
Induction of Lipocalin2 in a Rat Model of Lung Irradiation

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Serum Lipocalin2 (LCN2) Concentration after Lung Irradiation
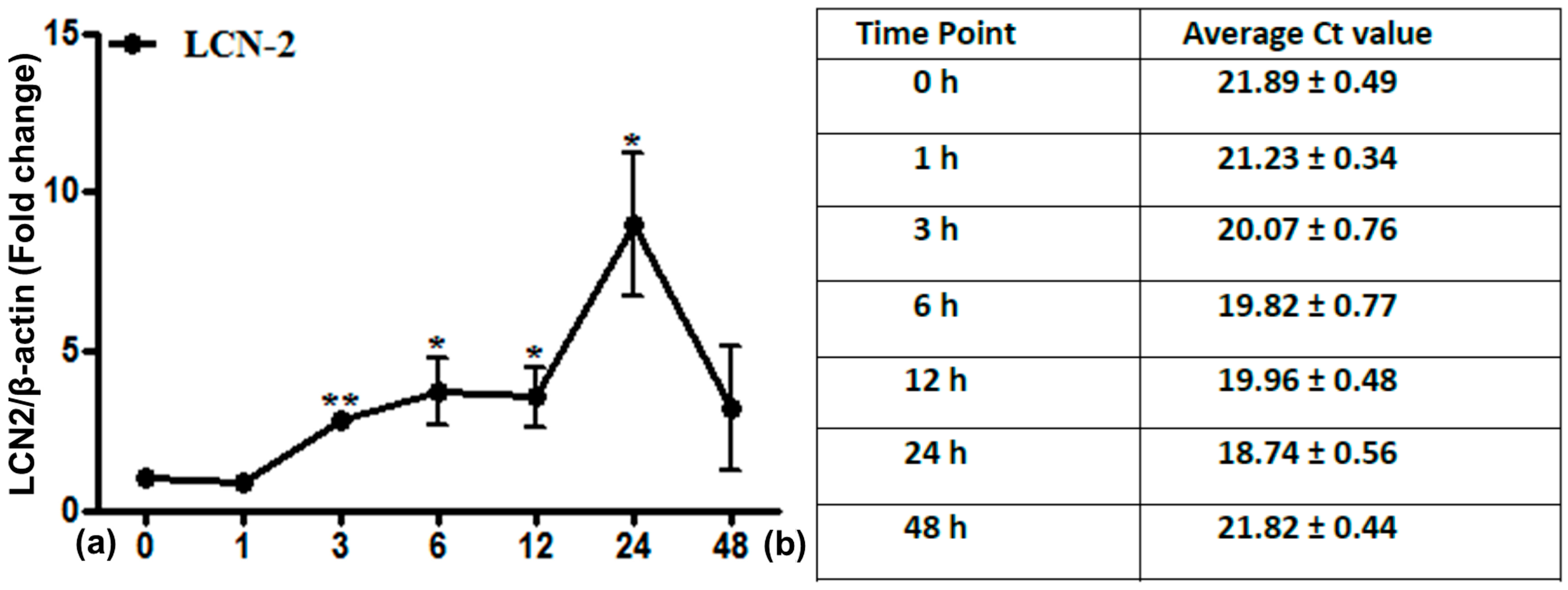
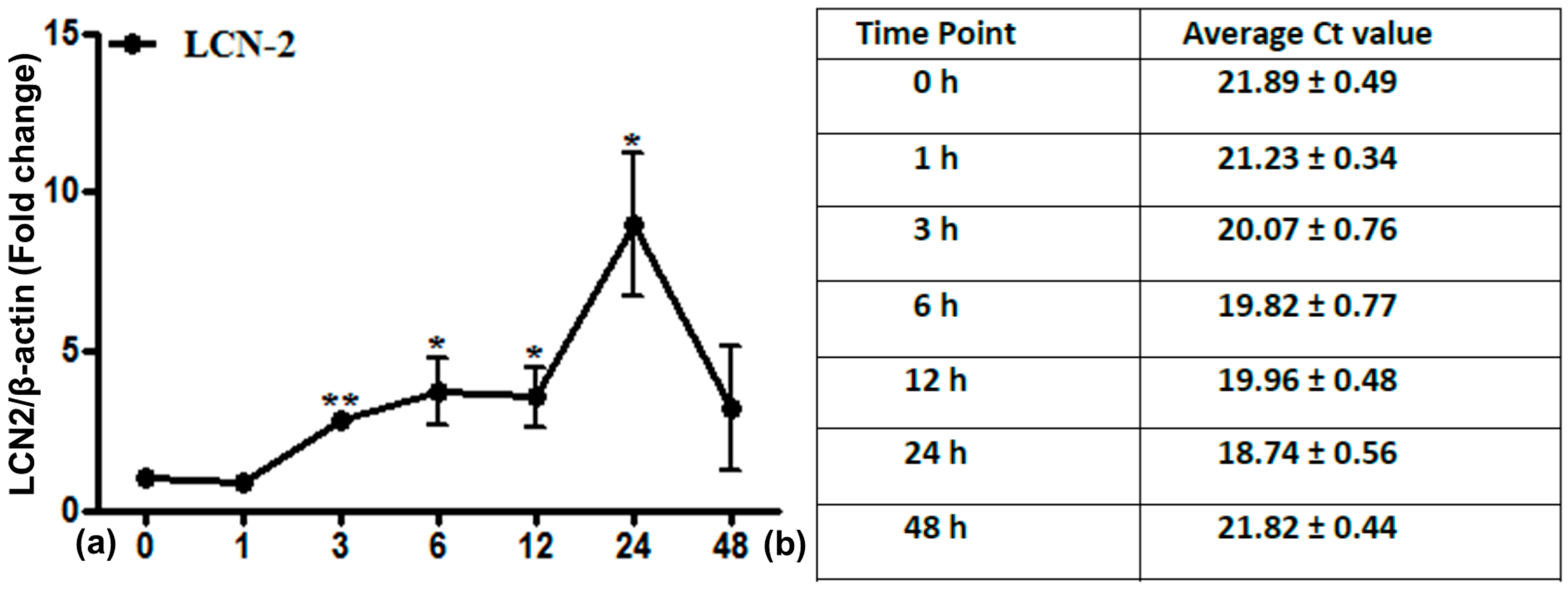
2.2. LCN2 Transcript Expression in Lung Tissue
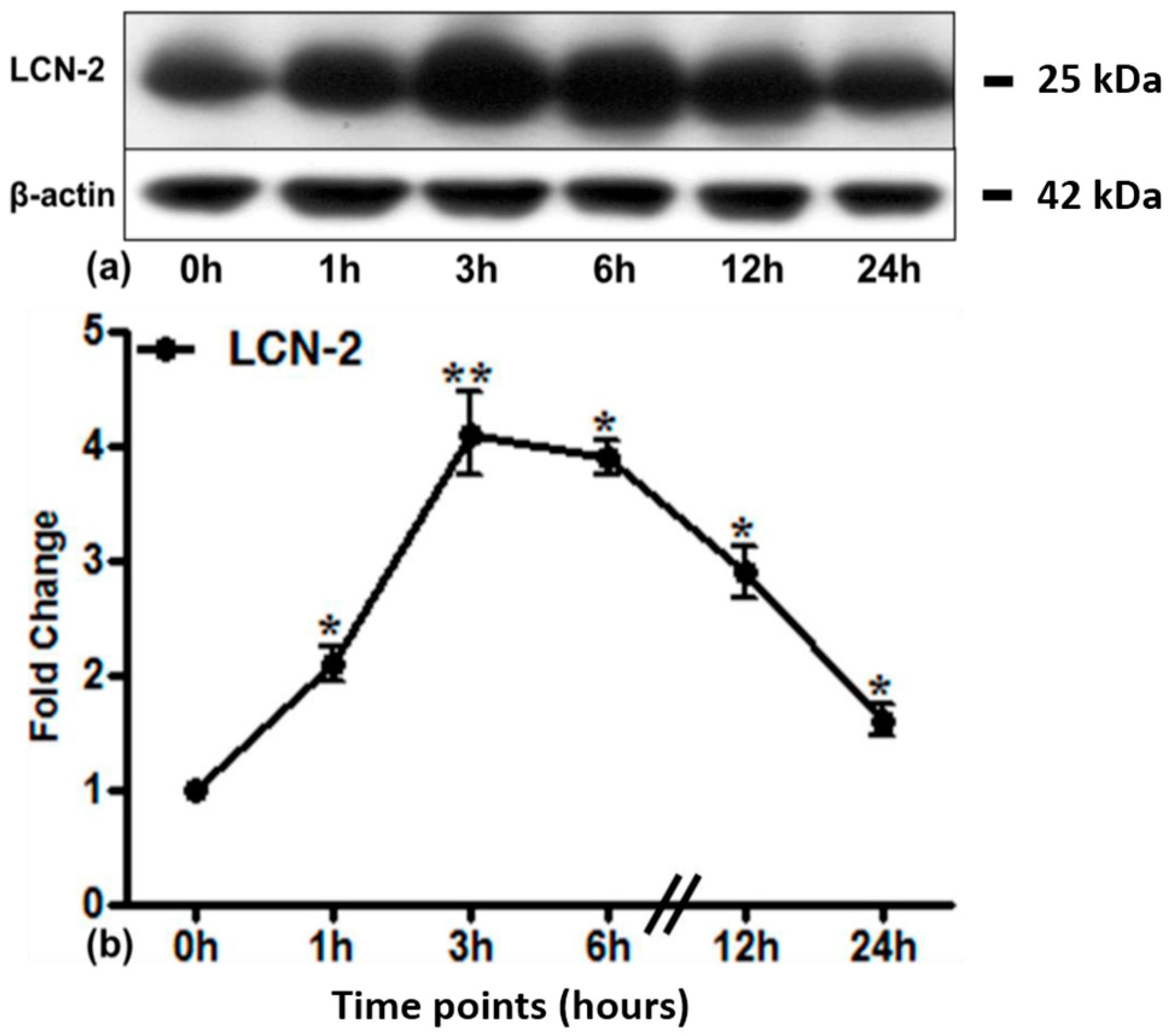
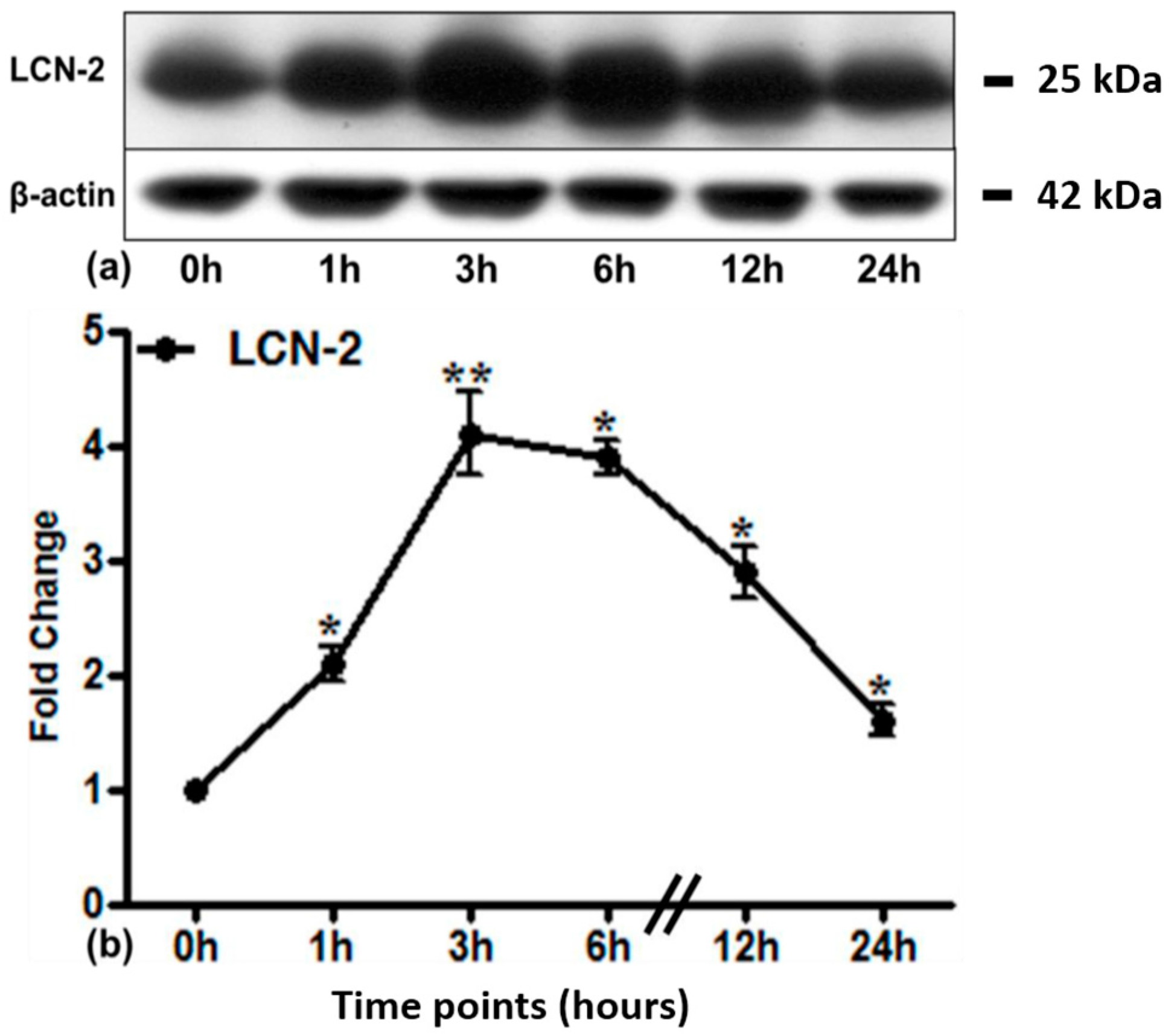
2.3. LCN2 Protein Expression in Lung Tissue
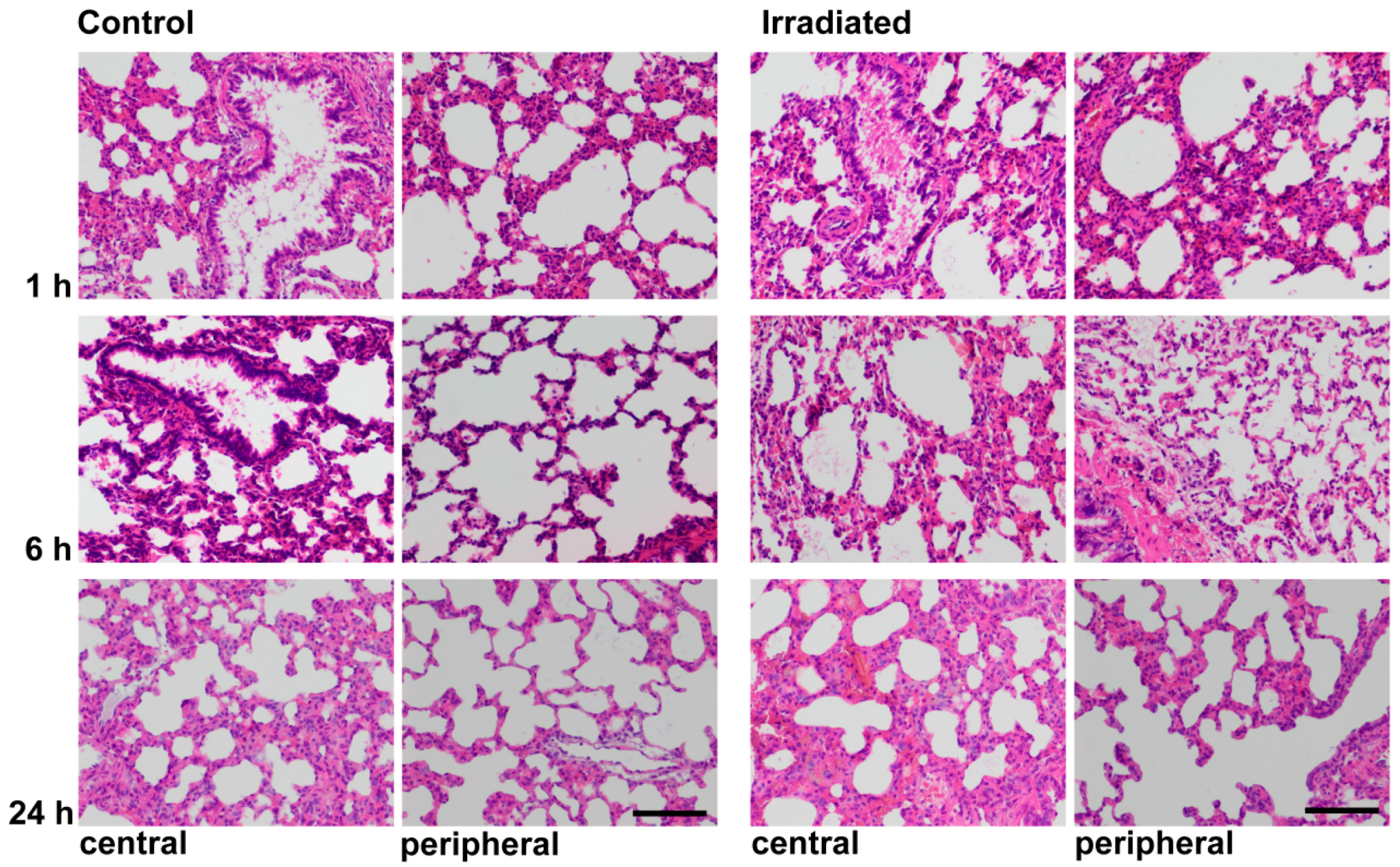
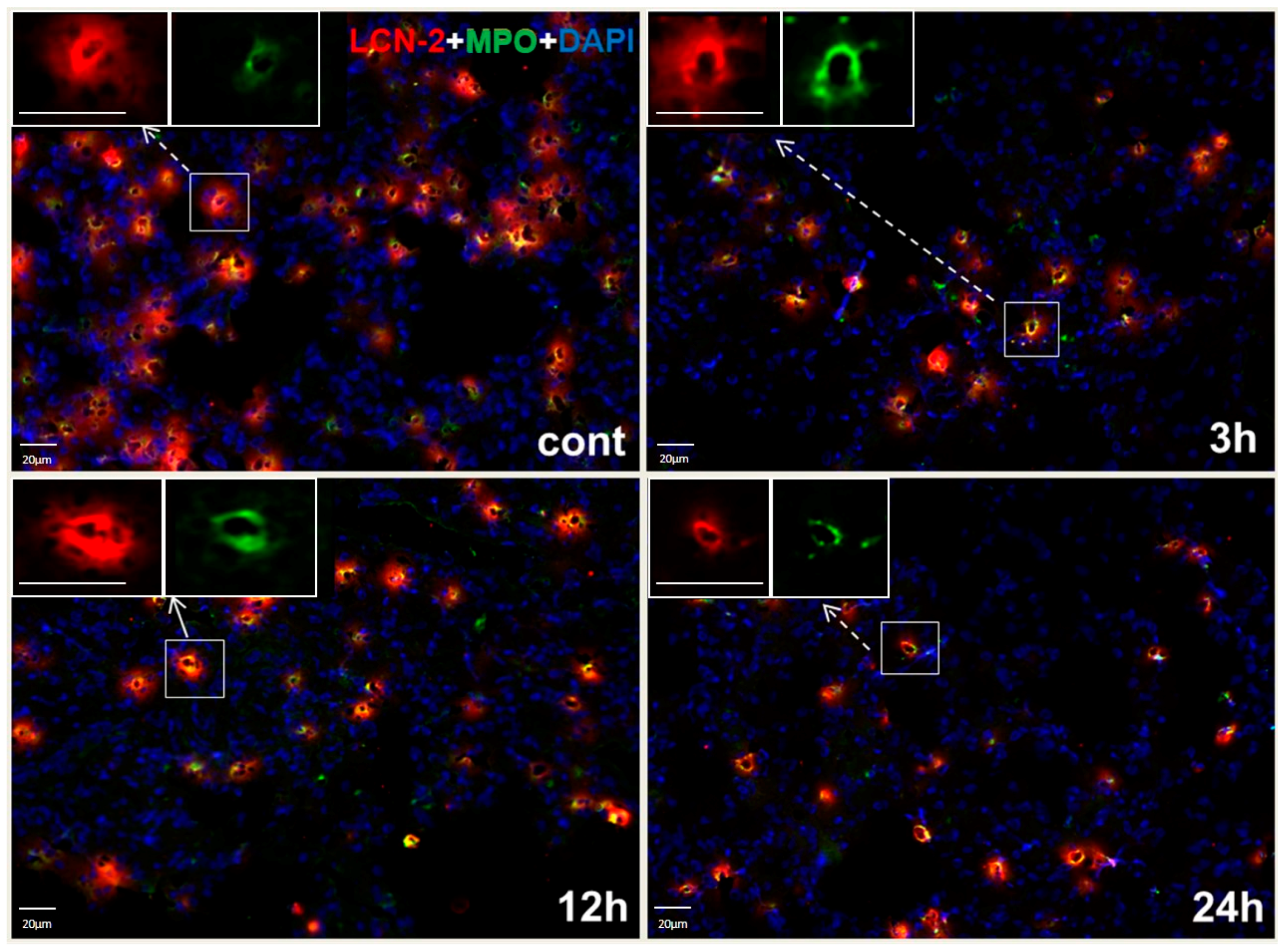
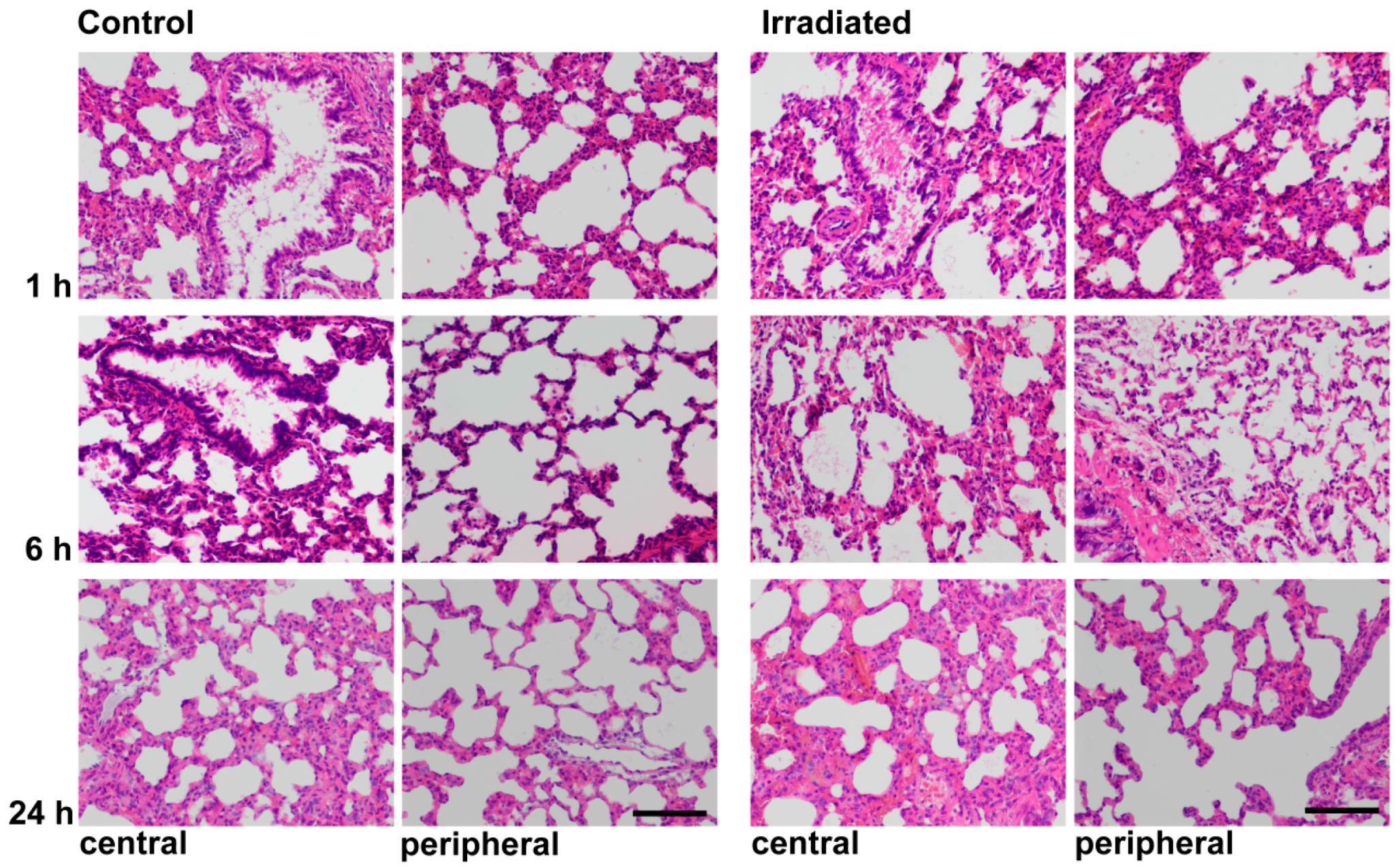
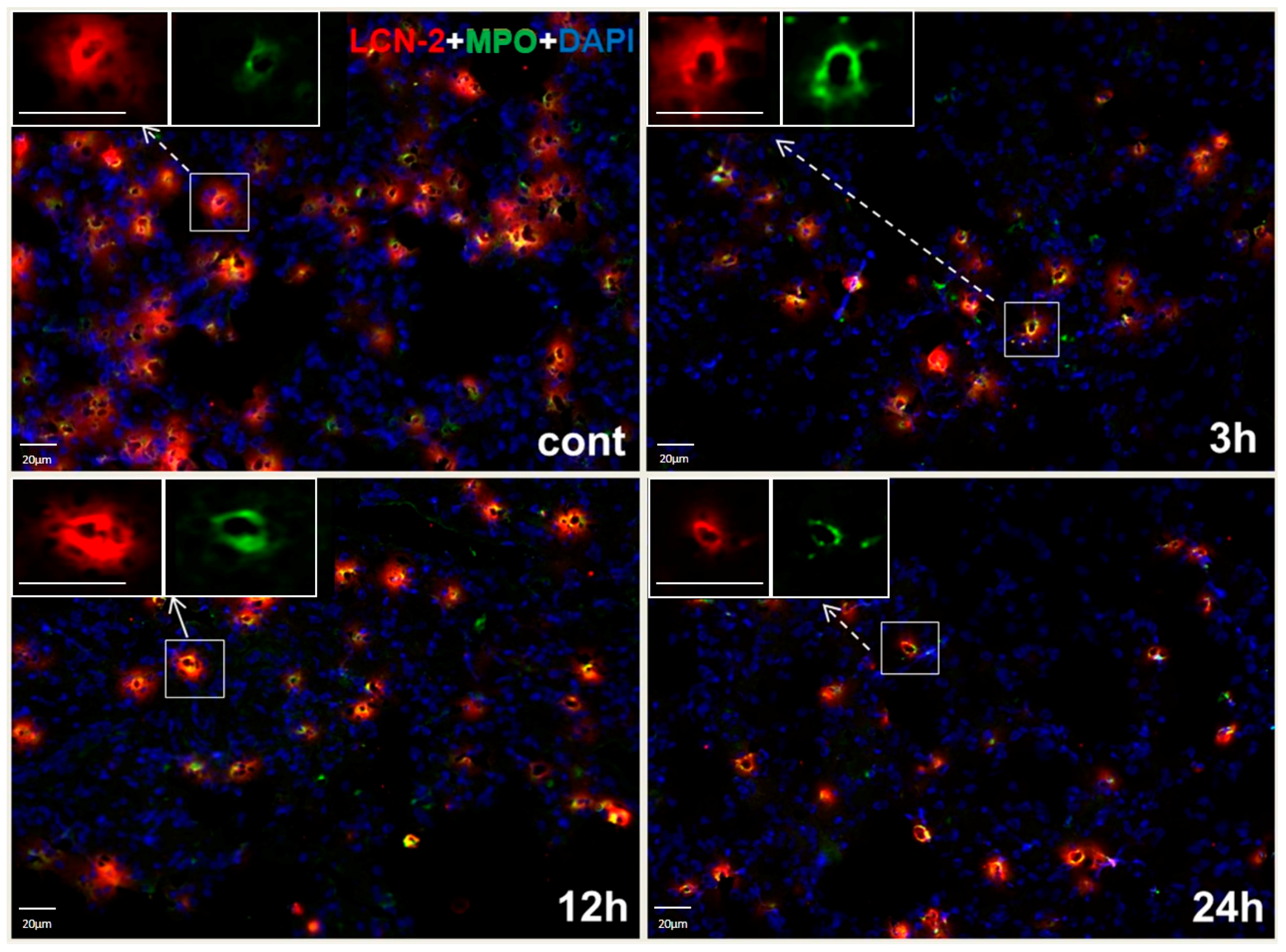
2.4. Histochemistry and LCN2 Immunofluorescence Staining in Sham and Irradiated Lung Tissue
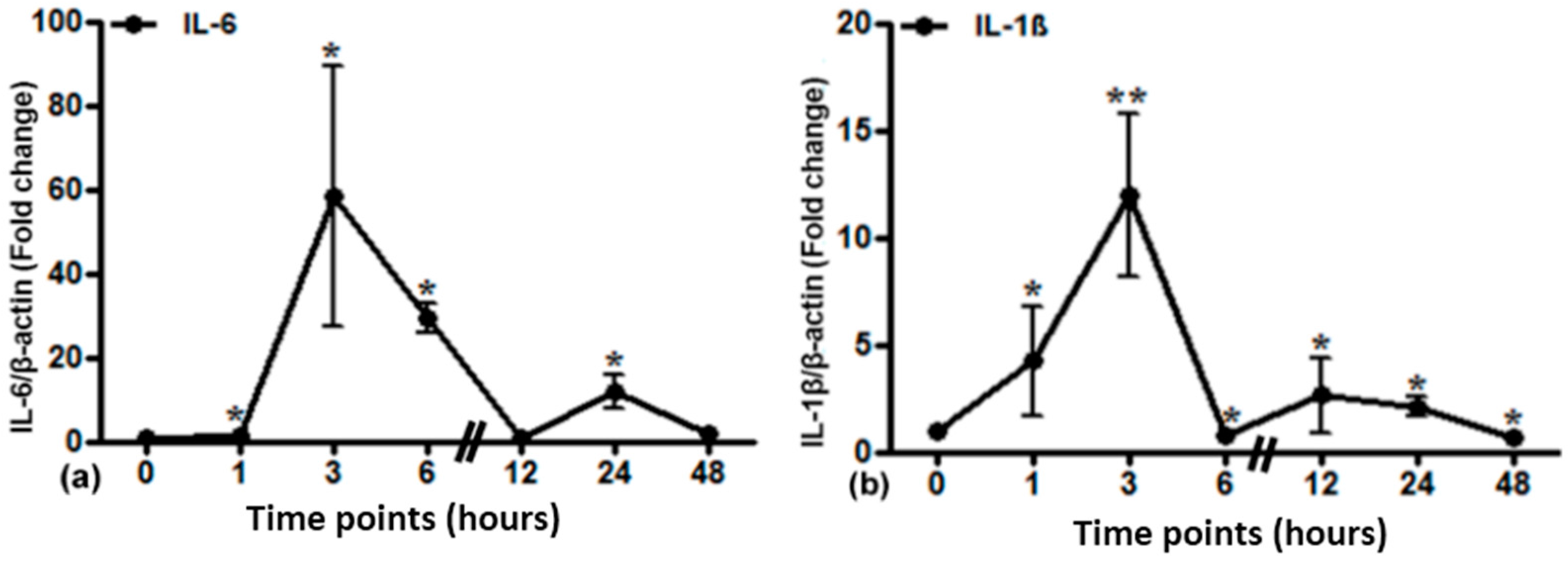
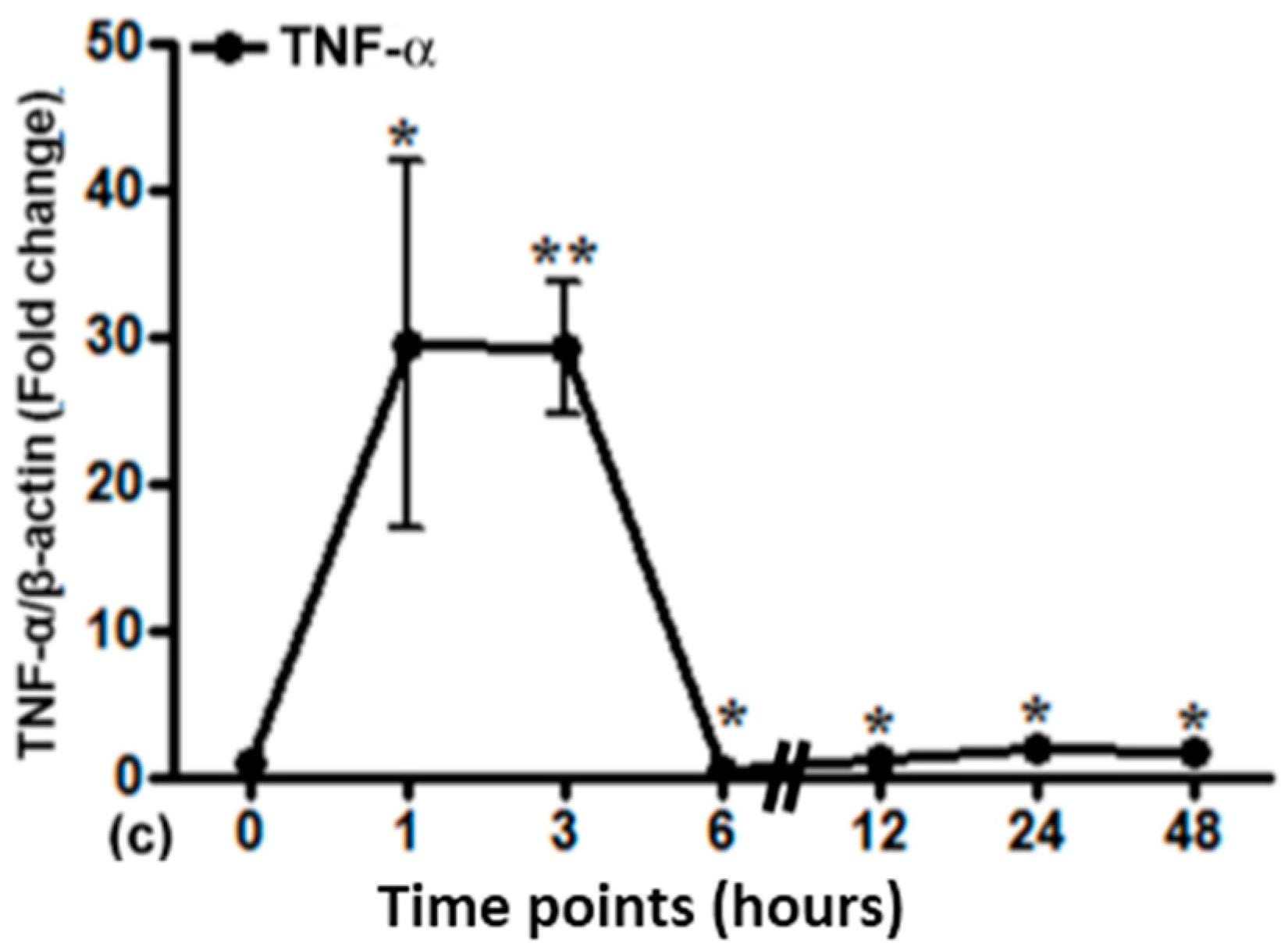
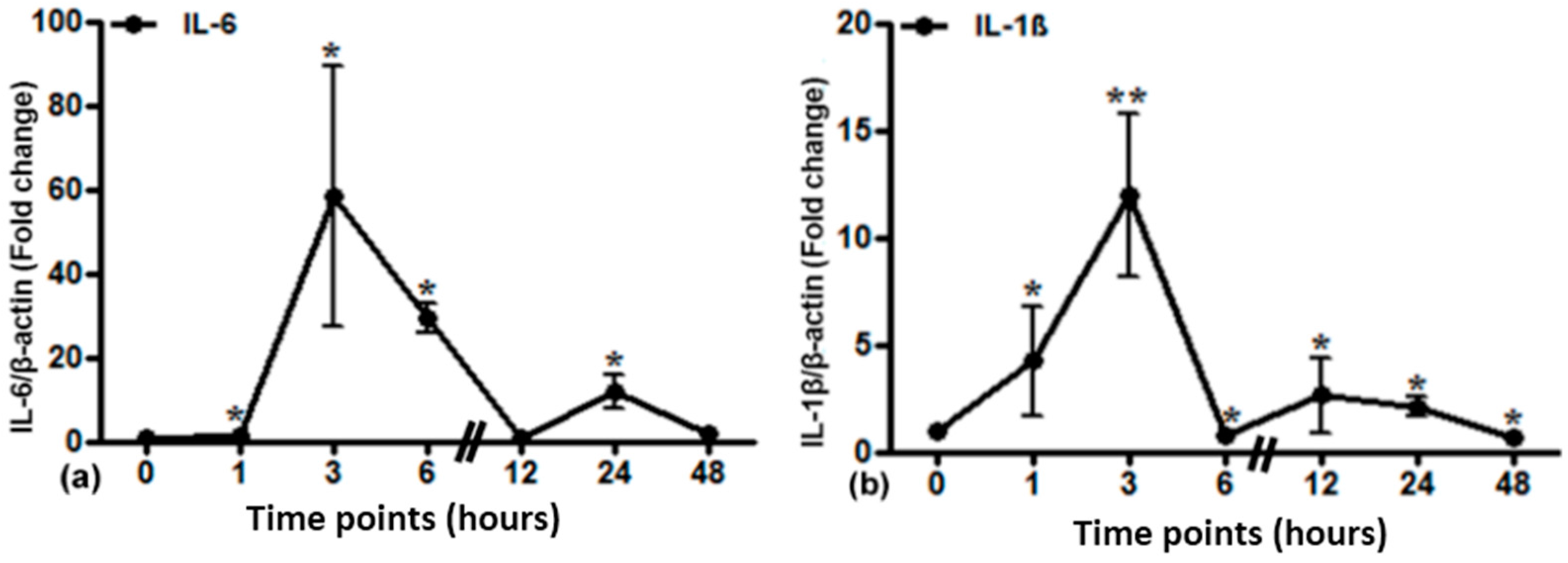
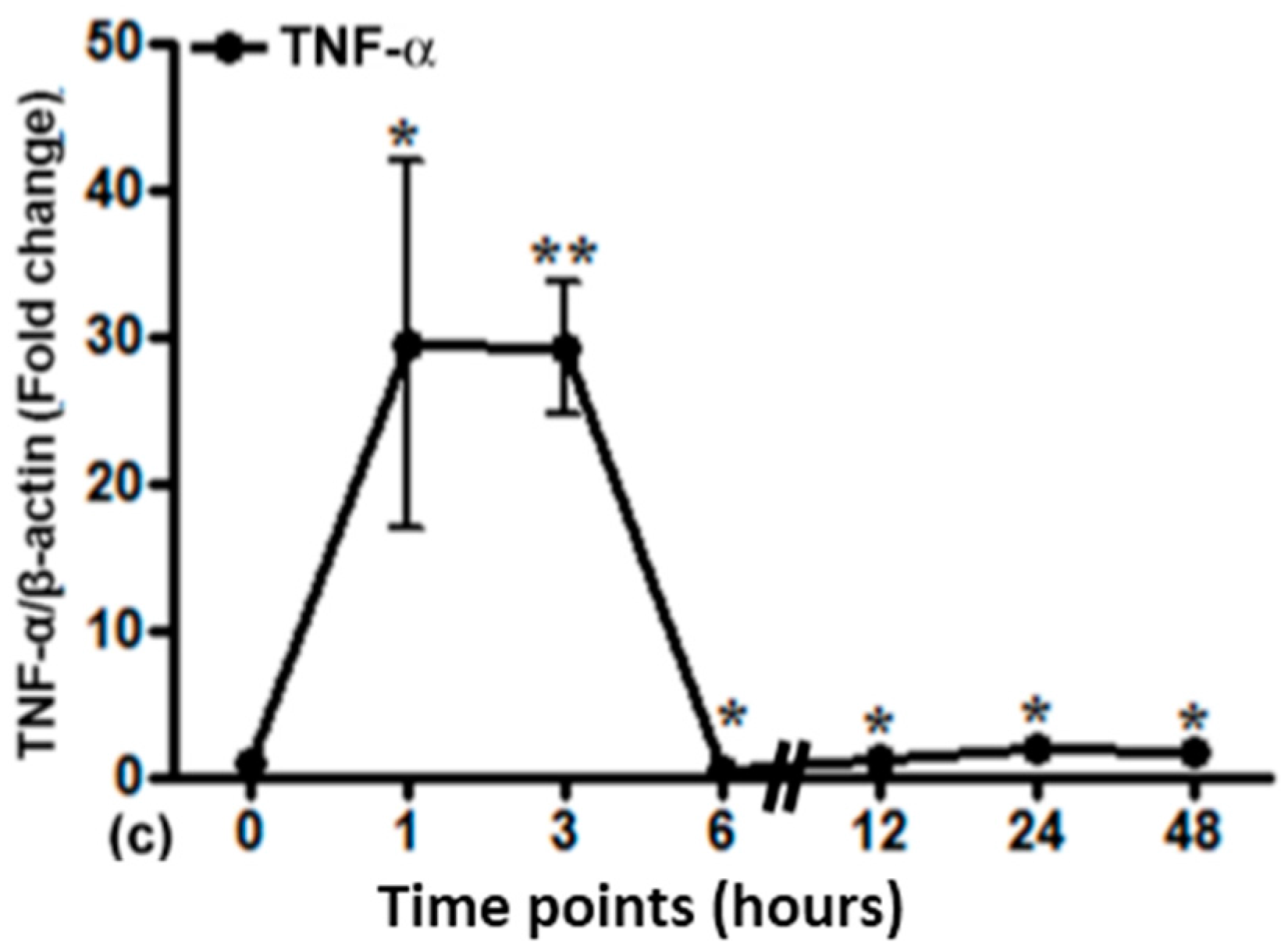
2.5. Gene Expression of Different Acute Phase Cytokines in Sham and Irradiated Lung Tissue
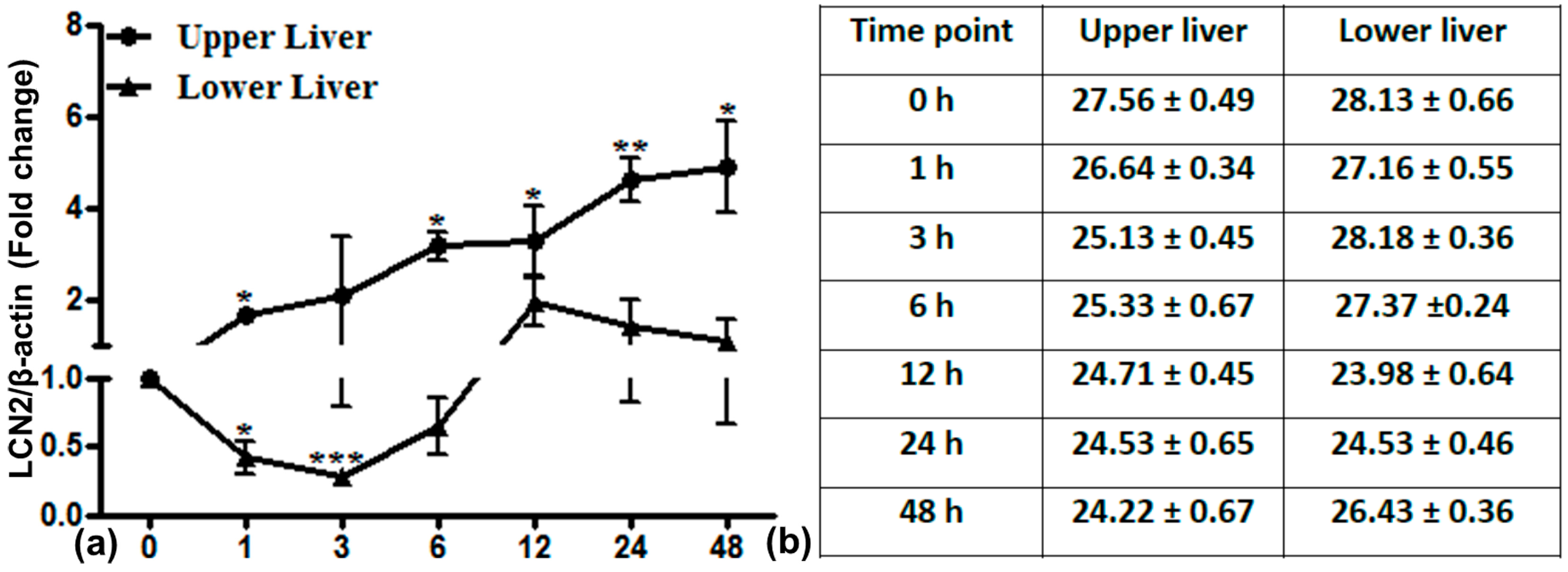
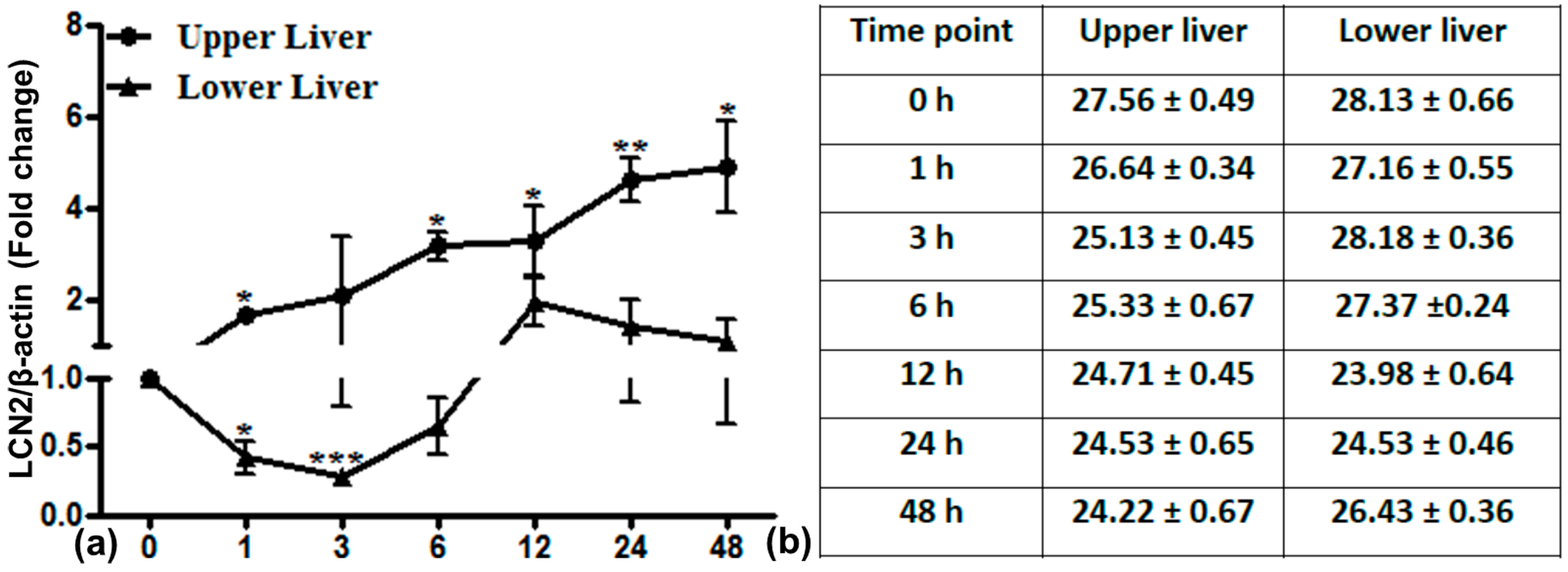
2.6. LCN2 Transcript Expression in the Upper and Lower Part of the Liver
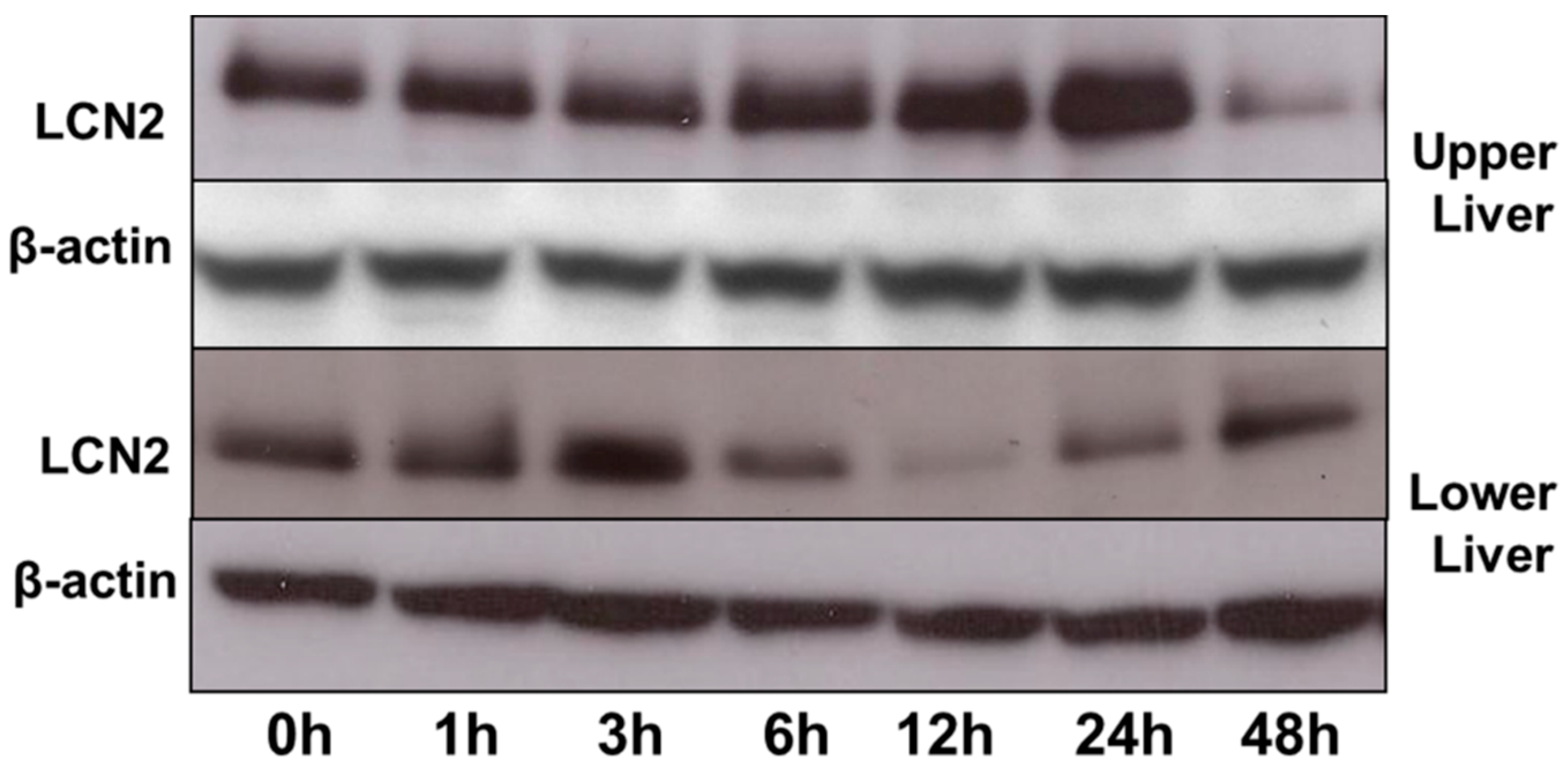
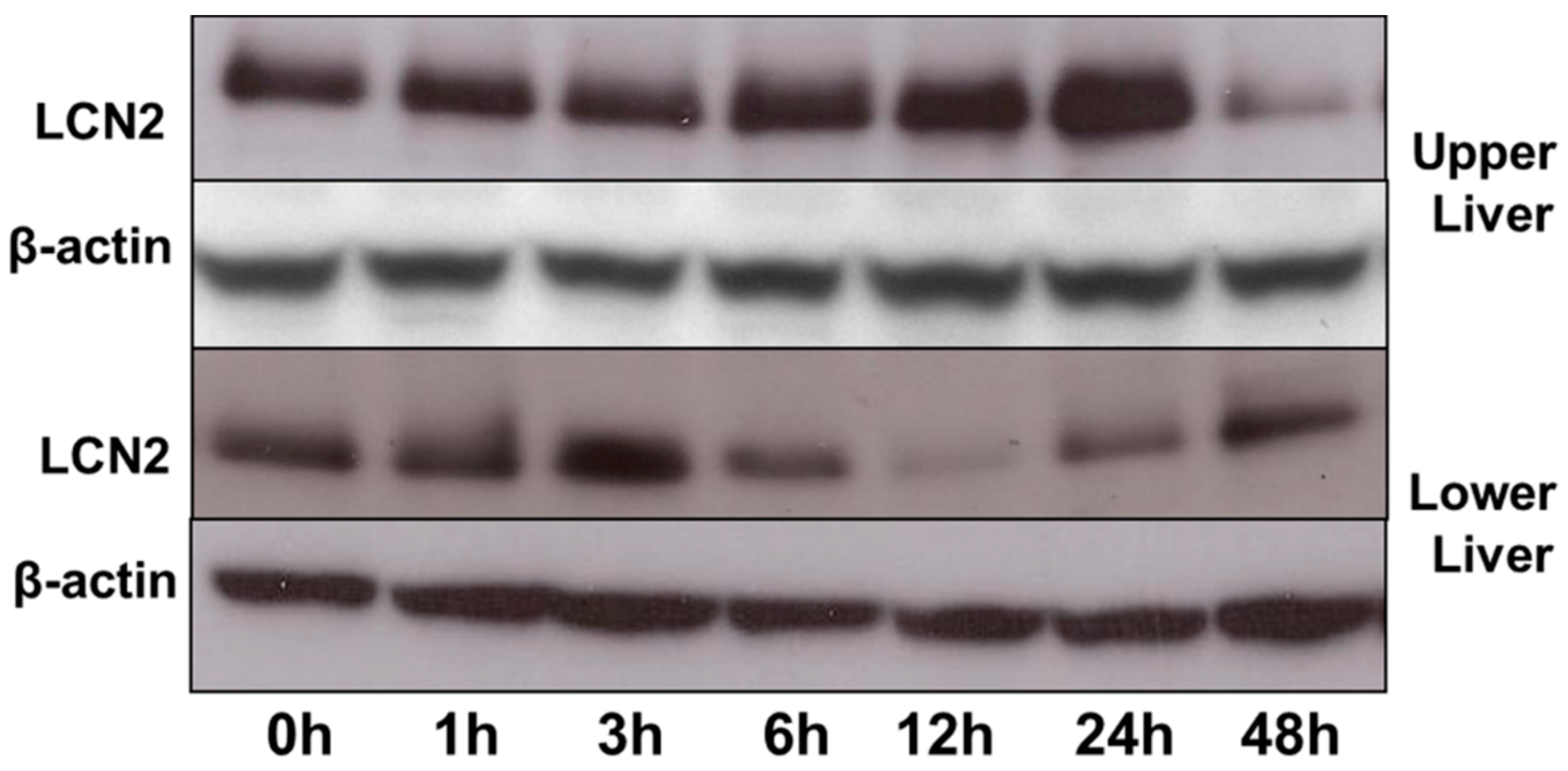
2.7. LCN2 Protein Expression in the Upper and Lower Part of the Liver
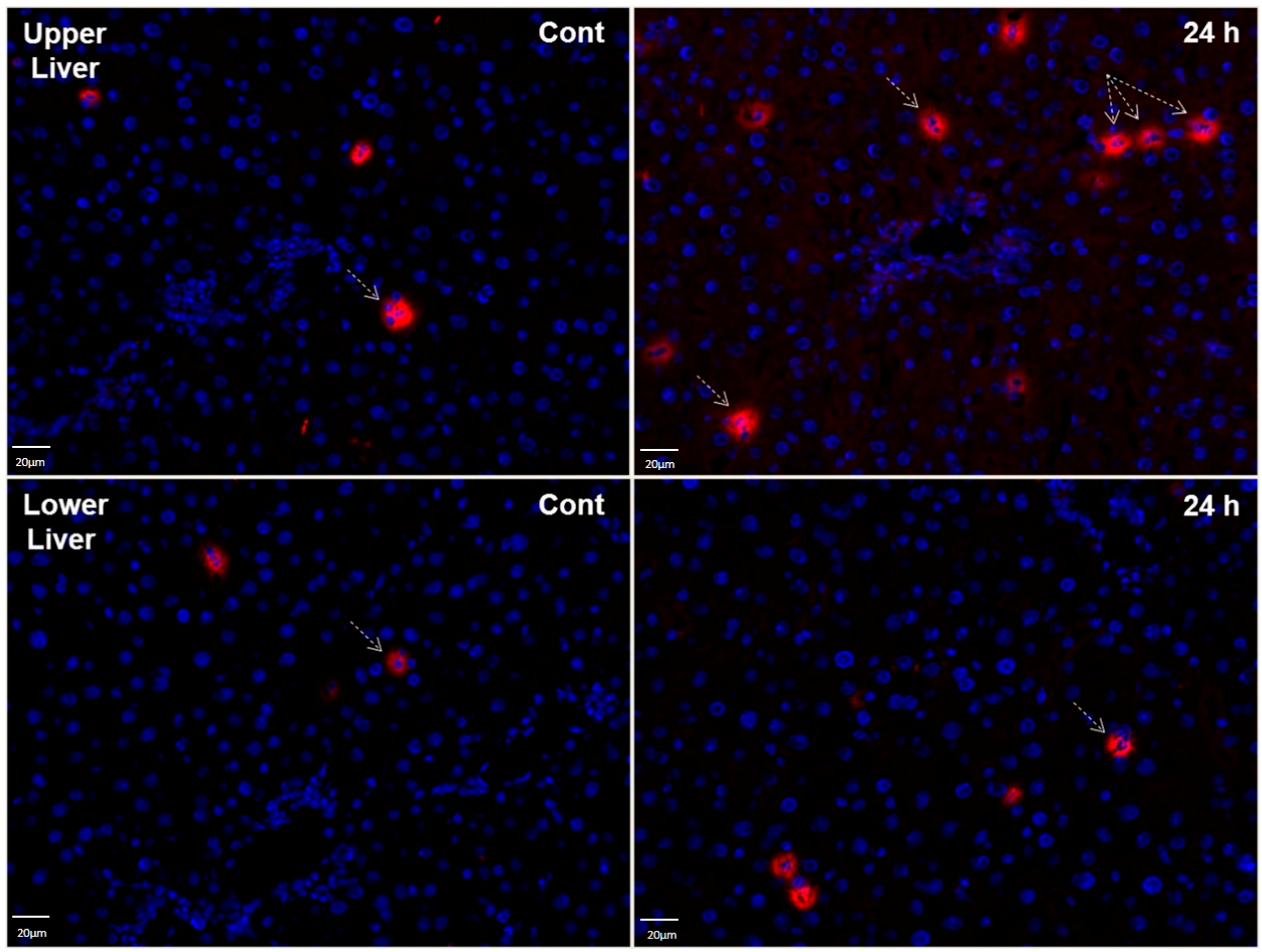
2.8. LCN2 Immunostaining of Liver Tissues after Lung Irradiation
