
Enhanced Autophagy in Polycystic Kidneys of AQP11 Null Mice



Abstract
:1. Introduction
2. Results
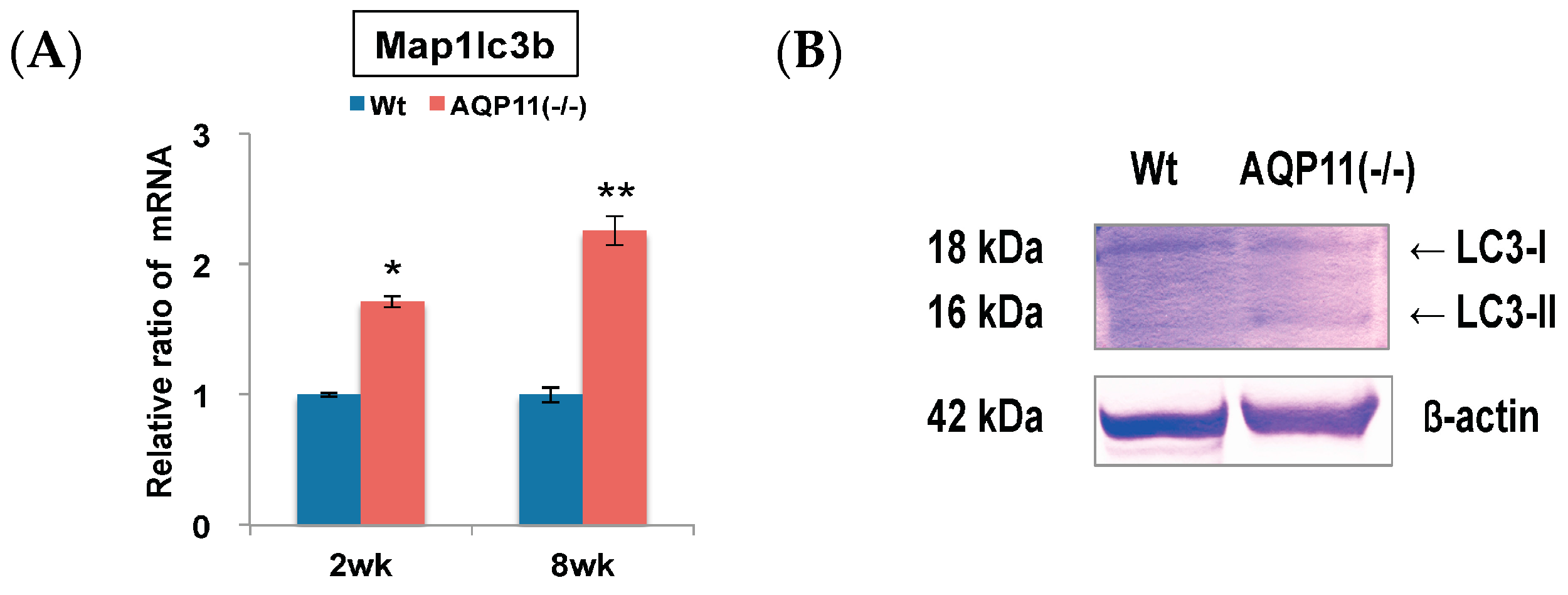
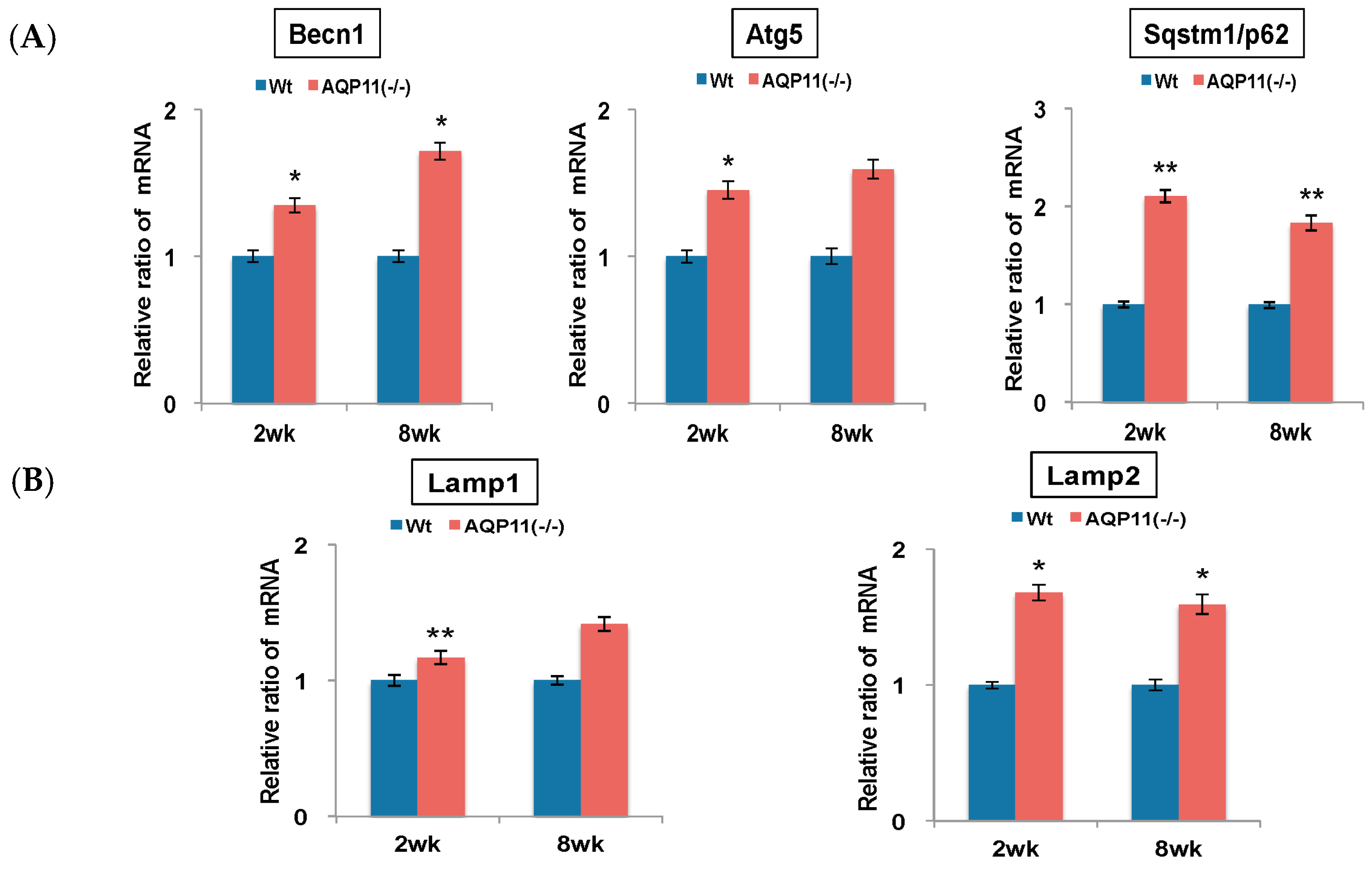
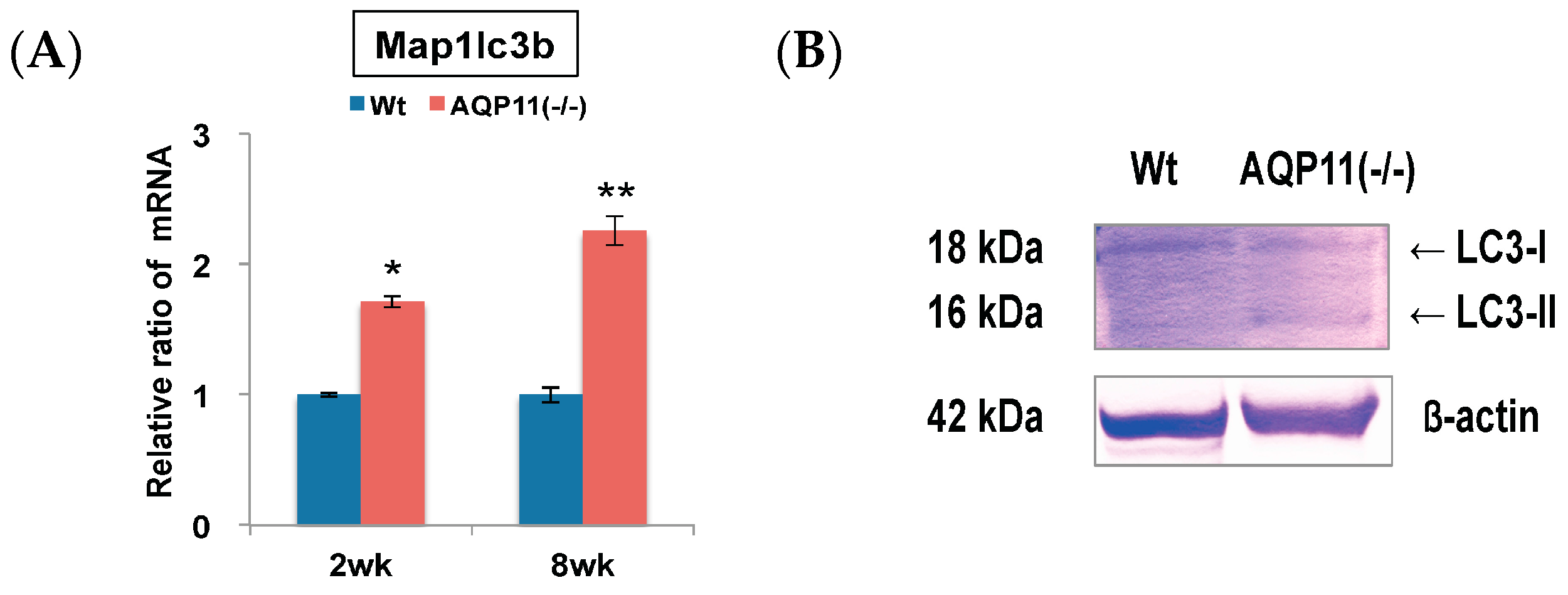
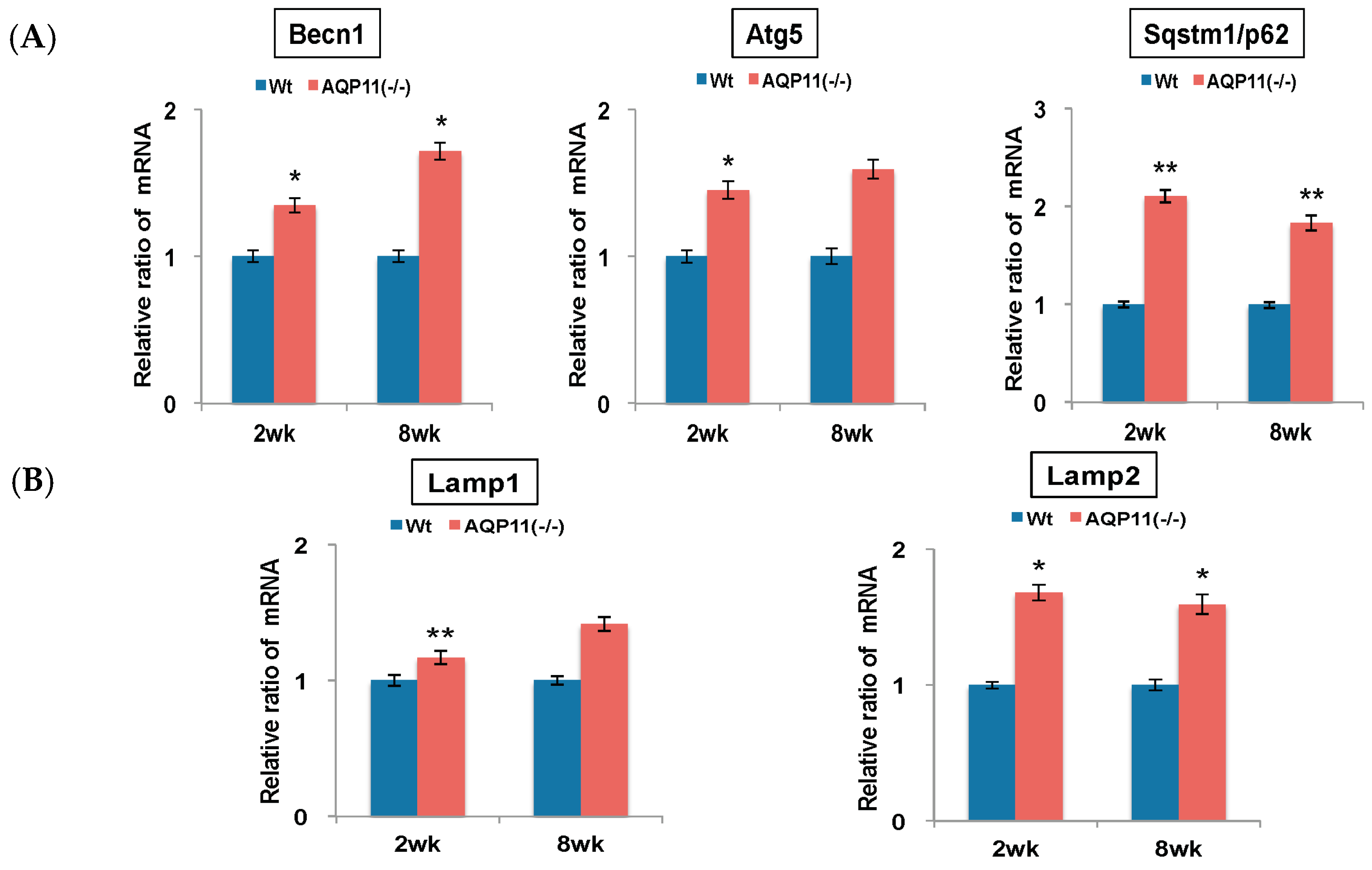
2.1. Autophagy-Related Genes in the AQP11(−/−) Kidney
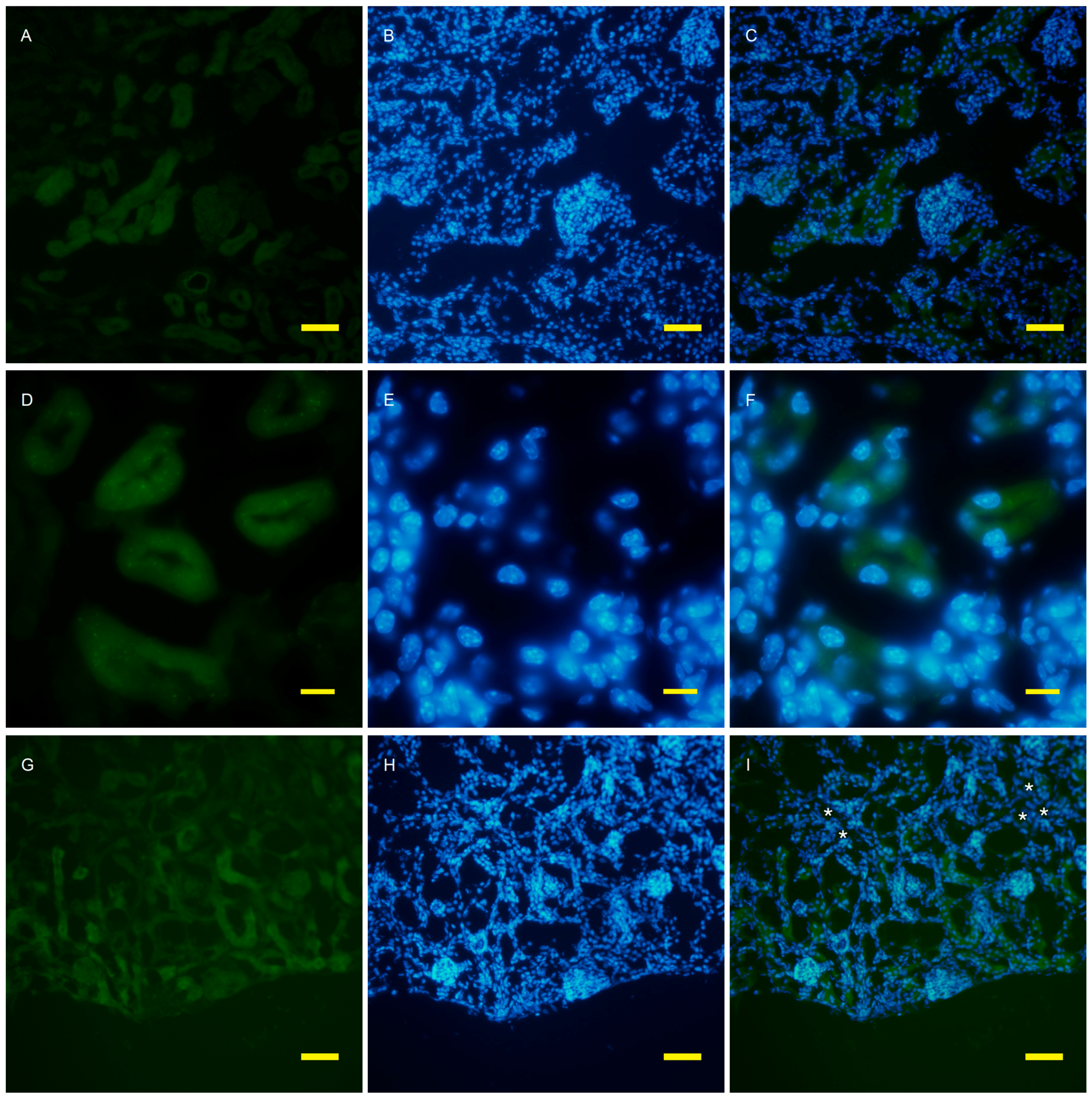
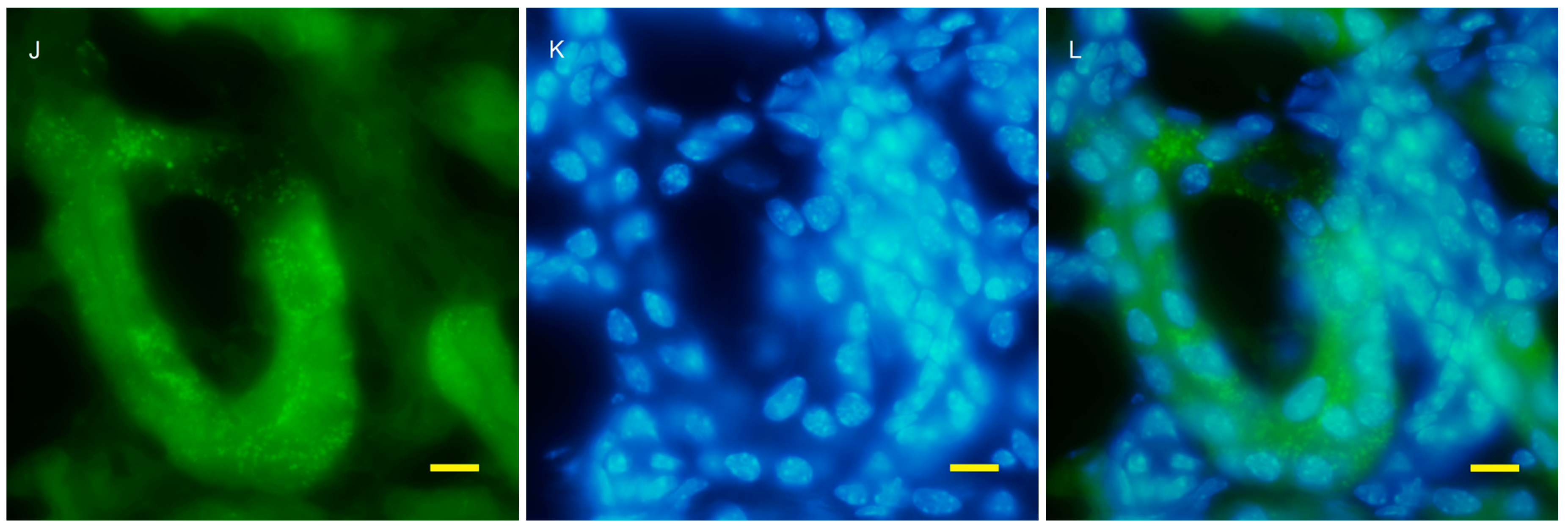
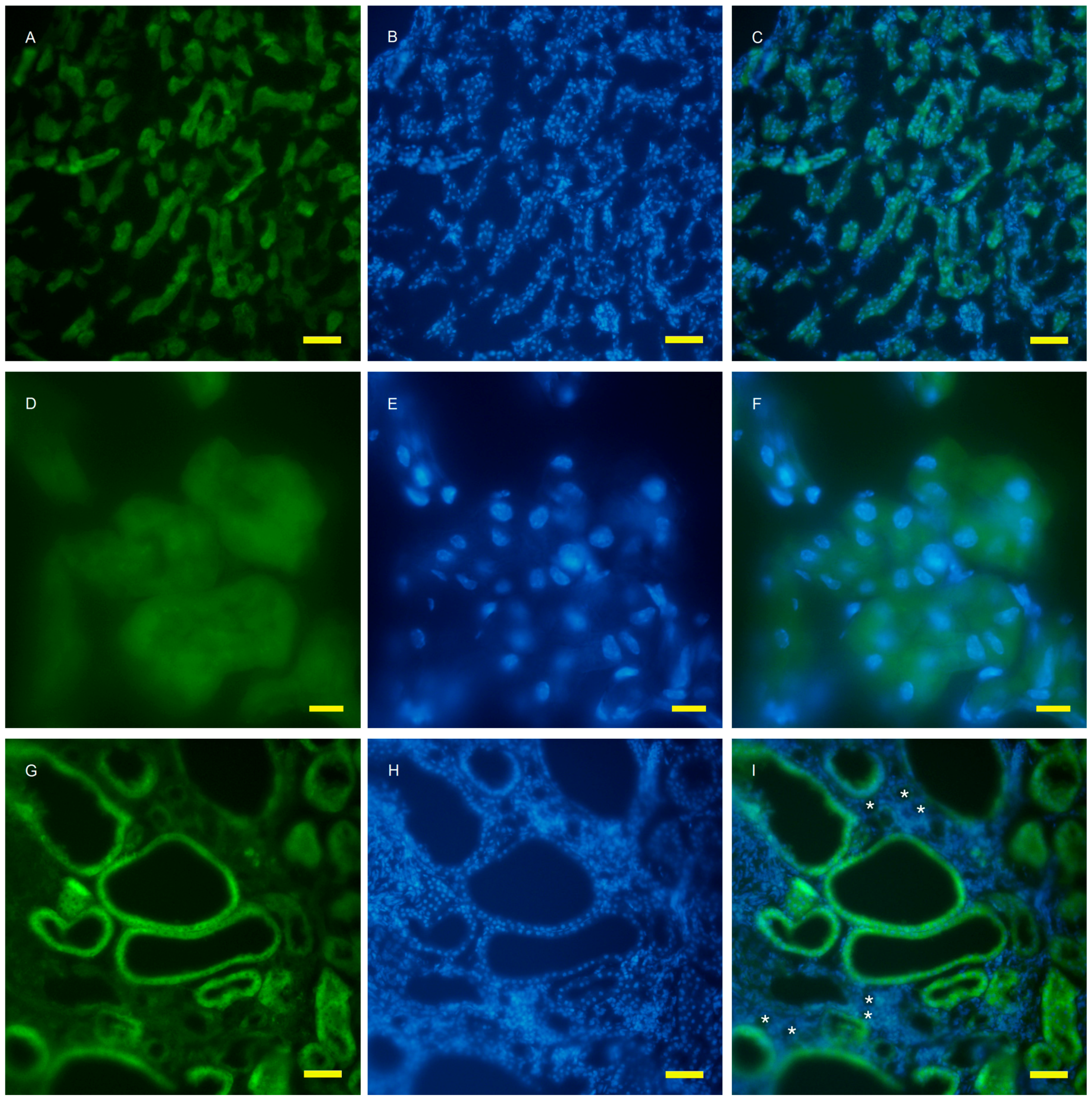
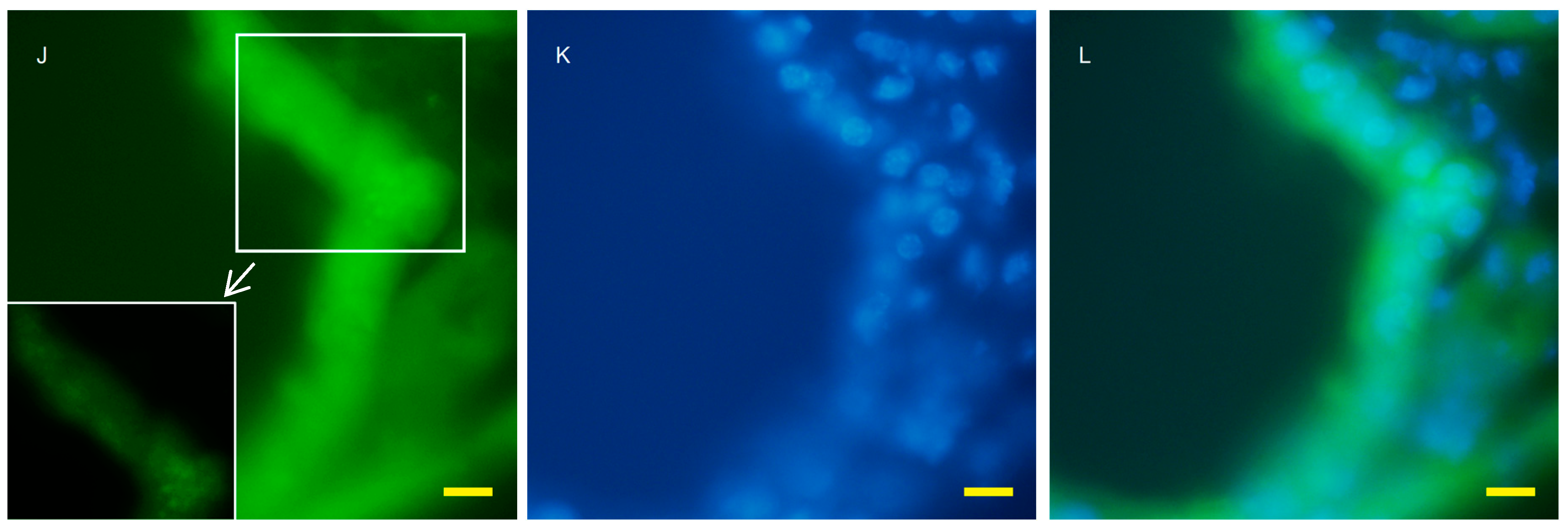
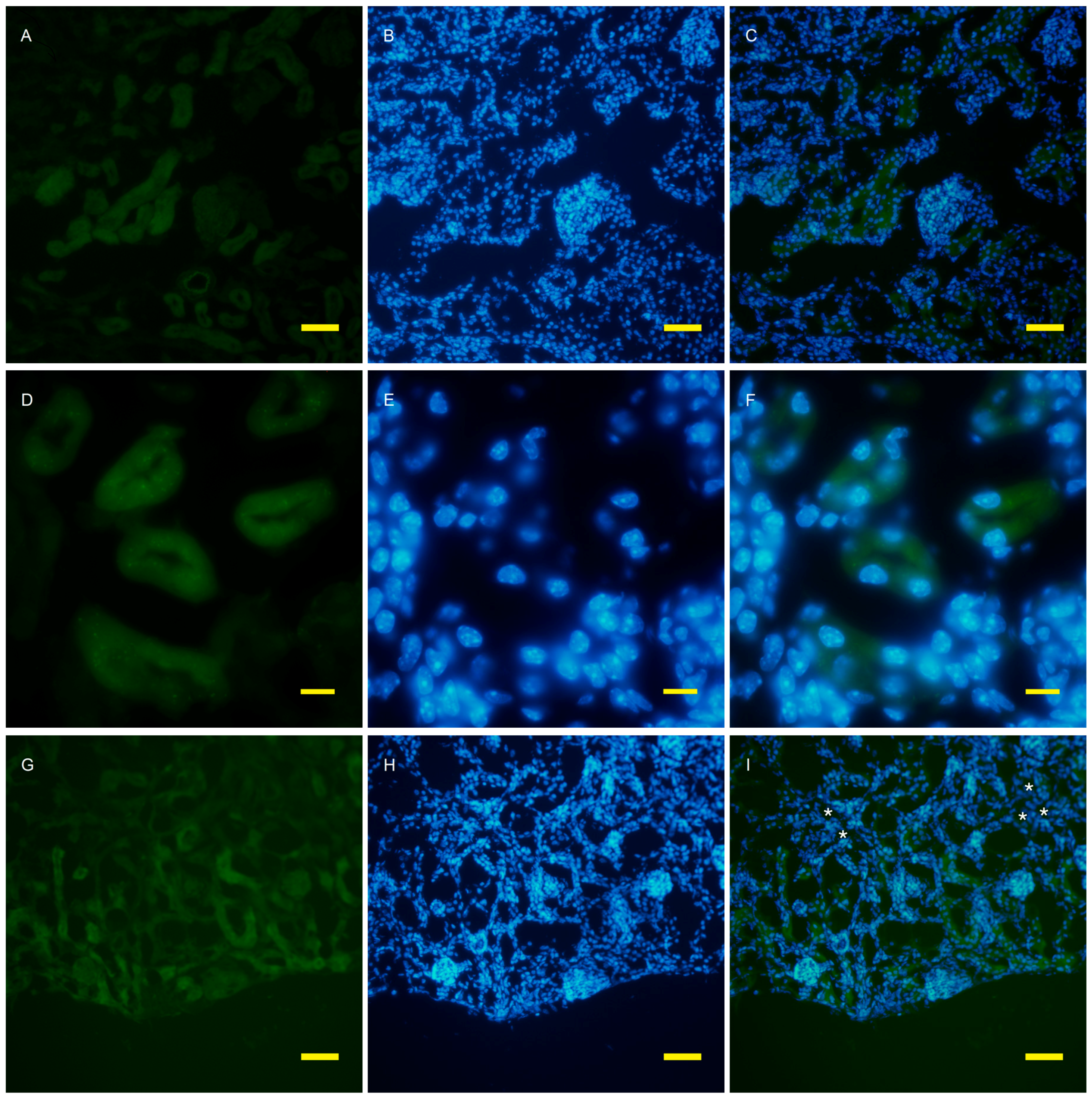
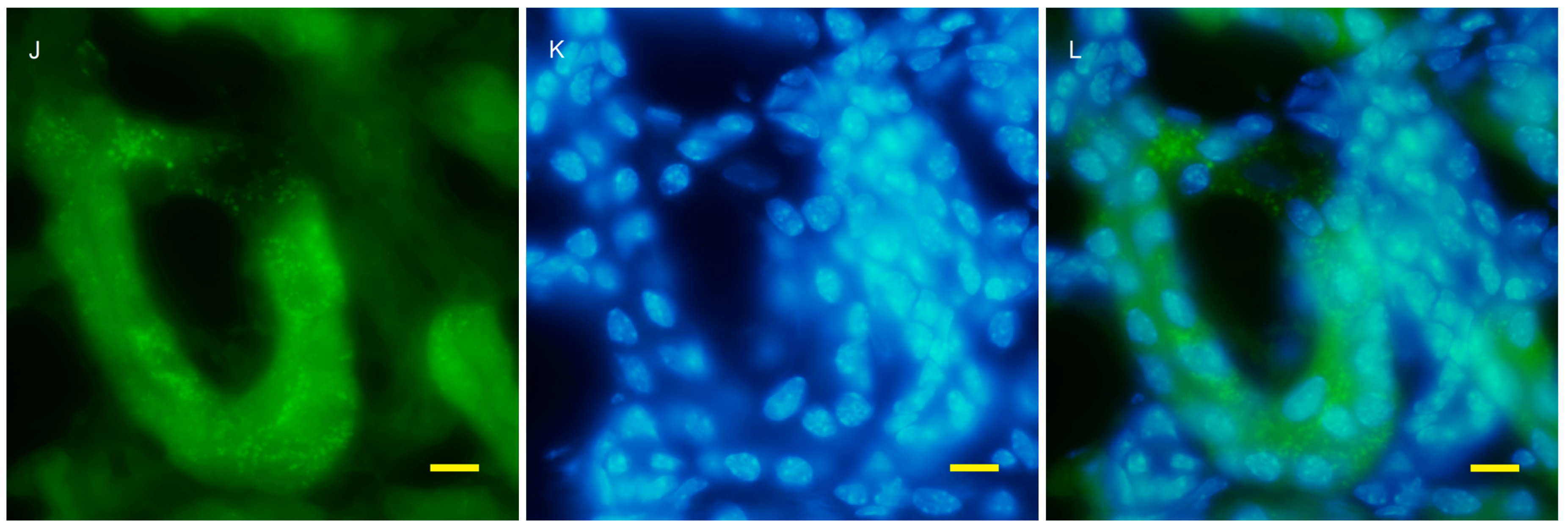
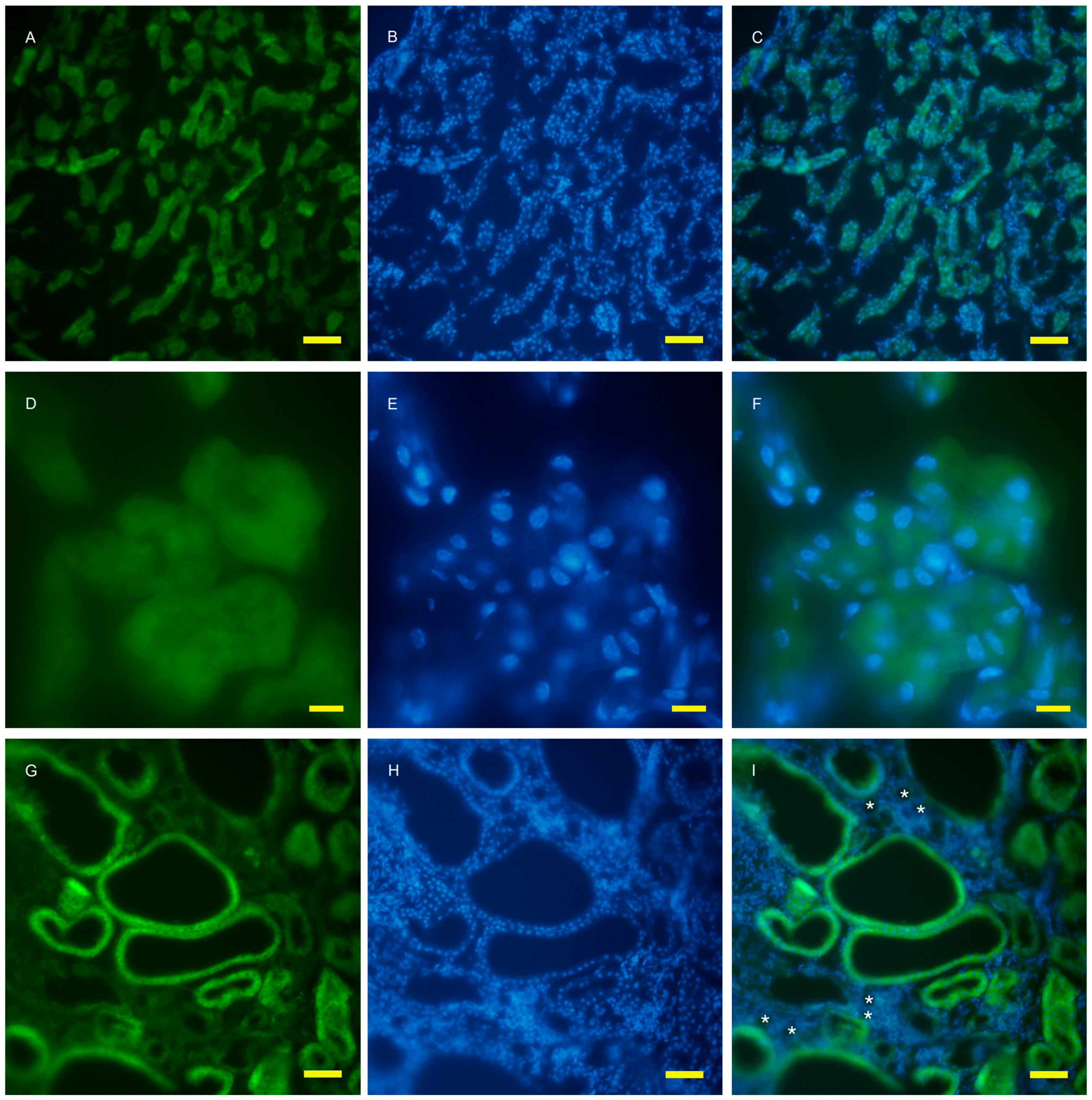
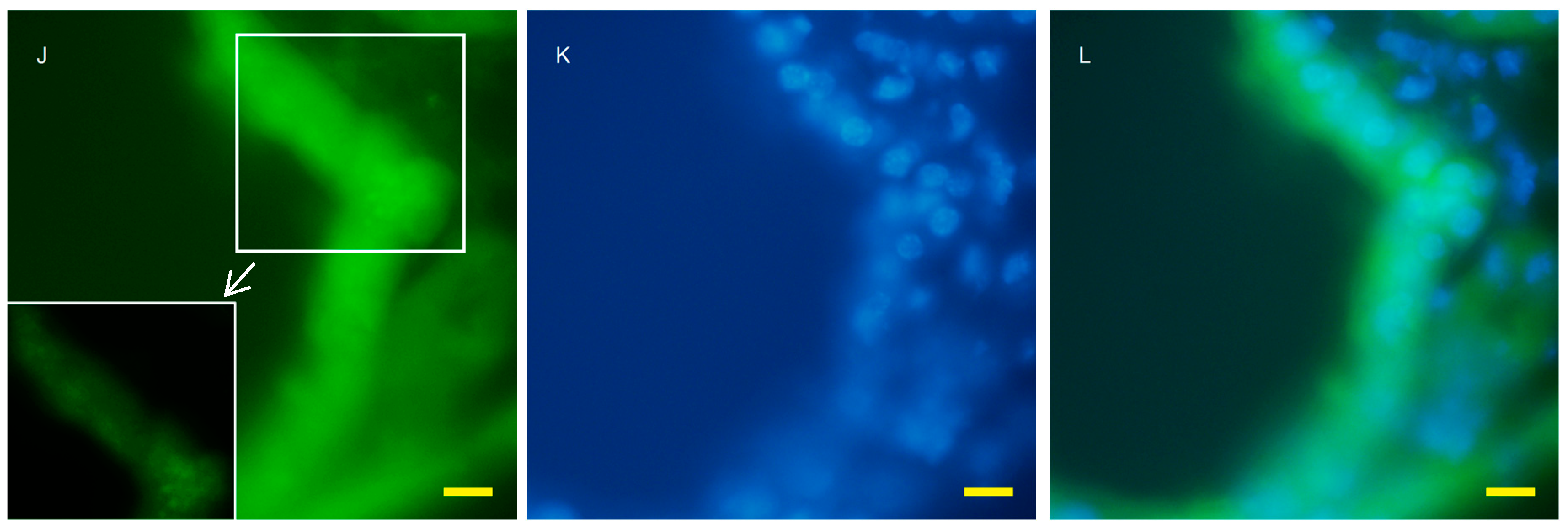
2.2. Autophagy Analysis in the AQP11(−/−) Kidney
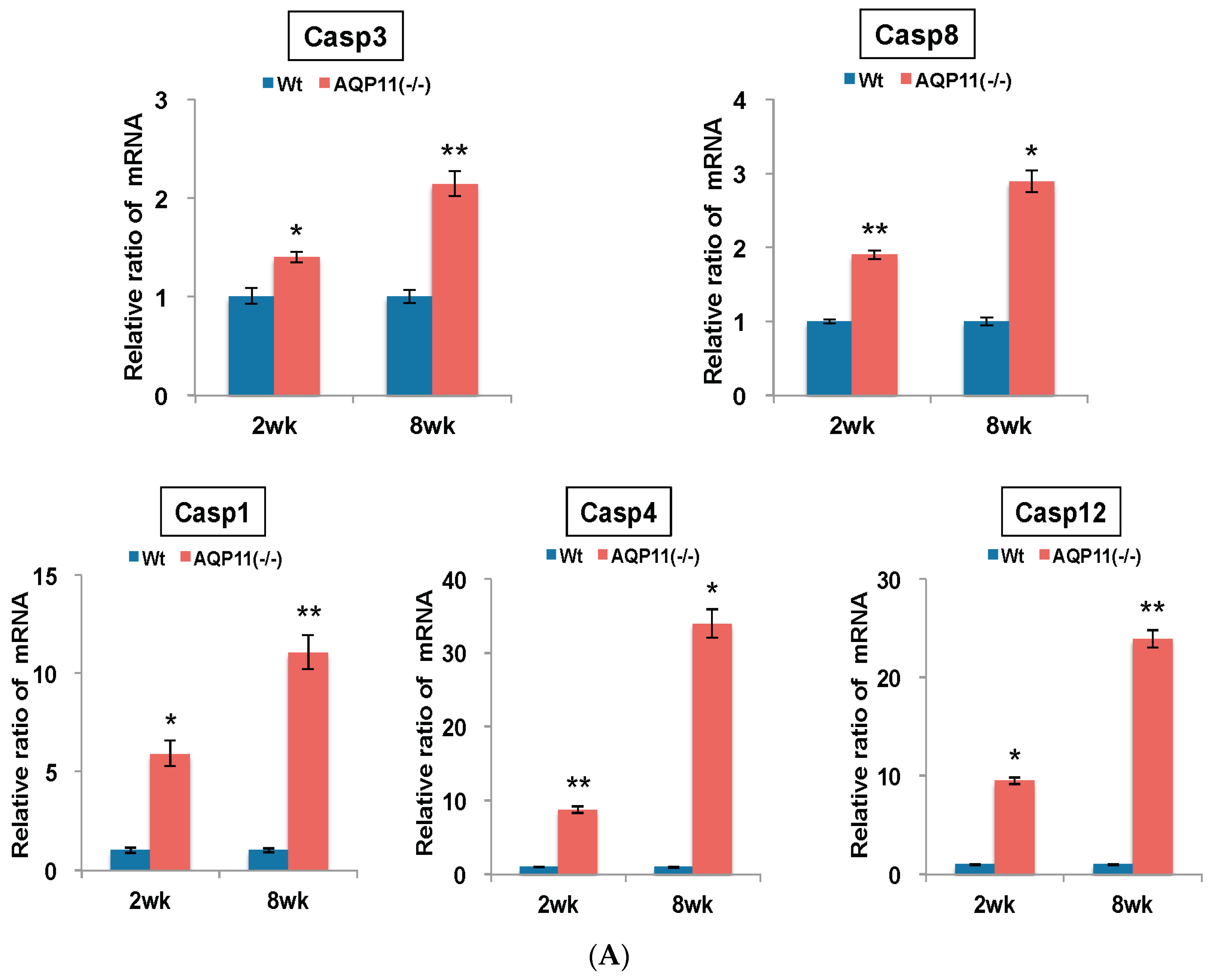
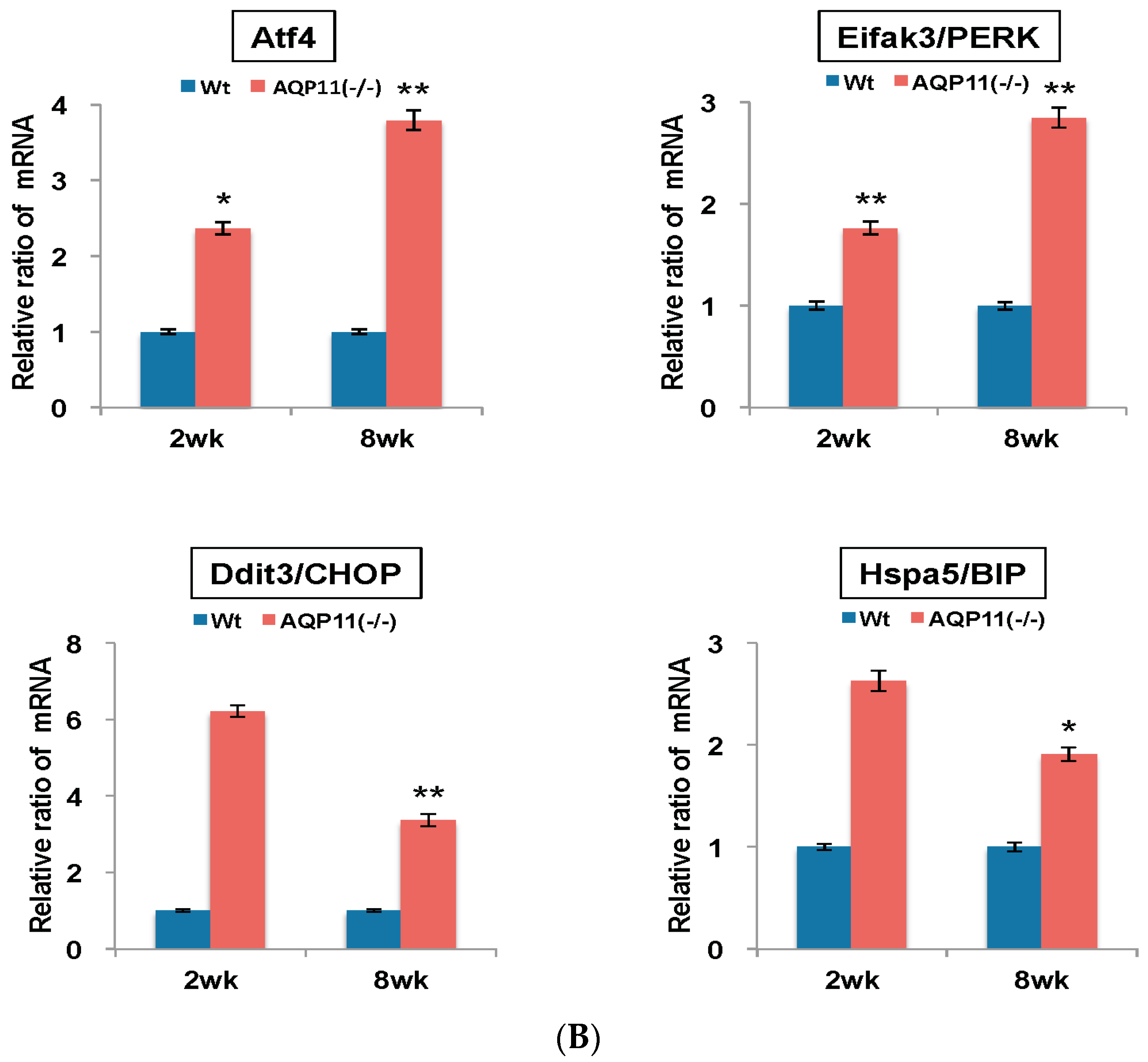
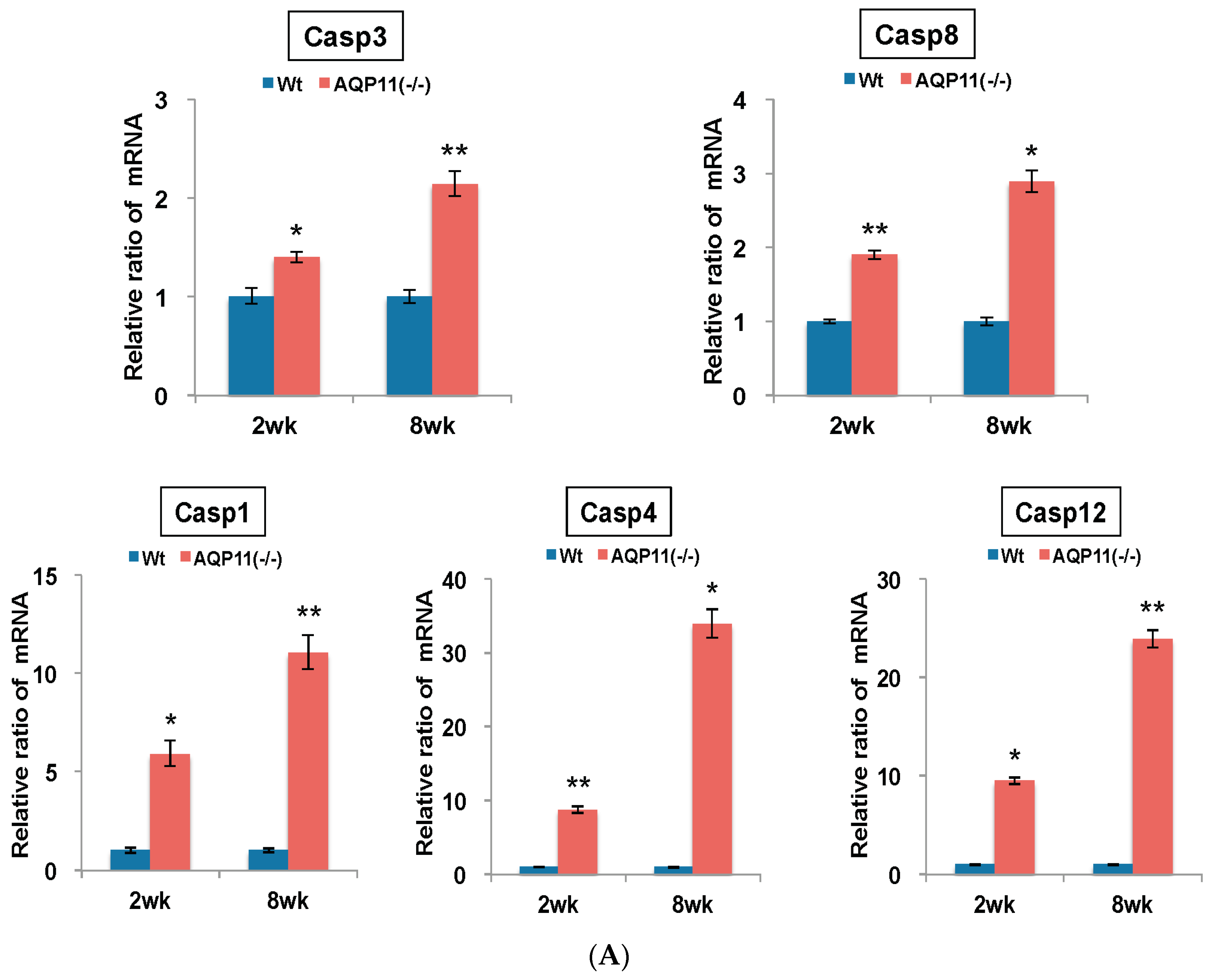
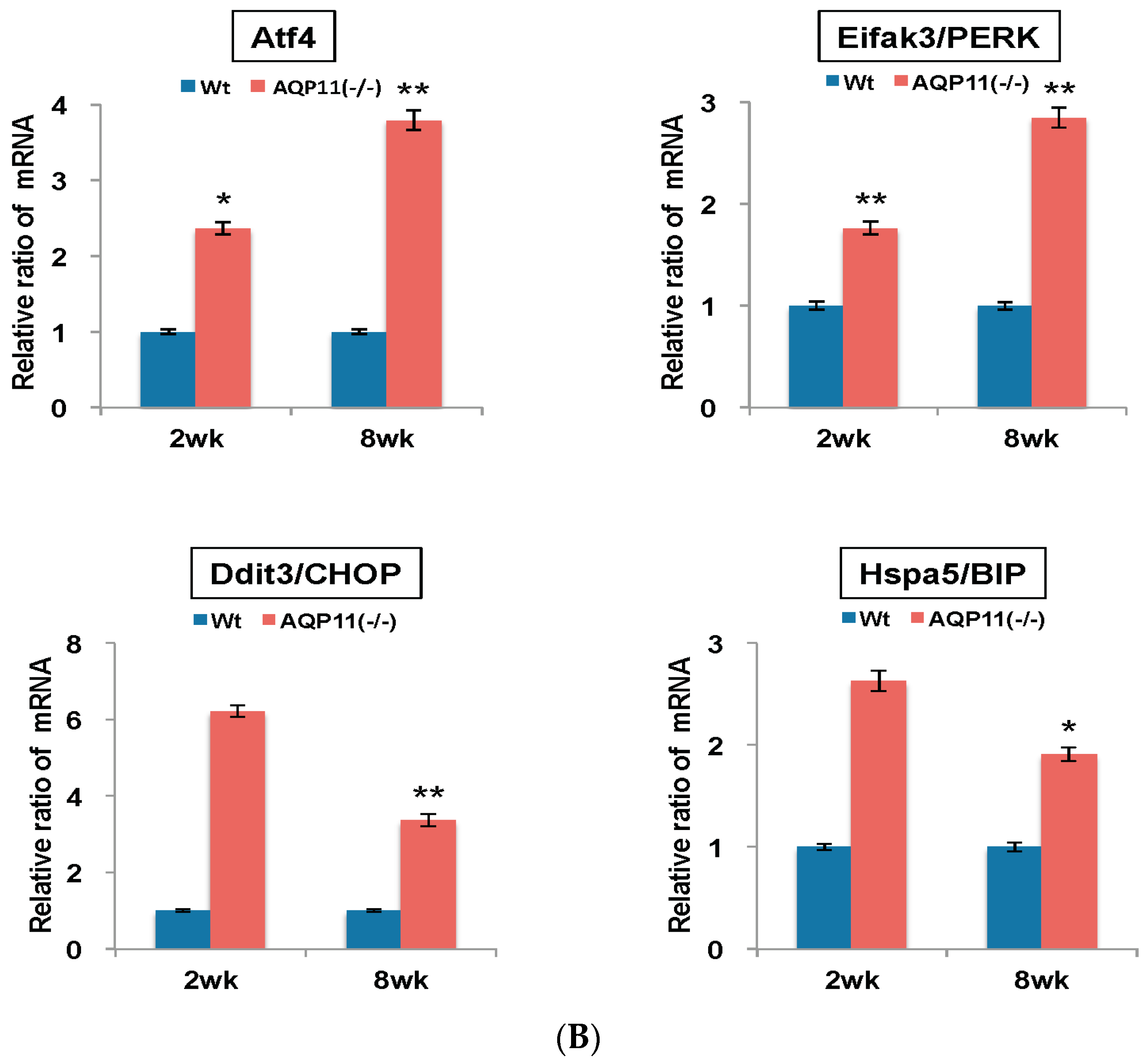
2.3. The Expression of Apoptosis-Related and ER Stress–Related Genes
