
Pigment Epithelium-Derived Factor (PEDF) Protects Osteoblastic Cell Line from Glucocorticoid-Induced Apoptosis via PEDF-R

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
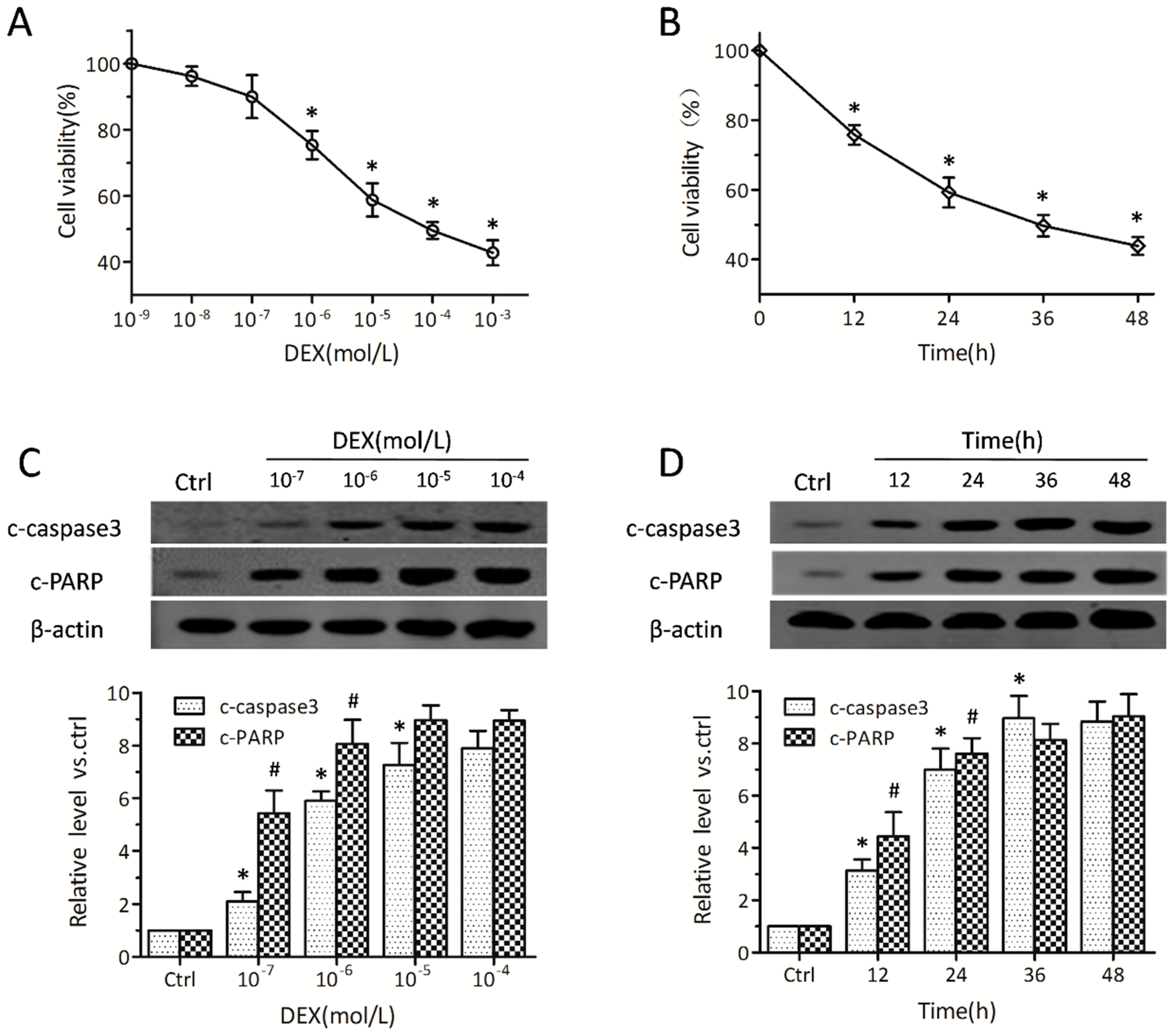
2.1. Dexamethasone Induces Apoptosis in MC3T3-E1 Pre-Osteoblasts
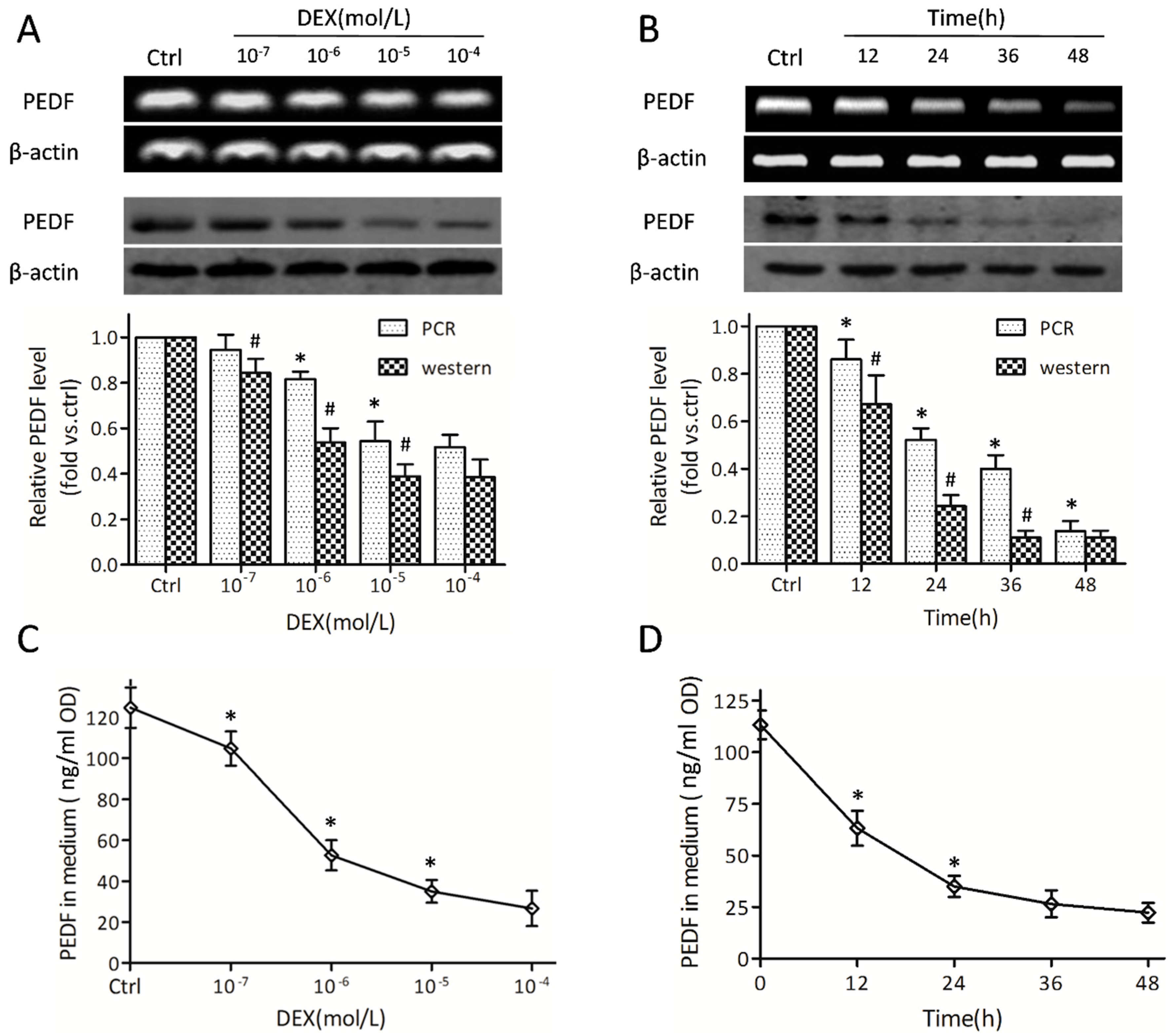
2.2. Dexamethasone Regulates Pigment Epithelial-Derived Factor (PEDF) at the Gene and Protein Levels
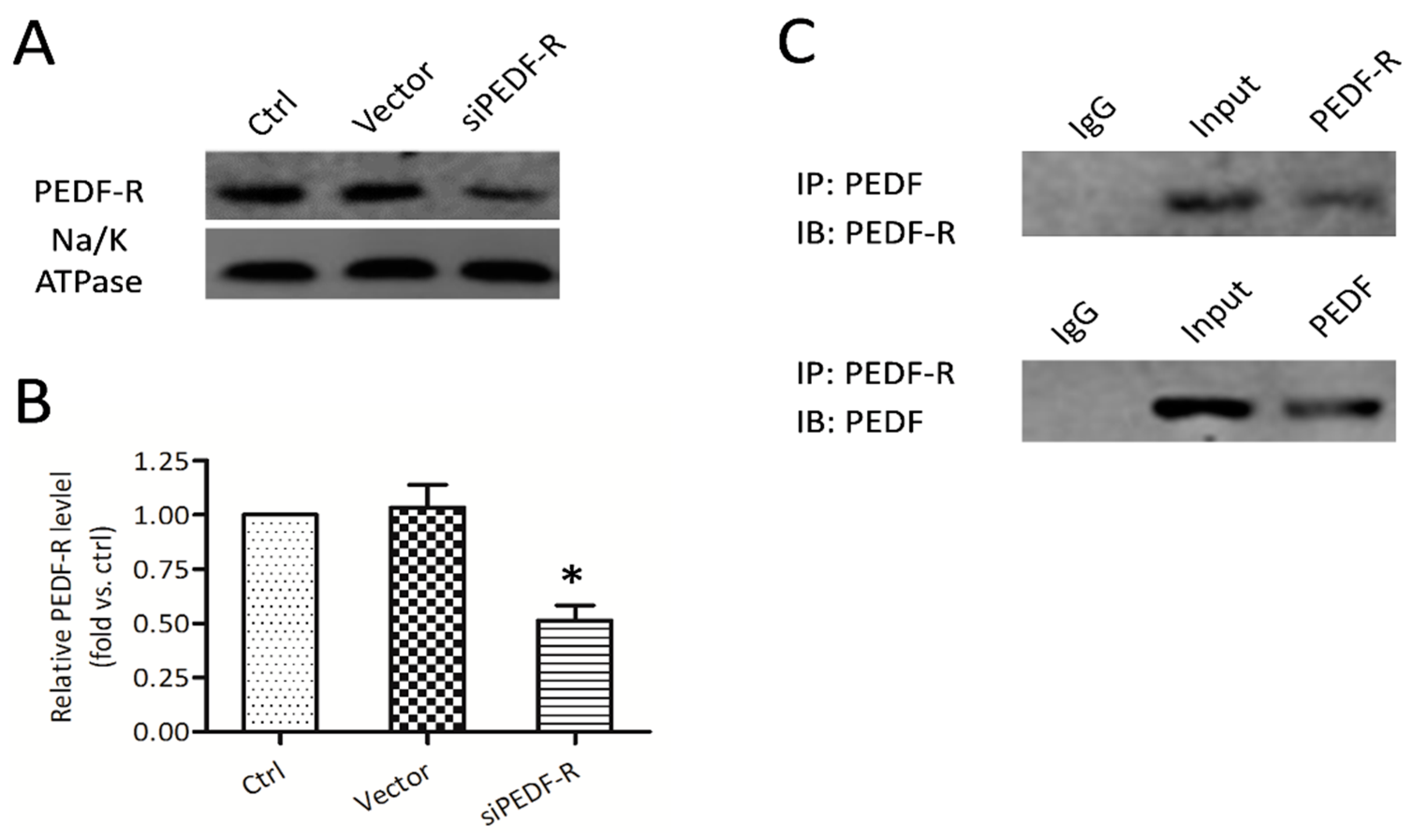
2.3. PEDF Binds to PEDF receptor (PEDF-R) on the Osteoblast Membrane
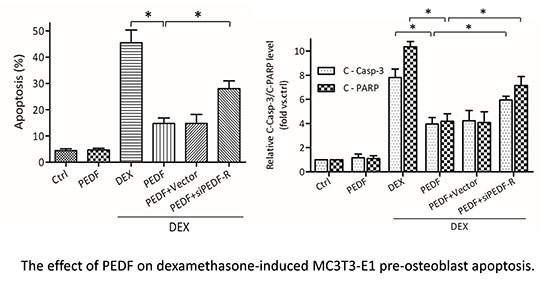
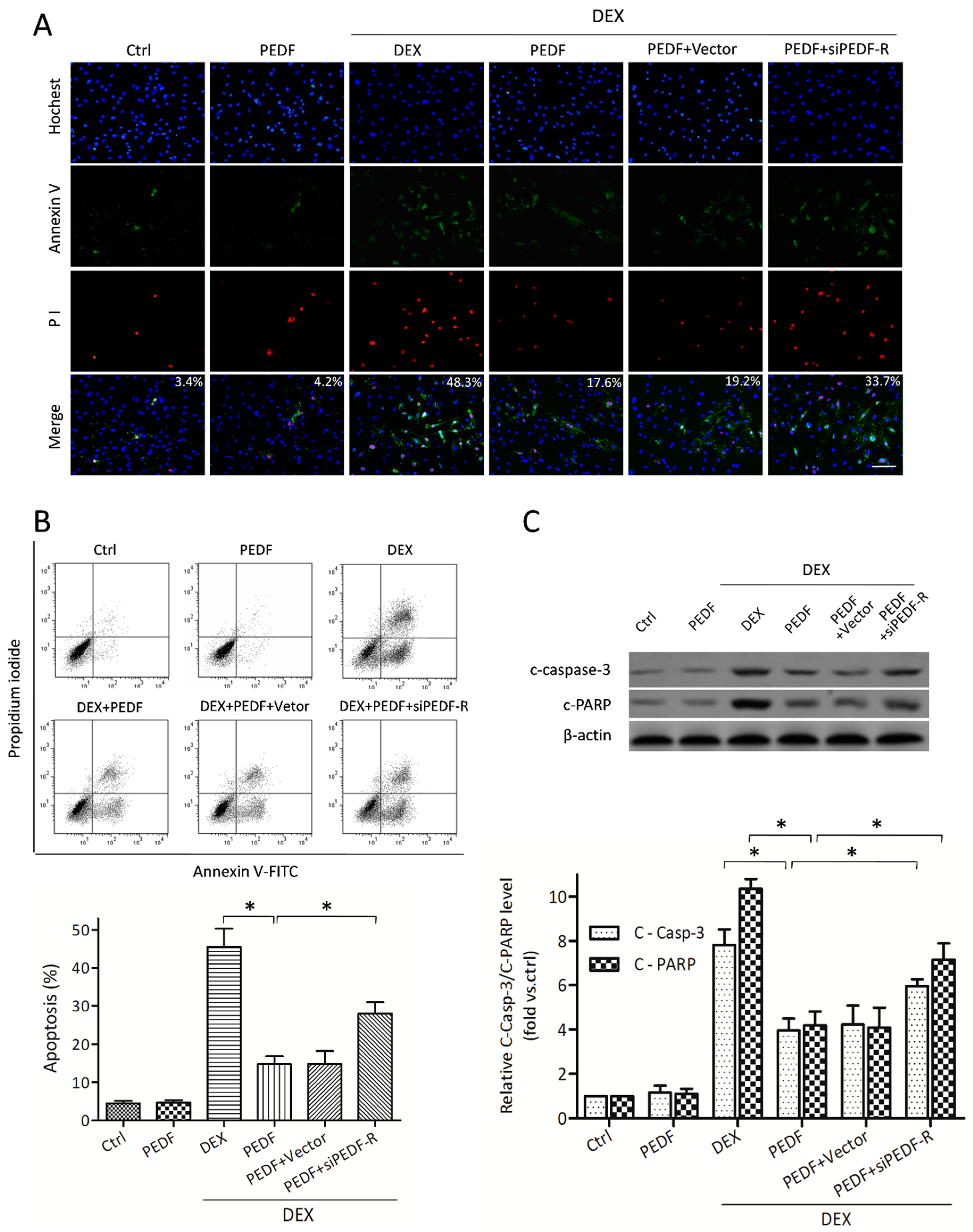
2.4. PEDF Attenuates DEX-Induced MC3T3-E1 Pre-Osteoblast Apoptosis via PEDF-R
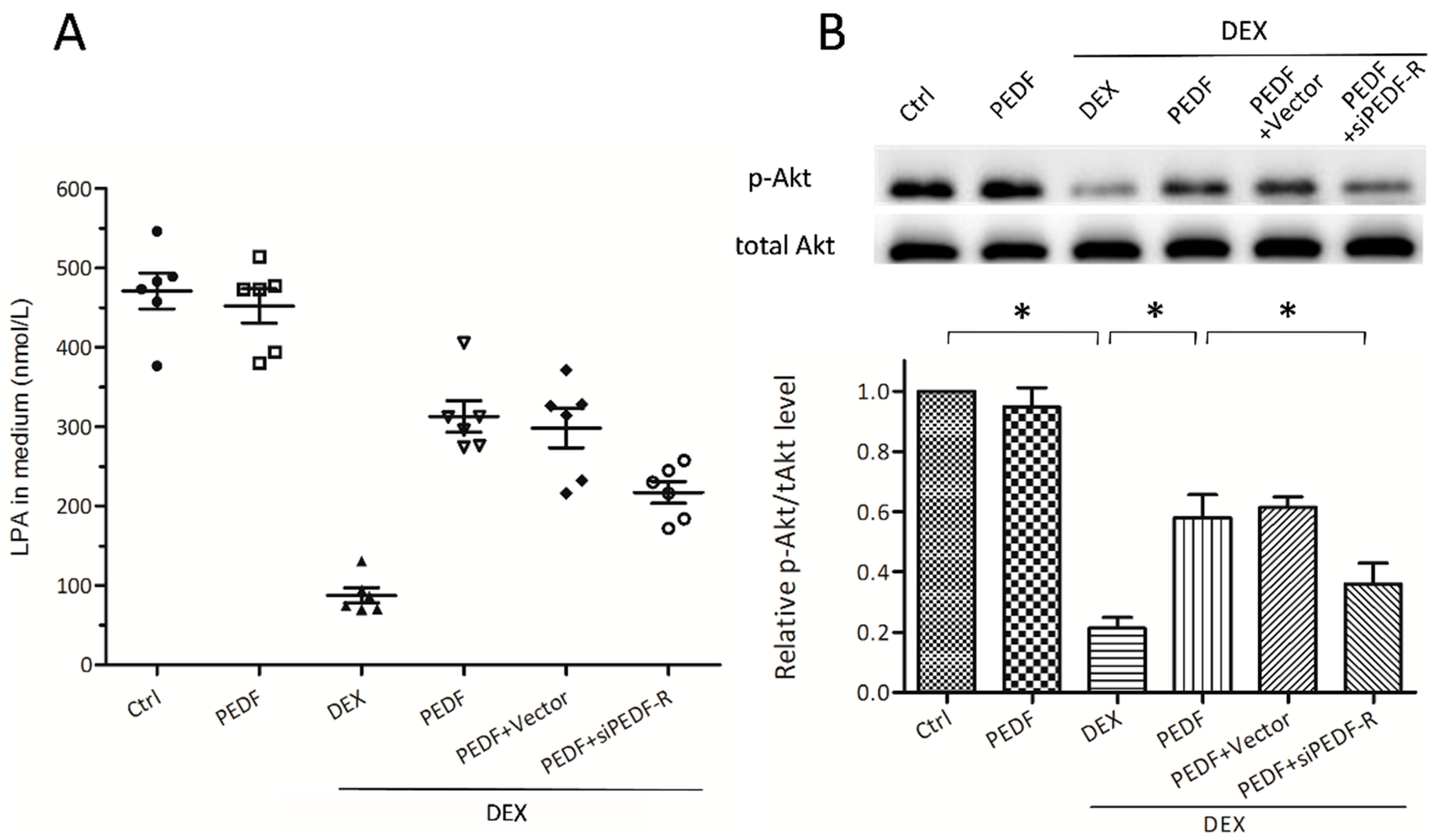
2.5. PEDF-R Mediates PEDF-Increased Lysophosphatidic Acid (LPA) Levels, Activating Phosphatidylinositol 3-Kinase (PI3K)/Akt Signaling in MC3T3-E1 Pre-Osteoblasts
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Recombinant Lentivirus Constructs and Viral Production
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Western Blot Analysis
4.6. Measurement of PEDF mRNA Expression
4.7. Co-Immunoprecipitation
4.8. Cell Apoptosis Assessment
4.9. Quantification of PEDF and LPA
4.10. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PEDF | Pigment epithelium-derived factor |
| DEX | Dexamethasone |
| rPEDF | Recombinant PEDF |
| PEDF-R | Pigment epithelial-derived factor receptor |
| GCs | Glucocorticoids |
