
RNA Sequencing Identifies Upregulated Kyphoscoliosis Peptidase and Phosphatidic Acid Signaling Pathways in Muscle Hypertrophy Generated by Transgenic Expression of Myostatin Propeptide

Abstract
:
1. Introduction
2. Results
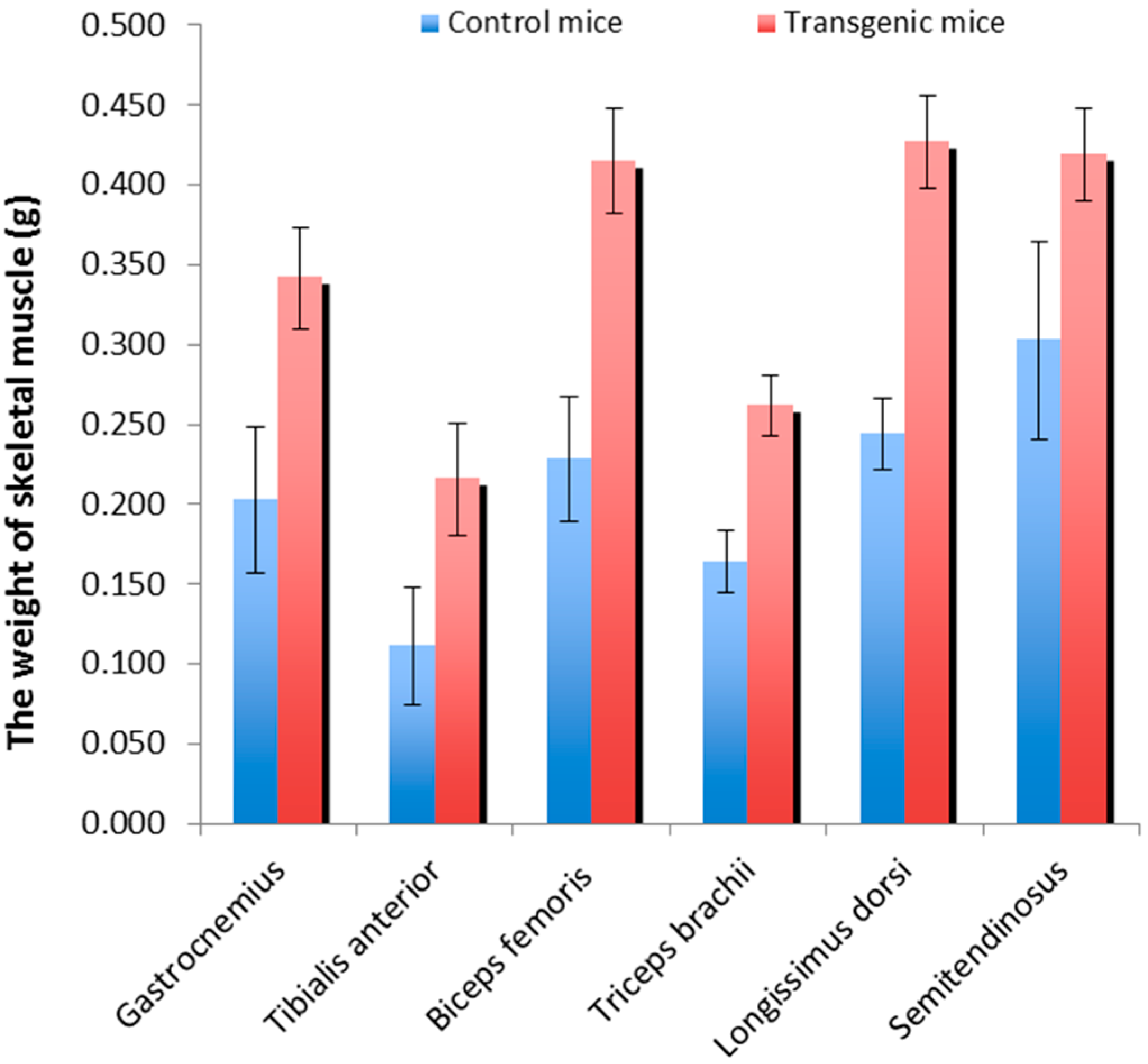
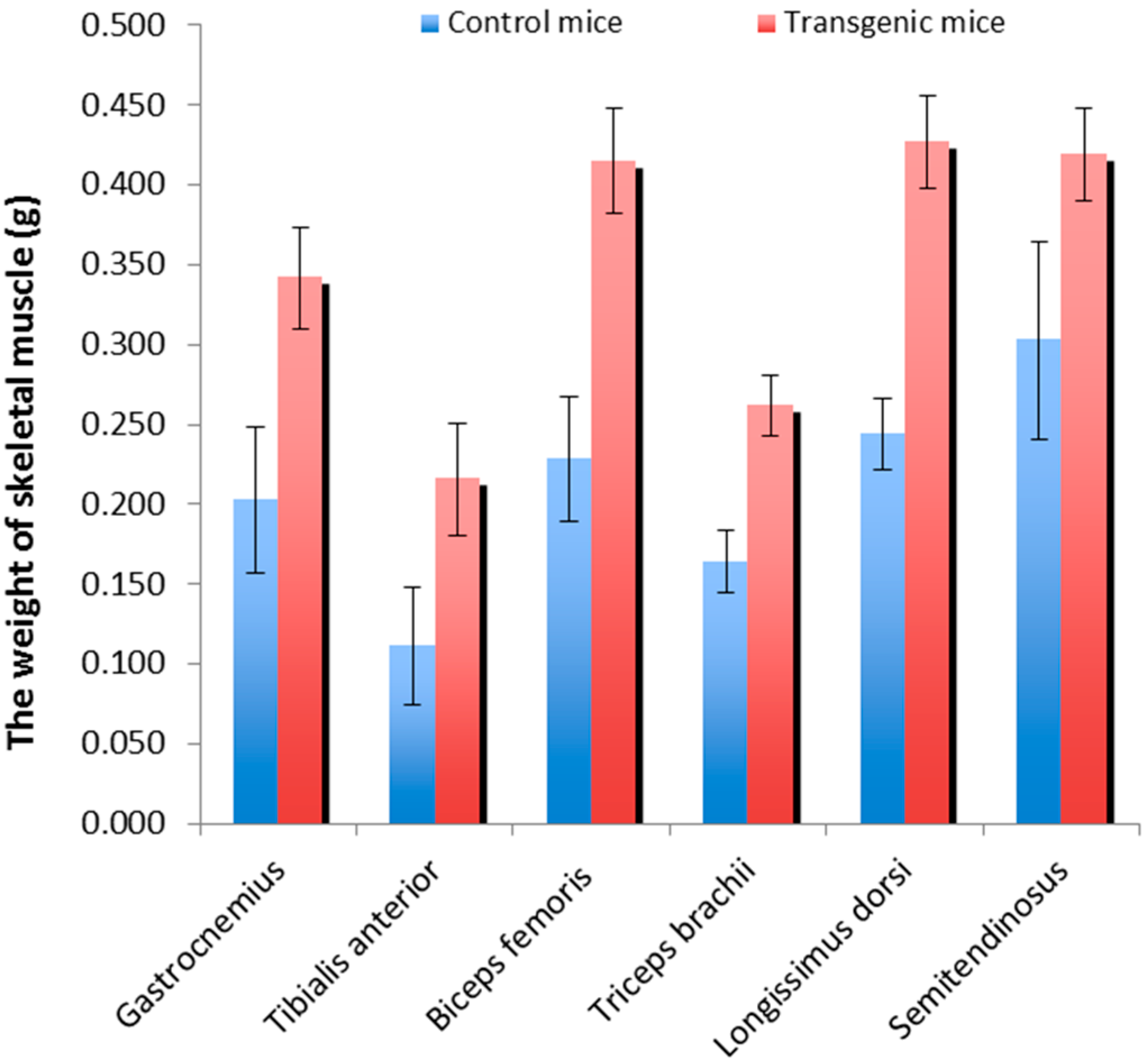
2.1. Skeletal Muscle Weights of the Transgenic Mice and Wild-Type Littermate Controls
2.2. Solexa Sequencing Data and Reads Assembly Using Tophat Software

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CN148 | CN164 | TN126 | TN135 | TN329 | |
|---|---|---|---|---|---|
| Raw reads | 9,710,772 | 10,371,693 | 9,991,395 | 10,258,517 | 10,076,991 |
| Clean reads | 8,717,967 | 9,418,459 | 9,054,439 | 9,301,124 | 9,032,266 |
| Ratio | 89.78% | 90.81% | 90.62% | 90.67% | 89.63% |
| CN148 | CN164 | TN126 | TN135 | TN329 | |
|---|---|---|---|---|---|
| Input | 8,717,967 | 9,418,459 | 9,054,439 | 9,301,124 | 9,032,266 |
| Output | 8,717,523 | 9,418,247 | 9,054,198 | 9,300,876 | 9,031,776 |
| Qualified | 99.99% | 99.99% | 99.99% | 99.99% | 99.99% |
| Mapped | 8,583,223 | 9,251,338 | 8,865,832 | 9,118,889 | 8,890,164 |
| Mapped% | 98.5% | 98.2% | 97.9% | 98% | 98.4% |
| Multiple alignments | 2,249,697 | 1,869,472 | 1,801,533 | 1,789,715 | 2,407,543 |
| Multiple alignments% | 26.2% | 20.2% | 20.3% | 19.6% | 27.1% |
| Min read length | 30 | 30 | 30 | 30 | 30 |
| Max read length | 50 | 50 | 50 | 50 | 50 |
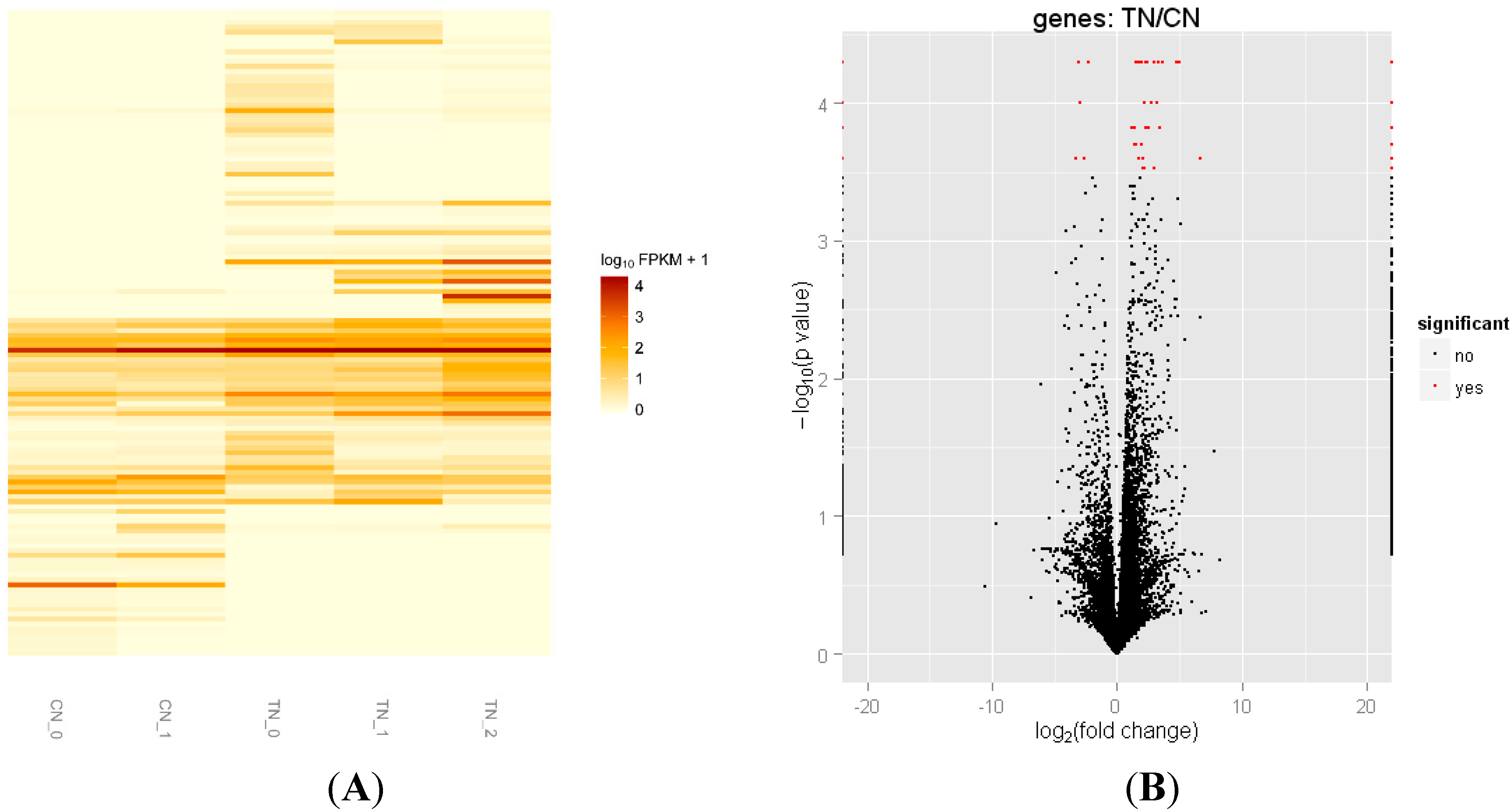
2.3. Differentially Expressed Genes of Skeletal Muscle between Transgenic Mice and Their Littermate Controls
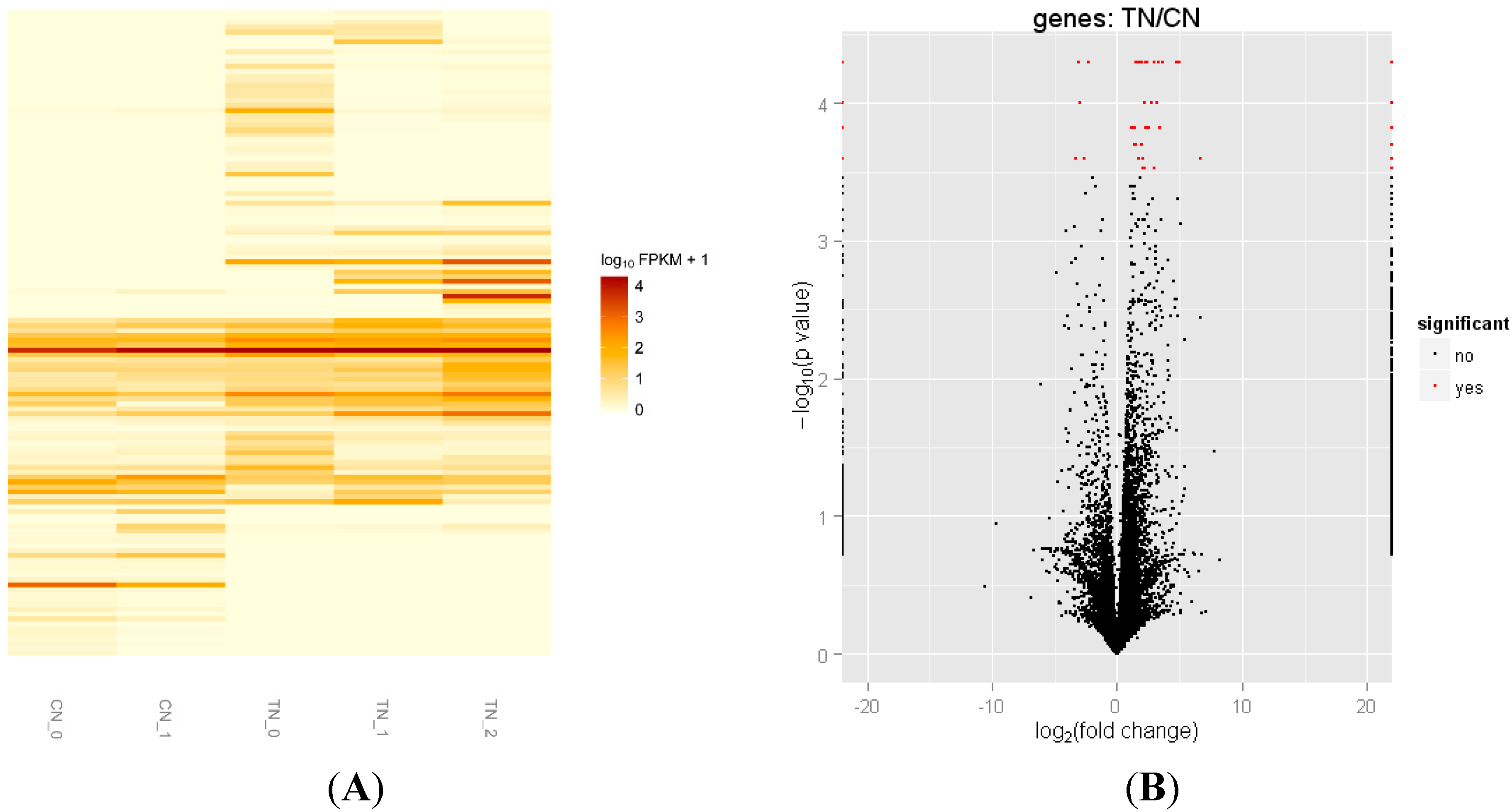
| Gene ID | Gene | Official Full Name | CN Value | TN Value | Fold Change | p-Value |
|---|---|---|---|---|---|---|
| Genes Related to Cytoskeleton and Muscle Development | ||||||
| XLOC_010673 | Myh6 | Myosin heavy polypeptide 6 | 5.49 | 0.70 | −7.83 | 0.00010 |
| XLOC_000666 | Mybph | Myosin binding protein H | 14.77 | 57.60 | 3.90 | 0.00005 |
| XLOC_003363 | Rmst | Rhabdomyosarcoma 2 associated transcript | 0.53 | 15.11 | 28.72 | 0.00005 |
| XLOC_006460 | Actg1 | Actin gamma cytoplasmic 1 | 11.26 | 355.42 | 31.56 | 0.00005 |
| XLOC_019664 | Actc1 | Actin alpha cardiac muscle 1 | 50.30 | 257.64 | 5.12 | 0.00005 |
| XLOC_034484 | Ky | Kyphoscoliosis peptidase (CD1) | 3.26 | 8.81 | 2.70 | 0.00015 |
| XLOC_026810 | Actb | Actin beta | 26.24 | 67.11 | 2.56 | 0.00020 |
| XLOC_000205 | Mstn-pro | Myostatin propeptide | 32.03 | 409.35 | 12.78 | 0.00005 |
| Genes Related to Lipoprotein Metabolic Process and Intracellular Signaling Cascade | ||||||
| XLOC_034105 | Apoa1 | Apolipoprotein A-I | 11.04 | 1.76 | −6.27 | 0.00025 |
| XLOC_019500 | Dgkz | Diacylglycerol kinase zeta | 22.14 | 90.10 | 4.07 | 0.00025 |
| XLOC_000353 | Plcd4 | Phospholipase C delta 4 | 3.76 | 17.38 | 4.63 | 0.00010 |