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Quantitative Real-Time PCR
4.3. Western Blotting
4.4. Fluorescent Microscopy
4.5. Statistical Analysis
4.6. Digital Imaging
Supplementary Materials
Acknowledgments
Author Contributions
Conflict of Interest
References
- Morishita, Y.; Matsuzaki, T.; Hara-chikuma, M.; Andoo, A.; Shimono, M.; Matsuki, A.; Kobayashi, K.; Ikeda, M.; Yamamoto, T.; Verkman, A.; et al. Disruption of aquaporin-11 produces polycystic kidneys following vacuolization of the proximal tubule. Mol. Cell. Biol. 2005, 25, 7770–7779. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, D.A.; Praetorius, J.; Tsunenari, T.; Nielsen, S.; Agre, P. Aquaporin-11: A channel protein lacking apparent transport function expressed in brain. BMC Biochem. 2006, 7, 14. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yakata, K.; Hiroaki, Y.; Ishibashi, K.; Sohara, E.; Sasaki, S.; Mitsuoka, K.; Fujiyoshi, Y. Aquaporin-11 containing a divergent NPA motif has normal water channel activity. Biochim. Biophys. Acta 2007, 1768, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Yakata, K.; Tani, K.; Fujiyoshi, Y. Water permeability and characterization of aquaporin. J. Struct. Biol. 2011, 174, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Madeira, A.; Fernández-Veledo, S.; Camps, M.; Zorzano, A.; Moura, T.F.; Ceperuelo Mallafré, V.; Vendrell, J.; Soveral, G. Human aquaporin-11 is a water and glycerol channel and localizes in the vicinity of lipid droplets in human adipocytes. Obesity 2014, 22, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Sohara, E.; Kobayashi, K.; Chiga, M.; Rai, T.; Ishibashi, K.; Horie, S.; Su, X.; Zhou, J.; Sasaki, S.; et al. Aberrant glycosylation and localization of polycystin-1 cause polycystic kidney in an AQP11 knockout model. J. Am. Soc. Nephrol. 2014, 25, 2789–2799. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Misaka, T.; Tanaka, Y.; Matsumoto, I.; Ishibashi, K.; Sasaki, S.; Abe, K. Aquaporin-11 knockout mice and polycystic kidney disease animals share a common mechanism of cyst formation. FASEB J. 2008, 22, 3672–3684. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.; Pscherer, A.; Roth, C.; Becker, J.; Mücher, G.; Zerres, K.; Dixkens, C.; Weis, J.; Guay-Woodford, L.; Buettner, R.; et al. Enhanced apoptotic cell death of renal epithelial cells in mice lacking transcription factor AP-2beta. Genes Dev. 1997, 11, 1938–1948. [Google Scholar] [CrossRef] [PubMed]
- Trudel, M.; D’Agati, V.; Costantini, F. C-myc as an inducer of polycystic kidney disease in transgenic mice. Kidney Int. 1991, 39, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Veis, D.J.; Sorenson, C.M.; Shutter, J.R.; Korsmeyer, S.J. Bcl-2-deficient mice demonstrate fulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented hair. Cell 1993, 75, 229–240. [Google Scholar] [CrossRef]
- Belibi, F.; Zafar, I.; Ravichandran, K.; Segvic, A.B.; Jani, A.; Ljubanovic, D.G.; Edelstein, C.L. Hypoxia-inducible factor-1α (HIF-1α) and autophagy in polycystic kidney disease (PKD). Am. J. Physiol. Ren. Physiol. 2011, 300, F1235–F1243. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Huber, T.B.; Edelstein, C.L.; Hartleben, B.; Inoki, K.; Jiang, M.; Koya, D.; Kume, S.; Lieberthal, W.; Pallet, N.; Quiroga, A.; et al. Emerging role of autophagy in kidney function, diseases and aging. Autophagy 2012, 8, 1009–1031. [Google Scholar] [CrossRef] [PubMed]
- Periyasamy-Thandavan, S.; Jiang, M.; Wei, Q.; Smith, R.; Yin, X.M.; Dong, Z. Autophagy is cytoprotective during cisplatin injury of renal proximal tubular cells. Kidney Int. 2008, 74, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wei, Q.; Dong, G.; Komatsu, M.; Su, Y.; Dong, Z. Autophagy in proximal tubules protects against acute kidney injury. Kidney Int. 2012, 82, 1271–1283. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Yamamoto, A.; Matsui, M.; Yoshimori, T.; Ohsumi, Y. In vivo analysis of autophagy in response to nutrient starvation using transgenic mice expressing a fluorescent autophagosome marker. Mol. Biol. Cell 2004, 15, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Kuma, A.; Hatano, M.; Matsui, M.; Yamamoto, A.; Nakaya, H.; Yoshimori, T.; Ohsumi, Y.; Tokuhisa, T.; Mizushima, N. The role of autophagy during the early neonatal starvation period. Nature 2004, 432, 1032–1036. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward | Reverse |
|---|---|---|
| Atf4 | 5′-aitgatggcttggccagtg-3′ | 5′-ccattttctccaacatccaatc-3′ |
| Atg5 | 5′-aagtctgtccttccgcagtc-3′ | 5′-tgaagaaagttatctgggtagctca-3′ |
| Becn1 | 5′-aggatggtgtctctcgaagatt-3′ | 5′-gatcagagtgaagctattagcactttc-3′ |
| Casp1 | 5′-cccactgctgatagggtgac-3′ | 5′-gcataggtacataagaatgaactgga-3′ |
| Casp3 | 5′-gaggctgacttcctgtatgctt-3′ | 5′-aaccacgacccgtccttt-3′ |
| Casp4 | 5′-tgtcatctctttgatatattcctgaag-3′ | 5′-caaggttgcccgatcaat-3′ |
| Casp8 | 5′-ttgaacaatgagatccccaaa-3′ | 5′-ccatttctacaaaaatttcaagcag-3′ |
| Casp12 | 5′-gggaattagcacaggcaact-3′ | 5′-ttcttttcttctcagctacagcaa-3′ |
| Ddit3/CHOP | 5′-gcgacagagccagaataaca-3′ | 5′-gatgcacttccttctggaaca-3′ |
| Eifak3/PERK | 5′-ccttggtttcatctagcctca-3′ | 5′-atccagggaggggatgat-3′ |
| GAPDH | 5′-tgtccgtcgtggatctgac-3′ | 5′-cctgcttcaccaccttcttg-3′ |
| Hspa5/BIP | 5′-ctgaggcgtatttgggaaag-3′ | 5′-tcatgacattcagtccagcaa-3′ |
| Lamp1 | 5′-cctacgagactgcgaatggt-3′ | 5′-ccacaagaactgccatttttc-3′ |
| Lamp2 | 5′-aaggtgcaaccttttaatgtgac-3′ | 5′-tgtcatcatccagcgaacac-3′ |
| Map11c3b | 5′-ccccaccaagatcccagt-3′ | 5′-cgctcatgttcacgtggt-3′ |
