
Expression of Castor LPAT2 Enhances Ricinoleic Acid Content at the sn-2 Position of Triacylglycerols in Lesquerella Seed

 and
and
Abstract
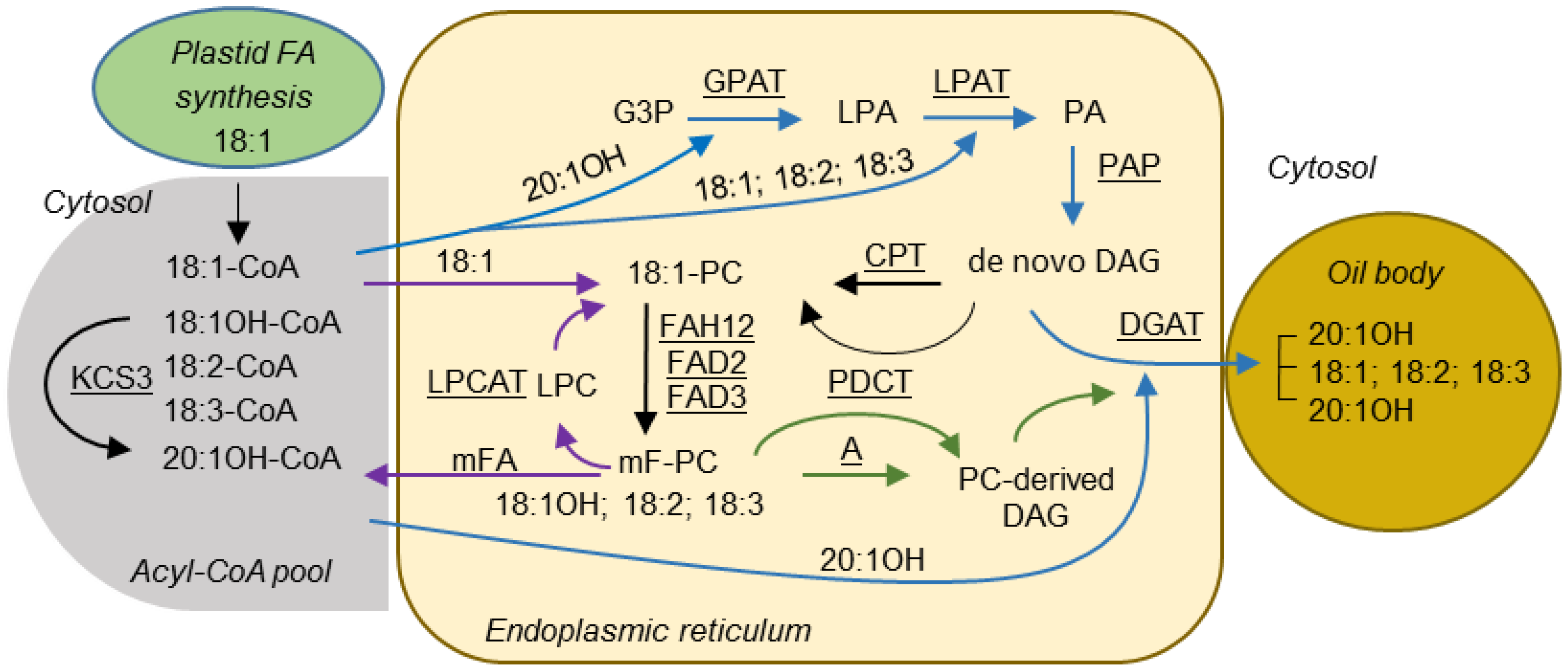
:1. Introduction
2. Results
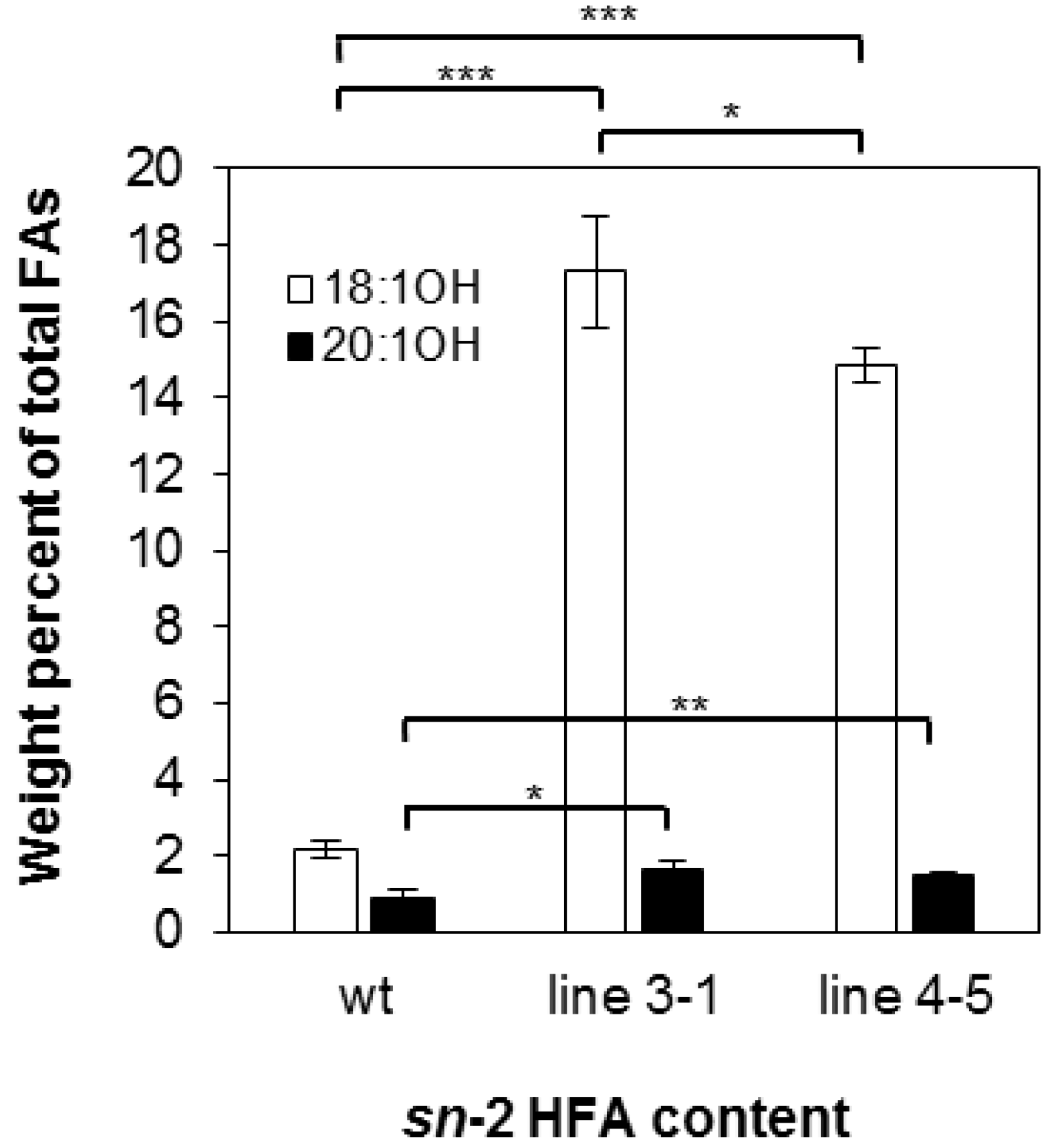
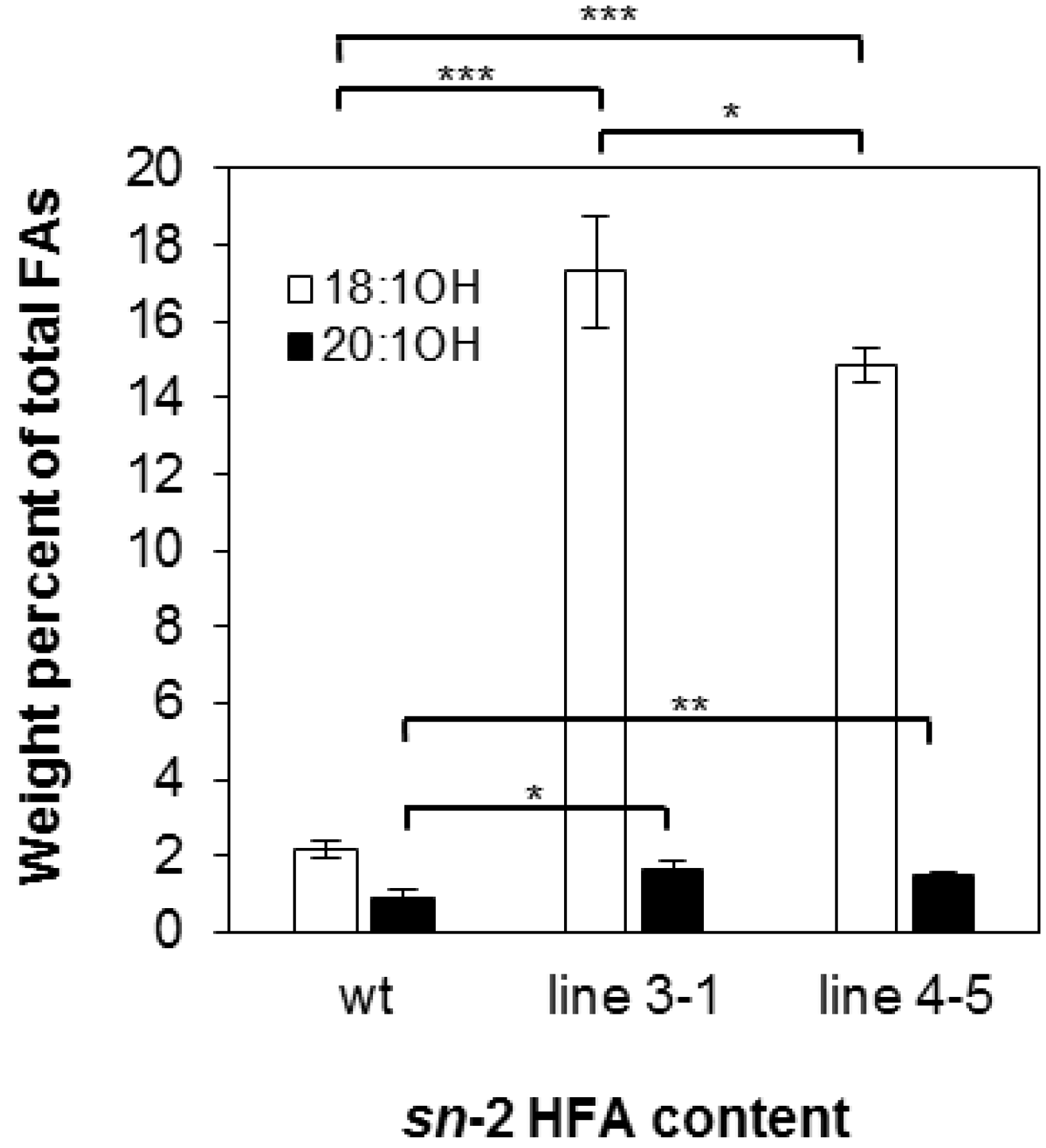
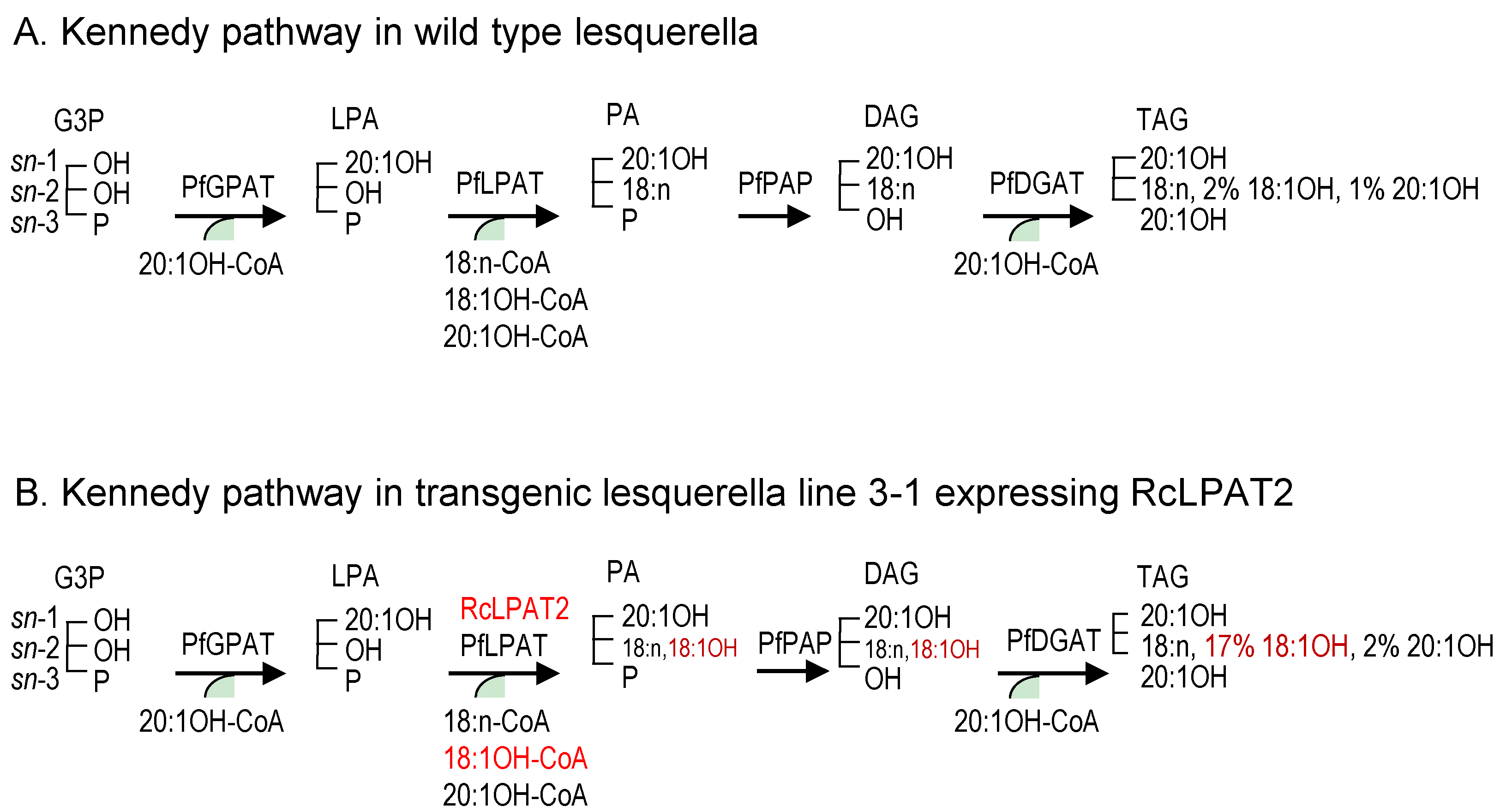
2.1. Castor Lysophosphatidic Acid Acyltransferase 2 (RcLPAT2) Increased 18:1OH Accumulation in Transgenic Seeds
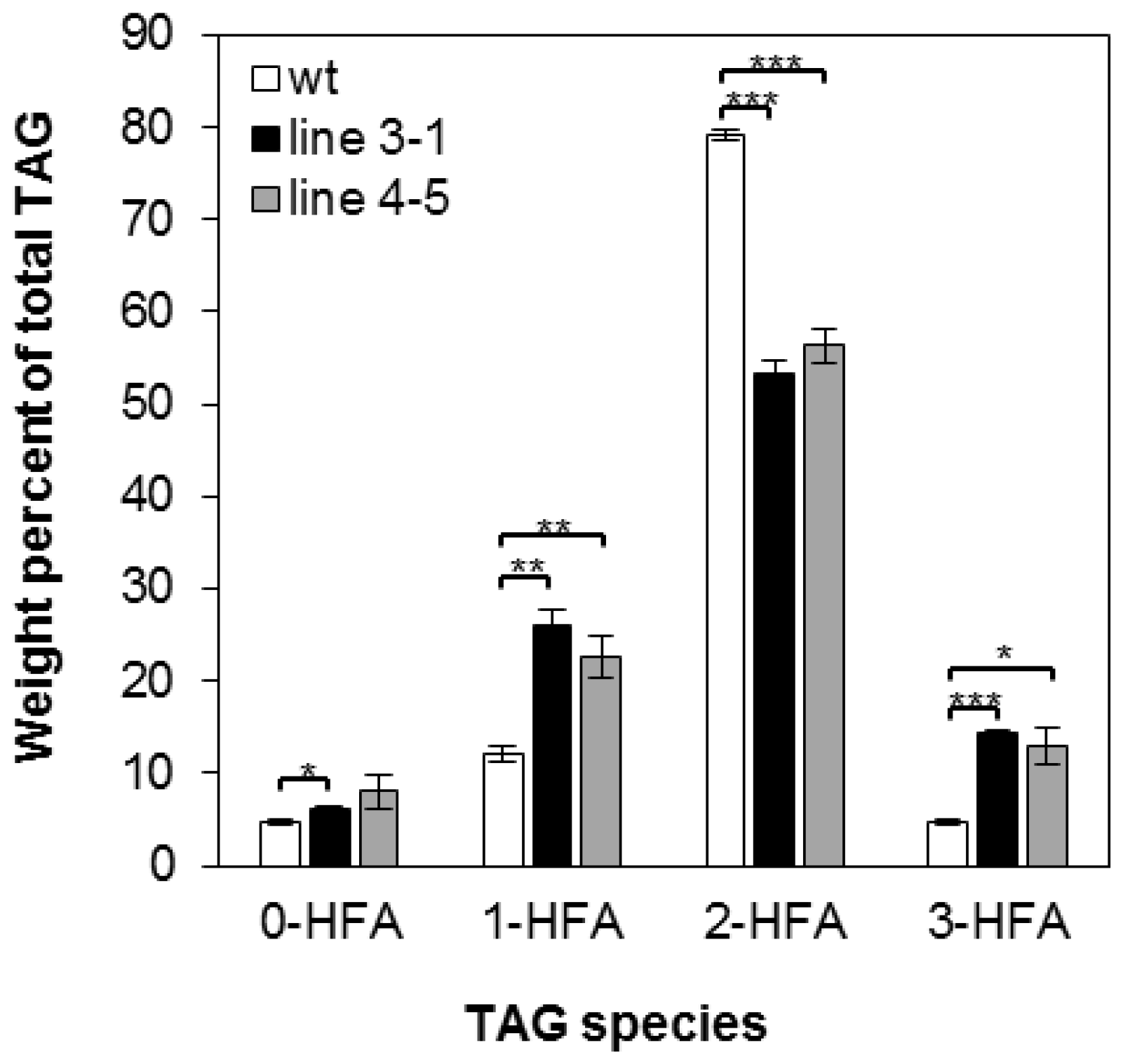
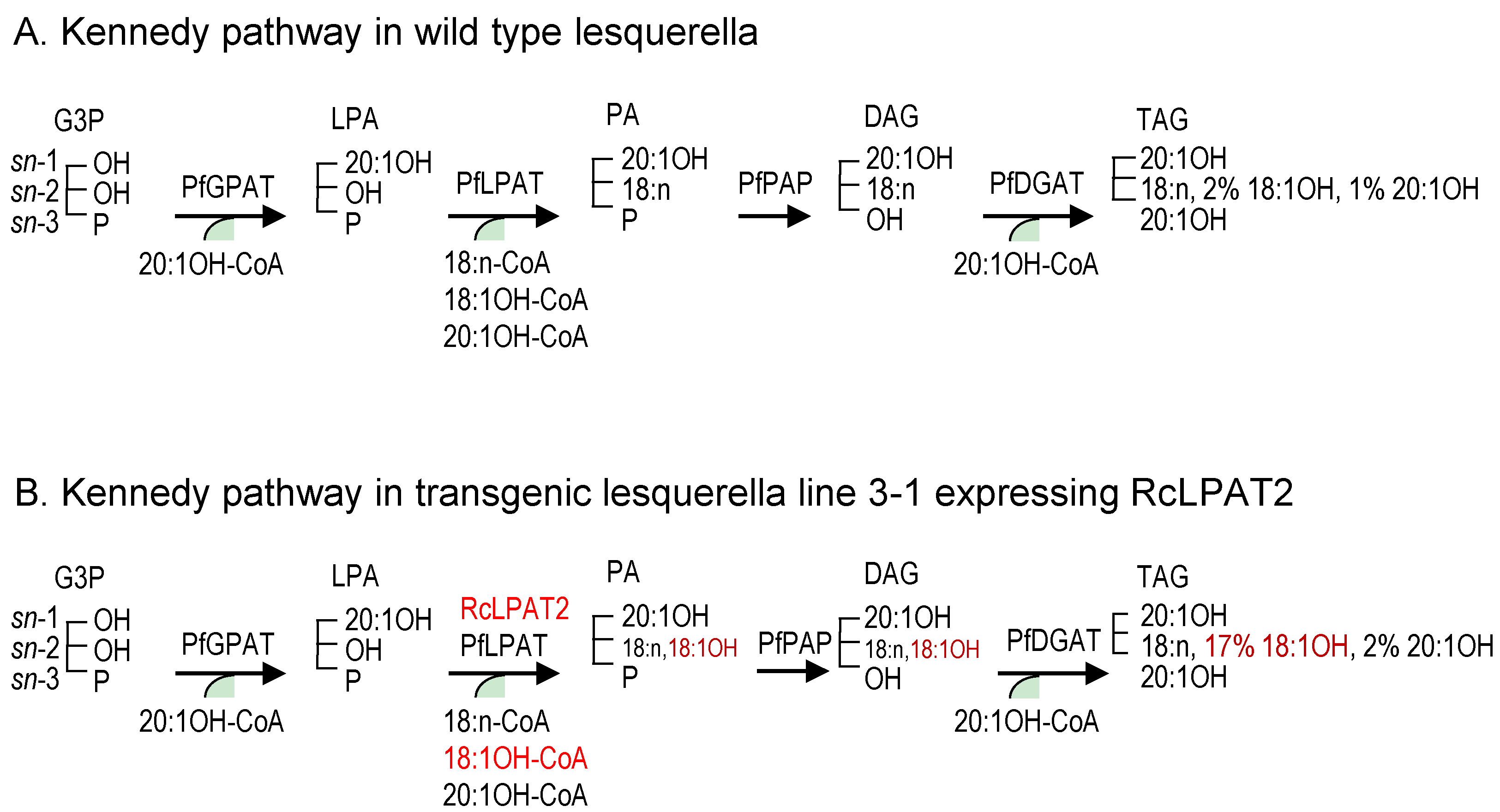
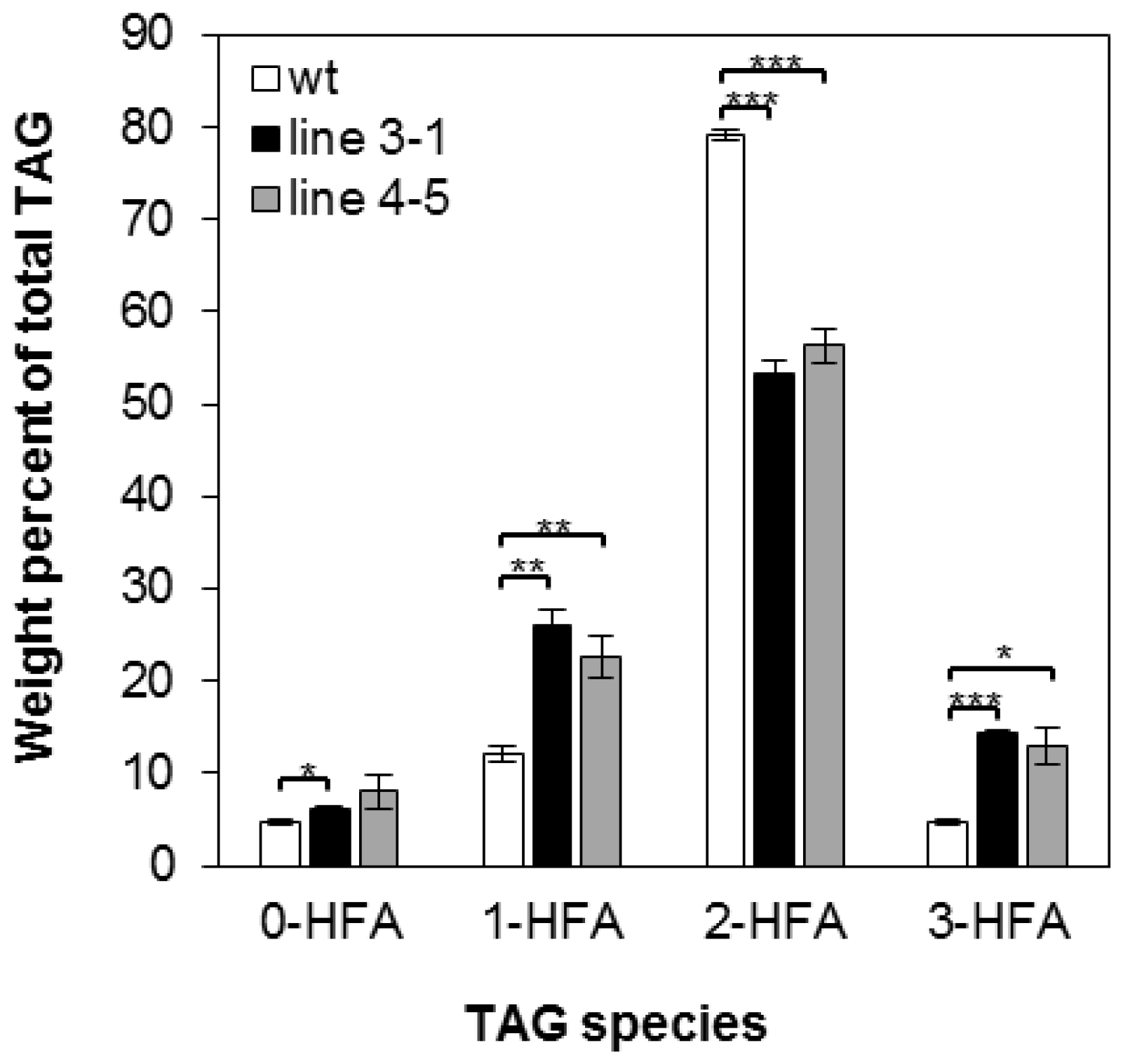
2.2. Overexpression of RcLPAT2 Altered the Composition of Hydroxy Fatty Acid (HFA)-Containing Triacylglycerol (TAG) Species and Increased the Percentage of 18:1OH at the sn-2 Position
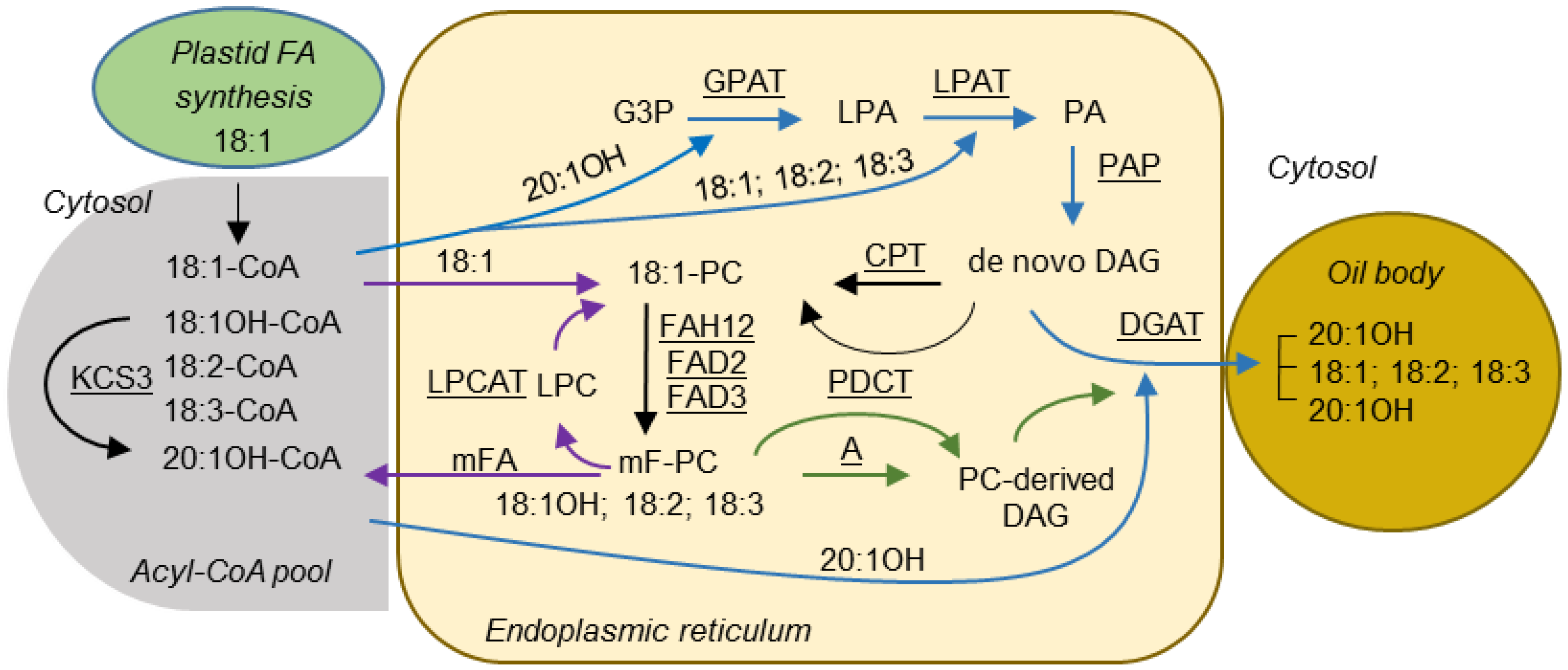
3. Discussion
4. Materials and Methods
4.1. Construction of pGPro4-PnapA-RcLPAT2 Binary Vector
4.2. Plant Transformation, and Estimation of Transgene Copy Number
4.3. Lipid Extraction, TAG Species, Regiochemical and GC Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Al-Shehbaz, I.A.; O’Kane, S.L., Jr. Lesquerella is united with Physaria (Brassicaceae). Novon 2002, 12, 319–329. [Google Scholar] [CrossRef]
- Smith, C.R., Jr.; Wilson, T.L.; Miwa, T.K.; Zobel, H.; Lohmar, R.L.; Wolff, I.A. Lesquerolic acid. A new hydroxy acid from Lesquerella seed oil. J. Org. Chem. 1961, 26, 2903–2905. [Google Scholar] [CrossRef]
- Hayes, D.G.; Kleiman, R. A detailed triglyceride analysis of Lesquerella fendleri oil: Column chromatographic fractionation followed by supercritical fluid chromatography. J. Am. Oil Chem. Soc. 1996, 73, 267–269. [Google Scholar] [CrossRef]
- Dierig, D.A.; Wang, G.; McCloskey, W.B.; Thorp, K.R.; Isbell, T.A.; Ray, D.T.; Foster, M.A. Lesquerella: New crop development and commercialization in the U.S. Ind. Crops Prod. 2011, 34, 1381–1385. [Google Scholar] [CrossRef]
- Isbell, T.A.; Mund, M.S.; Evangelista, R.L.; Dierig, D.A. Method for analysis of fatty acid distribution and oil content on a single Lesquerella fendleri seed. Ind. Crops Prod. 2008, 28, 231–236. [Google Scholar] [CrossRef]
- Chen, G.Q.; Lin, J.T.; Lu, C. Hydroxy fatty acid synthesis and lipid gene expression during seed development in Lesquerella fendleri. Ind. Crops Prod. 2010, 34, 1286–1292. [Google Scholar] [CrossRef]
- Hayes, D.G.; Kleiman, R. Recovery of hydroxy fatty acids from Lesquerella oil with lipases. J. Am. Oil Chem. Soc. 1992, 69, 982–985. [Google Scholar] [CrossRef]
