
Metabolic Profiling of Pyrrolizidine Alkaloids in Foliage of Two Echium spp. Invaders in Australia—A Case of Novel Weapons?


 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
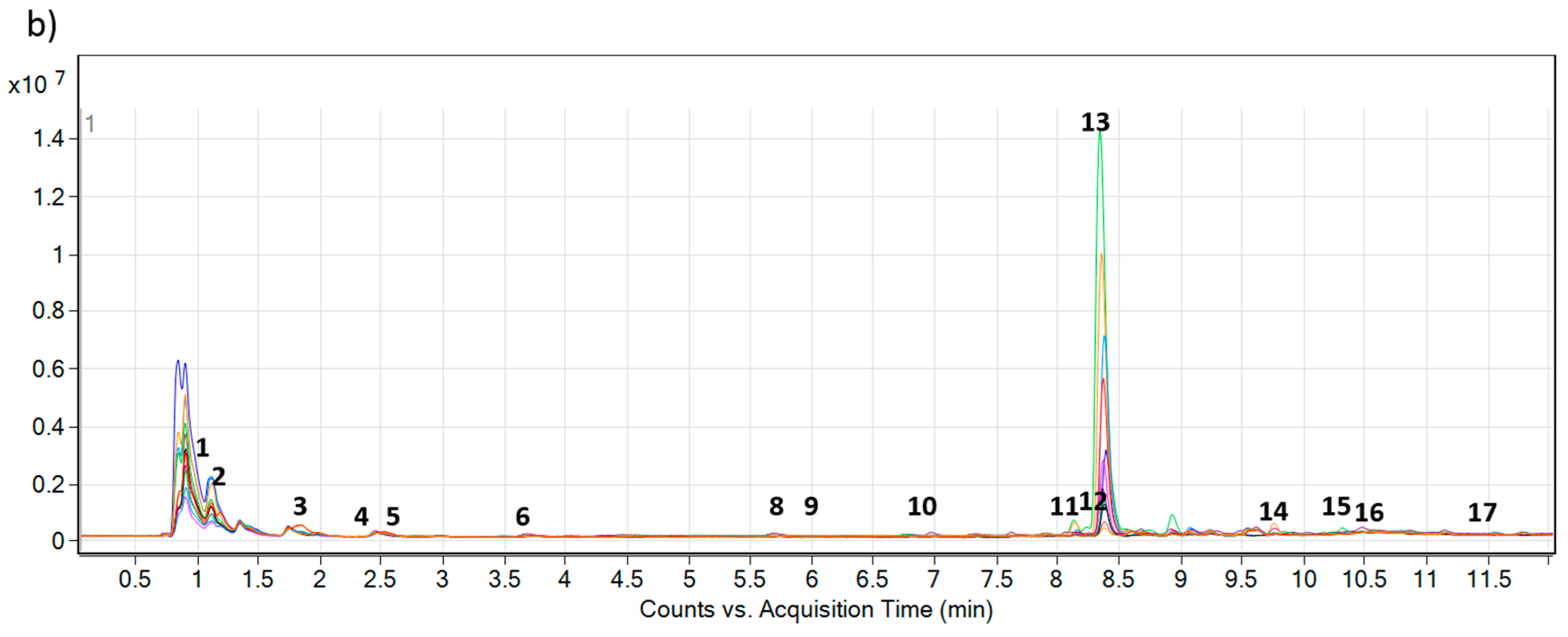
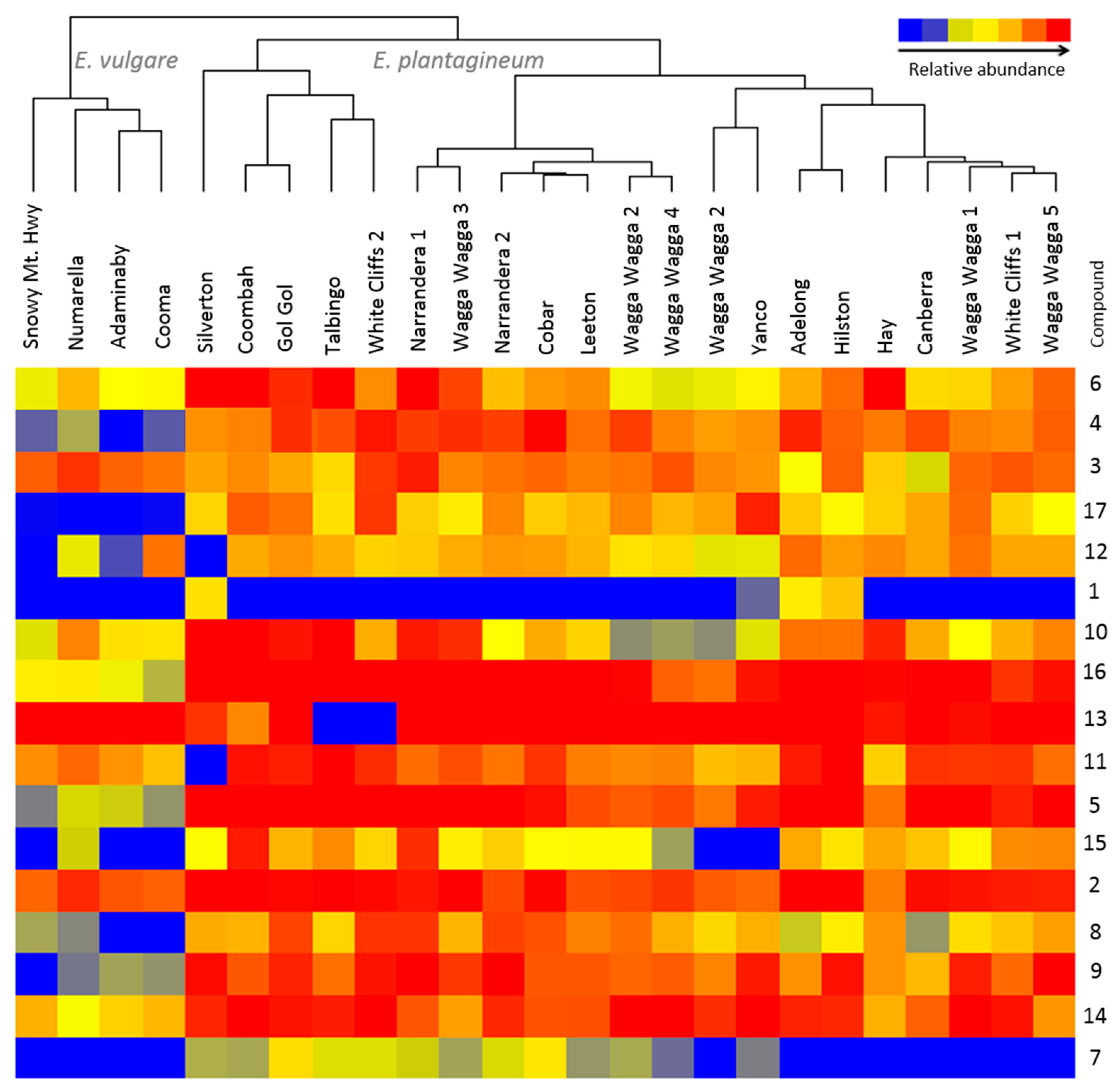
2.1. Profiling of Pyrrolizidine Alkaloids from Foliage
