
Function of Lipid Storage Droplet 1 (Lsd1) in Wing Development of Drosophila melanogaster





Abstract
:
1. Introduction
2. Results
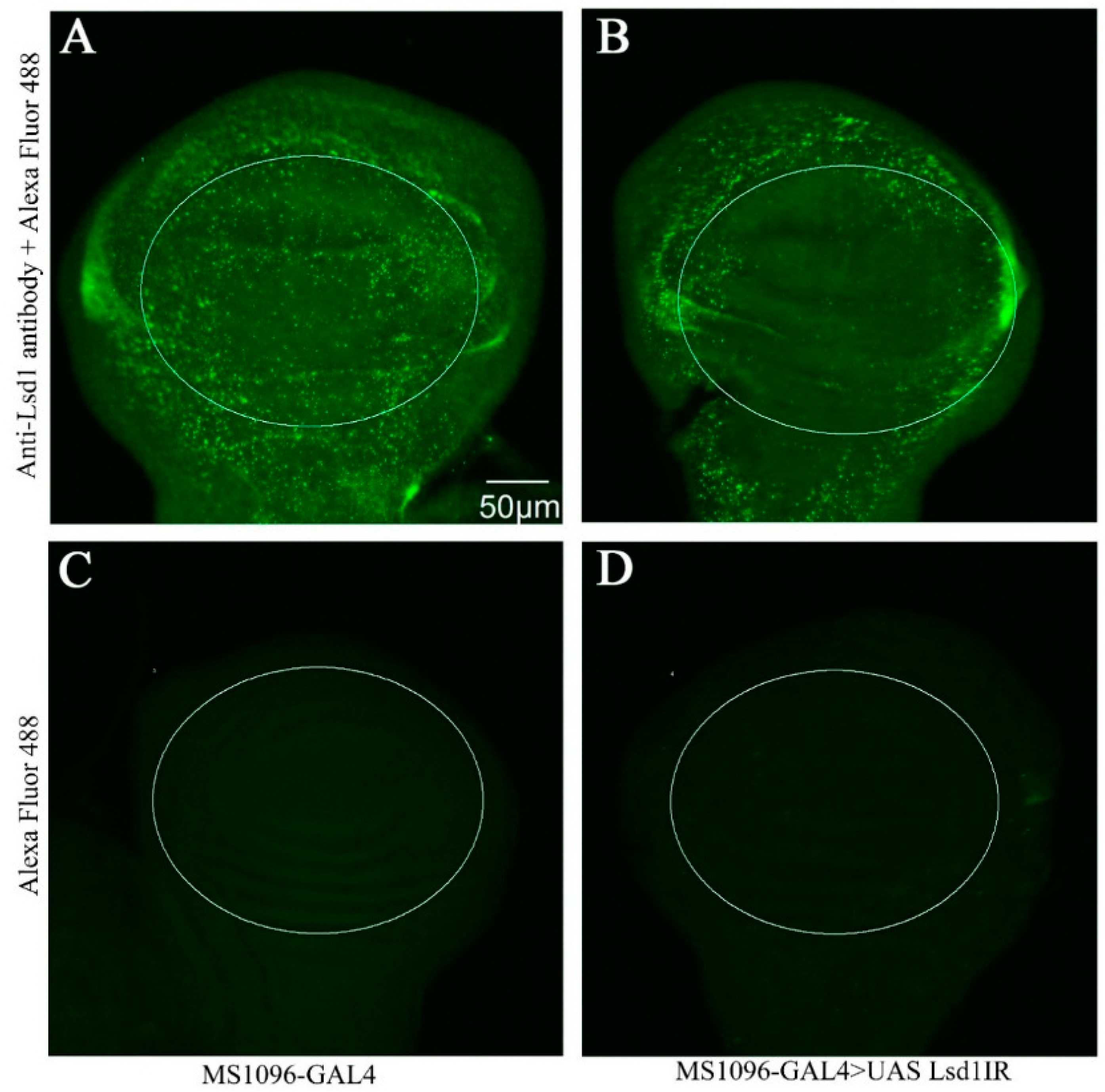
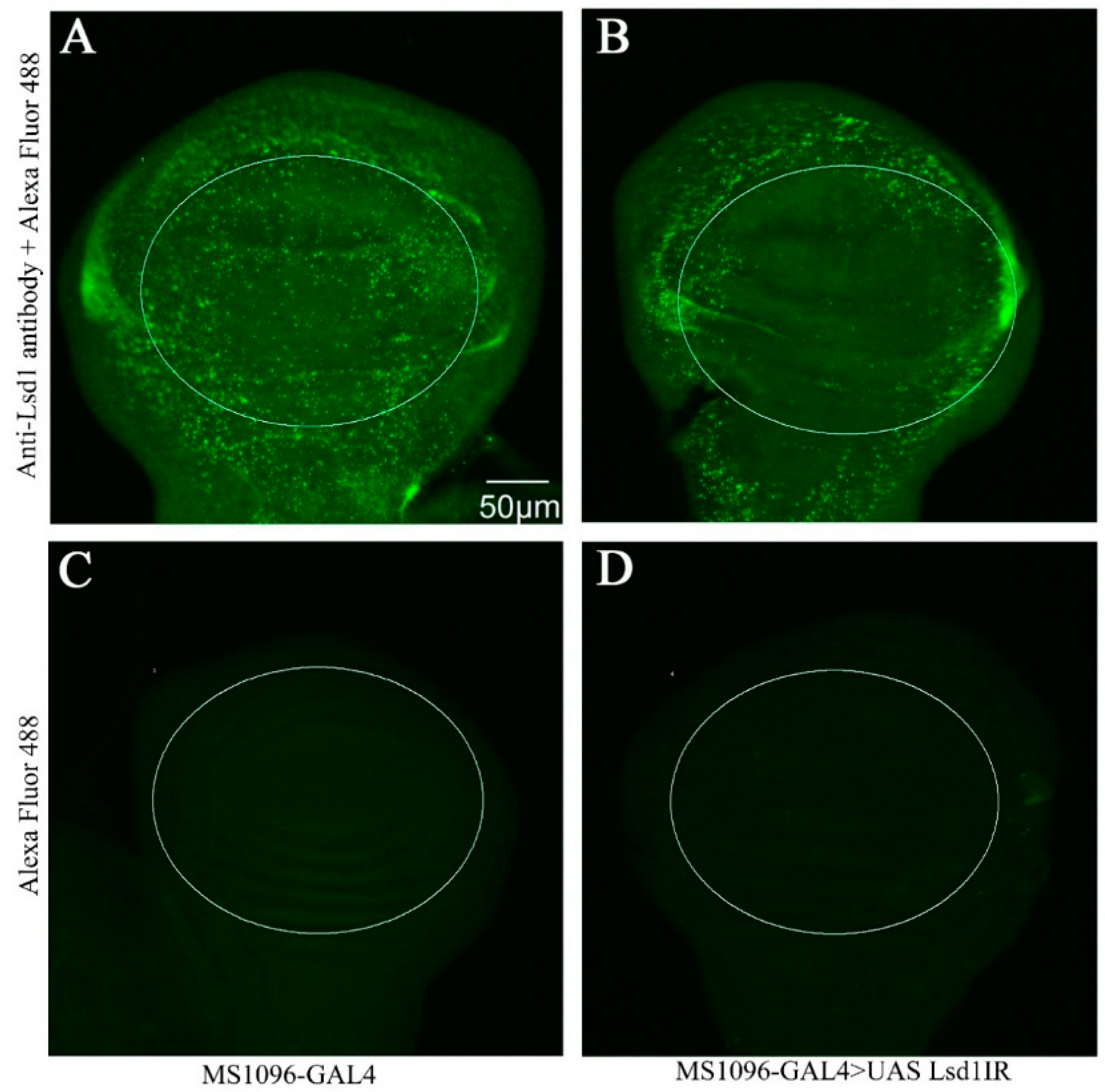
2.1. Effect of Lsd1 Knockdown in Various Tissues and Entire Drosophila
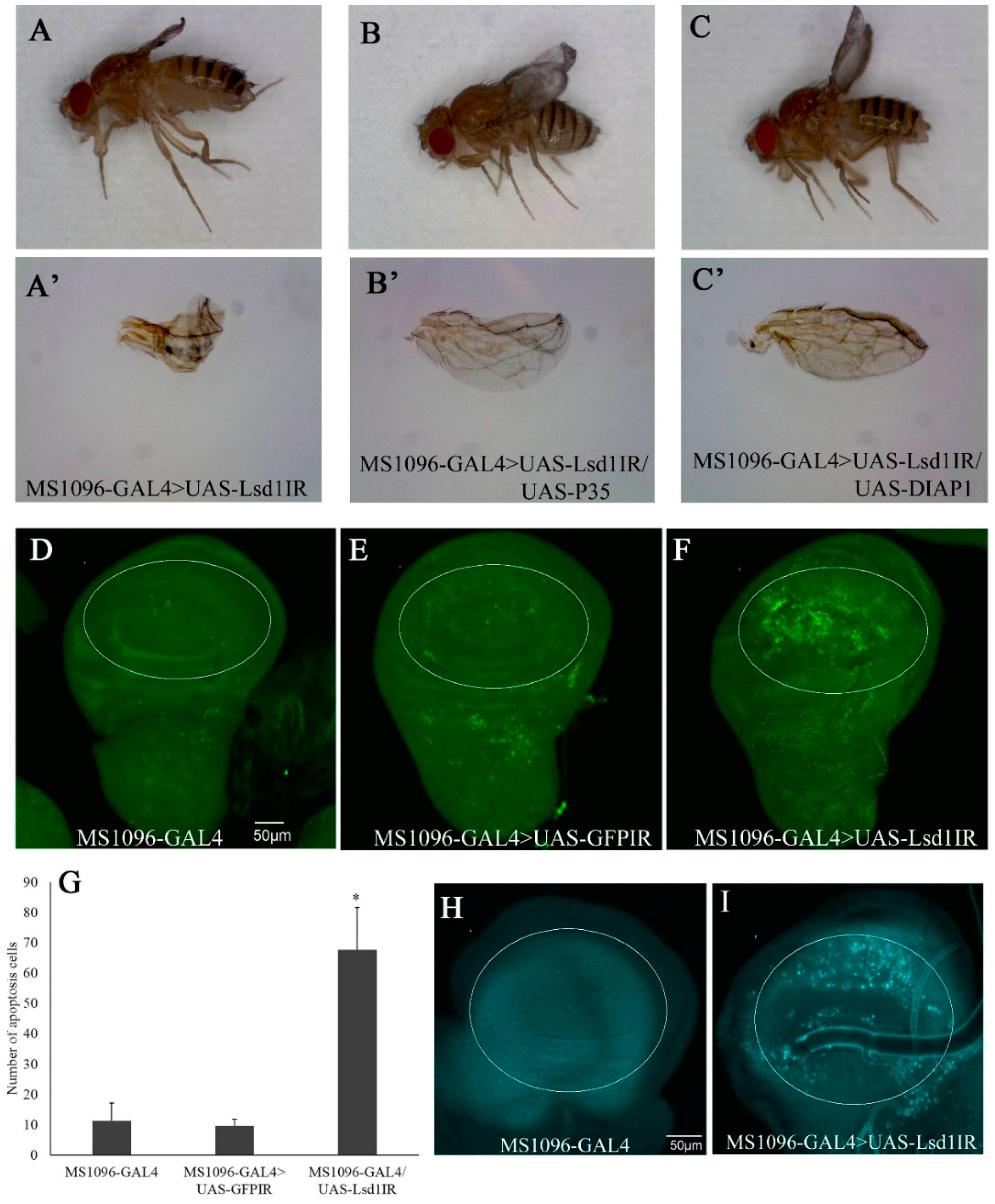
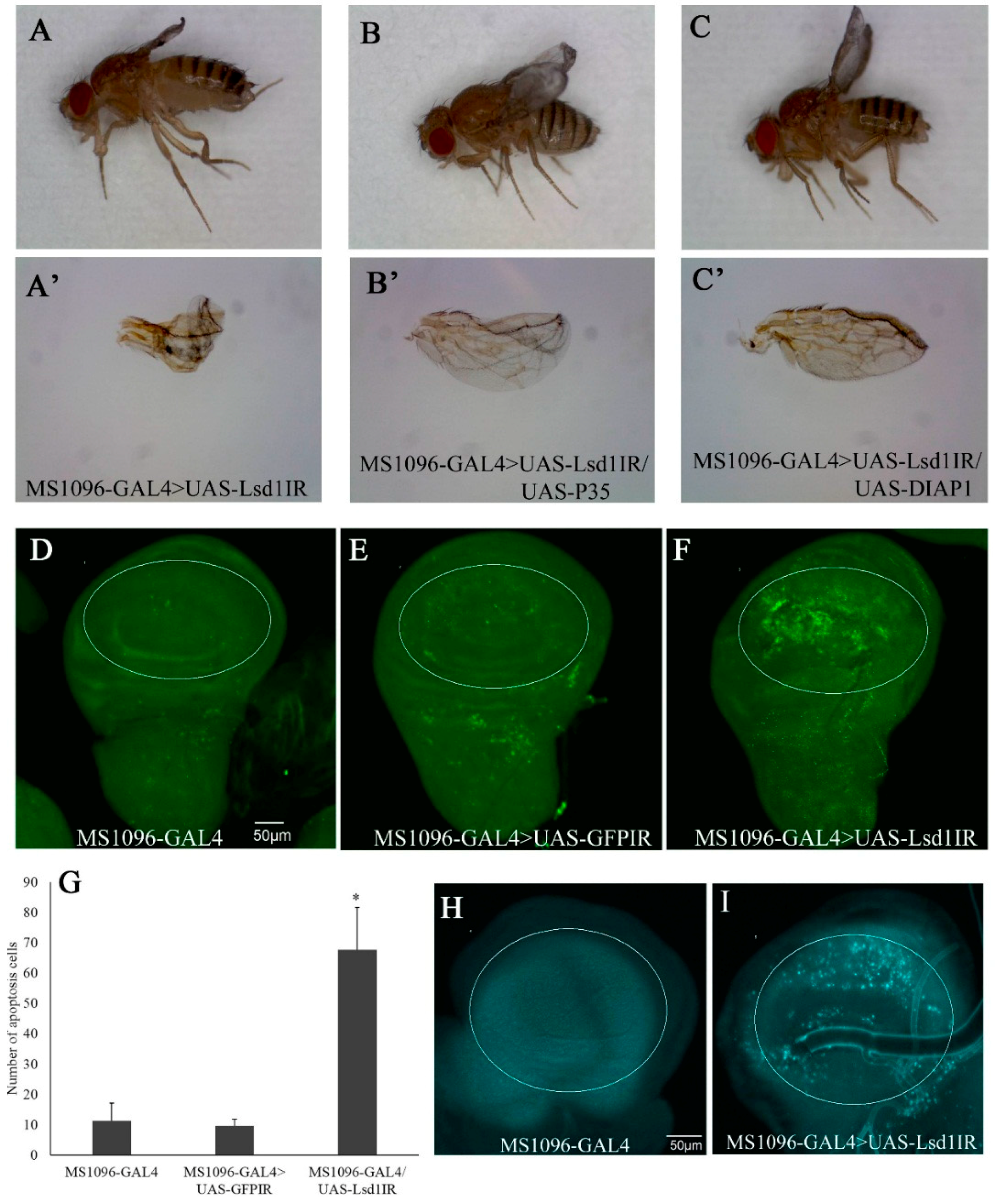
2.2. Knockdown of Lsd1 Disrupted Normal Wing Development
2.3. Knockdown of Lsd1 Led to Increased Cell Death
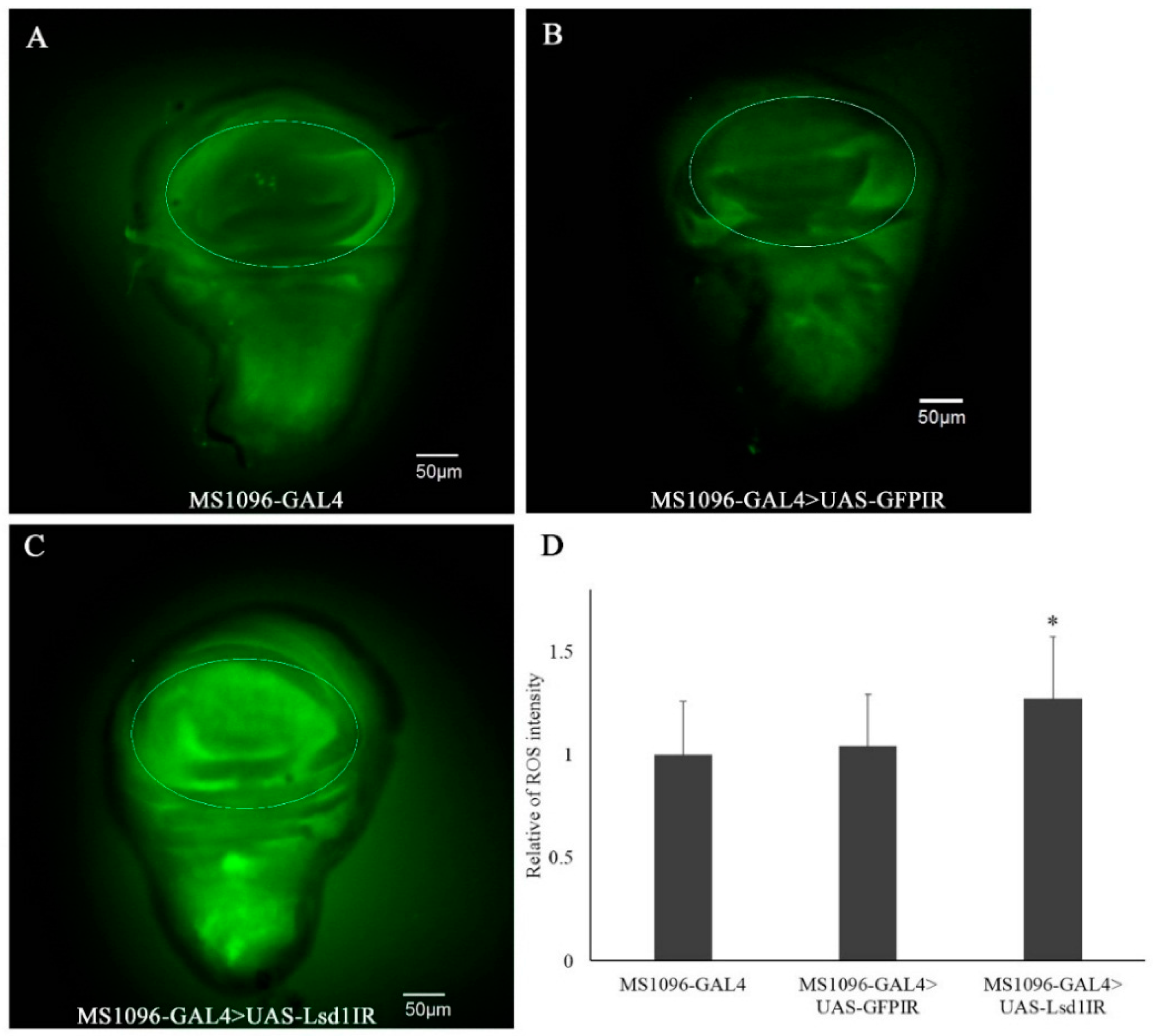
2.4. Knockdown of Lsd1 Increased ROS Generation
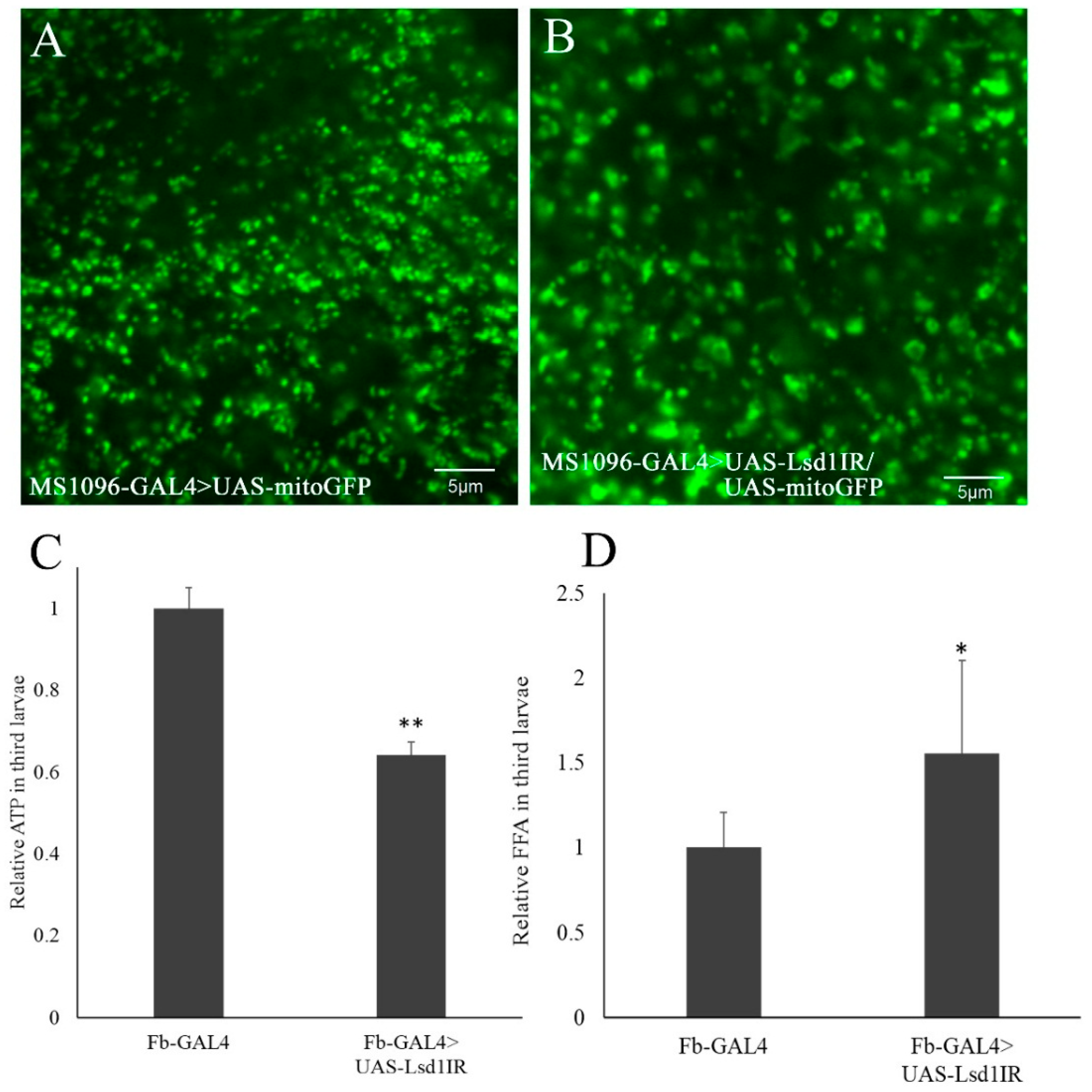
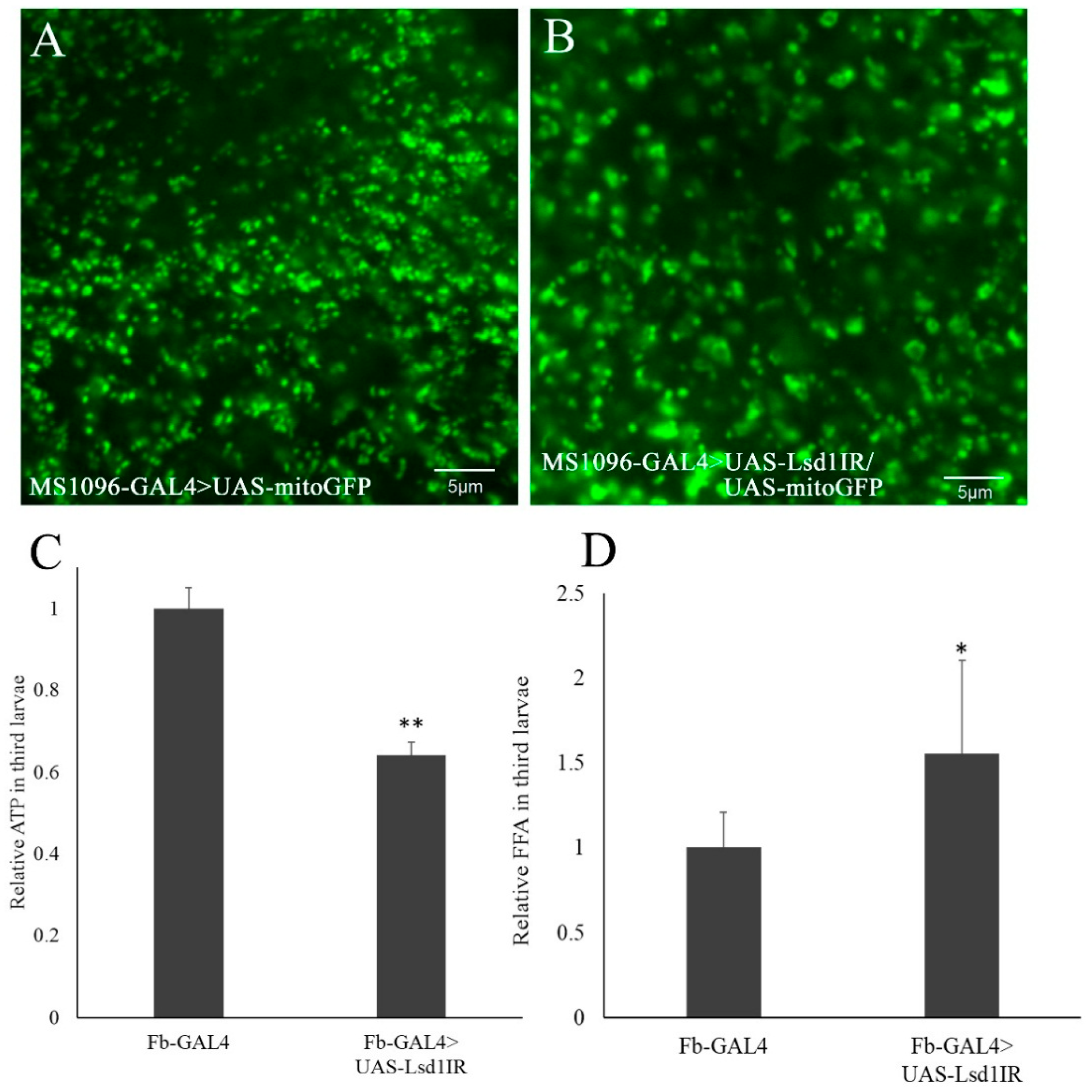
2.5. Knockdown of Lsd1 Caused Stress in Mitochondria and Defects in ATP Production
3. Discussion
4. Materials and Methods
4.1. Fly Stocks
4.2. Immunohistochemistry
4.3. In Vivo ROS Detection
4.4. Apoptotic Detection
4.5. Autophagy Assay
4.6. ATP Assay
4.7. FFA Level Measurement
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tansey, J.T.; Sztalryd, C.; Hlavin, E.M.; Kimmel, A.R.; Londos, C. The central role of perilipin a in lipid metabolism and adipocyte lipolysis. IUBMB Life 2004, 56, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Beller, M.; Bulankina, A.V.; Hsiao, H.H.; Urlaub, H.; Jackle, H.; Kuhnlein, R.P. PERILIPIN-dependent control of lipid droplet structure and fat storage in Drosophila. Cell Metab. 