
Effects of Microcystin-LR Exposure on Spermiogenesis in Nematode Caenorhabditis elegans

Abstract
:1. Introduction
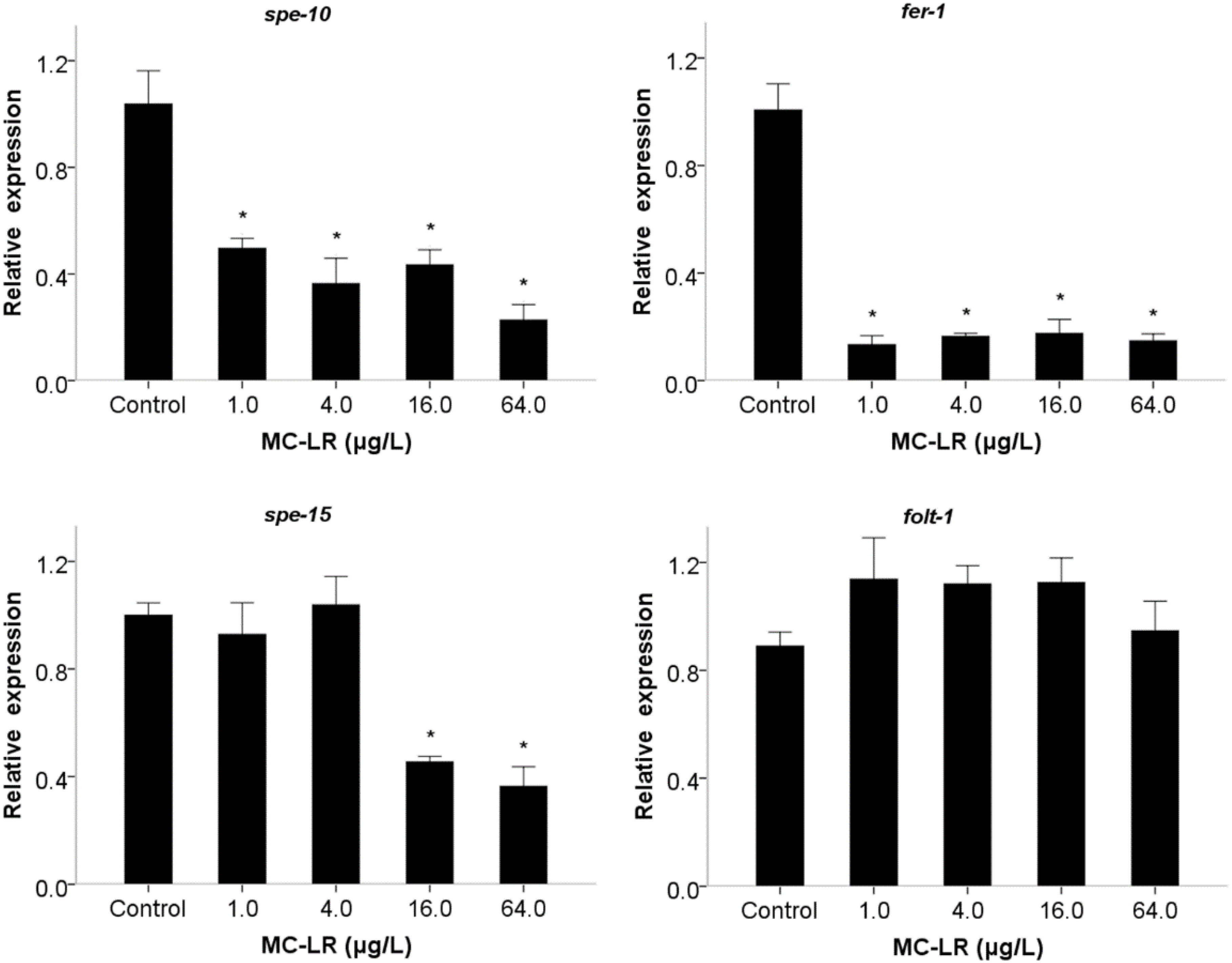
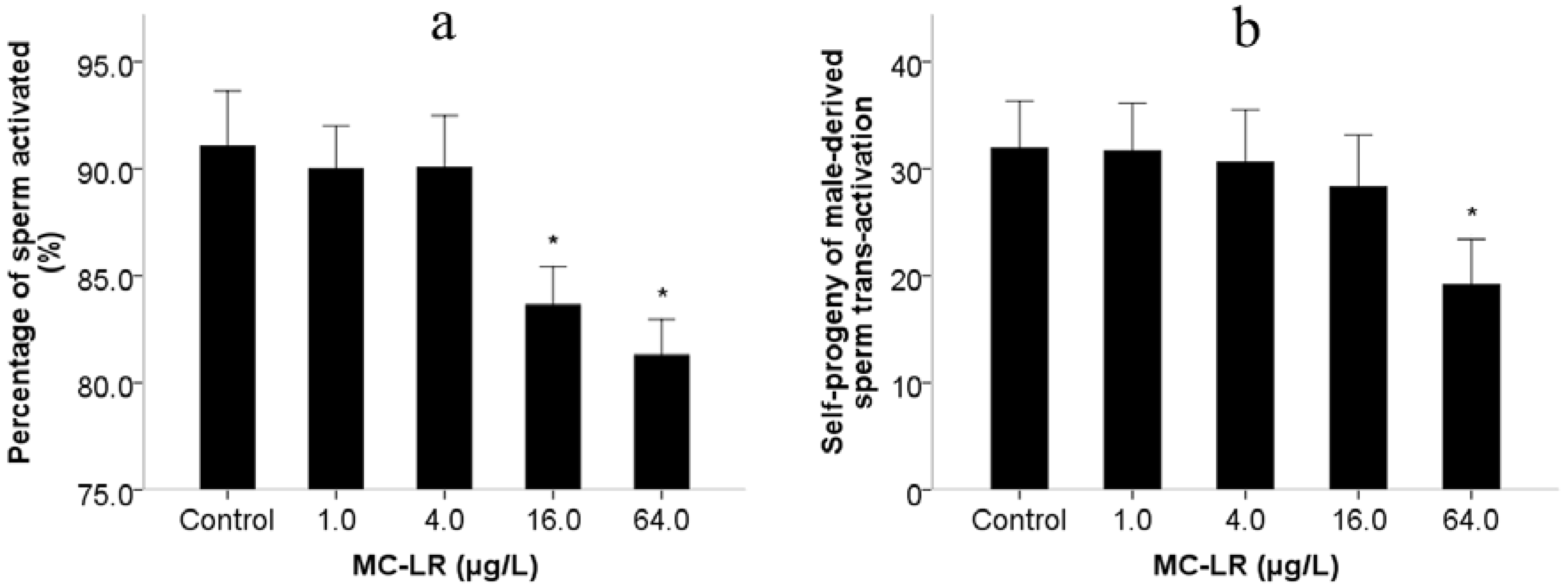
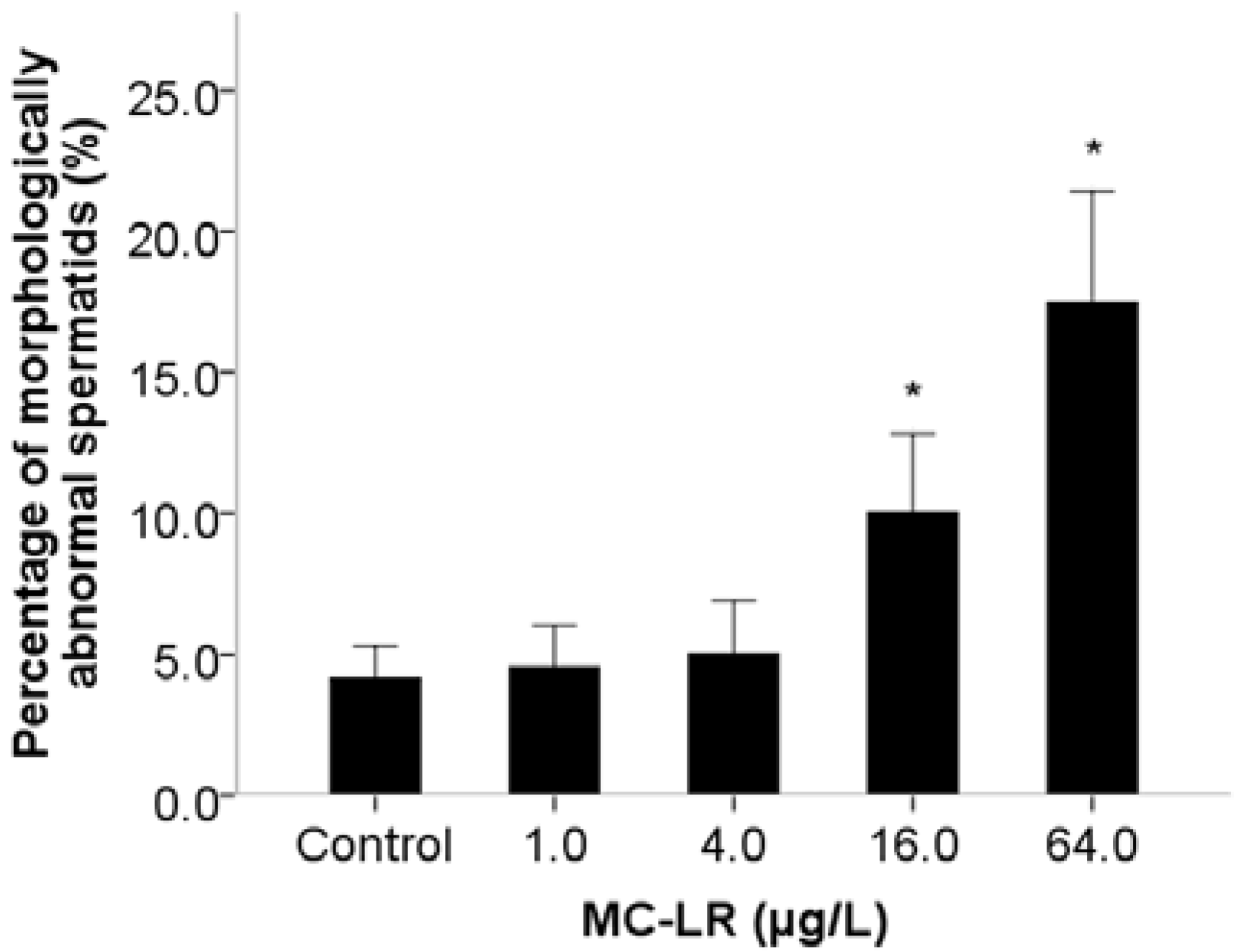
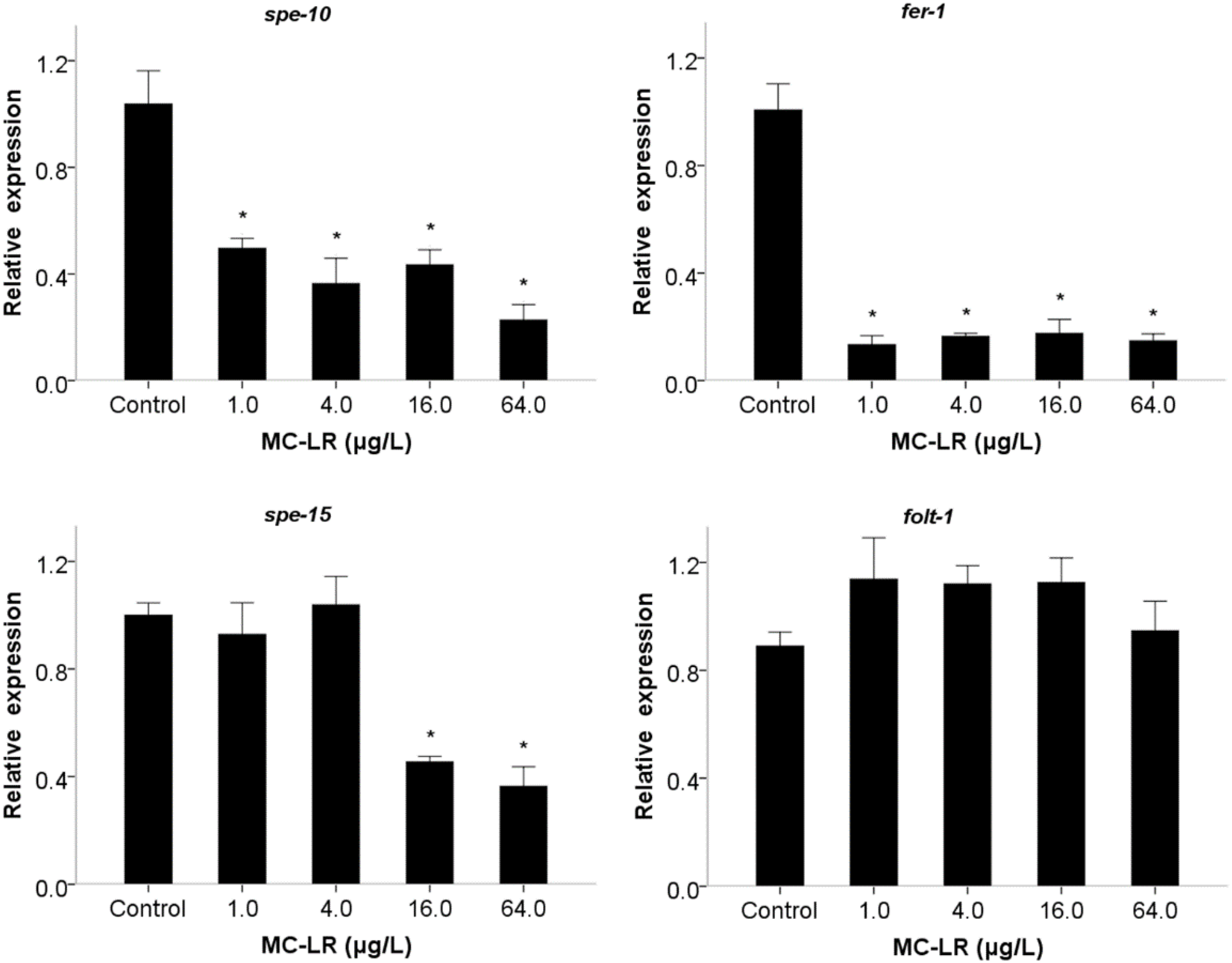
2. Results and Discussion
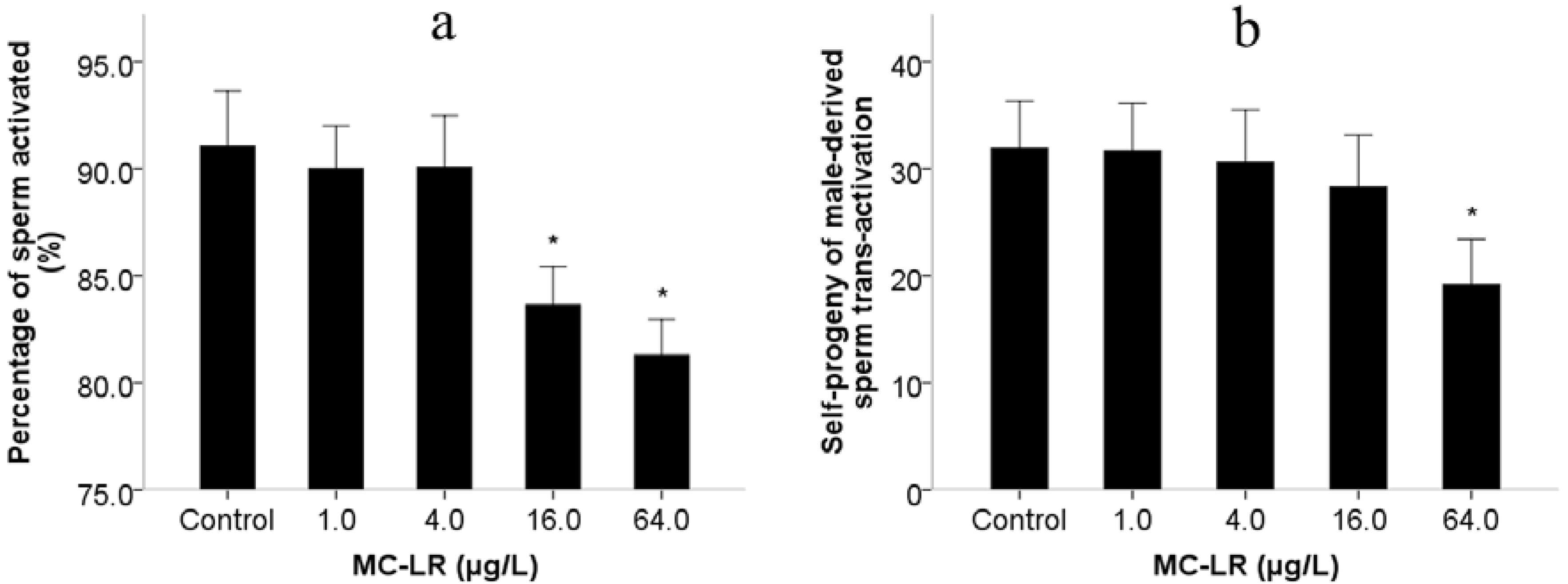
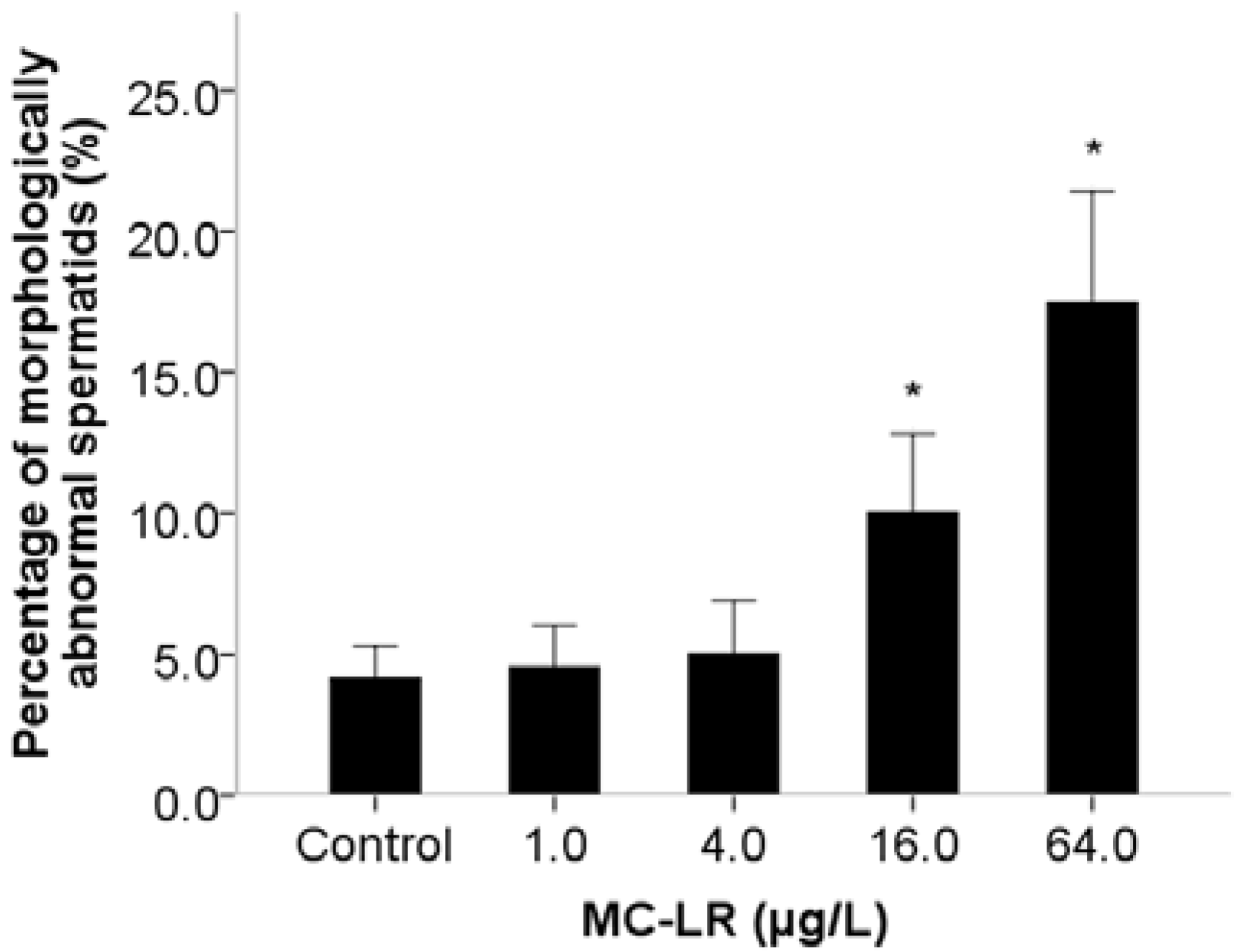

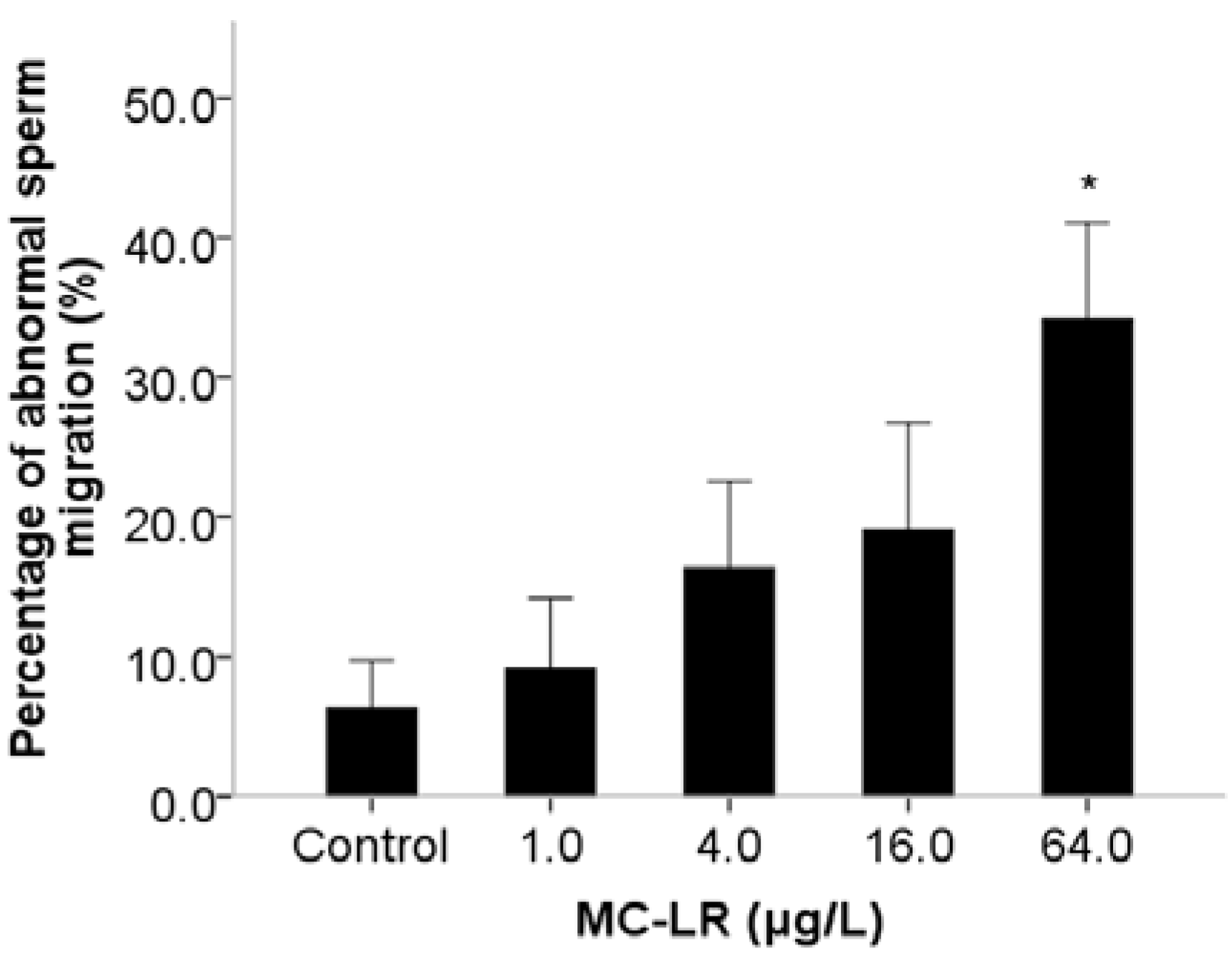
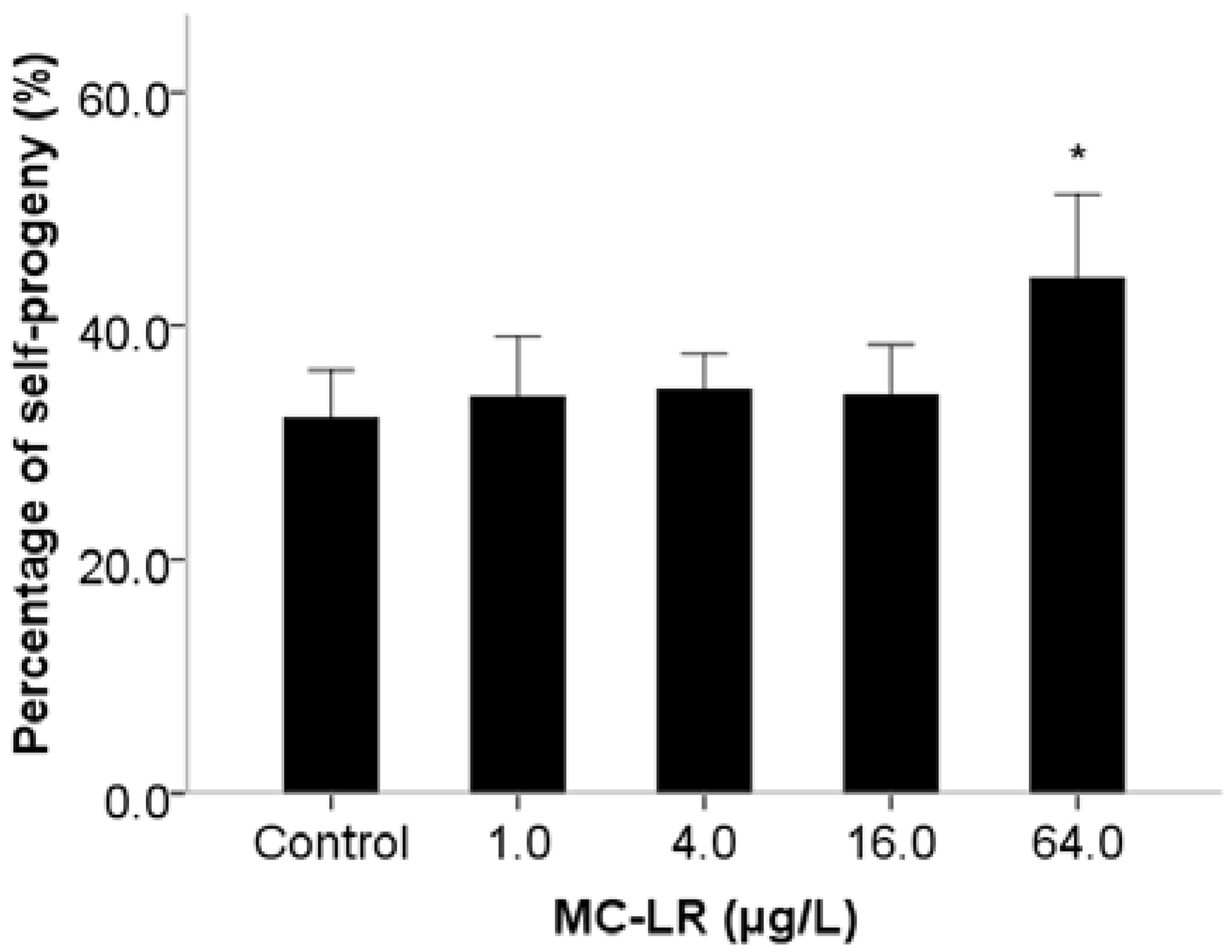
3. Experimental Section

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Primer | Sequence | Annealing Temperature | |
|---|---|---|---|
| spe-10 | Forward | TTTTATTGTCGGCGGAGTGT | 57.8 °C |
| Reverse | CGATGACTGCGAACTTTGAG | ||
| spe-15 | Forward | GGAGTTTTGGATGTCGCTGGTT | 60.4 °C |
| Reverse | GCTCTCTGGGTGAAATGTTGGA | ||
| fer-1 | Forward | AATGGATGGAATGCTGTTGGTC | 57.8 °C |
| Reverse | AACGCTTTCTGAAGTTGTGGTG | ||
| folt-1 | Forward | TCCATTCCTCACTCCGTTTCTA | 60.4 °C |
| Reverse | GCATCTGCCATACTCCTTTACC | ||
| act-1 | Forward | ATGTGTGACGACGAGGTT | 60.4 °C |
| Reverse | GAAGCACTTGCGGTGAAC | ||
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, V.M.; Sivonen, K.; Evans, W.R.; Carmichael, W.W.; Namikoshi, M. Hepatotoxic microcystin diversity in cyanobacterial blooms collected in Portuguese freshwaters. Water Res. 1996, 30, 2377–2384. [Google Scholar] [CrossRef]