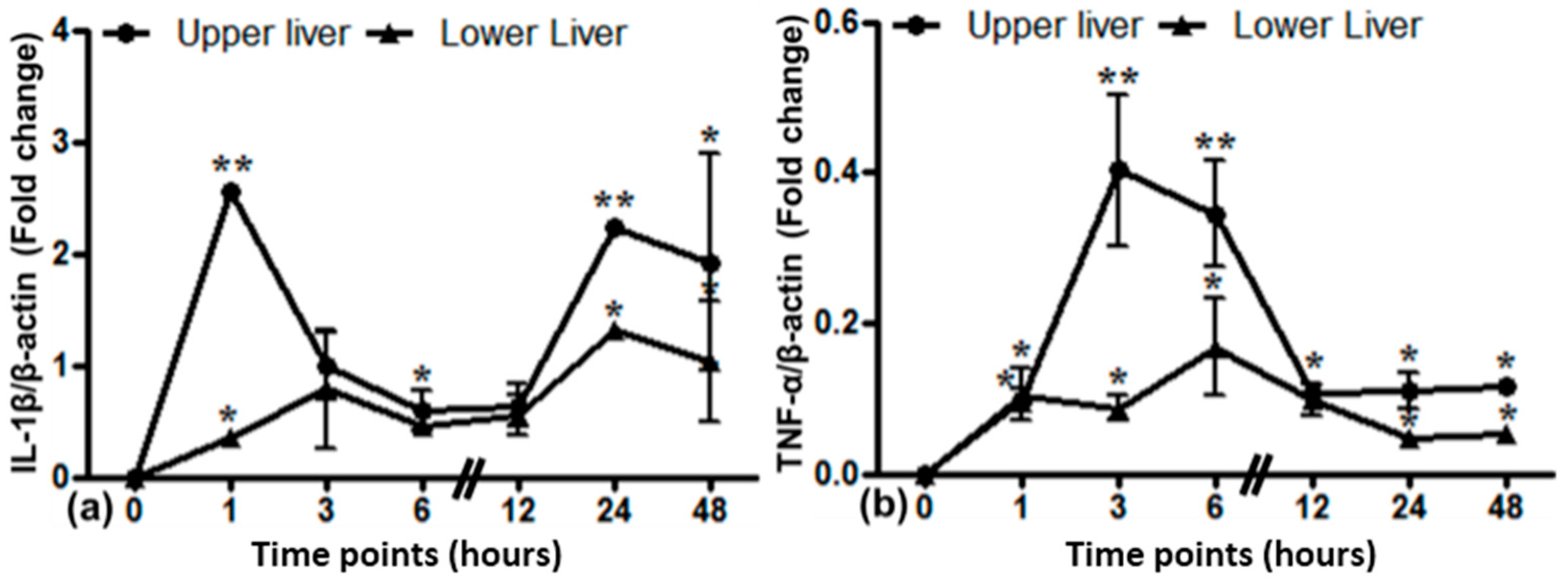
2.9. Gene Expression of Different Cytokines in the Upper and Lower Part of the Liver after Lung Irradiation
3. Discussion
4. Material and Methods
4.1. Animals
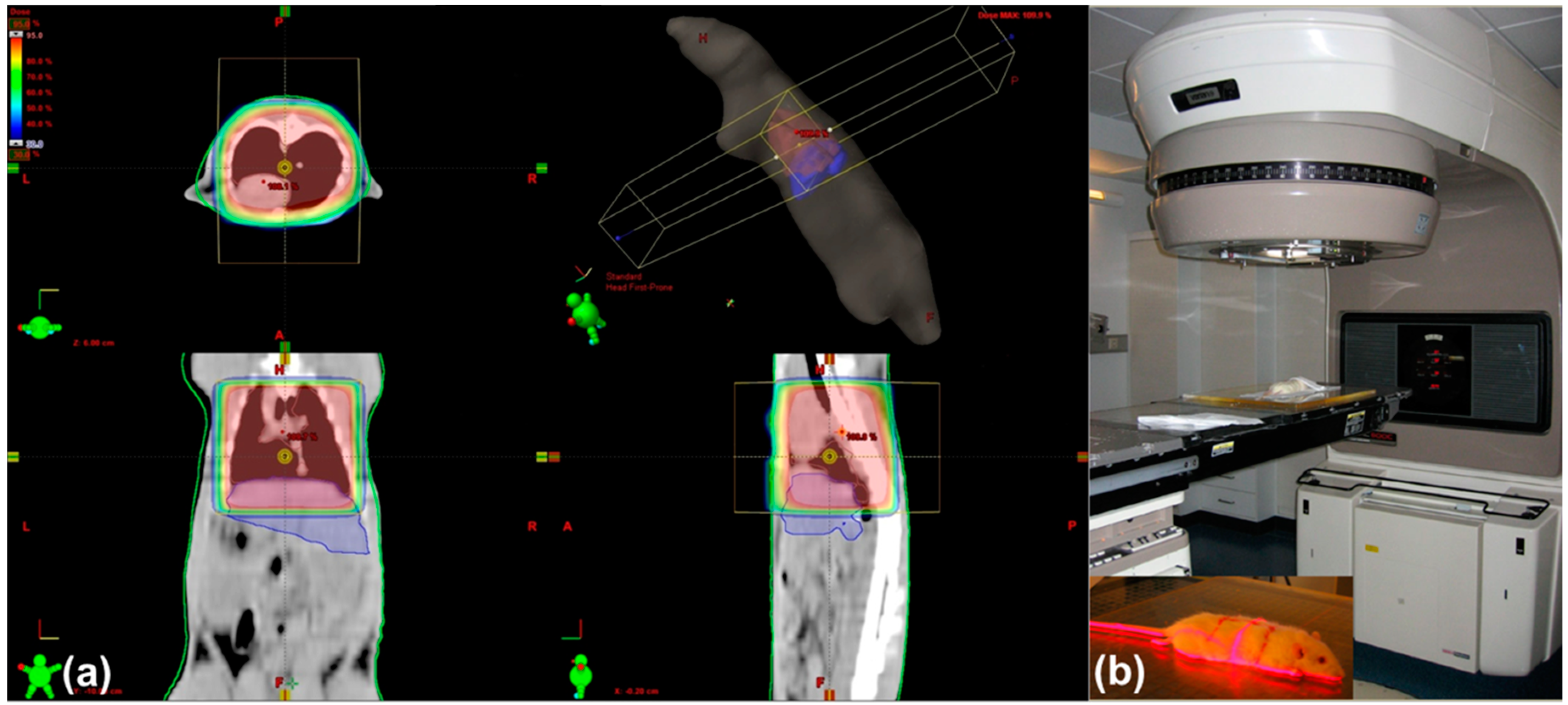
4.2. Irradiation of Rat Lung
4.3. Measurement of Serum LCN2 Levels
4.4. RNA Isolation and RT-PCR
4.5. Protein Extraction and Immunoblotting
4.6. Histology and Immunohistochemistry
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kjeldsen, L.; Johnsen, A.H.; Sengelov, H.; Borregaard, N. Isolation and primary structure of NGAL, a novel protein associated with human neutrophil gelatinase. J. Biol. Chem. 1993, 268, 10425–10432. [Google Scholar] [PubMed]
- Yang, J.; Goetz, D.; Li, J.Y.; Wang, W.; Mori, K.; Setlik, D.; Du, T.; Erdjument-Bromage, H.; Tempst, P.; Strong, R.; et al. An iron delivery pathway mediated by a lipocalin. Mol. Cell 2002, 10, 1045–1056. [Google Scholar] [CrossRef]
- Sultan, S.; Pascucci, M.; Ahmad, S.; Malik, I.A.; Bianchi, A.; Ramadori, P.; Ahmad, G.; Ramadori, G. LIPOCALIN-2 is a major acute-phase protein in a rat and mouse model of sterile abscess. Shock 2012, 37, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Cowland, J.B.; Borregaard, N. Molecular characterization and pattern of tissue expression of the gene for neutrophil gelatinase-associated lipocalin