2010, 12, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, A.R.; Brasaemle, D.L.; McAndrews-Hill, M.; Sztalryd, C.; Londos, C. Adoption of PERILIPIN as a unifying nomenclature for the mammalian PAT-family of intracellular lipid storage droplet proteins. J. Lipid Res. 2010, 51, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Granneman, J.G.; Moore, H.P.; Krishnamoorthy, R.; Rathod, M. Perilipin controls lipolysis by regulating the interactions of AB-hydrolase containing 5 (Abhd5) and adipose triglyceride lipase (Atgl). J. Biol. Chem. 2009, 284, 34538–34544. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, H.; Souza, S.C.; Zhang, H.H.; Strissel, K.J.; Christoffolete, M.A.; Kovsan, J.; Rudich, A.; Kraemer, F.B.; Bianco, A.C.; Obin, M.S.; et al. Perilipin promotes hormone-sensitive lipase-mediated adipocyte lipolysis via phosphorylation-dependent and -independent mechanisms. J. Biol. Chem. 2006, 281, 15837–15844. [Google Scholar] [CrossRef] [PubMed]
- Sztalryd, C.; Xu, G.; Dorward, H.; Tansey, J.T.; Contreras, J.A.; Kimmel, A.R.; Londos, C. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. J. Cell Biol. 2003, 161, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, R.; Strauss, J.G.; Haemmerle, G.; Schoiswohl, G.; Birner-Gruenberger, R.; Riederer, M.; Lass, A.; Neuberger, G.; Eisenhaber, F.; Hermetter, A.; et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science 2004, 306, 1383–1386. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Xiang, Y.; Chen, H.; Liu, Z.; Gronke, S.; Kuhnlein, R.P.; Huang, X. Opposite and redundant roles of the two Drosophila perilipins in lipid mobilization. J. Cell Sci. 2012, 125, 3568–3577. [Google Scholar] [CrossRef] [PubMed]
- Bickel, P.E.; Tansey, J.T.; Welte, M.A. PAT proteins, an ancient family of lipid droplet proteins that regulate cellular lipid stores. Biochim. Biophys. Acta 2009, 1791, 419–440. [Google Scholar] [CrossRef] [PubMed]
- Gandotra, S.; le Dour, C.; Bottomley, W.; Cervera, P.; Giral, P.; Reznik, Y.; Charpentier, G.; Auclair, M.; Delepine, M.; Barroso, I.; et al. Perilipin deficiency and autosomal dominant partial lipodystrophy. N. Engl. J. Med. 2011, 364, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.D.; Thummel, C.S. Diabetic larvae and obese flies-emerging studies of metabolism in Drosophila. Cell Metab. 2007, 6, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, A.; Stainier, D.Y. Lessons from “lower” organisms: What worms, flies, and zebrafish can teach us about human energy metabolism. PLoS Genet. 2007, 3, e199. [Google Scholar] [CrossRef] [PubMed]