- Maidana, M.; Carlis, V.; Galhardi, F.G.; Yunes, J.S.; Geracitano, L.A.; Monserrat, J.M.; Barros, D.M. Effects of microcystins over short- and long-term memory and oxidative stress generation in hippocampus of rats. Chem. Biol. Interact. 2006, 159, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, P.; Chen, J.; Liang, G. Distribution of microcystins in various organs (heart, liver, intestine, gonad, brain, kidney and lung) of Wistar rat via intravenous injection. Toxicon 2008, 52, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sheng, J.; Sha, J.; Han, X. The toxic effects of microcystin-LR on the reproductive system of male rats in vivo and in vitro. Reprod. Toxicol. 2008, 26, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, X. Microcystin-LR causes cytotoxicity effects in rat testicular Sertoli cells. Environ. Toxicol. Pharmacol. 2012, 33, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, Y.; Yuan, M.; Xiang, Z.; Han, X. In vivo study on the effects of microcystin-LR on the apoptosis; proliferation and differentiation of rat testicular spermatogenic cells of male rats injected i.p. with toxins. J. Toxicol. Sci. 2013, 38, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Trinchet, I.; Djediat, C.; Huet, H.; Dao, S.P.; Edery, M. Pathological modifications following sub-chronic exposure of medaka fish (Oryzias latipes) to microcystin-LR. Reprod. Toxicol. 