- Lin, J.T.; Fagerquist, C.K.; Chen, G.Q. Ratios of regioisomers of the molecular species of triacylglycerols in Lesquerella (Physaria fendleri) oil estimated by mass spectrometry. J. Am. Oil Chem. Soc. 2016, 93, 183–191. [Google Scholar] [CrossRef]
- Caupin, H.J. Products from Castor Oil: Past, Present and Future; Gunstone, F.D., Padley, F.B., Eds.; Marcel Dekker: New York, NY, USA, 1997; pp. 787–795. [Google Scholar]
- Chen, G.Q. Effective reduction of chimeric tissue in transgenics for the stable genetic transformation of Lesquerella fendleri. HortScience 2011, 46, 86–90. [Google Scholar]
- Chapman, K.D.; Ohlrogge, J.B. Compartmentation of triacylglycerol accumulation in plants. J. Biol. Chem. 2012, 287, 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.P. Biosynthesis of complex lipids. Fed. Proc. 1961, 20, 934–940. [Google Scholar] [PubMed]
- Weiss, S.B.; Kennedy, E.P.; Kiyasu, J.Y. The enzymatic synthesis of triglycerides. J. Biol. Chem. 1960, 235, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Barron, E.J.; Stumpf, P.K. Fat metabolism in higher plants XIX. The biosynthesis of triglycerides by avocado-mesocarp enzymes. Biochim. Biophys. Acta 1962, 60, 329–337. [Google Scholar] [CrossRef]
- Lands, W.E. Lipid metabolism. Annu. Rev. Biochem. 1965, 34, 313–346. [Google Scholar] [CrossRef] [PubMed]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; DeBono, A.; Durrett, T.P.; et al. Acyl-lipid metabolism. Arabidopsis Book 2013, 11, e0161. [Google Scholar] [CrossRef] [PubMed]
- Dahlqvist, A.; Stahl, U.; Lenman, M.; Banas, A.; Lee, M.; Sandager, L.; Ronne, H.; Stymne, S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Natl. Acad. Sci. USA 2000, 97, 6487–6492. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Durrett, T.P.; Ohlrogge, J.B.; Pollard, M. Analysis of acyl fluxes through multiple pathways of triacylglycerol synthesis in developing soybean embryos. Plant Physiol. 2009, 150, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Browse, J. The pathway of triacylglycerol synthesis through phosphatidylcholine in Arabidopsis produces a bottleneck for the accumulation of unusual fatty acids in transgenic seeds. Plant J. 2011, 68, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Ren, Z.; Lu, C. The phosphatidylcholine diacylglycerol cholinephosphotransferase is required for efficient hydroxy fatty acid accumulation in transgenic Arabidopsis. Plant Physiol. 2012, 158, 1944–1954. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Xin, Z.; Ren, Z.; Miquel, M.; Browse, J. An enzyme regulating triacylglycerol composition is encoded by the ROD1 gene of Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18837–18842. [Google Scholar] [CrossRef] [PubMed]
