
Liquid Hybridization and Solid Phase Detection: A Highly Sensitive and Accurate Strategy for MicroRNA Detection in Plants and Animals

 ,
,
Abstract
:1. Introduction
2. Results
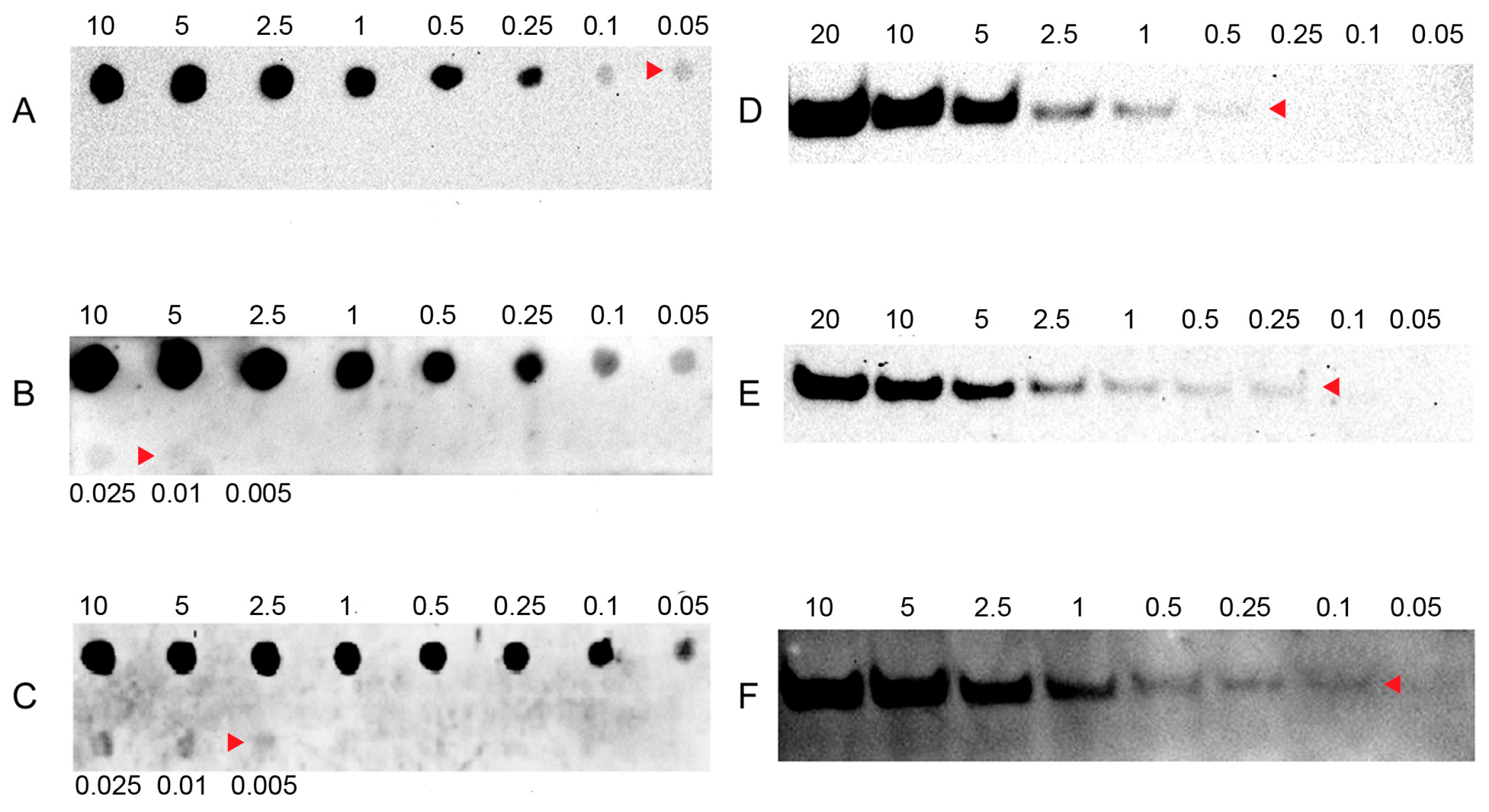
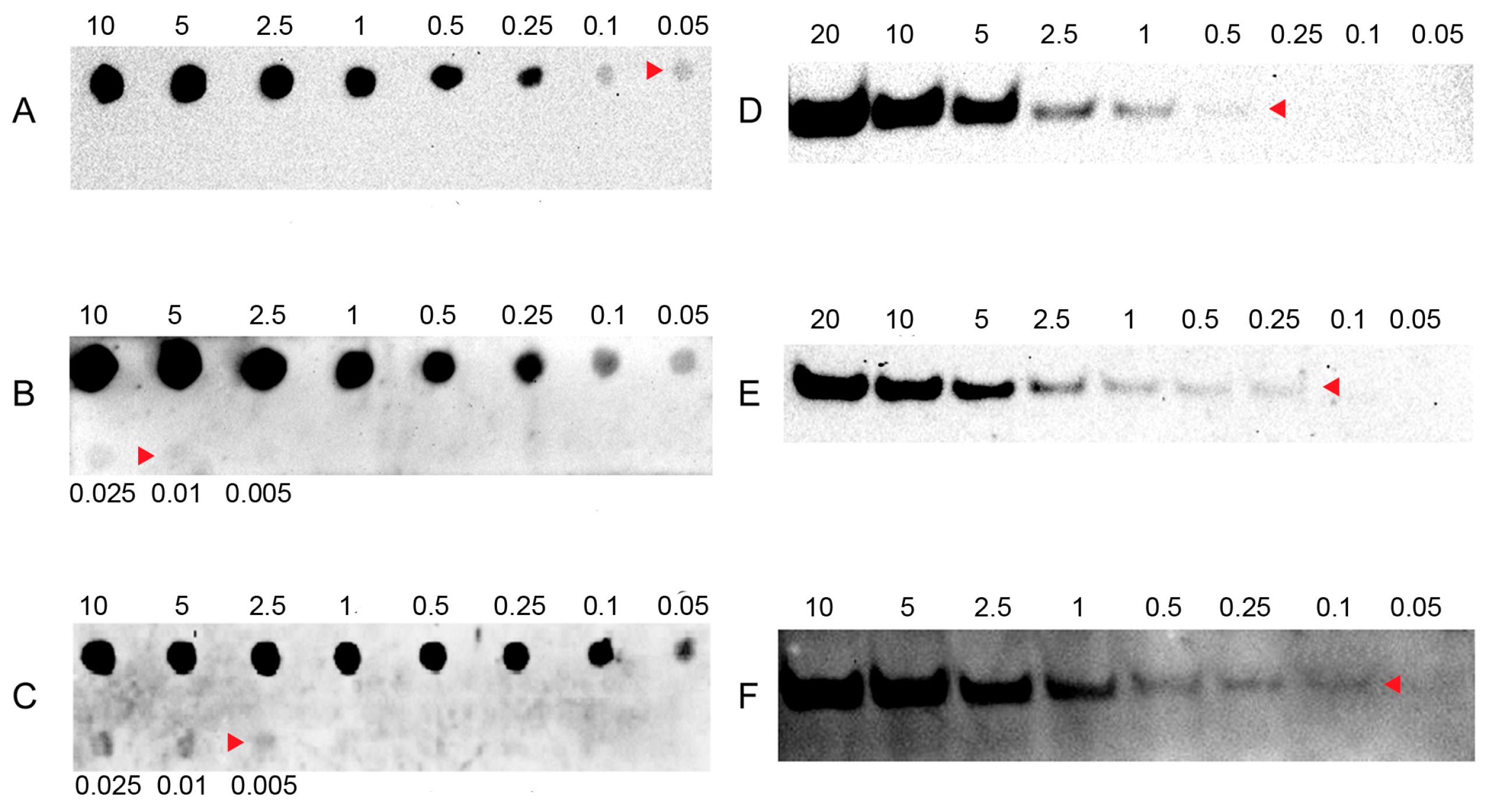
2.1. Sensitivity of LHSPD
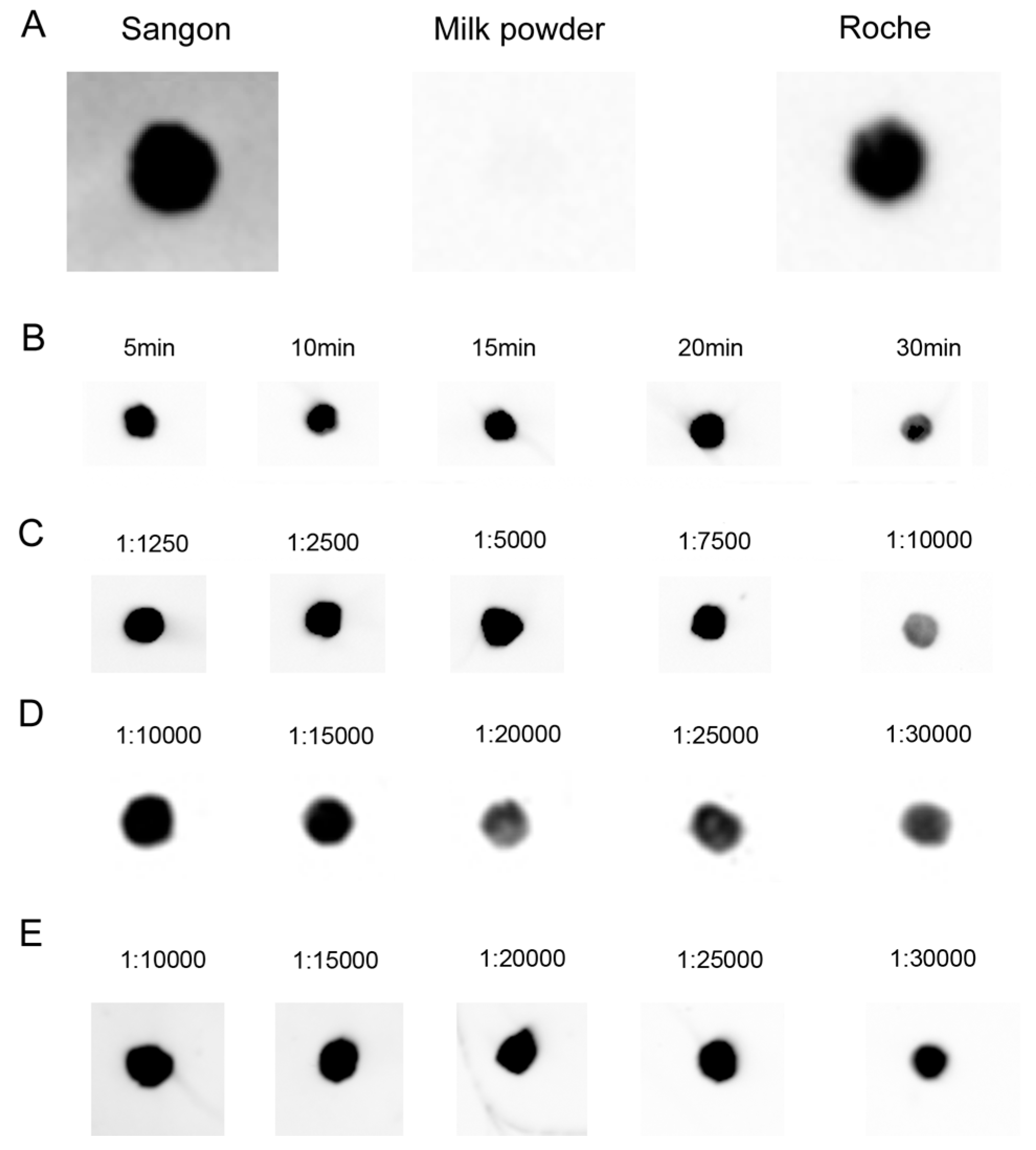
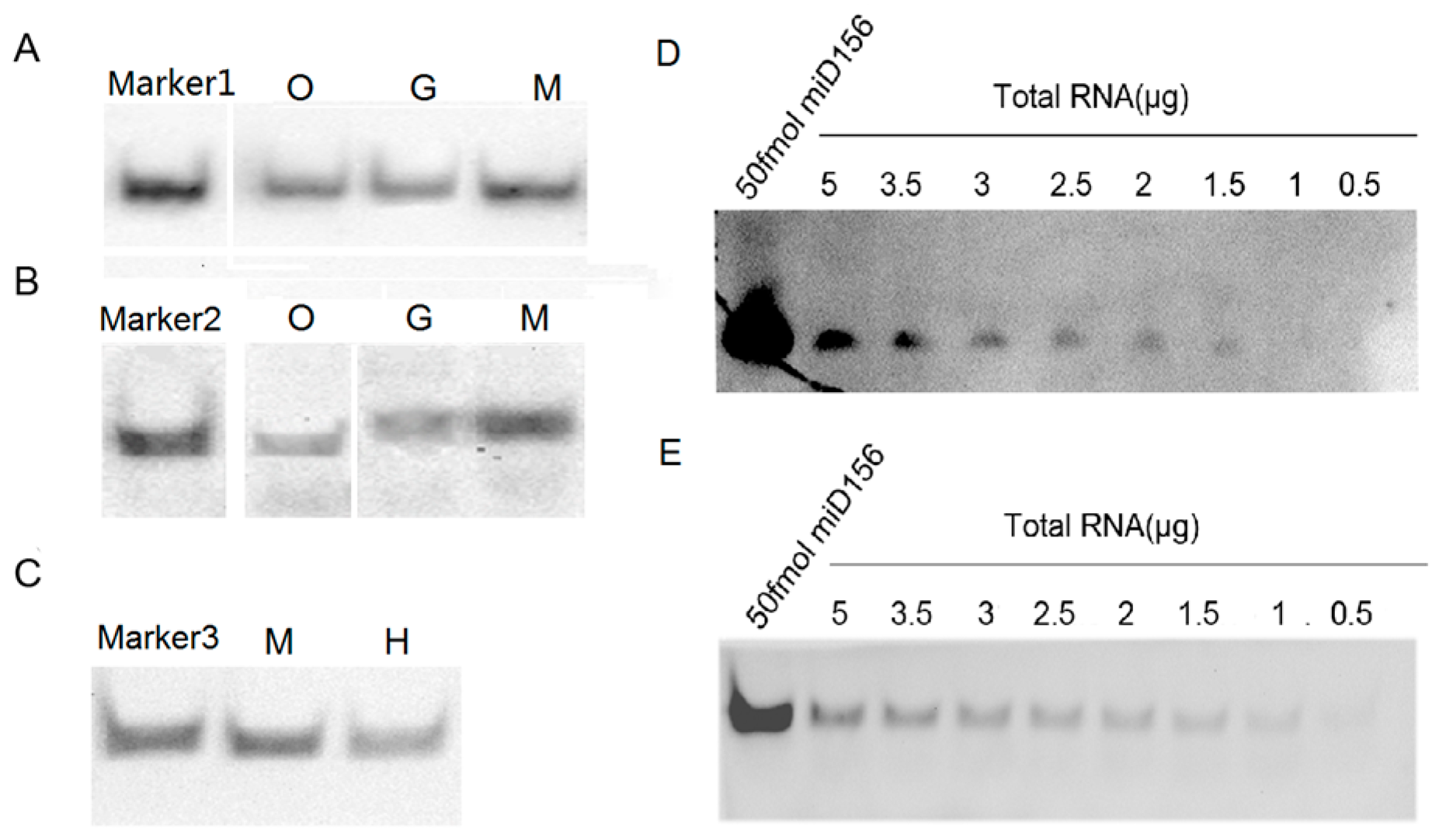
2.2. Specificity and Hybridization Buffer of LHSPD
2.3. Signal Detection on Solid Support Membrane
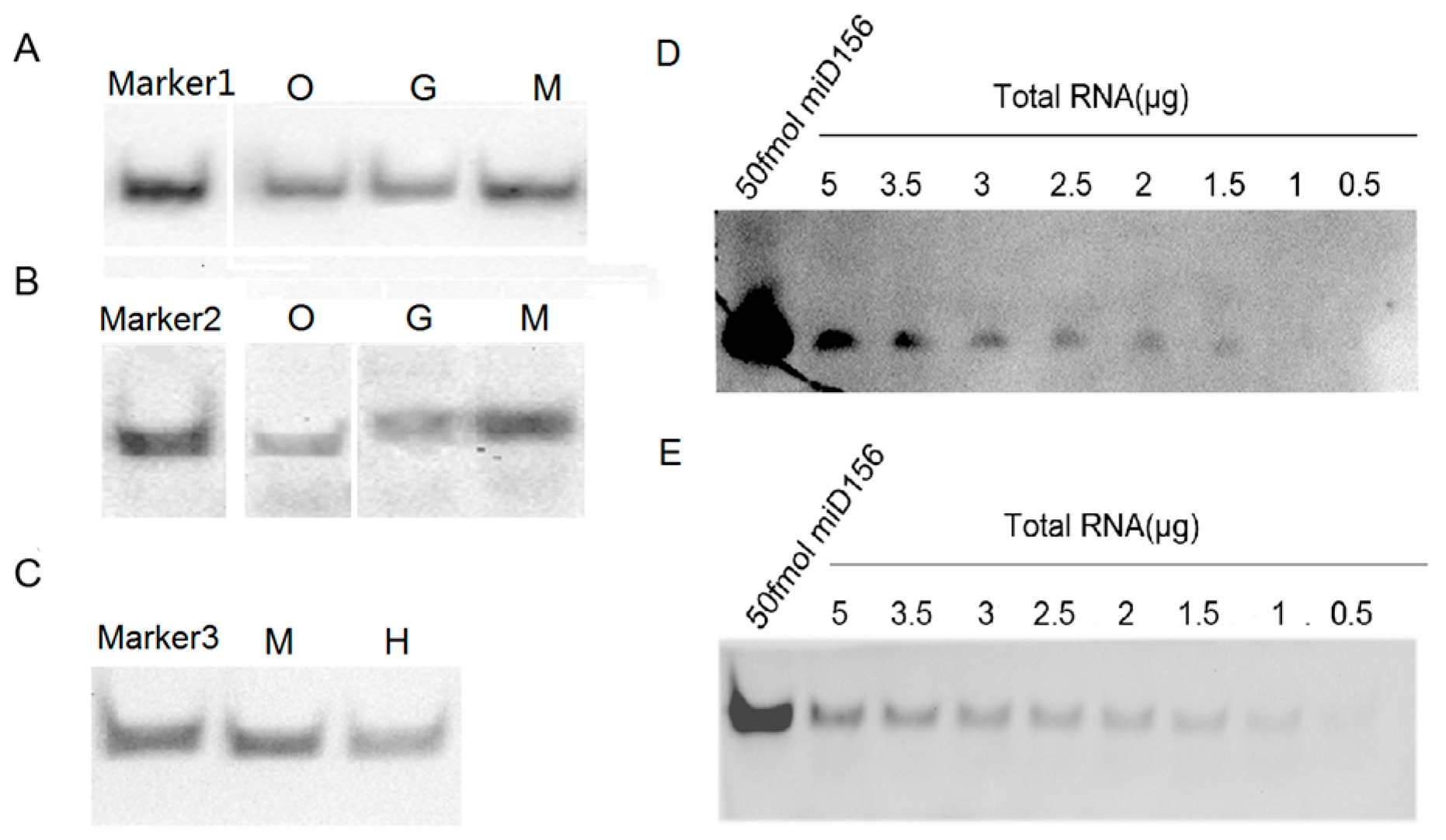
2.4. Detection of miRNAs with LHSPD
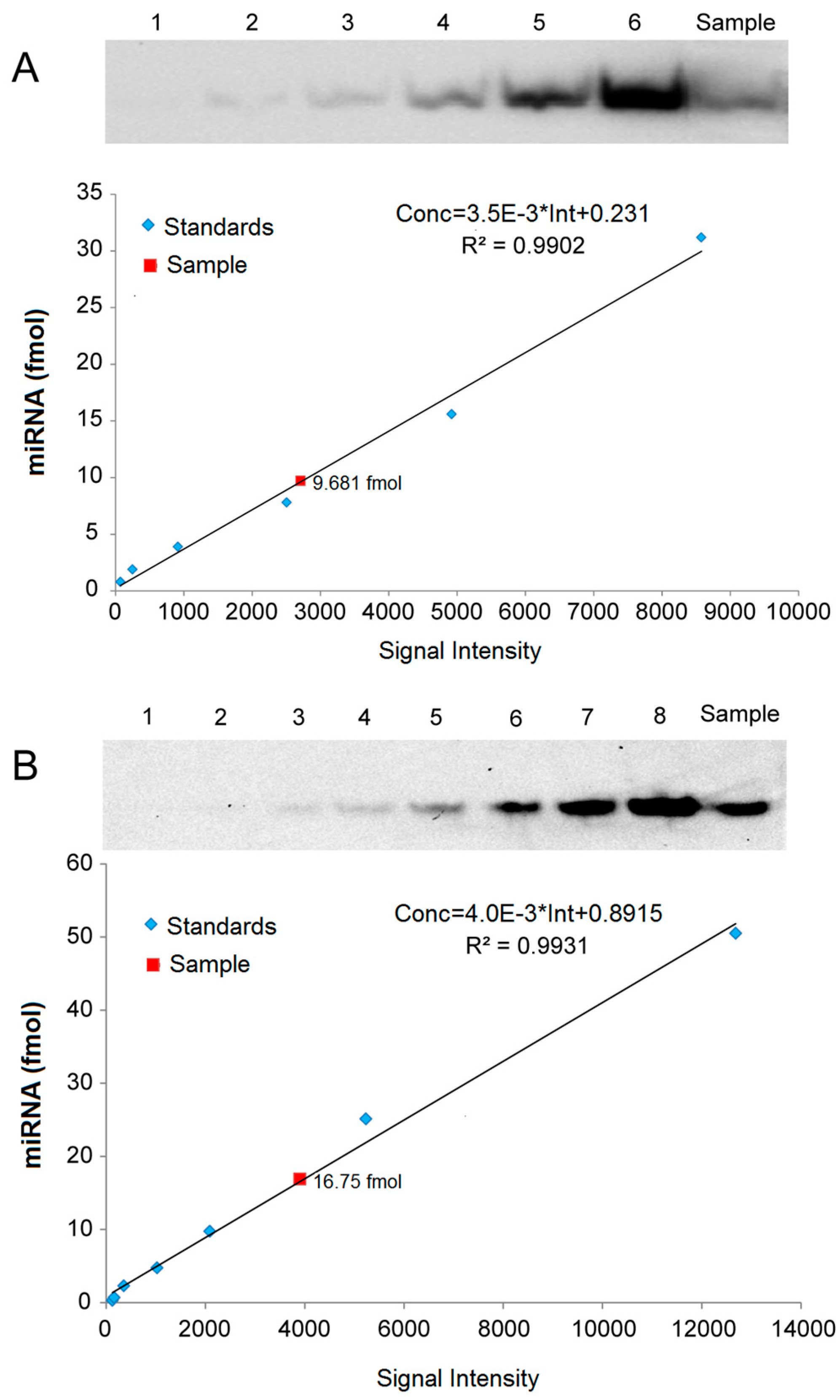
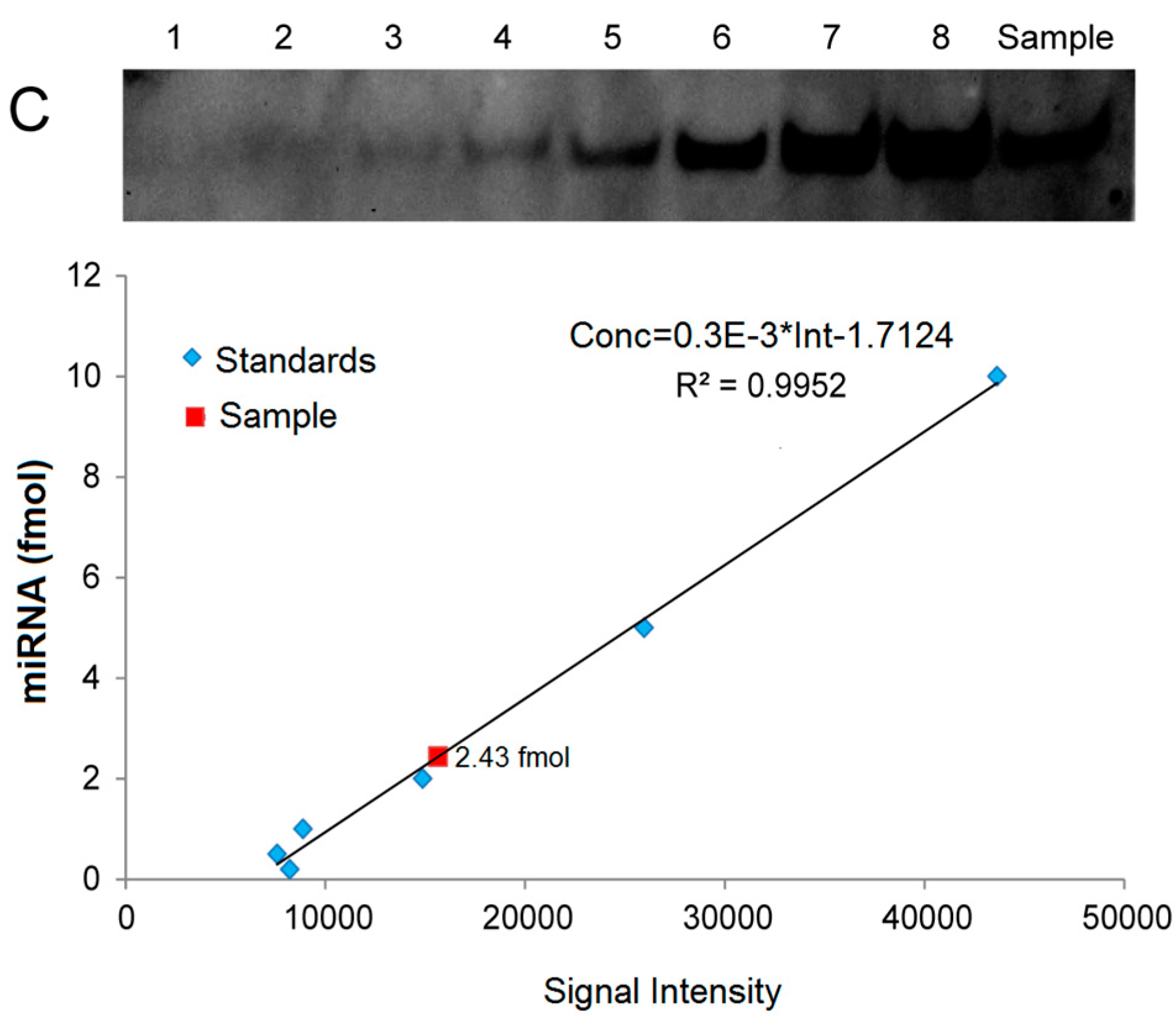
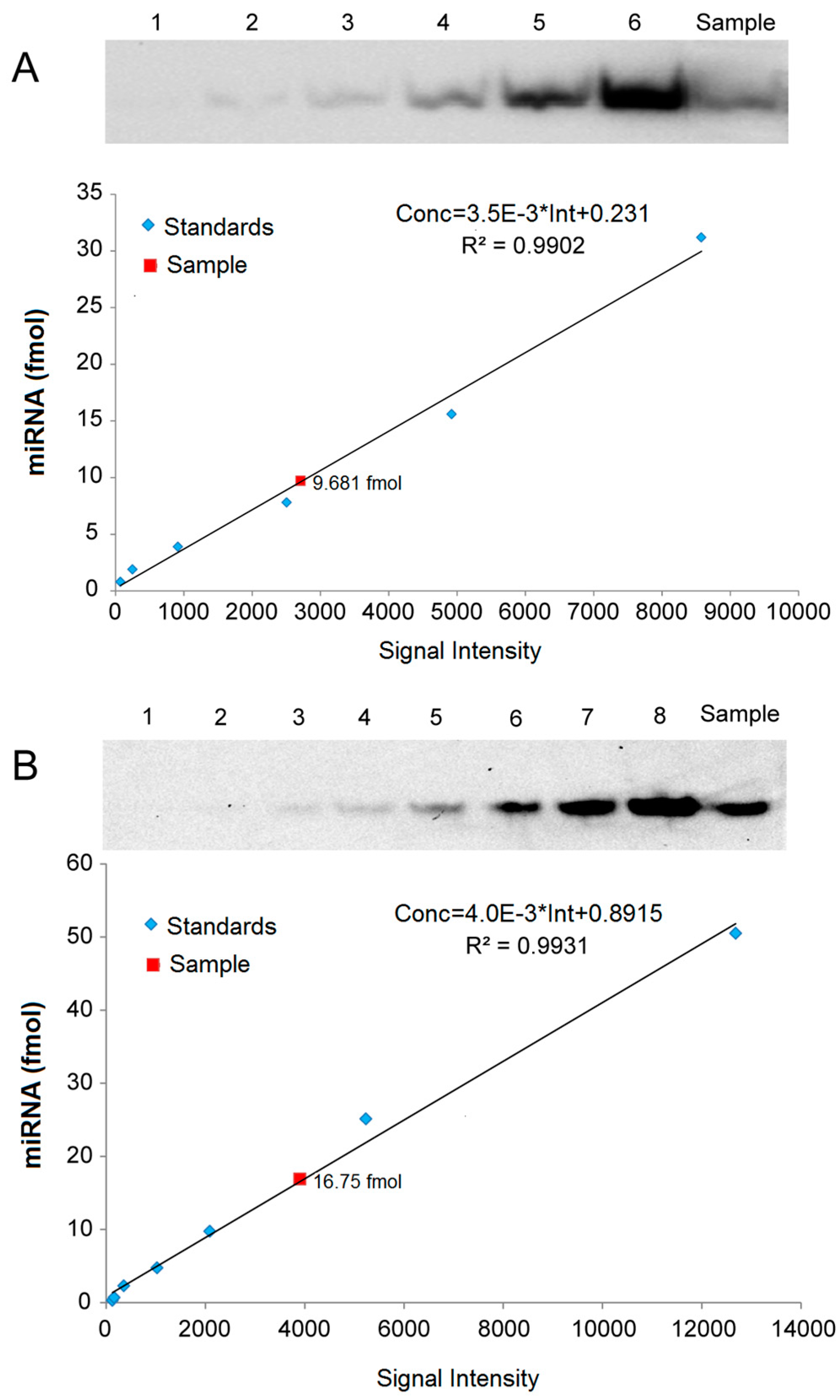
2.5. Quantitative Analysis of miRNAs by LHSPD
3. Discussion
4. Experimental Section
4.1. Plant and Animal Materials
4.2. Synthesis of Oligonucleotides and Hybridization Probes
4.3. Preparation of Small RNAs
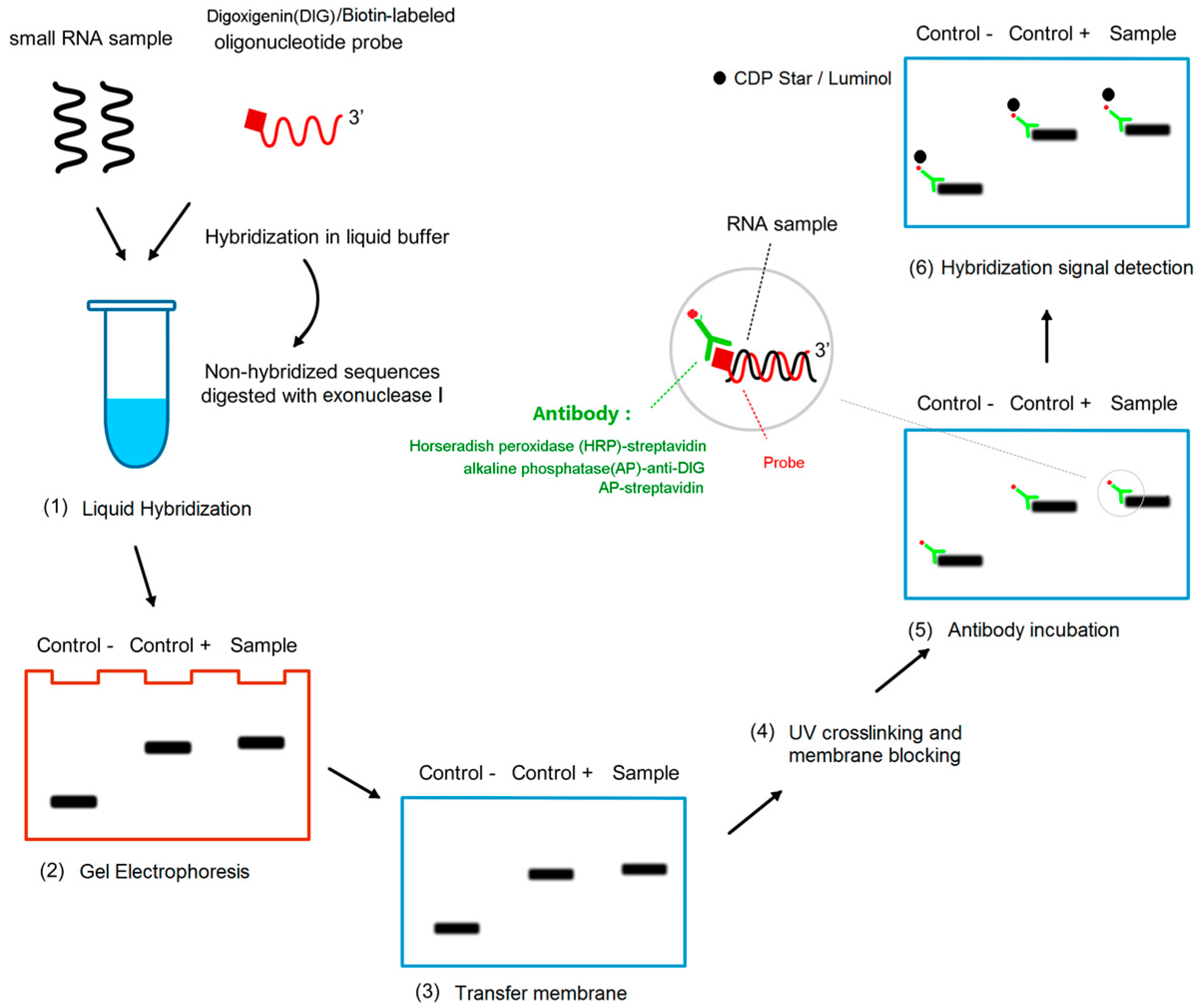
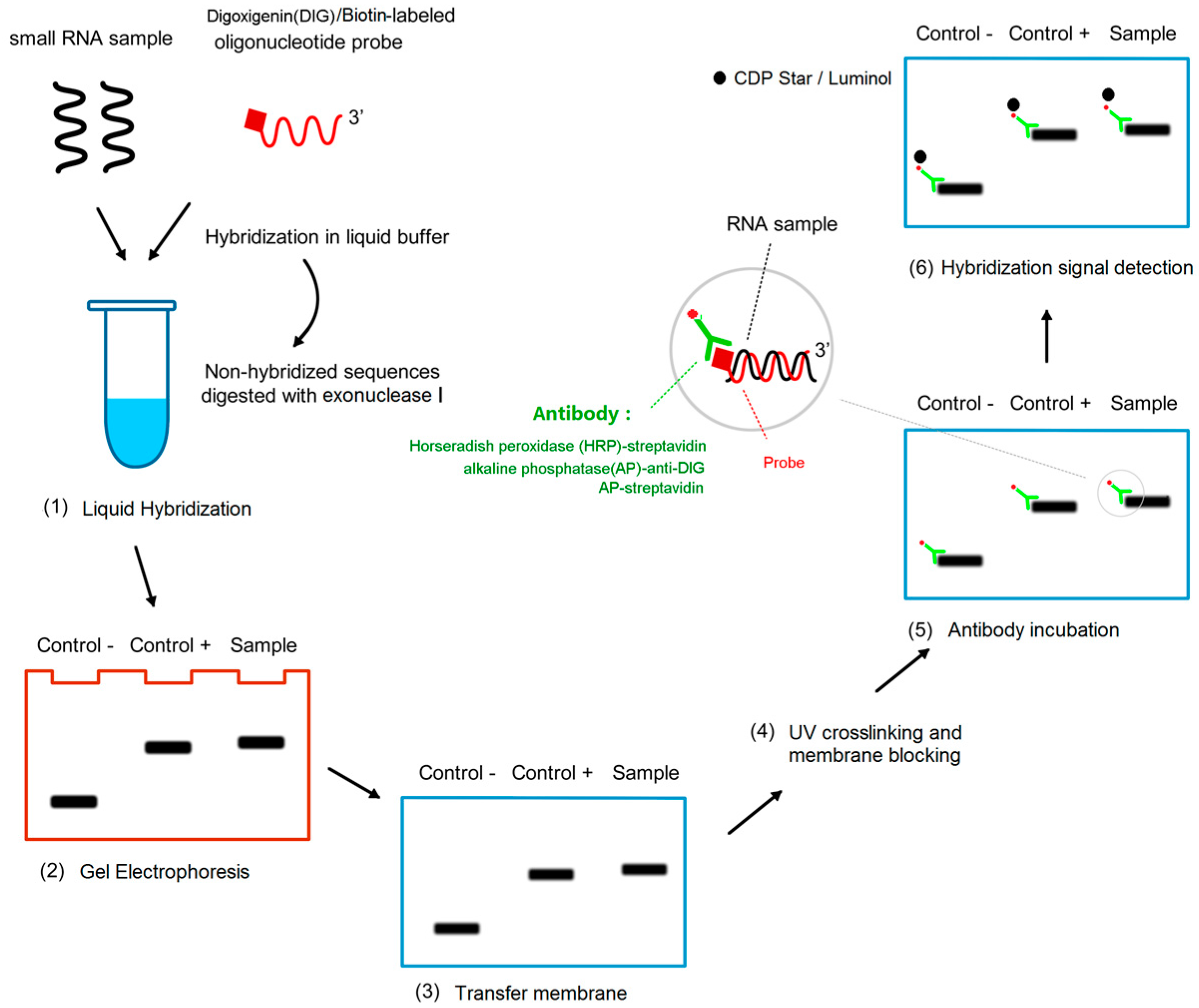
4.4. Protocol for miRNA Detection by LHSPD
4.4.1. Liquid Hybridization
4.4.2. Gel Electrophoresis
4.4.3. Transfer Membrane
4.4.4. UV Crosslinking and Membrane Blocking
4.4.5. Antibody Incubation
4.4.6. Hybridization Signal Detection
4.5. Traditional Northern Hybridization Strategy