2011, 32, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xiang, Z.; Li, D.; Han, X. Regulation of microcystin-LR-induced toxicity in mouse spermatogonia by miR-96. Environ. Sci. Technol. 2014, 48, 6383–6390. [Google Scholar] [CrossRef] [PubMed]
- Hermo, L.; Pelletier, R.M.; Cyr, D.G.; Smith, C.E. Surfing the wave, cycle, life history, and genes/proteins expressed by testicular germ cells. Part 1: Background to spermatogenesis, spermatogonia, and spermatocytes. Microsc. Res. Tech. 2010, 73, 241–278. [Google Scholar] [CrossRef] [PubMed]
- Chemes, H.E.; Rawe, V.Y. The making of abnormal spermatozoa: Cellular and molecular mechanisms underlying pathological spermiogenesis. Cell Tissue Res. 2010, 341, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.H. Rationale, interpretation, andrology lab corner validation, and uses of sperm function tests. J. Androl. 2000, 21, 10–30. [Google Scholar] [PubMed]
- Cheng, C.Y.; Mruk, D.D. Regulation of spermiogenesis, spermiation and blood-testis barrier dynamics: Novel insights from studies on Eps8 and Arp3. Biochem. J. 2011, 435, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.C.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Huguier, P.; Manier, N.; Meline, C.; Bauda, P.; Pandard, P. Improvement of the Caenorhabditis elegans growth and reproduction test to assess the ecotoxicity of soils and complex matrices. Environ. Toxicol. Chem. 2013, 32, 2100–2108. [Google Scholar] [CrossRef] [PubMed]
- Lesch, B.J.; Page, D.C. Genetics of germ cell development. Nat. Rev. Genet. 2012, 13, 781–794. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.S.; Shakes, D.C. Spermatogenesis. In Germ Cell Development in C. elegans; Springer: New York, NY, USA, 2013; Volume 757, pp. 171–203. [Google Scholar]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discov. 2006, 5, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.; Carrel, J.S. Fertilization and sperm competition in the nematode Caenorhabditis elegans. Dev. Biol. 1979, 73, 304–321. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Ruan, Q.L.; Ju, J.J.; Li, Y.H.; Li, X.B.; Liu, R.; Liang, G.Y.; Zhang, J.; Pu, Y.P.; Wang, D.Y.; Yin, L.H. Chlorpyrifos exposure reduces reproductive capacity owing to a damaging effect on gametogenesis in the nematode Caenorhabditis elegans. J. Appl. Toxicol. 