| GR | Glucocorticoid receptor |
| LPA | Lysophosphatidic acid |
| PNPLA2 | Patatin-like phospholipase domain-containing 2 |
| ATGL | Adipose triglyceride lipase |
| pAkt | Phosphorylated Akt |
| LBD | Ligand binding domain |
References
- Weinstein, R.S. Clinical practice. Glucocorticoid-induced bone disease. N. Engl. J. Med. 2011, 365, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Canalis, E.; Mazziotti, G.; Giustina, A.; Bilezikian, J.P. Glucocorticoid-induced osteoporosis: Pathophysiology and therapy. Osteoporos. Int. 2007, 18, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, R.S. Glucocorticoid-induced osteoporosis and osteonecrosis. Endocrinol. Metab. Clin. N. Am. 2012, 41, 595–611. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.A.; Zheng, H.X.; Wang, C.W.; Shi, D.; Li, J.J. Influence of glucocorticoids on the osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. BMC Musculoskelet. Disord. 2014, 15, 239. [Google Scholar] [CrossRef] [PubMed]
- Kósa, J.P.; Kis, A.; Bácsi, K.; Balla, B.; Nagy, Z.; Takács, I.; Speer, G.; Lakatos, P. The protective role of bone morphogenetic protein-8 in the glucocorticoid-induced apoptosis on bone cells. Bone 2011, 48, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, L.; OuYang, Y.; Guan, H.; Liu, Q.; Ni, B. Glucocorticoid induced osteoblast apoptosis by increasing E4BP4 expression via up-regulation of Bim. Calcif. Tissue Int. 2014, 94, 640–647. [Google Scholar] [PubMed]