| Sqstm1/p62 | 5′-agacccctcacaggaaggac-3′ | 5′-catctgggagagggactcaa-3′ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, Y.; Watari, M.; Saito, T.; Morishita, Y.; Ishibashi, K. Enhanced Autophagy in Polycystic Kidneys of AQP11 Null Mice. Int. J. Mol. Sci. 2016, 17, 1993. https://doi.org/10.3390/ijms17121993
Tanaka Y, Watari M, Saito T, Morishita Y, Ishibashi K. Enhanced Autophagy in Polycystic Kidneys of AQP11 Null Mice. International Journal of Molecular Sciences. 2016; 17(12):1993. https://doi.org/10.3390/ijms17121993
Chicago/Turabian StyleTanaka, Yasuko, Mayumi Watari, Tatsuya Saito, Yoshiyuki Morishita, and Kenichi Ishibashi. 2016. "Enhanced Autophagy in Polycystic Kidneys of AQP11 Null Mice" International Journal of Molecular Sciences 17, no. 12: 1993. https://doi.org/10.3390/ijms17121993
APA StyleTanaka, Y., Watari, M., Saito, T., Morishita, Y., & Ishibashi, K. (2016). Enhanced Autophagy in Polycystic Kidneys of AQP11 Null Mice. International Journal of Molecular Sciences, 17(12), 1993. https://doi.org/10.3390/ijms17121993