- Slack, C.R.; Campbell, L.C.; Browse, J.A.; Roughan, P.G. Some evidence for the reversibility of the cholinephosphotransferasecatalysed reaction in developing linseed cotyledons in vivo. Biochim. Biophys. Acta 1983, 754, 10–20. [Google Scholar] [CrossRef]
- Lin, J.T.; Turner, C.; Liao, L.P.; McKeon, T.A. Identification and quantification of the molecular species of acylglycerols in castor oil by HPLC using ELSD. J. Liquid Chromatogr. Relat. Technol. 2003, 26, 773–780. [Google Scholar] [CrossRef]
- Moreau, R.A.; Stumpf, P.K. Recent studies of the enzymic synthesis of ricinoleic acid by developing castor beans. Plant Physiol. 1981, 67, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Bafor, M.; Smith, M.A.; Jonsson, L.; Stobart, K.; Stymne, S. Ricinoleic acid biosynthesis and triacylglycerol assembly in microsomal preparations from developing castor-bean (Ricinus communis) endosperm. Biochem. J. 1991, 280, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Engeseth, N.; Stymne, S. Desaturation of oxygenated fatty acids in Lesquerella and other oil seeds. Planta 1996, 198, 238–245. [Google Scholar] [CrossRef]
- Reed, D.W.; Taylor, D.C.; Covello, P.S. Metabolism of hydroxy fatty acids in developing seeds in the genera Lesquerella (Brassicaceae) and Linum (Linaceae). Plant Physiol. 1997, 114, 63–68. [Google Scholar] [PubMed]
- Moon, H.; Smith, M.A.; Kunst, L. A condensing enzyme from the seeds of Lesquerella fendleri that specifically elongates hydroxy fatty acids. Plant Physiol. 2001, 127, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-R.; Chen, G.Q.; Kim, H.U. Current progress towards the metabolic engineering of plant seed oil for hydroxy fatty acids production. Plant Cell Rep. 2015, 34, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Van de Loo, F.J.; Broun, P.; Turner, S.; Somerville, C. An oleate 12-hydroxylase from Ricinus communis L. is a fatty acyl desaturase homolog. Proc. Natl. Acad. Sci. USA 1995, 92, 6743–6747. [Google Scholar] [CrossRef] [PubMed]
- Broun, P.; Boddupalli, S.; Somerville, C. A bifunctional oleate 12-hydroxylase: Desaturase from Lesquerella fendleri. Plant J. 1998, 13, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Broun, P.; Somerville, C. Accumulation of ricinoleic, lesquerolic, and densipolic acids in seeds of transgenic arabidopsis plants that express a fatty acyl hydroxylase cDNA from castor bean. Plant Physiol. 1997, 113, 933–942. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Smith, M.A.; Moon, H.; Chowrira, G.; Kunst, L. Heterologous expression of a fatty acid hydroxylase gene in developing seeds of Arabidopsis thaliana. Planta 2003, 217, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Wallis, J.G.; Skidmore, C.; Browse, J. A mutation in Arabidopsis cytochrome b5 reductase identified by high-throughput screening differentially affects hydroxylation and desaturation. Plant J. 2006, 48, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Fulda, M.; Wallis, J.G.; Browse, J. A high-throughput screen for genes from castor that boost hydroxy fatty acid accumulation in seed oils of transgenic Arabidopsis. Plant J. 2006, 45, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Burgal, J.; Shockey, J.; Lu, C.; Dyer, J.; Larson, T.; Graham, I.; Browse, J. Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol. J. 2008, 6, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Van Erp, H.; Bates, P.D.; Burgal, J.; Shockey, J.; Browse, J. Castor phospholipid:diacylglycerol acyltransferase facilitates efficient metabolism of hydroxy fatty acids in transgenic Arabidopsis. Plant Physiol. 2011, 155, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Lee, K.R.; Go, Y.S.; Jung, J.H.; Suh, M.C.; Kim, J.B. Endoplasmic reticulum-located PDAT1-2 from castor bean enhances hydroxy fatty acid accumulation in transgenic plants. Plant Cell Physiol. 2011, 52, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Van Erp, H.; Shockey, J.; Zhang, M.; Adhikari, N.D.; Browse, J. Reducing isozyme competition increases target fatty acid accumulation in seed triacylglycerols of transgenic Arabidopsis. Plant Physiol. 2015, 168, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Frentzen, M. Acyltransferases from basic science to modified seed oils. Lipid/Fett 1998, 100, 161–166. [Google Scholar] [CrossRef]
- Phillips, B.E.; Smith, C.R., Jr.; Tallent, W.H. Glycerides of Limnanthes douglasii seed oil. Lipids 1971, 6, 93–99. [Google Scholar] [CrossRef]
- Balachandran, C.; Arumughan, C.; Mathew, A.G. Distribution of major chemical constituents and fatty acids in different regions of coconut endosperm. J. Am. Oil Chem. Soc. 1985, 62, 1583–1586. [Google Scholar] [CrossRef]
- Brough, C.L.; Coventry, J.M.; Christie, W.W.; Kroon, J.T.M.; Brown, A.P.; Barsby, T.L.; Slabas, A.R. Towards the genetic engineering of triacylglycerols of defined fatty acid composition: Major changes in erucic acid content at the sn-2 position affected by the introduction of a 1-acyl-sn-glycerol-3-phosphate acyltransferase from Limnanthes douglasii into oil seed rape. Mol. Breed. 1996, 2, 133–142. [Google Scholar]