- Moutsatsou, P.; Kassi, E.; Papavassiliou, A.G. Glucocorticoid receptor signaling in bone cells. Trends Mol. Med. 2012, 18, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Jewell, C.M.; Scoltock, A.B.; Hamel, B.L.; Yudt, M.R.; Cidlowski, J.A. Complex human glucocorticoid receptor dim mutations define glucocorticoid induced apoptotic resistance in bone cells. Mol. Endocrinol. 2012, 26, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Rauch, A.; Seitz, S.; Baschant, U.; Schilling, A.F.; Illing, A.; Stride, B.; Kirilov, M.; Mandic, V.; Takacz, A.; Schmidt-Ullrich, R.; et al. Glucocorticoids suppress bone formation by attenuating osteoblast differentiation via the monomeric glucocorticoid receptor. Cell Metab. 2010, 11, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Quan, G.M.; Ojaimi, J.; Li, Y.; Kartsogiannis, V.; Zhou, H.; Choong, P.F. Localization of pigment epithelium-derived factor in growing mouse bone. Calcif. Tissue Int. 2005, 76, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Tombran-Tink, J.; Chader, G.G.; Johnson, L.V. PEDF: A pigment epithelium derived factor with potent neuronal differentiative activity. Exp. Eye Res. 1991, 53, 411–414. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Yamagishi, S.I.; Sata, M. Structure–Function Relationships of PEDF. Curr. Mol. Med. 2010, 10, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Yabe, T.; Wilson, D.; Schwartz, J.P. NFκB activation is required for the neuroprotective effects of pigment epithelium-derived factor (PEDF) on cerebellar granule neurons. J. Biol. Chem. 2001, 276, 43313–43319. [Google Scholar] [CrossRef] [PubMed]