2012, 32, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Allard, P.; Kleinstreuer, N.C.; Knudsen, T.B.; Colaiacovo, M.P.A. C. elegans screening platform for the rapid assessment of chemical disruption of germline function. Environ. Health Perspect. 2013, 121, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Geng, Z.Z.; Wang, Y.; Tong, Z.H.; Yu, H.Q. Essential roles of p53 and MAPK cascades in microcystin-LR-induced germline apoptosis in Caenorhabditis elegans. Environ. Sci. Technol. 2012, 46, 3442–3448. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhao, Y.; Sun, W.; Shimabukuro, K.; Miao, L. Transformation: How do nematode sperm become activated and crawl? Protein Cell 2012, 3, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Shakes, D.C.; Ward, S. Initiation of spermiogenesis in C. elegans: A pharmacological and genetic analysis. Dev. Biol. 1989, 134, 189–200. [Google Scholar] [CrossRef]
- Geldziler, B.; Chatterjee, I.; Singson, A. The genetic and molecular analysis of spe-19, a gene required for sperm activation in Caenorhabditis elegans. Dev. Biol. 2005, 283, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Kubagawa, H.M.; Watts, J.L.; Corrigan, C.; Edmonds, J.W.; Sztul, E.; Browse, J.; Miller, M.A. Oocyte signals derived from polyunsaturated fatty acids control sperm recruitment in vivo. Nat. Cell