- Knutzon, D.S.; Hayes, T.R.; Wyrick, A.; Xiong, H.; Davies, H.M.; Voelker, T.A. Lysophosphatidic acid acyltransferase from coconut endosperm mediates the insertion of laurate at the sn-2 position of triacylglycerols in lauric rapeseed oil and can increase total laurate levels. Plant Physiol. 1999, 120, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Lassner, M.W.; Levering, C.K.; Davies, H.M.; Knutzon, D.S. Lysophosphatidic acid acyltransferase from meadowfoam mediates insertion of erucic acid at the sn-2 position of triacylglycerol in transgenic rapeseed oil. Plant Physiol. 1995, 109, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Katavic, V.; Giblin, E.M.; Barton, D.L.; MacKenzie, S.L.; Keller, W.A.; Hu, X.; Taylor, D.C. Modification of seed oil content and acyl composition in the brassicaceae by expression of a yeast sn-2 acyltransferase gene. Plant Cell 1997, 9, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Huang, A.H.C. Plastid lysophosphatidyl acyltransferase is essential for embryo development in Arabidopsis. Plant Physiol. 2004, 134, 1206–1216. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Li, Y.; Huang, A.H. Ubiquitous and endoplasmic reticulum-located lysophosphatidyl acyltransferase, LPAT2, is essential for female but not male gametophyte development in Arabidopsis. Plant Cell 2005, 17, 1073–1089. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.H.; Prakash, R.R.; Sweet, M.; Shanklin, J. Coexpressing Escherichia coli cyclopropane synthase with Sterculia foetida Lysophosphatidic acid acyltransferase enhances cyclopropane fatty acid accumulation. Plant Physiol. 2014, 164, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Cagliari, A.; Margis-Pinheiro, M.; Loss, G.; Mastroberti, A.A.; de Araujo Mariath, J.E.; Margis, R. Identification and expression analysis of castor bean (Ricinus communis) genes encoding enzymes from the triacylglycerol biosynthesis pathway. Plant Sci. 2010, 179, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Troncoso-Ponce, M.A.; Kilaru, A.; Cao, X.; Durrett, T.P.; Fan, J.; Jensen, J.K.; Thrower, N.A.; Pauly, M.; Wilkerson, C.; Ohlrogge, J.B. Comparative deep transcriptional profiling of four developing oilseeds. Plant J. 2011, 68, 1014–1027. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.P.; Kroon, J.T.; Swarbreck, D.; Febrer, M.; Larson, T.R.; Graham, I.A.; Caccamo, M.; Slabas, A.R. Tissue-specific whole transcriptome sequencing in castor, directed at understanding triacylglycerol lipid biosynthetic pathways. PLoS ONE 2012, 7, e30100. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Caro, J.M.; Chileh, T.; Kazachkov, M.; Zou, J.; Alonso, D.L.; Garcia-Maroto, F. The multigene family of lysophosphatidate acyltransferase (LPAT)-related enzymes in Ricinus communis: Cloning and molecular characterization of two LPAT genes that are expressed in castor seeds. Plant Sci. 2013, 199–200, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.T.; Chen, G.Q. Identification of TAG and DAG and their FA constituents in Lesquerella (Physaria fendleri) oil by HPLC and MS. J. Am. Oil Chem. Soc. 2013, 90, 1819–1829. [Google Scholar] [CrossRef]
- Dauk, M.; Lam, P.; Kunst, L.; Smith, M.A. A FAD2 homologue from Lesquerella lindheimeri has predominantly fatty acid hydroxylase activity. Plant Sci. 2007, 173, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Lozinsky, S.; Yang, H.; Forseille, L.; Cook, G.R.; Ramirez-Erosa, I.; Smith, M.A. Characterization of an oleate 12-desaturase from Physaria fendleri and identification of 5′UTR introns in divergent FAD2 family genes. Plant Physiol. Biochem. PPB/Soc. Fr. Physiol. Veg. 2014, 75, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Chen, G.Q. Identification of hydroxy fatty acid and triacylglycerol metabolism-related genes in Lesquerella through seed transcriptome analysis. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Rice, A.; McGlew, K.; Shaw, V.; Park, H.; Clemente, T.; Pollard, M.; Ohlrogge, J.; Durrett, T.P. Metabolic engineering of oilseed crops to produce high levels of novel acetyl glyceride oils with reduced viscosity, freezing point and calorific value. Plant Biotechnol. J. 2015, 13, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Carlsson, A.S.; Francis, T.; Zhang, M.; Hoffman, T.; Giblin, M.E.; Taylor, D.C. Triacylglycerol synthesis by PDAT1 in the absence of DGAT1 activity is dependent on re-acylation of LPC by LPCAT2. BMC Plant Biol. 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Browse, J. The significance of different diacylgycerol synthesis pathways on plant oil composition and bioengineering. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shen, W.; Kazachkov, M.; Chen, G.; Chen, Q.; Carlsson, A.S.; Stymne, S.; Weselake, R.J.; Zou, J. Metabolic interactions between the lands cycle and the Kennedy pathway of glycerolipid synthesis in Arabidopsis developing seeds. Plant Cell 2012, 24, 4652–4669. [Google Scholar] [CrossRef] [PubMed]