| XLOC_025320 | Alb | Albumin | 83.44 | 9.80 | −8.51 | 0.00005 |
| XLOC_002012 | G0s2 | G0/G1 switch gene 2 | 4.35 | 23.62 | 5.42 | 0.00005 |
| Genes for Immune Response | ||||||
| XLOC_007714 | Ighm | Immunoglobulin heavy constant mu | 27.24 | 73.44 | 2.70 | 0.00020 |
| XLOC_007114 | Tnfaip2 | Tumor necrosis factor alpha-induced protein 2 | 3.90 | 17.93 | 4.60 | 0.00010 |
| XLOC_031749 | Trim12c | Tripartite motif-containing 12C | 0.34 | 34.88 | 103.84 | 0.00025 |
| Genes for Oxidation Reduction | ||||||
| XLOC_006482 | Cbr2 | Carbonyl reductase 2 | 4.44 | 41.62 | 9.37 | 0.00010 |
| XLOC_010336 | Mss51 | Mitochondrial translational activator | 77.38 | 16.04 | −4.83 | 0.00005 |
| XLOC_020605 | Kcnab1 | Potassium voltage-gated channel shaker-related Subfamily beta member 1 | 2.86 | 11.06 | 3.87 | 0.00020 |
| Genes for Cell Proliferation | ||||||
| XLOC_001651 | Ptprc | Protein tyrosine phosphatase receptor type C (CD45) | 0.81 | 7.90 | 9.77 | 0.00005 |
| XLOC_026557 | Gm13841 | Predicted gene 13841 (ribosomal protein L29) | 0.00 | 8.32 | inf | 0.00005 |
| Genes of Proteolysis | ||||||
| XLOC_024383 | Zmpste24 | Zinc metallopeptidase STE24 | 5.67 | 27.67 | 4.88 | 0.00015 |
| Genes of Carbohydrate Phosphatase Activity | ||||||
| XLOC_018711 | Pfkfb3 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 | 8.83 | 30.52 | 3.46 | 0.00005 |
| Genes of Integral to Membrane, Intrinsic to Membrane | ||||||
| XLOC_015439 | Aqp4 | Aquaporin 4 | 2.40 | 6.81 | 2.83 | 0.00020 |
| Genes of Extracellular Region | ||||||
| XLOC_025292 | Csn1s2a | casein alpha s2-like A | 0.00 | 8.54 | inf | 0.00005 |
| Others | ||||||
| XLOC_029580 | Josd2 | Josephin domain containing 2 | 109.35 | 22.64 | −4.83 | 0.00005 |
| XLOC_025924 | Gm15459 | Predicted gene 15459 (heat shock protein 8 pseudogene) | 7.46 | 21.77 | 2.92 | 0.00005 |
| XLOC_003234 | Odf3l2 | Outer dense fiber of sperm tails 3-like 2 | 14.39 | 57.11 | 3.97 | 0.00005 |
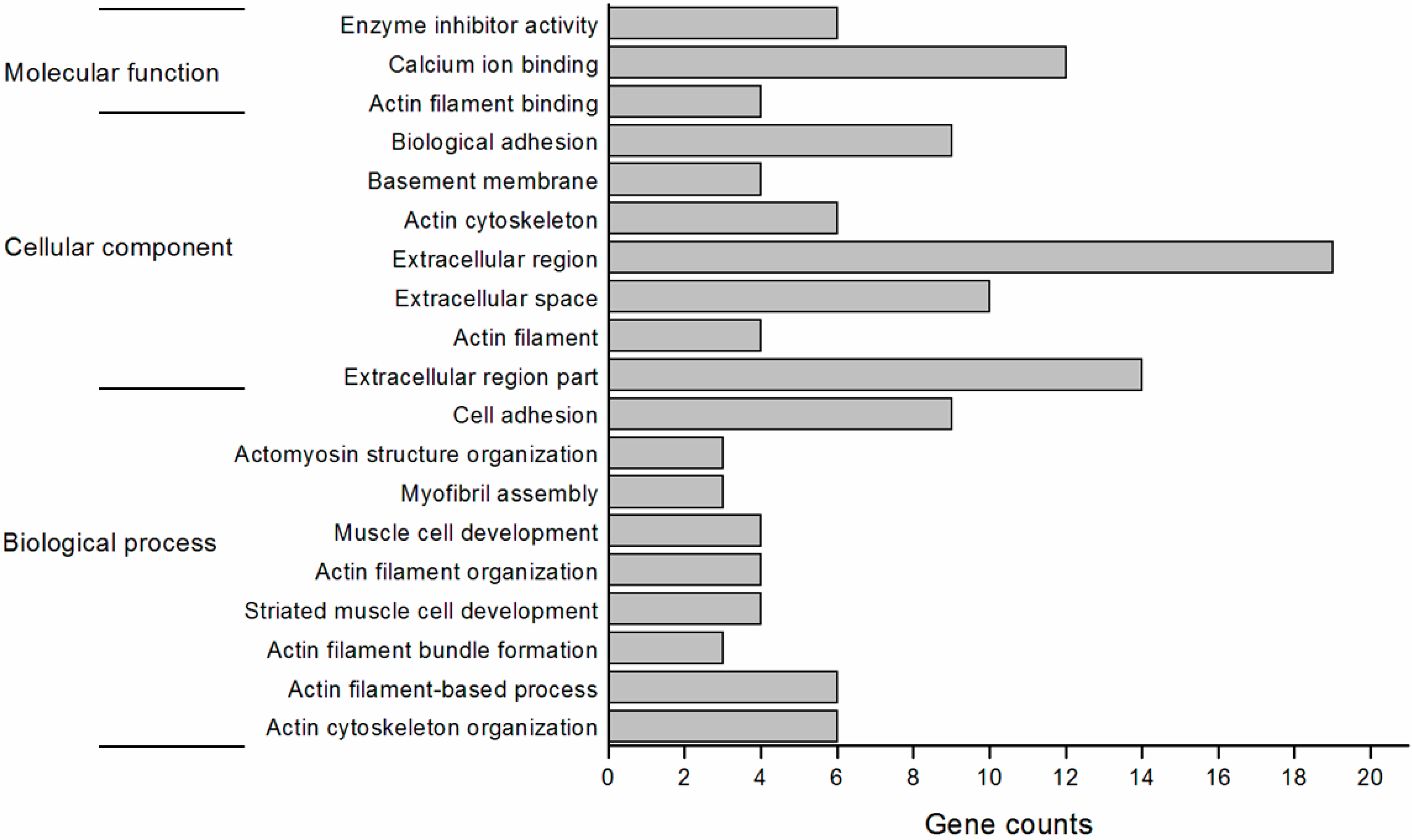
2.4. Function Annotation of the Differentially Expressed Genes
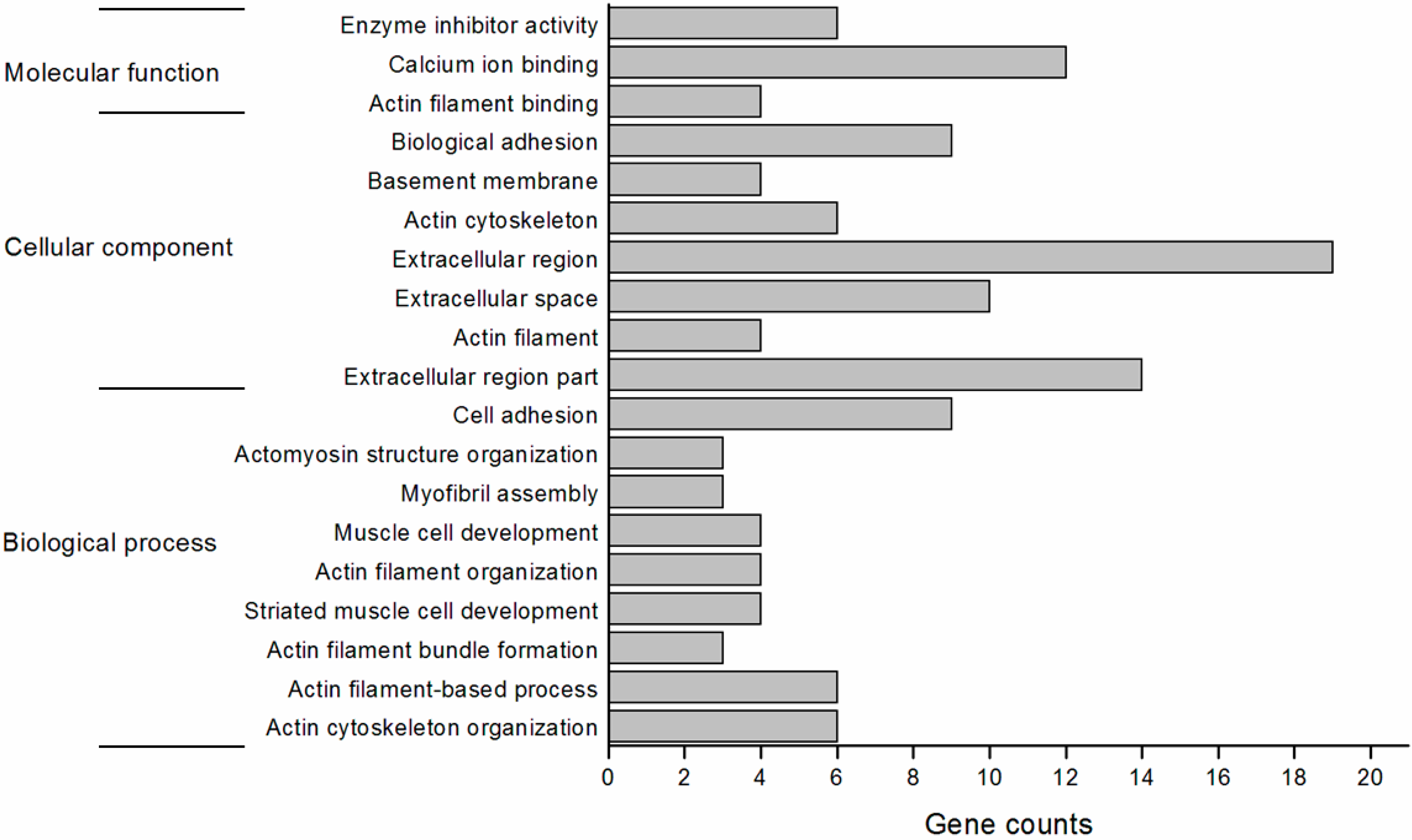
2.5. Validation of the Differentially Expressed Genes by RT-qPCR
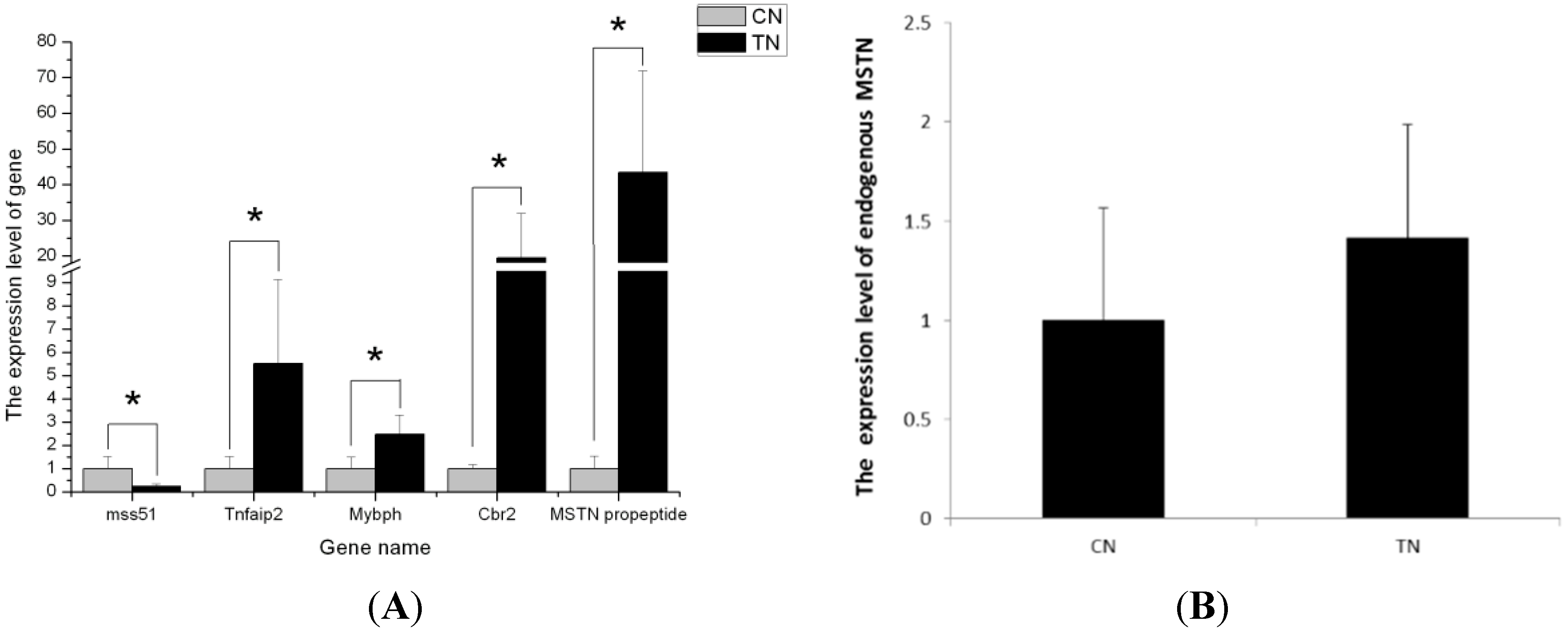
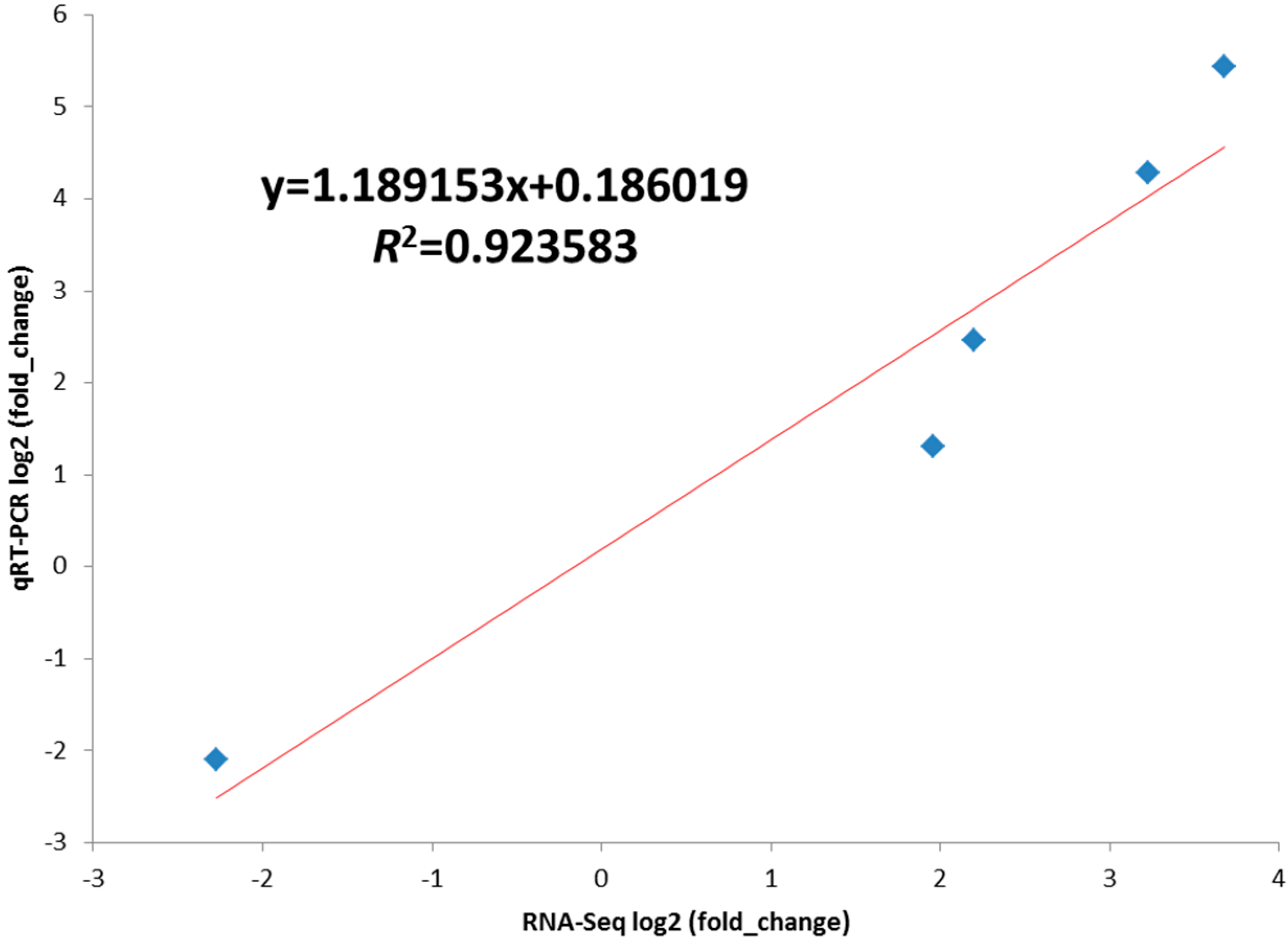
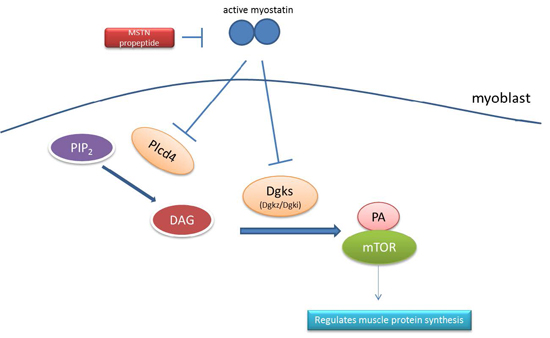
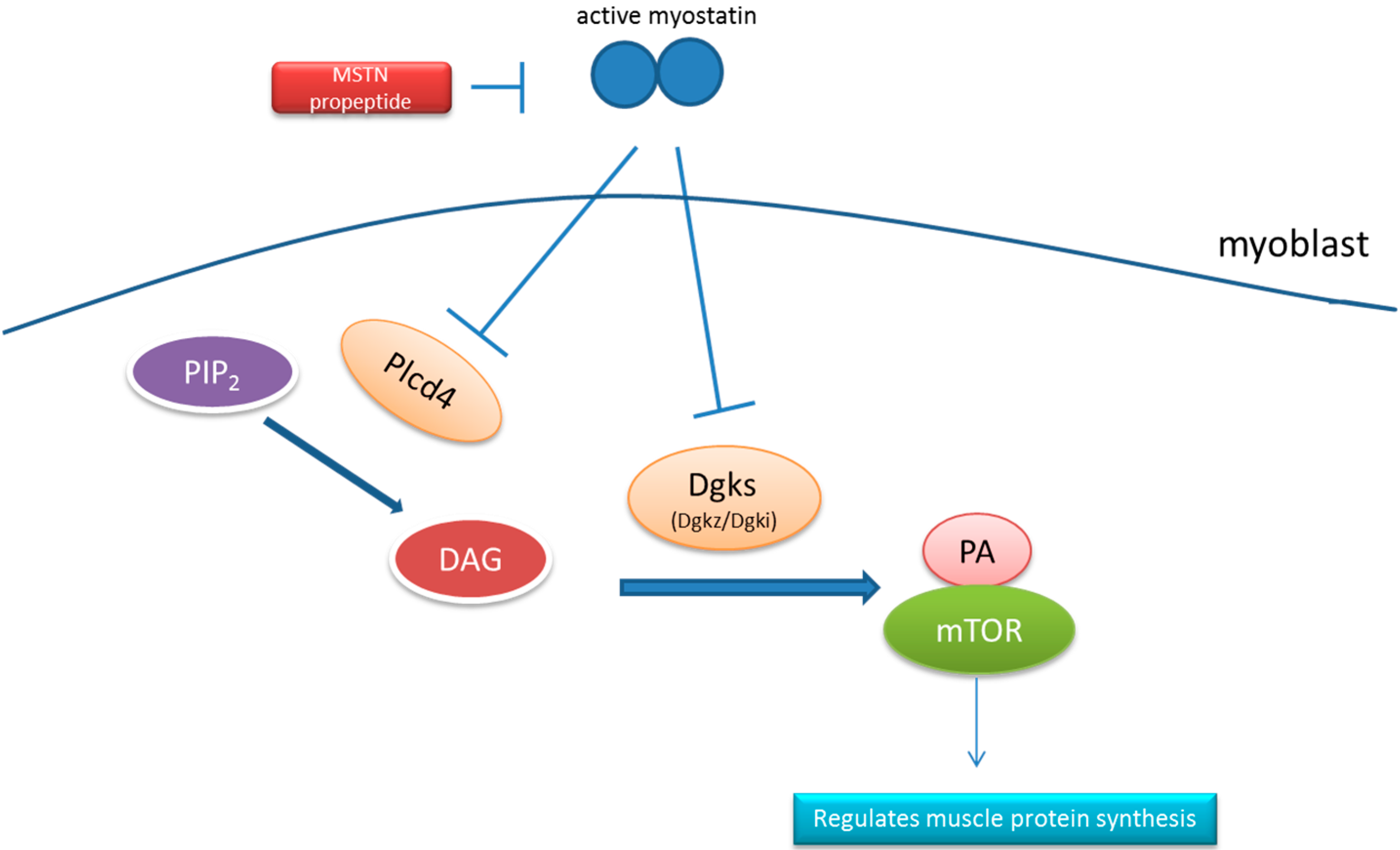
2.6. Signaling Pathway Analysis
| Category | Term | Gene Counts | Genes |
|---|---|---|---|
| KEGG_PATHWAY | Hypertrophic cardiomyopathy (HCM) | 5 | Actb, Itga4, Actg1, Myh6, Acta |
| KEGG_PATHWAY | Dilated cardiomyopathy | 5 | Actb, Itga4, Actg1, Myh6, Acta |
| KEGG_PATHWAY | Focal adhesion | 5 | Lama1, Lamc3, Actb, Itga4, Actg1 |
| KEGG_PATHWAY | Arrhythmogenic right ventricular cardiomyopathy (ARVC) | 3 | Actb, Itga4, Actg1 |
| KEGG_PATHWAY | Phosphatidic acid signaling system | 3 | Dgki, Dgkz, Plcd4 |
| KEGG_PATHWAY | Complement and coagulation cascades | 3 | C4bp, Proc, F12 |