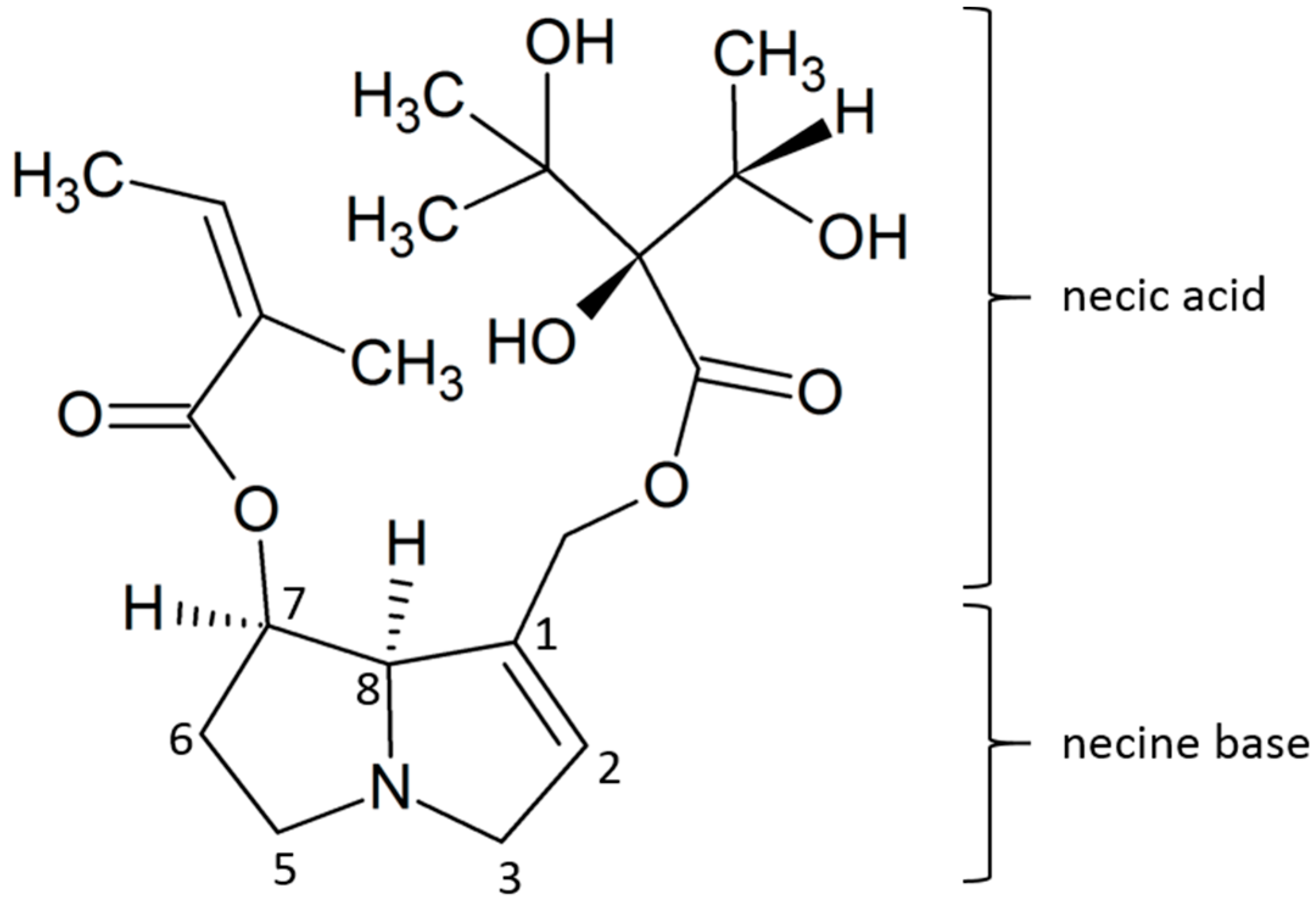
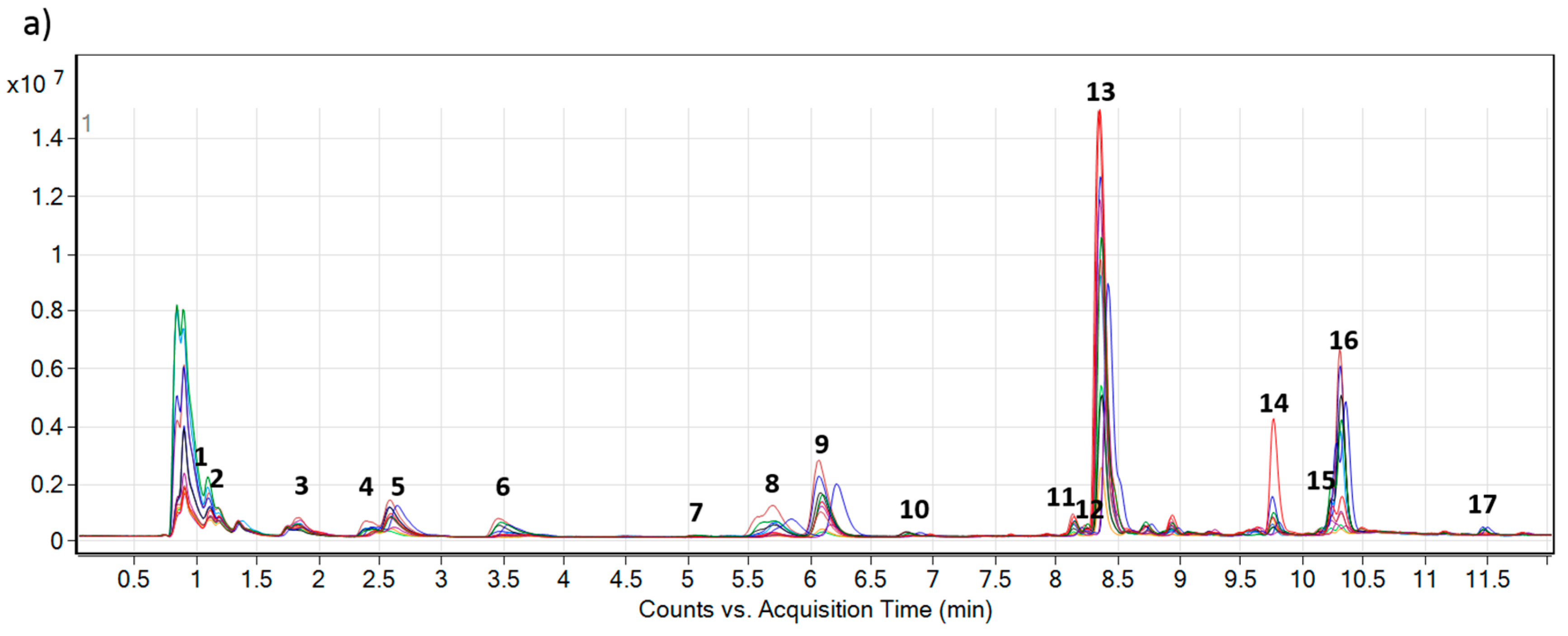
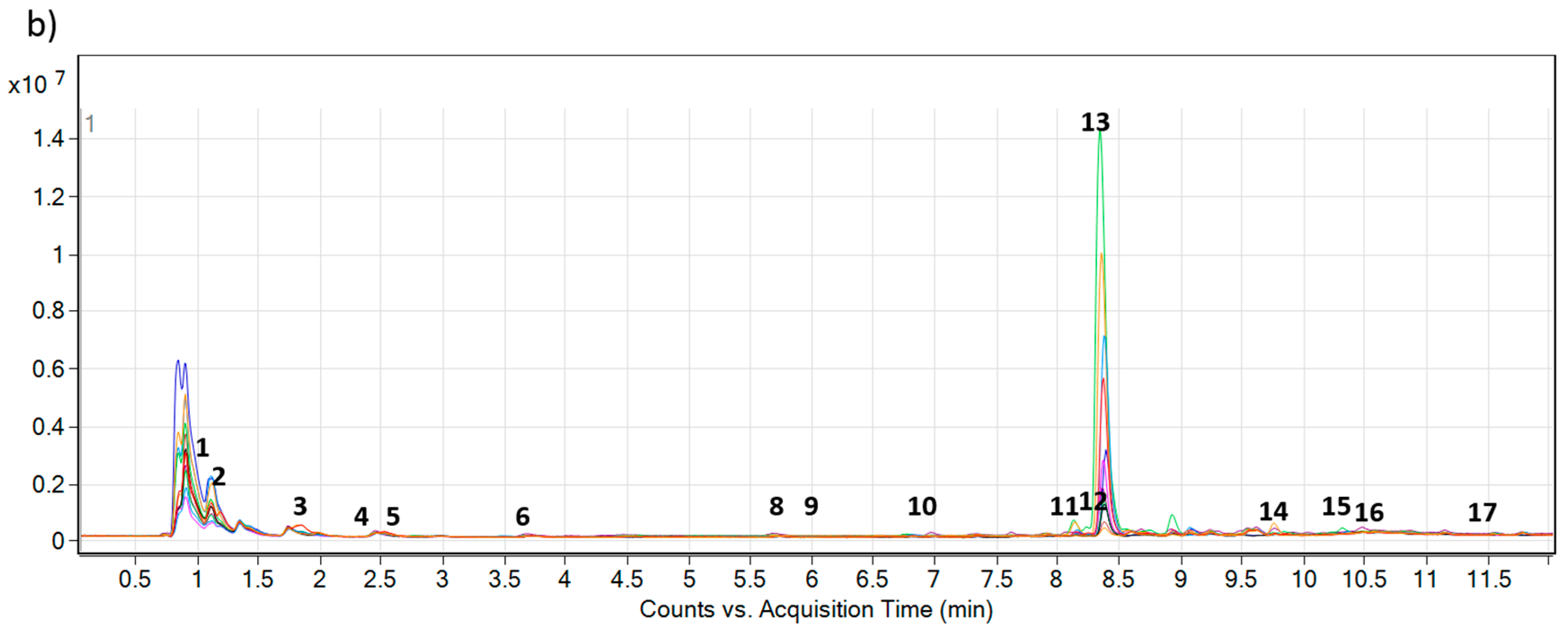
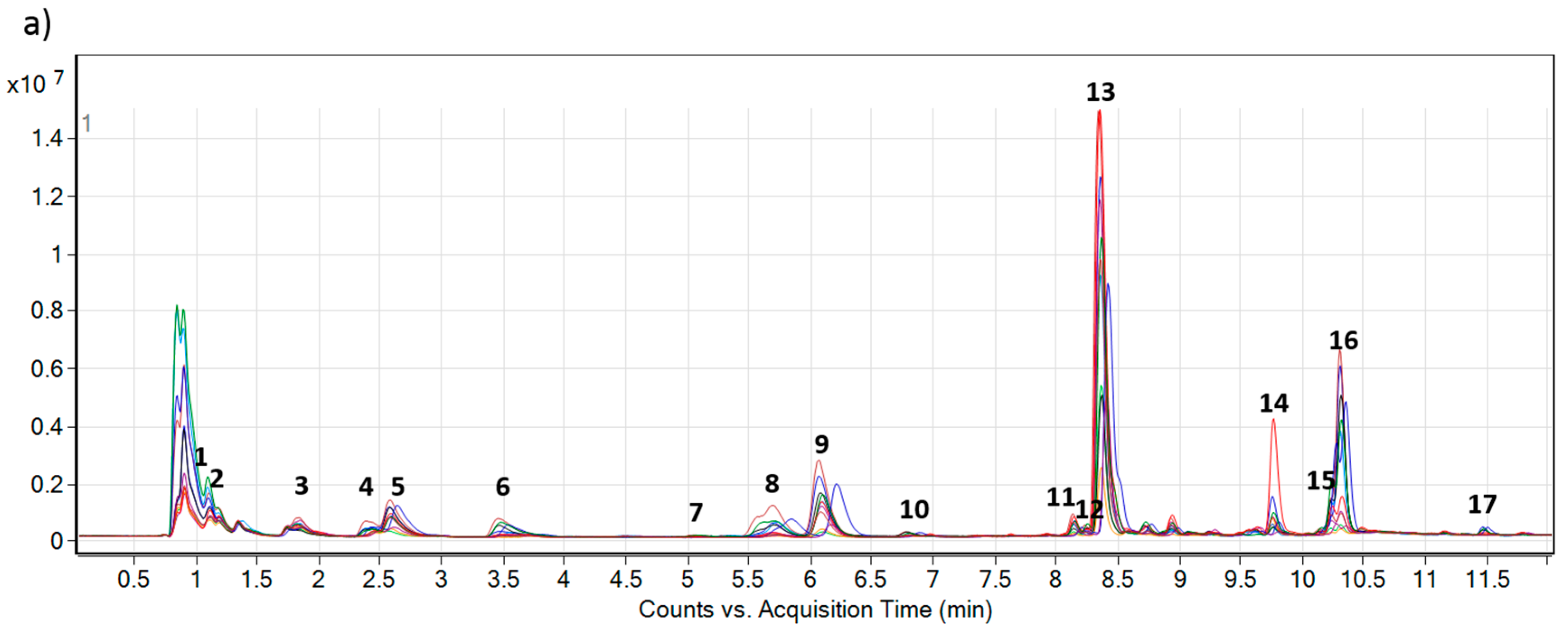
2.2. Accumulation of PAs and PANOs in the Field Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name | AM | Calculated [M + H] | Measured [M + H] | ∆ppm | Approx. RT | Formula |