- Imai, D.; Yoneya, S.; Gehlbach, P.L.; Wei, L.L.; Mori, K. Intraocular gene transfer of pigment epithelium-derived factor rescues photoreceptors from light-induced cell death. J. Cell. Physiol. 2005, 202, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Z.; Feng, S.J.; Xu, L.; Shi, H.X.; Chen, L.L.; Yuan, G.D.; Yan, W.; Zhuang, W.; Zhang, Y.Q.; et al. PEDF improves cardiac function in rats with acute myocardial infarction via inhibiting vascular permeability and cardiomyocyte apoptosis. Int. J. Mol. Sci. 2015, 16, 5618–5634. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Song, N.; Tombran-Tink, J.; Niyibizi, C. Pigment epithelium-derived factor enhances differentiation and mineral deposition of human mesenchymal stem cells. Stem Cells 2013, 31, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Broadhead, M.L.; Akiyama, T.; Choong, P.F.; Dass, C.R. The Pathophysiological Role of PEDF in Bone Diseases. Curr. Mol. Med. 2010, 10, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, P.; Locatelli-Hoops, S.; Kenealey, J.; DesJardin, J.; Notari, L.; Becerra, S.P. Pigment epithelium-derived factor (PEDF) prevents retinal cell death via PEDF Receptor (PEDF-R): Identification of a functional ligand binding site. J. Biol. Chem. 2013, 288, 23928–23942. [Google Scholar] [CrossRef] [PubMed]