Biol. 2006, 8, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- LaMunyon, C.W.; Ward, S. Larger sperm outcompete smaller sperm in the nematode Caenorhabditis elegans. Proc. R. Soc. Lond. B: Biol. Sci. 1998, 265, 1997–2002. [Google Scholar] [CrossRef] [PubMed]
- Gleason, E.J.; Lindsey, W.C.; Kroft, T.L.; Singson, A.W.; L’Hernault, S.W. spe-10 encodes a DHHC-CRD zinc-finger membrane protein required for endoplasmic reticulum/Golgi membrane morphogenesis during Caenorhabditis elegans spermatogenesis. Genetics 2006, 172, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, J.F.; Mandell, M.A.; Moulder, G.; Hill, K.L.; L’Hernault, S.W.; Barstead, R.; Titus, M.A. Myosin VI is required for asymmetric segregation of cellular components during C. elegans spermatogenesis. Curr. Biol. 2000, 10, 1489–1496. [Google Scholar] [CrossRef]
- Krajacic, P.; Hermanowski, J.; Lozynska, O.; Khurana, T.S.; Lamitina, T. C. elegans dysferlin homolog fer-1 is expressed in muscle, and fer-1 mutations initiate altered gene expression of muscle enriched genes. Physiol. Genom. 2009, 40, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, K.; Ashokkumar, B.; Moussaif, M.; Sze, J.Y.; Said, H.M. Cloning and functional characterization of a folate transporter from the nematode Caenorhabditis elegans. Am. J. Physiol. Cell Physiol. 