from humans. Genomics 1997, 45, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Alpizar-Alpizar, W.; Laerum, O.D.; Illemann, M.; Ramirez, J.A.; Arias, A.; Malespin-Bendana, W.; Ramírez, V.; Lund, L.R.; Borregaard, N.; Nielsen, B.S. Neutrophil gelatinase-associated lipocalin (NGAL/LCN2) is upregulated in gastric mucosa infected with Helicobacter pylori. Virchows Arch. 2009, 455, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.S.; Borregaard, N.; Bundgaard, J.R.; Timshel, S.; Sehested, M.; Kjeldsen, L. Induction of NGAL synthesis in epithelial cells of human colorectal neoplasia and inflammatory bowel diseases. Gut 1996, 38, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Sunil, V.R.; Patel, K.J.; Nilsen-Hamilton, M.; Heck, D.E.; Laskin, J.D.; Laskin, D.L. Acute endotoxemia is associated with upregulation of lipocalin 24p3/LCN2 in lung and liver. Exp. Mol. Pathol. 2007, 83, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, S.; Tschesche, H. Cloning and expression of human neutrophil lipocalin cDNA derived from bone marrow and ovarian cancer cells. FEBS Lett. 1995, 357, 255–259. [Google Scholar] [CrossRef]
- Furutani, M.; Arii, S.; Mizumoto, M.; Kato, M.; Imamura, M. Identification of a neutrophil gelatinase-associated lipocalin mRNA in human pancreatic cancers using a modified signal sequence trap method. Cancer Lett. 1998, 122, 209–214. [Google Scholar] [CrossRef]