- Miura, S.; Gan, J.W.; Brzostowski, J.; Parisi, M.J.; Schultz, C.J.; Londos, C.; Oliver, B.; Kimmel, A.R. Functional conservation for lipid storage droplet association among Perilipin, ADRP, and TIP47 (PAT)-related proteins in mammals, Drosophila, and Dictyostelium. J. Biol. Chem. 2002, 277, 32253–32257. [Google Scholar] [CrossRef] [PubMed]
- Arrese, E.L.; Rivera, L.; Hamada, M.; Mirza, S.; Hartson, S.D.; Weintraub, S.; Soulages, J.L. Function and structure of lipid storage droplet protein 1 studied in lipoprotein complexes. Arch. Biochem. Biophys. 2008, 473, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Phelps, C.B.; Brand, A.H. Ectopic gene expression in Drosophila using GAL4 system. Methods 1998, 14, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Crespo, M.; Pascual, M.; Fernandez-Costa, J.M.; Garcia-Lopez, A.; Monferrer, L.; Miranda, M.E.; Zhou, L.; Artero, R.D. Drosophila muscleblind is involved in troponin T alternative splicing and apoptosis. PLoS ONE 2008, 3, e1613. [Google Scholar] [CrossRef] [PubMed]
- Tseng, A.S.; Hariharan, I.K. An overexpression screen in Drosophila for genes that restrict growth or cell-cycle progression in the developing eye. Genetics 2002, 162, 229–243. [Google Scholar] [PubMed]