| KEGG_PATHWAY | ECM-receptor interaction | 3 | Lama1, Lamc3, Itga4 |
| KEGG_PATHWAY | Hematopoietic cell lineage | 3 | Itga4, Cd3d, Cd5 |

3. Discussion
4. Experimental Section
4.1. Muscle Tissue Sample Collections
4.2. RNA Isolation and RNA Sequencing
4.3. Quality Control for Raw Sequencing Data
4.4. Bioinformatics Analysis
4.5. Differential Expression Gene Validation by Real Time RT-PCR
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lee, S.J. Regulation of muscle mass by myostatin. Annu. Rev. Cell Dev. Biol. 2004, 20, 61–86. [Google Scholar] [CrossRef] [PubMed]
- Parsons, S.A.; Millay, D.P.; Sargent, M.A.; McNally, E.M.; Molkentin, J.D. Age-dependent effect of myostatin blockade on disease severity in a murine model of limb-girdle muscular dystrophy. Am. J. Pathol. 2006, 168, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new tgf-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Zimmers, T.A.; Davies, M.V.; Koniaris, L.G.; Haynes, P.; Esquela, A.F.; Tomkinson, K.N.; McPherron, A.C.; Wolfman, N.M.; Lee, S.J. Induction of cachexia in mice by systemically administered myostatin. Science 2002, 296, 1486–1488. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, K. Myostatin inhibition by a follistatin-derived peptide ameliorates the pathophysiology of muscular dystrophy model mice. Acta Myol. 2008, 27, 14–18. [Google Scholar] [PubMed]
- McPherron, A.C.; Lee, S.J. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA 1997, 94, 12457–12461. [Google Scholar] [CrossRef] [PubMed]
- Kambadur, R.; Sharma, M.; Smith, T.P.; Bass, J.J. Mutations in myostatin (gdf8) in double-muscled belgian blue and piedmontese cattle. Genome Res. 1997, 7, 910–916. [Google Scholar] [PubMed]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibe, B.; Bouix, J.; Caiment, F.; Elsen, J.M.; Eychenne, F.; et al. A mutation creating a potential illegitimate microrna target site in the myostatin gene affects muscularity in sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.S. Myostatin mutation associated with gross muscle hypertrophy in a child. N. Engl. J. Med. 2004, 351, 1030–1030. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.Q.; Jou, W.; Chanturiya, T.; Portas, J.; Gavrilova, O.; McPherron, A.C. Myostatin inhibition in muscle, but not adipose tissue, decreases fat mass and improves insulin sensitivity. PLoS ONE 2009, 4, e4937. [Google Scholar] [CrossRef] [PubMed]
- Bradley, L.; Yaworsky, P.J.; Walsh, F.S. Myostatin as a therapeutic target for musculoskeletal disease. Cell Mol. Life Sci. 2008, 65, 2119–2124. [Google Scholar] [CrossRef] [PubMed]
- Attisano, L.; Wrana, J.L. Signal transduction by the TGF-β superfamily. Science 2002, 296, 1646–1647. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [PubMed]