- Shi, H.; Gu, Y.; Yang, J.; Xu, L.; Mi, W.; Yu, W. Lipocalin2 promotes lung metastasis of murine breast cancer cells. J. Exp. Clin. Cancer Res. 2008, 27. [Google Scholar] [CrossRef] [PubMed]
- Travis, S.M.; Conway, B.A.; Zabner, J.; Smith, J.J.; Anderson, N.N.; Singh, P.K.; Greenberg, E.P.; Welsh, M.J. Activity of abundant antimicrobials of the human airway. Am. J. Respir. Cell Mol. Biol. 1999, 20, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H., Jr. Radiation therapy in the management of limited small cell lung cancer: When, where, and how much? Chest 1998, 113, 92S–100S. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.; Cameron, S.; Ahmad, S.; Malik, I.A.; Schultze, F.C.; Hielscher, R.; Rave-Fränk, M.; Hess, C.F.; Ramadori, G.; Christiansen, H. Serum Lipocalin2 is a potential biomarker of liver irradiation damage. Liver Int. 2013, 33, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Flower, D.R. The lipocalin protein family: Structure and function. Biochem. J. 1996, 318 Pt 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, H.Y. Lung inflammation: Normal host defense or a complication of some diseases? Annu. Rev. Med. 1987, 38, 295–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, Y.; Zhang, Y.; Leroith, D.; Bernlohr, D.A.; Chen, X. The role of lipocalin2 in the regulation of inflammation in adipocytes and macrophages. Mol. Endocrinol. 2008, 22, 1416–1426. [Google Scholar] [CrossRef] [PubMed]