4.5.1. Preparation of Denaturing PAGE and Gel Electrophoresis
4.5.2. Transfer Membrane and UV Crosslinking
4.5.3. Prehybridization and Hybridization
4.5.4. Washing Membrane
4.5.5. Antibody Incubation and Signal Detection
4.6. Quantitative Analysis of miRNA in Small RNA Samples
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhou, H.; Zhang, Q.; Zhang, J.; Ni, F.; Liu, C.; Qi, Y. DNA methylation mediated by a microRNA pathway. Mol. Cell 2010, 38, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Siomi, H.; Siomi, M.C. Expanding RNA physiology: MicroRNAs in a unicellular organism. Genes Dev. 2007, 21, 1153–1156. [Google Scholar] [CrossRef] [PubMed]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.C.; Vaucheret, H. Functions of microRNAs and related small RNAs in plants. Nat. Genet. 2006, 38, S31–S36. [Google Scholar] [CrossRef] [PubMed]
- Shukla, L.I.; Chinnusamy, V.; Sunkar, R. The role of microRNAs and other endogenous small RNAs in plant stress responses. Biochim. Biophys. Acta 2008, 1779, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Chinnusamy, V.; Zhu, J.; Zhu, J.K. Small RNAs as big players in plant abiotic stress responses and nutrient deprivation. Trends Plant Sci. 2007, 12, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. Plant microRNA: A small regulatory molecule with big impact. Dev. Biol. 2006, 289, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Griffiths-Jones, S. miRBase: The microRNA sequence database. Methods Mol. Biol. 2006, 342, 129–138. [Google Scholar] [PubMed]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 2004, 5, R13. [Google Scholar] [CrossRef] [PubMed]
- Grundhoff, A.; Sullivan, C.S.; Ganem, D. A combined computational and microarray-based approach identifies novel microRNAs encoded by human γ-herpesviruses. RNA 2006, 12, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Neely, L.A.; Patel, S.; Garver, J.; Gallo, M.; Hackett, M.; McLaughlin, S.; Nadel, M.; Harris, J.; Gullans, S.; Rooke, J. A single-molecule method for the quantitation of microRNA gene expression. Nat. Methods 2006, 3, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.J.; Li, X.B.; Li, Y.Y.; Zhao, L.F.; He, W.L.; Gao, Y.Q.; Wan, Y.J.; Xia, W.; Chen, T.; Zheng, H.; et al. Quantification of microRNA by gold nanoparticle probes. Anal. Biochem. 2008, 376, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Jonstrup, S.P.; Koch, J.; Kjems, J. A microRNA detection system based on padlock probes and rolling circle amplification. RNA 2006, 12, 1747–1752. [Google Scholar] [CrossRef] [PubMed]