2007, 293, C670–C681. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.U.; Liau, W.S.; Balamurugan, K.; Ashokkumar, B.; Said, H.M.; LaMunyon, C.W. Knockout of the folate transporter folt-1 causes germline and somatic defects in C. elegans. BMC Dev. Biol. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.; Ruan, Q.; Li, X.; Liu, R.; Li, Y.; Pu, Y.; Yin, L.; Wang, D. Neurotoxicological evaluation of microcystin-LR exposure at environmental relevant concentrations on nematode Caenorhabditis elegans. Environ. Sci. Pollut. Res. 2013, 20, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.L.; Boyd, W.A.; Williams, P.L. Assessment of sublethal endpoints for toxicity testing with the nematode Caenorhabditis elegans. Environ. Toxicol. Chem. 2001, 20, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Singaravelu, G.; Chatterjee, I.; Marcello, M.R.; Singson, A. Isolation and in vitro activation of Caenorhabditis elegans sperm. Annu. Rev. Cell Dev. Biol. 2007, 23, 405–433. [Google Scholar]
- Achanzar, W.E.; Ward, S. A nematode gene required for sperm vesicle fusion. J. Cell Sci. 1997, 110, 1073–1081. [Google Scholar] [PubMed]
- L’Hernault, S.W.; Roberts, T.M. Cell biology of nematode sperm. Methods Cell Biol. 1995, 48, 273–301. [Google Scholar] [PubMed]