- Schroll, A.; Eller, K.; Feistritzer, C.; Nairz, M.; Sonnweber, T.; Moser, P.A.; Rosenkranz, A.R.; Theurl, I.; Weiss, G. Lipocalin2 ameliorates granulocyte functionality. Eur. J. Immunol. 2012, 42, 3346–3357. [Google Scholar] [CrossRef] [PubMed]
- Warszawska, J.M.; Gawish, R.; Sharif, O.; Sigel, S.; Doninger, B.; Lakovits, K.; Mesteri, I.; Nairz, M.; Boon, L.; Spiel, A.; et al. Lipocalin2 deactivates macrophages and worsens pneumococcal pneumonia outcomes. J. Clin. Investig. 2013, 123, 3363–3372. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, T.; Sagawa, M.; Yamato, K.; Xian, M.; Yamamoto, T.; Suematsu, M.; Ikeda, Y.; Kizaki, M. Myeloperoxidase is a key regulator of oxidative stress mediated apoptosis in myeloid leukemic cells. Clin. Cancer Res. 2007, 13, 5436–5445. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Sultan, S.; Naz, N.; Ahmad, G.; Alwahsh, S.M.; Cameron, S.; Moriconi, F.; Ramadori, G.; Malik, I.A. Regulation of iron uptake in primary culture rat hepatocytes: The role of acute phase cytokines. Shock 2014, 41, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.J.; Feng, D.; Wu, H.; Wang, H.; Chan, Y.; Kolls, J.; Borregaard, N.; Porse, B.; Berger, T.; Mak, T.W.; et