- Allawi, H.T.; Dahlberg, J.E.; Olson, S.; Lund, E.; Olson, M.; Ma, W.P.; Takova, T.; Neri, B.P.; Lyamichev, V.I. Quantitation of microRNAs using a modified invader assay. RNA 2004, 10, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V.; Lee, R.C. Identification of microRNAs and other tiny noncoding RNAs by cDNA cloning. Methods Mol. Biol. 2004, 265, 131–158. [Google Scholar] [PubMed]
- Shi, R.; Chiang, V.L. Facile means for quantifying microRNA expression by real-time PCR. Biotechniques 2005, 39, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Cissell, K.A.; Deo, S.K. Trends in microRNA detection. Anal. Bioanal. Chem. 2009, 394, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Pall, G.S.; Hamilton, A.J. Improved Northern blot method for enhanced detection of small RNA. Nat. Protoc. 2008, 3, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Éva, V.; József, B.; Zoltán, H. MicroRNA detection by northern blotting using locked nucleic acid probes. Nat. Protoc. 2008, 3, 190–196. [Google Scholar]
- Válóczi, A.; Hornyik, C.; Varga, N.; Burgyán, J.; Kauppinen, S.; Havelda, Z. Sensitive and specific detection of microRNAs by Northern blot analysis using LNA-modified oligonucleotide probes. Nucleic Acids Res. 2004, 32, e175. [Google Scholar] [CrossRef] [PubMed]
- Éva, V.; Burgyán, J.; Havelda, Z. Detection of microRNAs by Northern blot analyses using LNA probes. Methods 2007, 43, 140–145. [Google Scholar]
- Ramkissoon, S.H.; Mainwaring, L.A.; Sloand, E.M.; Young, N.S.; Kajigaya, S. Nonisotopic detection of microRNA using digoxigenin labeled RNA probes. Mol. Cell. Probes 2006, 20, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Li, Z.; Moore, P.S.; Monaghan, A.P.; Chang, Y.; Nichols, M.; John, B. A sensitive non-radioactive Northern blot method to detect small RNAs. Nucleic Acids Res. 2010, 38, e98. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Gong, P.; Li, J.; Yang, J.; Zhang, G.; Li, H.; Yang, Z.; Zhang, X. Simple and nonradioactive detection of microRNAs using digoxigenin (DIG)-labeled probes with high sensitivity. RNA 2014, 20, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ni, M.; Zhang, Y. Detecting miRNAs by liquid hybridization and color development. Methods 2012, 58, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tong, Y.; Wang, S. Rapid and accurate detection of plant miRNAs by liquid Northern hybridization. Int. J. Mol. Sci. 2010, 11, 3138–3148. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.A.; Peng, H.; Zhan, C.; Fan, L.; Ai, T.; Wang, S. Genome-wide analysis reveals diversity of rice intronic miRNAs in sequence structure, biogenesis and function. PLoS ONE 2013, 8, e63938. [Google Scholar] [CrossRef] [PubMed]
- Chilkoti, A.; Stayton, P.S. Molecular origins of the slow streptavidin-biotin dissociation kinetics. J. Am. Chem. Soc. 1995, 117, 10622–10628. [Google Scholar] [CrossRef]
- Casey, J.; Davidson, N. Rates of formation and thermal stabilities of RNA:DNA and DNA:DNA duplexes at high concentrations of formamide. Nucleic Acids Res. 1977, 4, 1539–1552. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-time quantification of microRNAs by stem–loop RT-PCR. Nucleic Acids Res. 2005, 33, e179–e179. [Google Scholar] [CrossRef] [PubMed]
- Maroney, P.A.; Chamnongpol, S.; Souret, F.; Nilsen, T.W. A rapid, quantitative assay for direct detection of microRNAs and other smallRNAs using splinted ligation. RNA 2007, 13, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.T.; Baldwin, D.A.; Scearce, L.M.; Oberholtzer, J.C.; Tobias, J.W.; Mourelatos, Z. Microarray-based, high-throughput gene expression profiling