- Singson, A.; Hill, K.L.; L’Hernault, S.W. Sperm competition in the absence of fertilization in Caenorhabditis elegans. Genetics 1999, 152, 201–208. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, M.; Chen, P.; Liu, R.; Liang, G.; Yin, L.; Pu, Y. Effects of Microcystin-LR Exposure on Spermiogenesis in Nematode Caenorhabditis elegans. Int. J. Mol. Sci. 2015, 16, 22927-22937. https://doi.org/10.3390/ijms160922927
Li Y, Zhang M, Chen P, Liu R, Liang G, Yin L, Pu Y. Effects of Microcystin-LR Exposure on Spermiogenesis in Nematode Caenorhabditis elegans. International Journal of Molecular Sciences. 2015; 16(9):22927-22937. https://doi.org/10.3390/ijms160922927
Chicago/Turabian StyleLi, Yunhui, Minhui Zhang, Pan Chen, Ran Liu, Geyu Liang, Lihong Yin, and Yuepu Pu. 2015. "Effects of Microcystin-LR Exposure on Spermiogenesis in Nematode Caenorhabditis elegans" International Journal of Molecular Sciences 16, no. 9: 22927-22937. https://doi.org/10.3390/ijms160922927
APA StyleLi, Y., Zhang, M., Chen, P., Liu, R., Liang, G., Yin, L., & Pu, Y. (2015). Effects of Microcystin-LR Exposure on Spermiogenesis in Nematode Caenorhabditis elegans. International Journal of Molecular Sciences, 16(9), 22927-22937. https://doi.org/10.3390/ijms160922927