al. Liver is the major source of elevated serum lipocalin-2 levels after bacterial infection or partial hepatectomy: A critical role for IL-6/STAT3. Hepatology 2015, 61, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.A.; Ten Haken, R.K.; Lawrence, T.S. Partial irradiation of the liver. Semin. Radiat. Oncol. 2001, 11, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Palma, D.A.; IASLC Advanced Radiation Technology Committee. Follow-up of patients after stereotactic radiation for lung cancer: A primer for the nonradiation oncologist. J. Thorac. Oncol. 2015, 10, 412–419. [Google Scholar] [CrossRef] [PubMed]
- La Manna, G.; Ghinatti, G.; Tazzari, P.L.; Alviano, F.; Ricci, F.; Capelli, I.; Cuna, V.; Todeschini, P.; Brunocilla, E.; Pagliaro, P.; et al. Neutrophil gelatinase-associated lipocalin increases HLA-G+/FoxP3+ T-regulatory cell population in an in vitro model of PBMC. PLoS ONE 2014, 9, e89497. [Google Scholar]
- Rodvold, J.J.; Mahadevan, N.R.; Zanetti, M. Lipocalin2 in cancer: When good immunity goes bad. Cancer Lett. 2012, 316, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S. Serum Lipocalin-2 (LCN-2) as a Major Acute Phase Protein under Different Pathological Conditions: In vivo and in vitro Studies. Ph.D. Thesis, University of Göttingen, Göttingen, Germany, 2012. [Google Scholar]