of microRNAs. Nat. Methods 2004, 1, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Deo, M.; Turner, D.L. Analysis of microRNA expression by in situ hybridization with RNA oligonucleotide probe. Methods 2007, 43, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Zheng, Y.F.; Wang, S.H.; Chen, F. A RNA isolation method suitable for a wide range of materials. Prog. Biochem. Biophys. 2008, 35, 591–597. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names | Sequence (5′–3′) | Tm (°C) | Mw |
|---|---|---|---|
| Osa-miR156 | UGA CAG AAG AGA GUG AGC AC | 57.3 | 6279 |
| (Biotin)-miD156rk | (Biotin)-A CGT GCT CAC TCT CTT CTG TCA | 60.5 | 6587+ |
| (DIG)-miD156rk | (DIG)-A CGT GCT CAC TCT CTT CTG TCA | 60.5 | 6587+ |
| miD156 | TGA CAG AAG AGA GTG AGC AC | 57.3 | 6201+ |
| miD156(−1) | TGA gAG AAG AGA GTG AGC AC | 55.4 | 6241 |
| miD156(−3) | aGA CAG gAG gGA GTG AGC AC | 57.4 | 6242 |
| miD156(−5) | aGA CAG gAG tGA GTc AGC gC | 59.5 | 6193+ |
| let-7a | UGA GGU AGU AGG UUG UAU AGU U | 46.1 | 7244 |
| (Biotin)-let-7rk | (Biotin)-ACA ACT ATA CAA CCT ACT ACC TCA | 52.6 | 7166+ |