- Joulia-Ekaza, D.; Cabello, G. Myostatin regulation of muscle development: Molecular basis, natural mutations, physiopathological aspects. Exp. Cell Res. 2006, 312, 2401–2414. [Google Scholar] [CrossRef] [PubMed]
- Langley, B.; Thomas, M.; Bishop, A.; Sharma, M.; Gilmour, S.; Kambadur, R. Myostatin inhibits myoblast differentiation by down-regulating myod expression. J. Biol. Chem. 2002, 277, 49831–49840. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Shinozaki, M.; Hara, T.; Furuya, T.; Miyazono, K. Two major smad pathways in TGF-β superfamily signalling. Genes Cells 2002, 7, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.H.; Chen, Y.G. Smad7: Not only a regulator, but also a cross-talk mediator of TGF-β signalling. Biochem. J. 2011, 434, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.A.; McNally, R.M.; Hoffmann, F.M.; Hornberger, T.A. Smad3 induces atrogin-1, inhibits mtor and protein synthesis, and promotes muscle atrophy in vivo. Mol. Endocrinol. 2013, 27, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Lokireddy, S.; McFarlane, C.; Ge, X.J.; Zhang, H.M.; Sze, S.K.; Sharma, M.; Kambadur, R. Myostatin induces degradation of sarcomeric proteins through a smad3 signaling mechanism during skeletal muscle wasting. Mol. Endocrinol. 2011, 25, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Trendelenburg, A.U.; Meyer, A.; Rohner, D.; Boyle, J.; Hatakeyama, S.; Glass, D.J. Myostatin reduces Akt/TORC1/p70S6K signaling, inhibiting myoblast differentiation and myotube size. Am. J. Physiol. Cell Physiol. 2009, 296, C1258–C1270. [Google Scholar] [CrossRef] [PubMed]
- Bogdanovich, S.; Krag, T.O.; Barton, E.R.; Morris, L.D.; Whittemore, L.A.; Ahima, R.S.; Khurana, T.S. Functional improvement of dystrophic muscle by myostatin blockade. Nature 2002, 420, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Diel, P.; Schiffer, T.; Geisler, S.; Hertrampf, T.; Mosler, S.; Schulz, S.; Wintgens, K.F.; Adler, M. Analysis of the effects of androgens and training on myostatin propeptide and follistatin concentrations in blood and skeletal muscle using highly sensitive immuno PCR. Mol. Cell. Endocrinol. 2010, 330, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.J.; Davies, M.V.; Pearson, A.A.; Wang, J.H.; Hewick, R.M.; Wolfman, N.M.; Qiu, Y. The myostatin propeptide and the follistatin-related gene are inhibitory binding proteins of myostatin in normal serum. J. Biol. Chem. 2002, 277, 40735–40741. [Google Scholar] [CrossRef] [PubMed]
- Kishioka, Y.; Thomas, M.; Wakamatsu, J.I.; Hattori, A.; Sharma, M.; Kambadur, R.; Nishimura, T. Decorin enhances the proliferation and differentiation of myogenic cells through suppressing myostatin activity. J. Cell. Physiol. 2008, 215, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, M.; Li, Y.; Zhu, J.; Ambrosio, F.; Uehara, K.; Fu, F.H.; Huard, J. Improved muscle healing after contusion injury by the inhibitory effect of suramin on myostatin, a negative regulator of muscle growth. Am. J. Sports Med. 2008, 36, 2354–2362. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.Q.; Chen, X.L.; Yu, B.; He, J.; Chen, D.W. Microrna-27a promotes myoblast proliferation by targeting myostatin. Biochem. Biophys. Res. Commun. 2012, 423, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Miretti, S.; Martignani, E.; Accornero, P.; Baratta, M. Functional effect of mir-27b on myostatin expression: A relationship in piedmontese cattle with double-muscled phenotype. BMC Genomics 2013, 14, 194. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, B. Postnatal expression of myostatin propeptide cdna maintained high muscle growth and normal adipose tissue mass in transgenic mice fed a high-fat diet. Mol. Reprod. Dev. 2006, 73, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ratovitski, T.; Brady, J.P.; Solomon, M.B.; Wells, K.D.; Wall, R.J. Expression of myostatin pro domain results in muscular transgenic mice. Mol. Reprod. Dev. 2001, 60, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Li, E.J.; Wall, R.J.; Yang, J. Coordinated patterns of gene expressions for adult muscle build-up in transgenic mice expressing myostatin propeptide. BMC Genomics 2009, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Laxalt, A.M.; Munnik, T. Phospholipid signalling in plant defence. Curr. Opin. Plant Biol. 2002, 5, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K.; Takenaka, K.; Nagano, K.; Takenawa, T. Growth factor-induced promoter activation of murine phospholipase C δ4 gene. Eur. J. Biochem. 2000, 267, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Fukami, K.; Yu, H.; Takenawa, T. A new phospholipase C δ4 is induced at s-phase of the cell cycle and appears in the nucleus. J. Biol. Chem. 1996, 271, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Veverka, V.; Crabbe, T.; Bird, I.; Lennie, G.; Muskett, F.W.; Taylor, R.J.; Carr, M.D. Structural characterization of the interaction of mTOR with phosphatidic acid and a novel class of inhibitor: Compelling evidence for a central role of the FRB domain in small molecule-mediated regulation of mTOR. Oncogene 2008, 27, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Vilella-Bach, M.; Bachmann, R.; Flanigan, A.; Chen, J. Phosphatidic acid-mediated mitogenic activation of mTOR signaling. Science 2001, 294, 1942–1945. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.M.; Kiriazis, H.; Moore, X.L.; Feng, X.H.; Sheppard, K.; Dart, A.; Du, X.J. Regression of pressure overload-induced left ventricular hypertrophy in mice. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H2702–H2707. [Google Scholar] [CrossRef] [PubMed]
- Swynghedauw, B. Developmental and functional adaptation of contractile proteins in cardiac and skeletal muscles. Physiol. Rev. 1986, 66, 710–771. [Google Scholar] [PubMed]