- Malik, I.A.; Moriconi, F.; Sheikh, N.; Naz, N.; Khan, S.; Dudas, J.; Mansuroglu, T.; Hess, C.F.; Rave-Fränk, M.; Christiansen, H.; et al. Single-dose γ-irradiation induces up-regulation of chemokine gene expression and recruitment of granulocytes into the portal area but not into other regions of rat hepatic tissue. Am. J. Pathol. 2010, 176, 1801–1815. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, G.; Moebius, U.; Dienes, H.P.; Meuer, S.; Meyer zum Buschenfelde, K.H. Lymphocytes from hepatic inflammatory infiltrate kill rat hepatocytes in primary culture. Comparison with peripheral blood lymphocytes. Virchows Arch. B Cell. Pathol. Incl. Mol. Pathol. 1990, 59, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, P.; Ahmad, G.; Ramadori, G. Cellular and molecular mechanisms regulating the hepatic erythropoietin expression during acute-phase response: A role for IL-6. Lab. Investig. 2010, 90, 1306–1324. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zhang, H.; Jiang, L.; Chi, Y.; Tian, J.; Du, W.; Yu, B.; Han, Z. Down-regulation of lipocalin2 suppresses the growth of human lung adenocarcinoma through oxidative stress involving Nrf2/HO-1 signaling. Acta Biochim. Biophys. Sin. (Shanghai) 2015, 47, 805–814. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Rat) | Forward 5′–3′ | Reverse 5’–3’ |
|---|---|---|
| LCN2 | GGAATATTCACAGCTACCCTC | TTGTTATCCTTGAGGCCCAG |