|---|---|---|---|---|---|---|---|
| 1 | Leptanthine-N-oxide | 331.1631 | 332.1704 | 332.1701 | 0.90 | 1.08 | C15H25NO7 |
| 2 | Echimiplatine-N-oxide | 331.1631 | 332.1704 | 332.1701 | 0.90 | 1.27 | C15H25NO7 |
| 3 | Uplandicine-N-oxide | 373.1737 | 374.1809 | 374.1808 | 0.27 | 1.91 | C17H27NO8 |
| 4 | Intermedine-N-oxide | 315.1682 | 316.1755 | 316.1754 | 0.32 | 2.35 | C15H25NO6 |
| 5 | Lycopsamine-N-oxide | 315.1682 | 316.1755 | 316.1753 | 0.63 | 2.54 | C15H25NO6 |
| 6 | 7-Angeloylretronencine-N-oxide | 253.1314 | 254.1387 | 254.1384 | 1.18 | 3.42 | C13H19NO4 |
| 7 | 7-O-Acetyllycopsamine * | 341.1838 | 342.1911 | 342.1907 | 1.16 | 5.30 | C17H27NO6 |
| 8 | 7-O-Acetyllycopsamine-N-oxide A * | 357.1788 | 358.1860 | 358.1858 | 0.56 | 5.59 | C17H27NO7 |
| 9 | 7-O-Acetyllycopsamine-N-oxide B * | 357.1788 | 358.1860 | 358.1854 | 1.67 | 5.95 | C17H27NO7 |
| 10 | 9-O-Angelylretronencine-N-oxide | 253.1314 | 254.1387 | 254.1384 | 1.18 | 6.70 | C13H19NO4 |