- Ojamaa, K.; Petrie, J.F.; Balkman, C.; Hong, C.; Klein, I. Posttranscriptional modification of myosin heavy-chain gene expression in the hypertrophied rat myocardium. Proc. Natl. Acad. Sci. USA 1994, 91, 3468–3472. [Google Scholar] [CrossRef] [PubMed]
- Amthor, H.; Macharia, R.; Navarrete, R.; Schuelke, M.; Brown, S.C.; Otto, A.; Voit, T.; Muntoni, F.; Vrbova, G.; Partridge, T.; et al. Lack of myostatin results in excessive muscle growth but impaired force generation. Proc. Natl. Acad. Sci. USA 2007, 104, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Tyska, M.J.; Warshaw, D.M. The myosin power stroke. Cell Motil Cytoskeleton 2002, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Blanco, G.; Coulton, G.R.; Biggin, A.; Grainge, C.; Moss, J.; Barrett, M.; Berquin, A.; Marechal, G.; Skynner, M.; van Mier, P.; et al. The kyphoscoliosis (ky) mouse is deficient in hypertrophic responses and is caused by a mutation in a novel muscle-specific protein. Hum. Mol. Genet. 2001, 10, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.D.; Culetto, E.; Ponting, C.P.; Miguel-Aliaga, I.; Davies, K.E.; Sattelle, D.B. Cloning and developmental expression analysis of ltd-1, the caenorhabditis elegans homologue of the mouse kyphoscoliosis (ky) gene. Mech. Dev. 2002, 117, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Beatham, J.; Romero, R.; Townsend, S.K.M.; Hacker, T.; van der Ven, P.F.M.; Blanco, G. Filamin C interacts with the muscular dystrophy KY protein and is abnormally distributed in mouse KY deficient muscle fibres. Hum. Mol. Genet. 2004, 13, 2863–2874. [Google Scholar] [CrossRef] [PubMed]
- Conti, A.; Riva, N.; Pesca, M.; Iannaccone, S.; Cannistraci, C.V.; Corbo, M.; Previtali, S.C.; Quattrini, A.; Alessio, M. Increased expression of myosin binding protein H in the skeletal muscle of amyotrophic lateral sclerosis patients. Biochim. Biophys. Acta 2014, 1842, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ishikawa, R.; Okagaki, T.; Ye, L.H.; Kohama, K. Stimulation of the ATP-dependent interaction between actin and myosin by a myosin-binding fragment of smooth muscle caldesmon. Cell Motil. Cytoskeleton 1994, 29, 250–258. [Google Scholar] [CrossRef] [PubMed]
- dos Remedios, C.G.; Chhabra, D.; Kekic, M.; Dedova, I.V.; Tsubakihara, M.; Berry, D.A.; Nosworthy, N.J. Actin binding proteins: Regulation of cytoskeletal microfilaments. Physiol. Rev. 2003, 83, 433–473. [Google Scholar] [CrossRef] [PubMed]
- Doherty, G.J.; McMahon, H.T. Mediation, modulation, and consequences of membrane-cytoskeleton interactions. Annu. Rev. Biophys. 2008, 37, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Devaiah, S.P.; Zhang, W.; Welti, R. Signaling functions of phosphatidic acid. Prog. Lipid Res. 2006, 45, 250–278. [Google Scholar] [CrossRef]