| β-actin | TGTCACCAACTGGGACGATA | AACACAGCCTGGATGGCTAC |
| IL-6 | GTCAACTCCATCTGCCCTTCAG | GGCAGTGGCTGTCAACAACAT |
| IL-1β | TACCTATGTCTGGCCCGTGGAG | ATCATCCCACGAGTCACACAGG |
| TNF-α | ACAAGGCTGCCCCGACTAT | CTCCTGGTATGAAGTGGCAAATC |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultan, S.; Ahmad, S.; Rave-Fränk, M.; Malik, I.A.; Hess, C.F.; Christiansen, H.; Cameron, S. Induction of Lipocalin2 in a Rat Model of Lung Irradiation. Int. J. Mol. Sci. 2016, 17, 637. https://doi.org/10.3390/ijms17050637
Sultan S, Ahmad S, Rave-Fränk M, Malik IA, Hess CF, Christiansen H, Cameron S. Induction of Lipocalin2 in a Rat Model of Lung Irradiation. International Journal of Molecular Sciences. 2016; 17(5):637. https://doi.org/10.3390/ijms17050637
Chicago/Turabian StyleSultan, Sadaf, Shakil Ahmad, Margret Rave-Fränk, Ihtzaz Ahmed Malik, Clemens F. Hess, Hans Christiansen, and Silke Cameron. 2016. "Induction of Lipocalin2 in a Rat Model of Lung Irradiation" International Journal of Molecular Sciences 17, no. 5: 637. https://doi.org/10.3390/ijms17050637
APA StyleSultan, S., Ahmad, S., Rave-Fränk, M., Malik, I. A., Hess, C. F., Christiansen, H., & Cameron, S. (2016). Induction of Lipocalin2 in a Rat Model of Lung Irradiation. International Journal of Molecular Sciences, 17(5), 637. https://doi.org/10.3390/ijms17050637