- Wang, X.; Ma, Y.; Zhao, Y.; Chen, Y.; Hu, Y.; Chen, C.; Shao, Y.; Xue, L. APLP1 promotes dFoxO-dependent cell death in Drosophila. Apoptosis Int. J. Program. Cell Death 2015, 20, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.G.; Tan, S.X.; Dawes, I.W. Reactive oxygen species and yeast apoptosis. Biochim. Biophys. Acta 2008, 1783, 1354–1368. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Yang, Y.; Ming, M.; Liu, B. Mitochondrial ROS generation for regulation of autophagic pathways in cancer. Biochem. Biophys. Res. Commun. 2011, 414, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkov, A.A. Mitochondrial metabolism of reactive oxygen species. Biochem. Biokhimiia 2005, 70, 200–214. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Phys. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D. The sites and topology of mitochondrial superoxide production. Exp. Gerontol. 2010, 45, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Ratnaparkhi, A. Signaling by Folded gastrulation is modulated by mitochondrial fusion and fission. J. Cell Sci. 2013, 126, 5369–5376. [Google Scholar] [CrossRef] [PubMed]
- De la Cova, C.; Senoo-Matsuda, N.; Ziosi, M.; Wu, D.C.; Bellosta, P.; Quinzii, C.M.; Johnston, L.A. Supercompetitor status of Drosophila Myc cells requires p53 as a fitness sensor to reprogram metabolism and promote viability. Cell Metab. 2014, 19, 470–483. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Li, H.; Zhang, K.; Jian, F.; Tang, J.; Song, Z. Loss of Yme1L perturbates mitochondrial dynamics. Cell Death Dis. 2013, 4, e896. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H. Mitochondrial disease. Lancet 2006, 368, 70–82. [Google Scholar] [CrossRef]
- Griffiths, E.J.; Rutter, G.A. Mitochondrial calcium as a key regulator of mitochondrial ATP production in mammalian cells. Biochim. Biophys. Acta 2009, 1787, 1324–1333. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, E.; Wiggins, D.; Fielding, B.; Gould, A.P. Specialized hepatocyte-like cells regulate Drosophila lipid metabolism. Nature 2007, 445, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Bugger, H.; Abel, E.D. Molecular mechanisms for myocardial mitochondrial dysfunction in the metabolic syndrome. Clin. Sci. (Lond.) 2008, 114, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Kuhnlein, R.P. Thematic review series: Lipid droplet synthesis and metabolism: From yeast to man. Lipid droplet-based storage fat metabolism in Drosophila. J. Lipid Res. 2012, 53, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- Pierce, G.B.; Parchment, R.E.; Lewellyn, A.L. Hydrogen peroxide as a mediator of programmed cell death in the blastocyst. Differ. Res. Biol. Divers. 1991, 46, 181–186. [Google Scholar] [CrossRef]