- Lager, I.; Yilmaz, J.L.; Zhou, X.R.; Jasieniecka, K.; Kazachkov, M.; Wang, P.; Zou, J.; Weselake, R.; Smith, M.A.; Bayon, S.; et al. Plant acyl-CoA:lysophosphatidylcholine acyltransferases (LPCATs) have different specificities in their forward and reverse reactions. J. Biol. Chem. 2013, 288, 36902–36914. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Thilmony, R.; Lin, J.-T. Transformation of Lesquerella fendleri with the new binary vector pGPro4-35s. OnLine J. Biol. Sci. 2011, 11, 90–95. [Google Scholar]
- Chen, G.Q.; Lin, J.-T. A napin promoter activates gene expression in developing seeds of Lesquerella fendleri. OnLine J. Biol. Sci. 2012, 12, 113–117. [Google Scholar] [CrossRef]
- Dierig, D.A.; Tomasi, P.M.; Dahlquist, G.H. Registration of WCL-LY2 high oil Lesquerella fendleri germplasm. Crop Sci. 2001, 41, 604–605. [Google Scholar] [CrossRef]
- Lazo, G.R.; Stein, P.A.; Ludwig, R.A. A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. Nat. Biotechnol. 1991, 9, 963–967. [Google Scholar] [CrossRef]
- Chen, G.Q.; Lin, J.-T. Use of quantitative polymerase chain reaction for determining copy numbers of transgenes in Lesquerella fendleri. Am. J. Agric. Biol. Sci. 2010, 5, 415–421. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HFA Content | 18:1OH | 18:2OH | 20:1OH | 20:2OH | Total HFA | |
|---|---|---|---|---|---|---|
| Wild type | 1.20 ± 0.09 | 0.24 ± 0.06 | 51.74 ± 0.98 | 3.07 ± 0.39 | 56.18 ± 0.48 | |
| Transgenics | ||||||
| ID | Copy number | |||||
| Line 1 | 2 | 3.79 ± 0.04 *** | 0.53 ± 0.04 ** | 45.64 ± 1.32 * | 3.11 ± 0.15 | 53.08 ± 1.46 |
| Line 2 | 1 | 1.74 ± 0.08 * | 0.30 ± 0.08 | 51.43 ± 0.50 | 3.45 ± 0.21 | 56.91 ± 0.33 |
| Line 3 | 1 | 4.08 ± 0.31 *** | 0.74 ± 0.12 * | 48.99 ± 0.69 | 3.60 ± 0.11 | 57.81 ± 0.34 |
| Line 4 | 2 | 3.81 ± 0.06 *** | 0.60 ± 0.03 ** | 50.02 ± 0.48 | 3.38 ± 0.24 | 57.81 ± 0.34 |
| Line 5 | 1 | 3.97 ± 0.13 *** | 0.75 ± 0.20 * | 48.35 ± 1.88 | 3.58 ± 0.31 | 56.64 ± 1.27 |
| Line 7 | 1 | 3.29 ± 0.12 *** | 0.34 ± 0.03 | 51.09 ± 0.22 | 2.84 ± 0.15 | 57.56 ± 0.21 |
| Line 8 | 1 | 1.42 ± 0.08 ** | 0.15 ± 0.07 | 48.24 ± 1.41 | 2.75 ± 0.20 | 52.56 ± 1.26 |
| Line 9 | 1 | 2.87 ± 0.21 *** | 0.53 ± 0.07 * | 53.51 ± 0.33 | 3.53 ± 0.31 | 60.44 ± 0.36 ** |
| Line 10 | 1 | 2.66 ± 0.12 | 0.43 ± 0.06 * | 48.98 ± 1.11 | 3.09 ± 0.06 | 55.17 ± 1.00 |
| Line 11 | 1 | 1.42 ± 0.08 | 0.23 ± 0.04 | 49.57 ± 0.46 | 3.28 ± 0.09 | 54.50 ± 0.31 * |
| Line 12 | 1 | 1.35 ± 0.05 | 0.15 ± 0.03 | 51.40 ± 1.75 | 3.26 ± 0.26 | 56.15 ± 1.58 |