- Alberdi, E.; Aymerich, M.S.; Becerra, S.P. Binding of pigment epithelium-derived factor (PEDF) to retinoblastoma cells and cerebellar granule neurons. Evidence for a PEDF receptor. J. Biol. Chem. 1999, 274, 31605–31612. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.M.; Mancuso, D.J.; Yan, W.; Sims, H.F.; Gibson, B.; Gross, R.W. Identification, cloning, expression, and purification of three novel human calcium-independent phospholipase A2 family members possessing triacylglycerol lipase and acylglycerol transacylase activities. J. Biol. Chem. 2004, 279, 48968–48975. [Google Scholar] [CrossRef] [PubMed]
- Borg, M.L.; Andrews, Z.B.; Duh, E.J.; Zechner, R.; Meikle, P.J.; Watt, M.J. Pigment Epithelium–Derived Factor Regulates Lipid Metabolism via Adipose Triglyceride Lipase. Diabetes 2011, 60, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, P.; Notario, P.M.; Becerra, S.P. Pigment epithelium-derived factor receptor (PEDF-R). A plasma membrane-linked phospholipase with PEDF binding affinity. Adv. Exp. Med. Biol. 2010, 664, 29–37. [Google Scholar] [PubMed]
- Grey, A.; Chen, Q.; Callon, K.; Xu, X.; Reid, I.R.; Cornish, J. The phospholipids sphingosine-1-phosphate and lysophosphatidic acid prevent apoptosis in osteoblastic cells via a signaling pathway involving Gi proteins and phosphatidylinositol-3 kinase. Endocrinology 2002, 143, 4755–4763. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.Z.; Collins, J.B.; Grissom, S.F.; Cidlowski, J.A. Selective regulation of bone cell apoptosis by translational isoforms of the glucocorticoid receptor. Mol. Cell. Biol. 2007, 27, 7143–7160. [Google Scholar] [CrossRef] [PubMed]
- Espina, B.; Liang, M.; Russell, R.G.; Hulley, P.A. Regulation of bim in glucocorticoid-mediated osteoblast apoptosis. J. Cell. Physiol. 2008, 215, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, T.; Jiang, X.; Yu, H.; Wang, M.; Wei, T.; Cui, H.; Zhuang, W.; Liu, Z.; Zhang, Z.; et al. PEDF and PEDF-derived peptide 44mer stimulate cardiac triglyceride degradation via ATGL. J. Transl. Med. 2015, 13, 68. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Y.; Lu, P.; Zhang, H.; Li, Y.; Dong, H.; Zhang, Z. PEDF attenuates hypoxia-induced apoptosis and necrosis in H9c2 cells by inhibiting p53 mitochondrial translocation via PEDF-R. Biochem. Biophys. Res. Commun. 2015, 465, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Fan, X.S.; Cai, L.; Liu, S.; Cong, X.F.; Chen, X. Lysophosphatidic acid rescues bone mesenchymal stem cells from hydrogen peroxide-induced apoptosis. Apoptosis 2015, 20, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Gattu, A.K.; Swenson, E.S.; Iwakiri, Y.; Samuel, V.T.; Troiano, N.; Berry, R.; Church, C.D.; Rodeheffer, M.S.; Carpenter, T.O.; Chung, C. Determination of mesenchymal stem cell fate by pigment epithelium-derived factor (PEDF) results in increased adiposity and reduced bone mineral content. FASEB J. 2013, 27, 4384–4394. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Zhan, X.; Li, X.; Zhao, J.; Chen, Y. Vanadate inhibits dexamethasone-induced apoptosis of rat bone marrow-derived mesenchymal stem cells. Ann. Clin. Lab. Sci. 2015, 45, 173–180. [Google Scholar] [PubMed]
- He, X.; Cheng, R.; Benyajati, S.; Ma, J.X. PEDF and its roles in physiological and pathological conditions: Implication in diabetic and hypoxia-induced angiogenic diseases. Clin. Sci. 2015, 128, 805–823. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Dass, C.R.; Shinoda, Y.; Kawano, H.; Tanaka, S.; Choong, P.F. PEDF regulates osteoclasts via osteoprotegerin and RANKL. Biochem. Biophys. Res. Commun. 2010, 391, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.H.; Shih, S.C.; Ho, T.C.; Ma, H.I.; Liu, M.Y.; Chen, S.L.; Tsao, Y.P. Pigment epithelium-derived factor 34-mer peptide prevents liver fibrosis and hepatic stellate cell activation through down-regulation of the PDGF receptor. PLoS ONE 2014, 9, e95443. [Google Scholar] [CrossRef] [PubMed]
- Bilak, M.M.; Becerra, S.P.; Vincent, A.M.; Moss, B.H.; Aymerich, M.S.; Kuncl, R.W. Identification of the neuroprotective molecular region of pigment epithelium-derived factor and its binding sites on motor neurons. J. Neurosci. 2002, 22, 9378–9386. [Google Scholar] [PubMed]