| (DIG)-let-7rk | (DIG)-ACA ACT ATA CAA CCT ACT ACC TCA | 52.6 | 7166+ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Mei, L.; Zhan, C.; Mao, Q.; Yao, M.; Wang, S.; Tang, L.; Chen, F. Liquid Hybridization and Solid Phase Detection: A Highly Sensitive and Accurate Strategy for MicroRNA Detection in Plants and Animals. Int. J. Mol. Sci. 2016, 17, 1457. https://doi.org/10.3390/ijms17091457
Li F, Mei L, Zhan C, Mao Q, Yao M, Wang S, Tang L, Chen F. Liquid Hybridization and Solid Phase Detection: A Highly Sensitive and Accurate Strategy for MicroRNA Detection in Plants and Animals. International Journal of Molecular Sciences. 2016; 17(9):1457. https://doi.org/10.3390/ijms17091457
Chicago/Turabian StyleLi, Fosheng, Lanju Mei, Cheng Zhan, Qiang Mao, Min Yao, Shenghua Wang, Lin Tang, and Fang Chen. 2016. "Liquid Hybridization and Solid Phase Detection: A Highly Sensitive and Accurate Strategy for MicroRNA Detection in Plants and Animals" International Journal of Molecular Sciences 17, no. 9: 1457. https://doi.org/10.3390/ijms17091457
APA StyleLi, F., Mei, L., Zhan, C., Mao, Q., Yao, M., Wang, S., Tang, L., & Chen, F. (2016). Liquid Hybridization and Solid Phase Detection: A Highly Sensitive and Accurate Strategy for MicroRNA Detection in Plants and Animals. International Journal of Molecular Sciences, 17(9), 1457. https://doi.org/10.3390/ijms17091457