- Kasahara, Y.; Iwai, K.; Yachie, A.; Ohta, K.; Konno, A.; Seki, H.; Miyawaki, T.; Taniguchi, N. Involvement of reactive oxygen intermediates in spontaneous and CD95 (Fas/APO-1)-mediated apoptosis of neutrophils. Blood 1997, 89, 1748–1753. [Google Scholar] [PubMed]
- Gruber, A.; Cornaciu, I.; Lass, A.; Schweiger, M.; Poeschl, M.; Eder, C.; Kumari, M.; Schoiswohl, G.; Wolinski, H.; Kohlwein, S.D.; et al. The N-terminal region of comparative gene identification-58 (CGI-58) is important for lipid droplet binding and activation of adipose triglyceride lipase. J. Biol. Chem. 2010, 285, 12289–12298. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.J.; Kloppenburg, M.; Toes, R.E.; Ioan-Facsinay, A. Fatty acids, lipid mediators, and T-cell function. Front. Immunol. 2014, 5, 483. [Google Scholar] [CrossRef] [PubMed]
- Moreira, P.I.; Carvalho, C.; Zhu, X.; Smith, M.A.; Perry, G. Mitochondrial dysfunction is a trigger of Alzheimer’s disease pathophysiology. Biochim. Biophys. Acta 2010, 1802, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Gronke, S.; Beller, M.; Fellert, S.; Ramakrishnan, H.; Jackle, H.; Kuhnlein, R.P. Control of fat storage by a Drosophila PAT domain protein. Curr. Biol. 2003, 13, 603–606. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GAL4 Line | Expression Pattern | Phenotype with UAS-Lsd1IR |
|---|---|---|
| MS1096 | Dorsal wing disc | Atrophied wing at 25 and 28 °C |