- Kenealey, J.; Subramanian, P.; Comitato, A.; Bullock, J.; Keehan, L.; Polato, F.; Hoover, D.; Marigo, V.; Becerra, S.P. Small retinoprotective peptides reveal a receptor-binding region on pigment epithelium-derived factor. J. Biol. Chem. 2015, 290, 25241–25253. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, S.; Zhang, Y.; Wang, X.; Zhao, F.; Sun, M.; Zheng, X.; Dong, H.; Guo, K. Pigment Epithelium-Derived Factor (PEDF) Protects Osteoblastic Cell Line from Glucocorticoid-Induced Apoptosis via PEDF-R. Int. J. Mol. Sci. 2016, 17, 730. https://doi.org/10.3390/ijms17050730
Yao S, Zhang Y, Wang X, Zhao F, Sun M, Zheng X, Dong H, Guo K. Pigment Epithelium-Derived Factor (PEDF) Protects Osteoblastic Cell Line from Glucocorticoid-Induced Apoptosis via PEDF-R. International Journal of Molecular Sciences. 2016; 17(5):730. https://doi.org/10.3390/ijms17050730
Chicago/Turabian StyleYao, Shengcheng, Yingnan Zhang, Xiaoyu Wang, Fengchao Zhao, Maji Sun, Xin Zheng, Hongyan Dong, and Kaijin Guo. 2016. "Pigment Epithelium-Derived Factor (PEDF) Protects Osteoblastic Cell Line from Glucocorticoid-Induced Apoptosis via PEDF-R" International Journal of Molecular Sciences 17, no. 5: 730. https://doi.org/10.3390/ijms17050730
APA StyleYao, S., Zhang, Y., Wang, X., Zhao, F., Sun, M., Zheng, X., Dong, H., & Guo, K. (2016). Pigment Epithelium-Derived Factor (PEDF) Protects Osteoblastic Cell Line from Glucocorticoid-Induced Apoptosis via PEDF-R. International Journal of Molecular Sciences, 17(5), 730. https://doi.org/10.3390/ijms17050730