- Janda, M.; Planchais, S.; Djafi, N.; Martinec, J.; Burketova, L.; Valentova, O.; Zachowski, A.; Ruelland, E. Phosphoglycerolipids are master players in plant hormone signal transduction. Plant Cell Rep. 2013, 32, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Munnik, T. Phosphatidic acid: An emerging plant lipid second messenger. Trends Plant Sci. 2001, 6, 227–233. [Google Scholar] [CrossRef] [PubMed]
- You, J.S.; Lincoln, H.C.; Kim, C.R.; Frey, J.W.; Goodman, C.A.; Zhong, X.P.; Hornberger, T.A. The role of diacylglycerol kinase zeta and phosphatidic acid in the mechanical activation of mammalian target of rapamycin (mTOR) signaling and skeletal muscle hypertrophy. J. Biol. Chem. 2014, 289, 1551–1563. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.A.; Frey, J.W.; Mabrey, D.M.; Jacobs, B.L.; Lincoln, H.C.; You, J.S.; Hornberger, T.A. The role of skeletal muscle mTOR in the regulation of mechanical load-induced growth. J. Physiol. 2011, 589, 5485–5501. [Google Scholar] [CrossRef] [PubMed]
- Hornberger, T.A. Mechanotransduction and the regulation of mTORC1 signaling in skeletal muscle. Int. J. Biochem. Cell Biol. 2011, 43, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, S.I.; Rincon, E.; Avila-Flores, A.; Torres-Ayuso, P.; Almena, M.; Cobos, M.A.; Albar, J.P.; Merida, I. Diacylglycerol kinase zeta controls diacylglycerol metabolism at the immunological synapse. Mol. Biol. Cell 2011, 22, 4406–4414. [Google Scholar] [CrossRef] [PubMed]
- Toschi, A.; Lee, E.; Xu, L.; Garcia, A.; Gadir, N.; Foster, D.A. Regulation of mTORC1 and mTORC2 complex assembly by phosphatidic acid: Competition with rapamycin. Mol. Cell. Biol. 2009, 29, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Hulmi, J.J.; Silvennoinen, M.; Lehti, M.; Kivela, R.; Kainulainen, H. Altered redd1, myostatin, and akt/mtor/foxo/mapk signaling in streptozotocin-induced diabetic muscle atrophy. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E307–E315. [Google Scholar] [CrossRef] [PubMed]
- Abramovici, H.; Mojtabaie, P.; Parks, R.J.; Zhong, X.P.; Koretzky, G.A.; Topham, M.K.; Gee, S.H. Diacylglycerol kinase zeta regulates actin cytoskeleton reorganization through dissociation of Rac1 from RhoGDI. Mol. Biol. Cell 2009, 20, 2049–2059. [Google Scholar] [CrossRef] [PubMed]
- Soto, I.C.; Fontanesi, F.; Myers, R.S.; Hamel, P.; Barrientos, A. A heme-sensing mechanism in the translational regulation of mitochondrial cytochrome c oxidase biogenesis. Cell Metab. 2012, 16, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, F.; Soto, I.C.; Horn, D.; Barrientos, A. Mss51 and Ssc1 facilitate translational regulation of cytochrome c oxidase biogenesis. Mol. Cell. Biol. 2010, 30, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, J.; Ishikura, S.; Asami, J.; Isaji, T.; Usami, N.; Hara, A.; Sakurai, T.; Tsuritani, K.; Oda, K.; Takahashi, M.; et al. Molecular characterization of mammalian dicarbonyl/l-xylulose reductase and its localization in kidney. J. Biol. Chem. 2002, 277, 17883–17891. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ma, L.; Huang, W.; Shai, Y.; Ji, X.; Ding, L.; Liu, Y.; Yu, L.; Zhao, S. Decreased expression of the human carbonyl reductase 2 gene hcr2 in hepatocellular carcinoma. Cell Mol. Biol. Lett. 2006, 11, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, F.; Maser, E. Carbonyl reductases and pluripotent hydroxysteroid dehydrogenases of the short-chain dehydrogenase/reductase superfamily. Drug Metab. Rev. 2007, 39, 87–144. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, E.; Sestili, P. Reactive oxygen species in skeletal muscle signaling. J. Signal Transduct. 2012, 2012. [Google Scholar] [CrossRef]
- Lee, K.-P.; Shin, Y.J.; Cho, S.C.; Lee, S.-M.; Bahn, Y.J.; Kim, J.Y.; Kwon, E.-S.; Park, S.C.; Rhee, S.G.; Woo, H.A.; et al. Peroxiredoxin 3 has a crucial role in the contractile function of skeletal muscle by regulating mitochondrial homeostasis. Free Radic. Biol. Med. 2014, 77, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Sriram, S.; Subramanian, S.; Juvvuna, P.K.; Ge, X.; Lokireddy, S.; McFarlane, C.D.; Wahli, W.; Kambadur, R.; Sharma, M. Myostatin augments muscle-specific ring finger protein-1 expression through an Nf-kB independent mechanism in SMAD3 null muscle. Mol. Endocrinol. 2014, 28, 317–330. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, Y.; Yang, J.; Xu, Z.; Jing, L.; Zhao, S.; Li, X. RNA Sequencing Identifies Upregulated Kyphoscoliosis Peptidase and Phosphatidic Acid Signaling Pathways in Muscle Hypertrophy Generated by Transgenic Expression of Myostatin Propeptide. Int. J. Mol. Sci. 2015, 16, 7976-7994. https://doi.org/10.3390/ijms16047976
Miao Y, Yang J, Xu Z, Jing L, Zhao S, Li X. RNA Sequencing Identifies Upregulated Kyphoscoliosis Peptidase and Phosphatidic Acid Signaling Pathways in Muscle Hypertrophy Generated by Transgenic Expression of Myostatin Propeptide. International Journal of Molecular Sciences. 2015; 16(4):7976-7994. https://doi.org/10.3390/ijms16047976
Chicago/Turabian StyleMiao, Yuanxin, Jinzeng Yang, Zhong Xu, Lu Jing, Shuhong Zhao, and Xinyun Li. 2015. "RNA Sequencing Identifies Upregulated Kyphoscoliosis Peptidase and Phosphatidic Acid Signaling Pathways in Muscle Hypertrophy Generated by Transgenic Expression of Myostatin Propeptide" International Journal of Molecular Sciences 16, no. 4: 7976-7994. https://doi.org/10.3390/ijms16047976
APA StyleMiao, Y., Yang, J., Xu, Z., Jing, L., Zhao, S., & Li, X. (2015). RNA Sequencing Identifies Upregulated Kyphoscoliosis Peptidase and Phosphatidic Acid Signaling Pathways in Muscle Hypertrophy Generated by Transgenic Expression of Myostatin Propeptide. International Journal of Molecular Sciences, 16(4), 7976-7994. https://doi.org/10.3390/ijms16047976