: The first 50 years. J. Nutr. 2006, 136, 2131–2134. [Google Scholar] [PubMed]
- Turunen, S. Metabolic pathways in the midgut epithelium of Pieris brassicae during carbohydrate and lipid assimilation. Insect Biochem. Mol. Biol. 1992, 23, 681–689. [Google Scholar] [CrossRef]
- Zhou, L.; Li, H.; Hao, F.; Li, N.; Liu, X.; Wang, G.; Wang, G.; Tang, H. Developmental changes for the hemolymph metabolome of silkworm (Bombyx mori L.). J. Proteome Res. 2015, 14, 2331–2347. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.B.; Wang, M.Q.; Zhang, G. Effects of putrescine on diapause induction and intensity, and post-diapause development of Helicoverpa armigera. Entomol. Exp. Appl. 2010, 136, 199–205. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, Y.X.; Xu, W.H. Proteomic and metabolomic profiles of larval hemolymph associated with diapause in the cotton bollworm, Helicoverpa armigera. BMC Genom. 2013, 14, 751. [Google Scholar] [CrossRef] [PubMed]
- Osawa, N. Sibling cannibalism in the ladybird beetle Harmonia axyridis—Fitness consequences for mother and offspring. Res. Popul. Ecol. 1992, 34, 45–55. [Google Scholar] [CrossRef]
- Belles-Isles, J.C.; Fitzgerald, G.J. A fitness advantage of cannibalism in female sticklebacks (Gasterosteus aculeatus L.). Ethol. Ecol. Evol. 1993, 5, 187–191. [Google Scholar] [CrossRef]
- Church, S.C.; Sherratt, T.N. The selective advantages of cannibalism in a Neotropical mosquito. Behav. Ecol. Sociobiol. 1996, 39, 117–123. [Google Scholar] [CrossRef]
- Mayntz, D.; Toft, S. Nutritional value of cannibalism and the role of starvation and nutrient imbalance for cannibalistic tendencies in a generalist predator. J. Anim. Ecol. 2006, 75, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.W.; Williams, T.; Escribano, A.; Caballero, P.; Cave, R.D.; Goulson, D. Fitness consequences of cannibalism in the fall armyworm, Spodoptera frugiperda. Behav. Ecol. 1999, 10, 298–303. [Google Scholar] [CrossRef]
- Santana, A.F.K.; Zago, R.C.; Zucoloto, F.S. Effects of sex, host-plant deprivation and presence of conspecific immatures on the cannibalistic behavior of wild Ascia monuste orseis (Godart) (Lepidoptera: Pieridae). Rev. Bras. Entomol. 2011, 55, 95–101. [Google Scholar] [CrossRef]
- Simpson, S.; Clissold, F.; Lihoreau, M.; Ponton, F.; Wilder, S.; Raubenheimer, D. Recent advances in the integrative nutrition of arthropods. Annu. Rev. Entomol. 2015, 60, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Ma, W.H.; Li, G.Q. Age- and nutrition-related cannibalism in larvae of the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). Acta Entomol. Sin. 2015, 58, 175–180. [Google Scholar]
- Tang, T.; Zhao, C.; Xu, L.; Qiu, L. Factors affecting larval cannibalism in the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Orient. Insects 2016, 50, 23–33. [Google Scholar] [CrossRef]
- Krȩżel, A.; Bal, W. A formula for correcting pKa values determined in D2O and H2O. J. Inorg. Biochem. 2004, 98, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Njock, G.; Pegnyemb, D.; Bartholomeusz, T.; Christen, P.; Vitorge, B.; Nuzillard, J.; Shivapurkar, R.; Foroozandeh, M.; Jeannerat, D. Spectral aliasing: A super zoom for 2D-NMR spectra. Principles and applications. Chimia 2010, 64, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Jeannerat, D. Rapid multidimensional NMR: High resolution by spectral aliasing. In Encyclopedia of Magnetic Resonance; John Wiley & Sons, Ltd.: Indianapolis, IN, USA, 2011. [Google Scholar] [CrossRef]
- Wang, T.; Shao, K.; Chu, Q.; Ren, Y.; Mu, Y.; Qu, L.; He, J.; Jin, C.; Xia, B. Automics: An integrated platform for NMR-based metabonomics spectral processing and data analysis. BMC Bioinform. 2009, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Gaude, E.; Chignola, F.; Spiliotopoulos, D.; Spitaleri, A.; Ghitti, M.; García-Manteiga, J.M.; Mari, S.; Musco, G. Muma, an R package for metabolomics univariate and multivariate statistical analysis. Curr. Metabol. 2013, 1, 180–189. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vergara, F.; Shino, A.; Kikuchi, J. Cannibalism Affects Core Metabolic Processes in Helicoverpa armigera Larvae—A 2D NMR Metabolomics Study. Int. J. Mol. Sci. 2016, 17, 1470. https://doi.org/10.3390/ijms17091470
Vergara F, Shino A, Kikuchi J. Cannibalism Affects Core Metabolic Processes in Helicoverpa armigera Larvae—A 2D NMR Metabolomics Study. International Journal of Molecular Sciences. 2016; 17(9):1470. https://doi.org/10.3390/ijms17091470
Chicago/Turabian StyleVergara, Fredd, Amiu Shino, and Jun Kikuchi. 2016. "Cannibalism Affects Core Metabolic Processes in Helicoverpa armigera Larvae—A 2D NMR Metabolomics Study" International Journal of Molecular Sciences 17, no. 9: 1470. https://doi.org/10.3390/ijms17091470
APA StyleVergara, F., Shino, A., & Kikuchi, J. (2016). Cannibalism Affects Core Metabolic Processes in Helicoverpa armigera Larvae—A 2D NMR Metabolomics Study. International Journal of Molecular Sciences, 17(9), 1470. https://doi.org/10.3390/ijms17091470