| Line 13 | 1 | 2.08 ± 0.11 ** | 0.29 ± 0.08 | 47.43 ± 2.07 | 3.20 ± 0.49 | 53.00 ± 2.07 |
| Line 14 | 1 | 1.97 ± 0.07 ** | 0.14 ± 0.07 | 53.09 ± 0.82 | 2.00 ± 0.13 | 57.19 ± 0.75 |
| Line 15 | 1 | 1.99 ± 0.10 ** | 0.24 ± 0.13 | 52.62 ± 0.70 | 2.93 ± 0.26 | 57.77 ± 0.47 |
| Line 16 | 2 | 1.48 ± 0.04 | 0.28 ± 0.03 | 48.96 ± 1.07 | 3.94 ± 0.10 | 54.66 ± 1.12 |
| Line 17 | 2 | 1.42 ± 0.04 | 0.23 ± 0.04 | 51.18 ± 0.59 | 3.61 ± 0.26 | 56.44 ± 0.41 |
| Average among lines | 2.46 ± 0.26 | 0.37 ± 0.05 | 50.03 ± 0.54 | 3.22 ± 0.11 | 56.08 ± 0.54 | |
| FA | Wild Type | Line 3-1 | Line 4-5 |
|---|---|---|---|
| 16:0 | 2.75 ± 0.08 | 3.51 ± 0.28 | 2.87 ± 0.07 |
| 16:1 | 1.16 ± 0.10 | 1.02 ± 0.09 | 1.00 ± 0.17 |
| 18:0 | 3.42 ± 0.83 | 3.31 ± 0.17 | 3.43 ± 0.20 |
| 18:1 | 15.19 ± 0.04 | 14.62 ± 0.30 | 14.96 ± 0.24 |
| 18:2 | 8.07 ± 0.28 | 8.43 ± 0.35 | 8.78 ± 0.48 |
| 18:3 | 13.90 ± 0.05 | 11.56 ± 0.65 | 11.40 ± 0.64 |
| 20:1 | 0.88 ± 0.12 | 0.37 ± 0.03 * | 0.68 ± 0.21 |
| 18:1OH | 1.43 ± 0.07 | 4.27 ± 0.10 *** | 4.23 ± 0.41 ** |
| 18:2OH | 0.02 ± 0.01 | 0.86 ± 0.14 * | 0.49 ± 0.25 |
| 20:1OH | 50.35 ± 0.59 | 47.82 ± 0.89 | 47.20 ± 0.62 * |
| 20:2OH | 2.63 ± 0.18 | 3.17 ± 0.12 | 3.50 ± 0.18 * |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, G.Q.; Van Erp, H.; Martin-Moreno, J.; Johnson, K.; Morales, E.; Browse, J.; Eastmond, P.J.; Lin, J.-T. Expression of Castor LPAT2 Enhances Ricinoleic Acid Content at the sn-2 Position of Triacylglycerols in Lesquerella Seed. Int. J. Mol. Sci. 2016, 17, 507. https://doi.org/10.3390/ijms17040507
Chen GQ, Van Erp H, Martin-Moreno J, Johnson K, Morales E, Browse J, Eastmond PJ, Lin J-T. Expression of Castor LPAT2 Enhances Ricinoleic Acid Content at the sn-2 Position of Triacylglycerols in Lesquerella Seed. International Journal of Molecular Sciences. 2016; 17(4):507. https://doi.org/10.3390/ijms17040507
Chicago/Turabian StyleChen, Grace Q., Harrie Van Erp, Jose Martin-Moreno, Kumiko Johnson, Eva Morales, John Browse, Peter J. Eastmond, and Jiann-Tsyh Lin. 2016. "Expression of Castor LPAT2 Enhances Ricinoleic Acid Content at the sn-2 Position of Triacylglycerols in Lesquerella Seed" International Journal of Molecular Sciences 17, no. 4: 507. https://doi.org/10.3390/ijms17040507
APA StyleChen, G. Q., Van Erp, H., Martin-Moreno, J., Johnson, K., Morales, E., Browse, J., Eastmond, P. J., & Lin, J.-T. (2016). Expression of Castor LPAT2 Enhances Ricinoleic Acid Content at the sn-2 Position of Triacylglycerols in Lesquerella Seed. International Journal of Molecular Sciences, 17(4), 507. https://doi.org/10.3390/ijms17040507