| En | Posterior wing disc and embryo | Lethal at 25 °C |
| GMR | Eye disc | No detectable phenotype |
| Fb | Fat body | Delay in growth at 25 °C, lethal at 28 °C |
| Act5C | All tissues | Lethal at 25 °C |
| Tubp | All tissues | Lethal at 25 °C |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Men, T.T.; Binh, T.D.; Yamaguchi, M.; Huy, N.T.; Kamei, K. Function of Lipid Storage Droplet 1 (Lsd1) in Wing Development of Drosophila melanogaster. Int. J. Mol. Sci. 2016, 17, 648. https://doi.org/10.3390/ijms17050648
Men TT, Binh TD, Yamaguchi M, Huy NT, Kamei K. Function of Lipid Storage Droplet 1 (Lsd1) in Wing Development of Drosophila melanogaster. International Journal of Molecular Sciences. 2016; 17(5):648. https://doi.org/10.3390/ijms17050648
Chicago/Turabian StyleMen, Tran Thanh, Tran Duy Binh, Masamitsu Yamaguchi, Nguyen Tien Huy, and Kaeko Kamei. 2016. "Function of Lipid Storage Droplet 1 (Lsd1) in Wing Development of Drosophila melanogaster" International Journal of Molecular Sciences 17, no. 5: 648. https://doi.org/10.3390/ijms17050648
APA StyleMen, T. T., Binh, T. D., Yamaguchi, M., Huy, N. T., & Kamei, K. (2016). Function of Lipid Storage Droplet 1 (Lsd1) in Wing Development of Drosophila melanogaster. International Journal of Molecular Sciences, 17(5), 648. https://doi.org/10.3390/ijms17050648