| 11 | Echimidine-N-oxide A | 413.2050 | 414.2122 | 414.2121 | 0.24 | 8.12 | C20H31NO8 |
| 12 | Echiuplatine-N-oxide | 397.2101 | 398.2173 | 398.2170 | 0.75 | 8.18 | C20H31NO7 |
| 13 | Echimidine-N-oxide B | 413.2050 | 414.2122 | 414.2120 | 0.48 | 8.32 | C20H31NO8 |
| 14 | 3’-O-Acetylechimidine-N-oxide | 455.2155 | 456.2228 | 456.2228 | 0.00 | 9.77 | C22H33NO9 |
| 15 | Echiumine-N-oxide A | 397.2101 | 398.2173 | 398.2172 | 0.25 | 10.20 | C20H31NO7 |
| 16 | Echiumine-N-oxide B | 397.2101 | 398.2173 | 398.2173 | 0.00 | 10.32 | C20H31NO7 |
| 17 | 3’-O-Acetylechiumine-N-oxide | 439.2206 | 440.2279 | 440.2276 | 0.68 | 11.53 | C22H33NO8 |
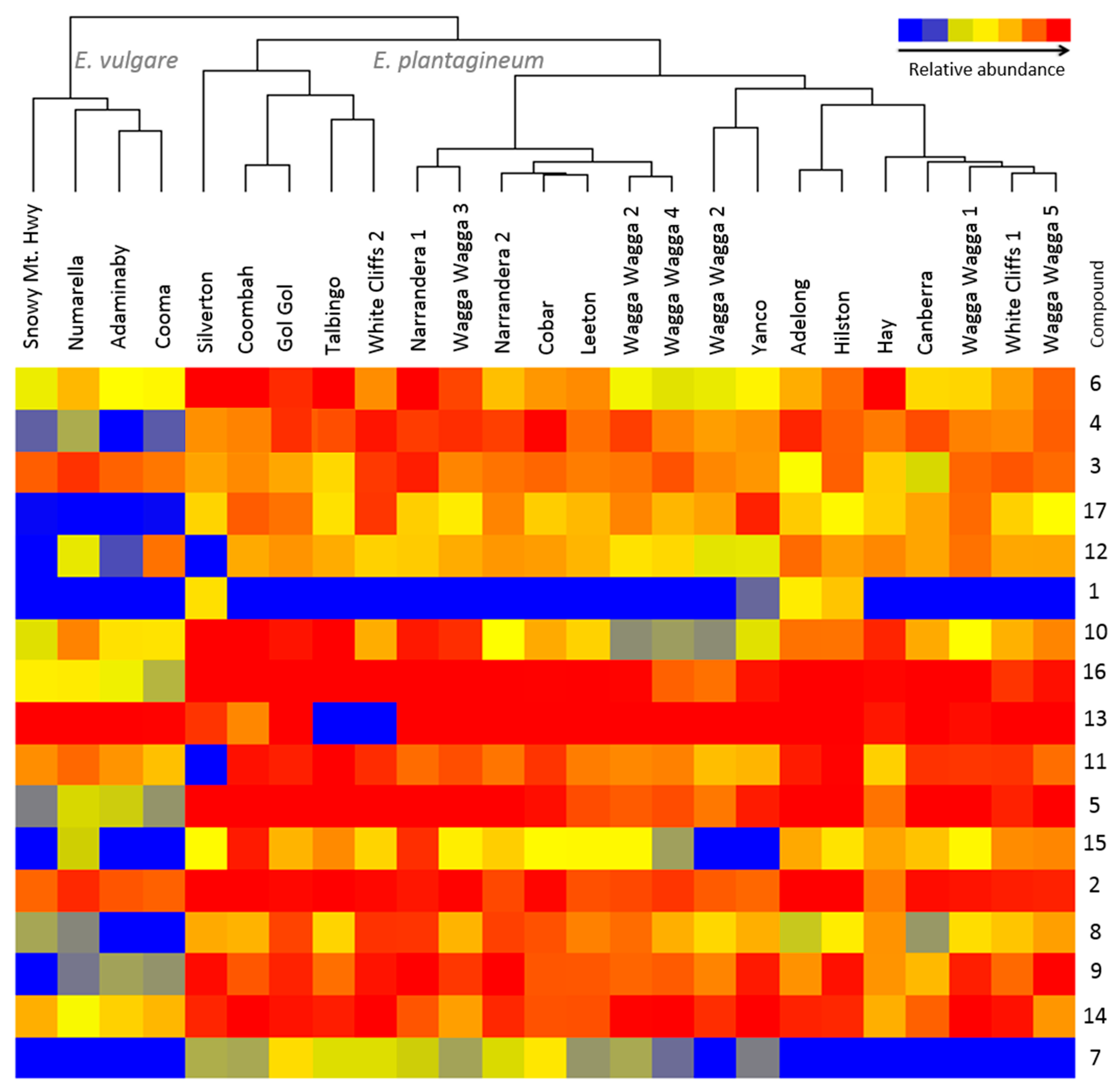
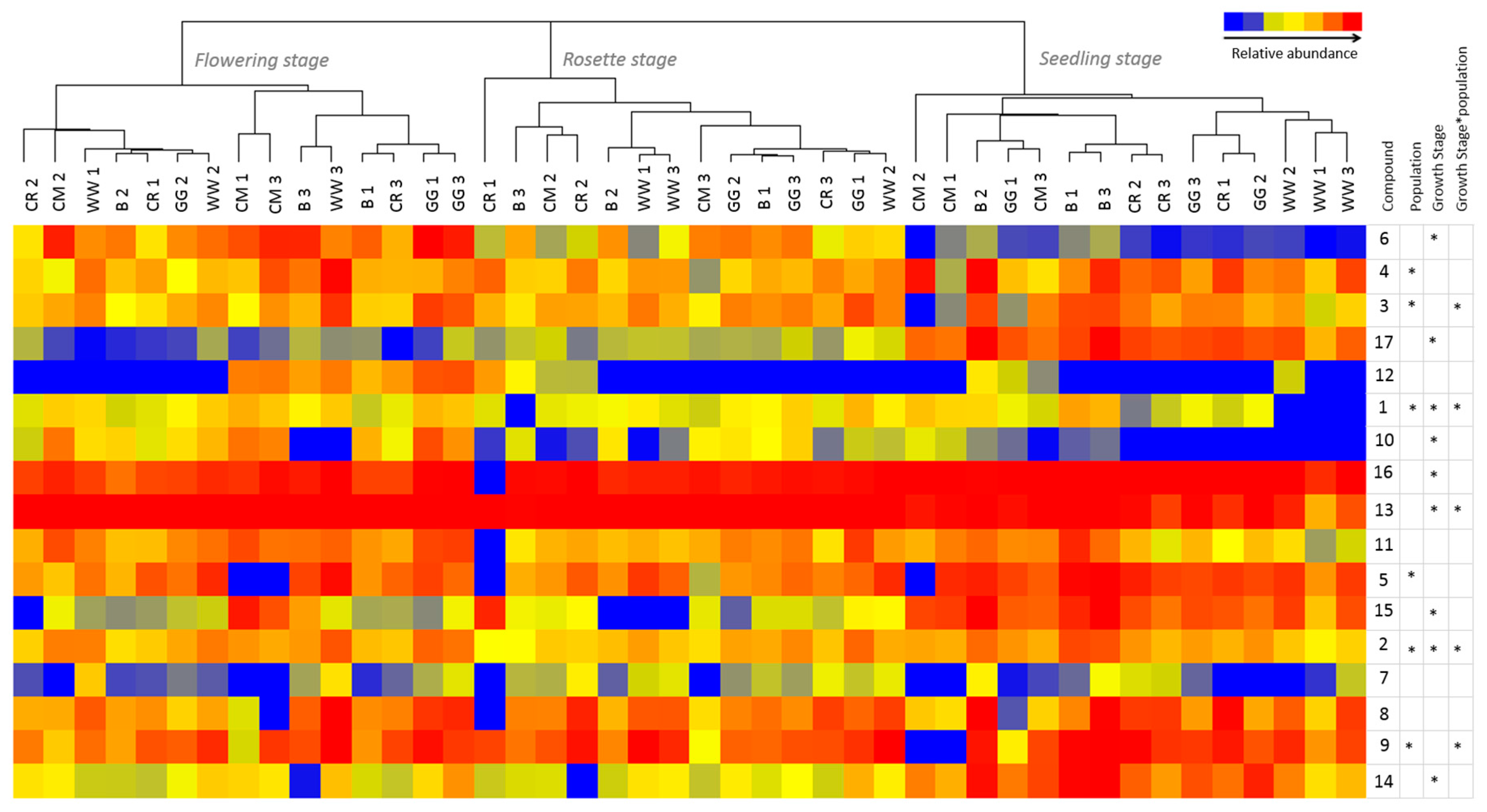
2.3. Growth Stage and Genotype-Dependent Accumulation of PAs and PANOs
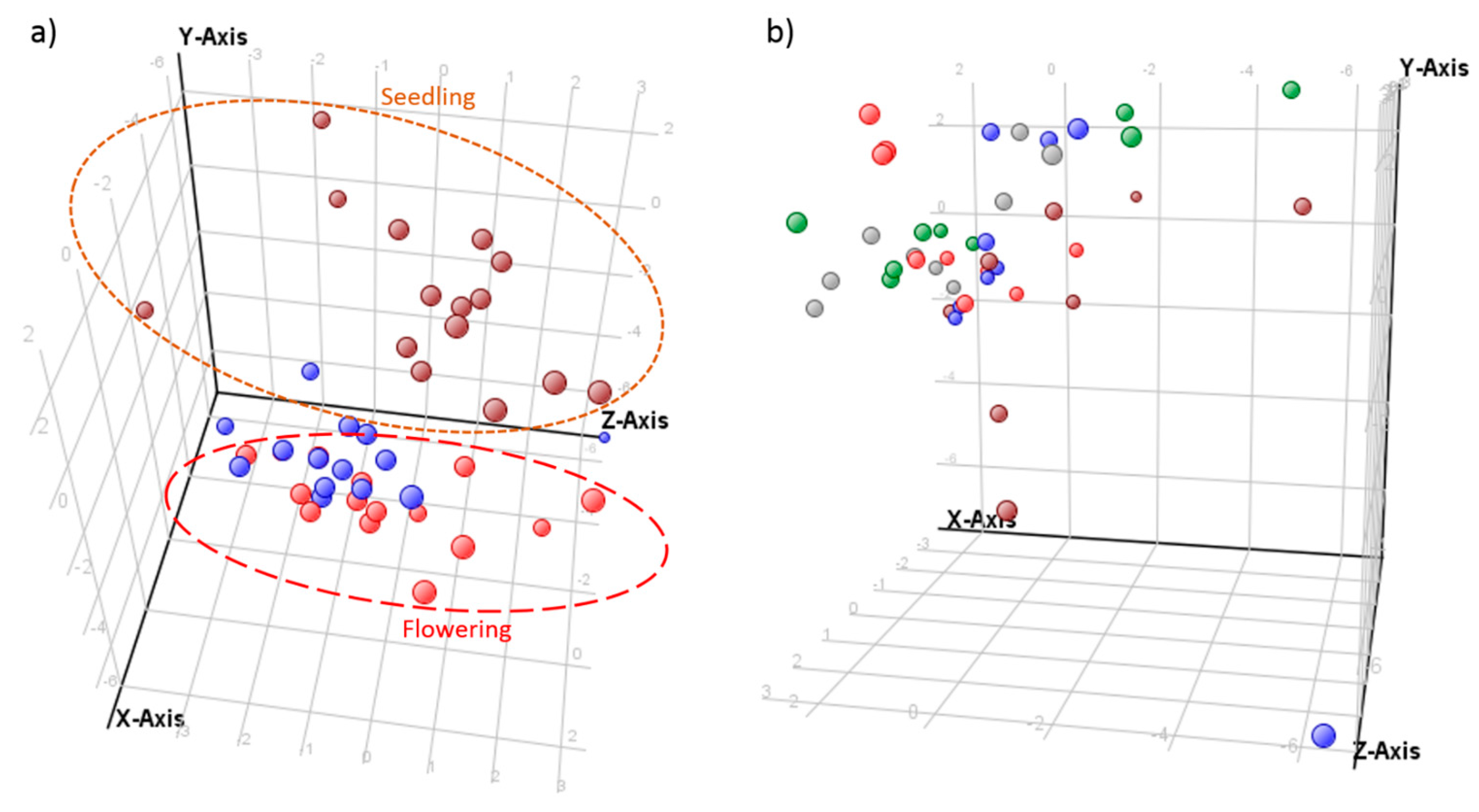
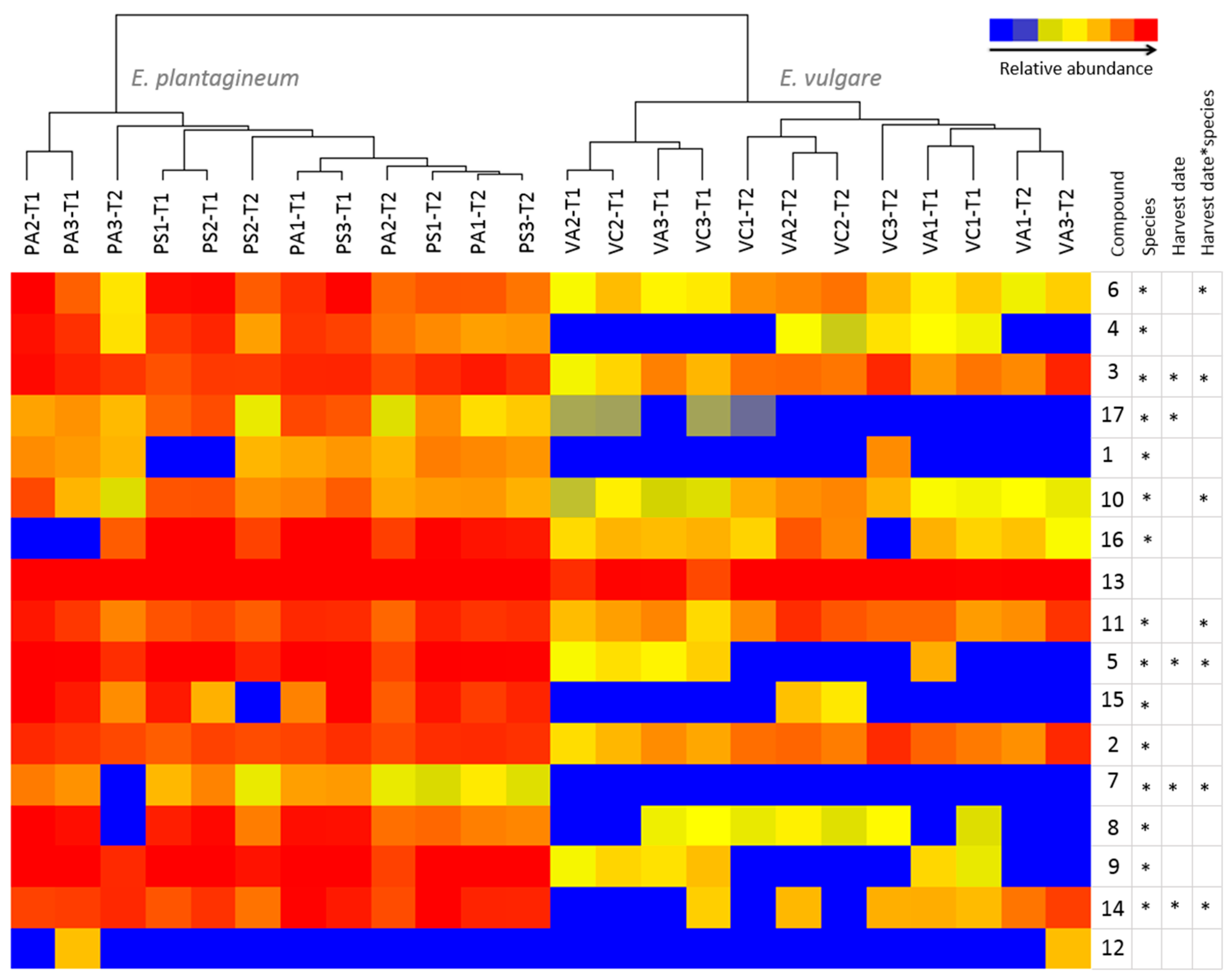
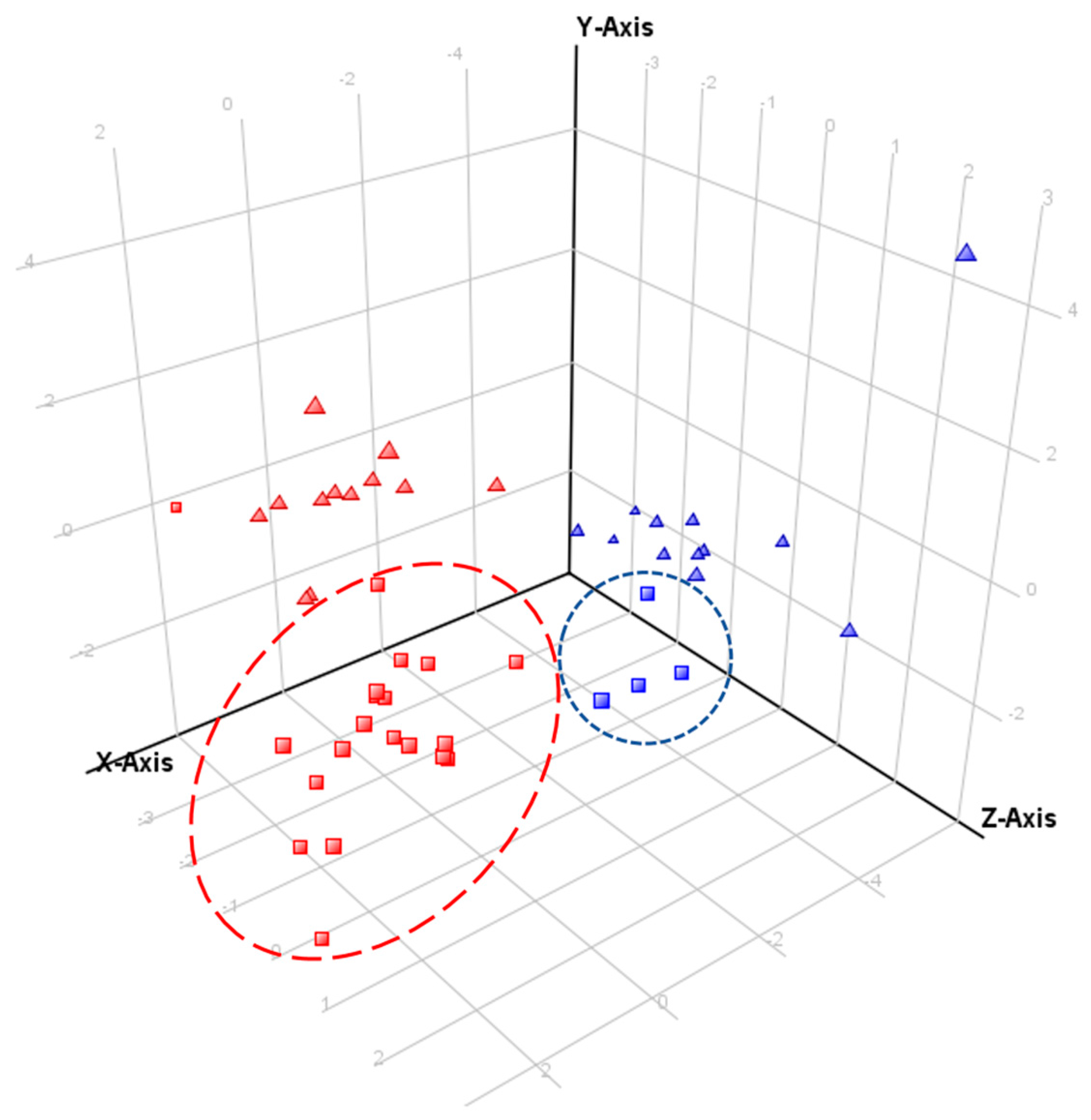
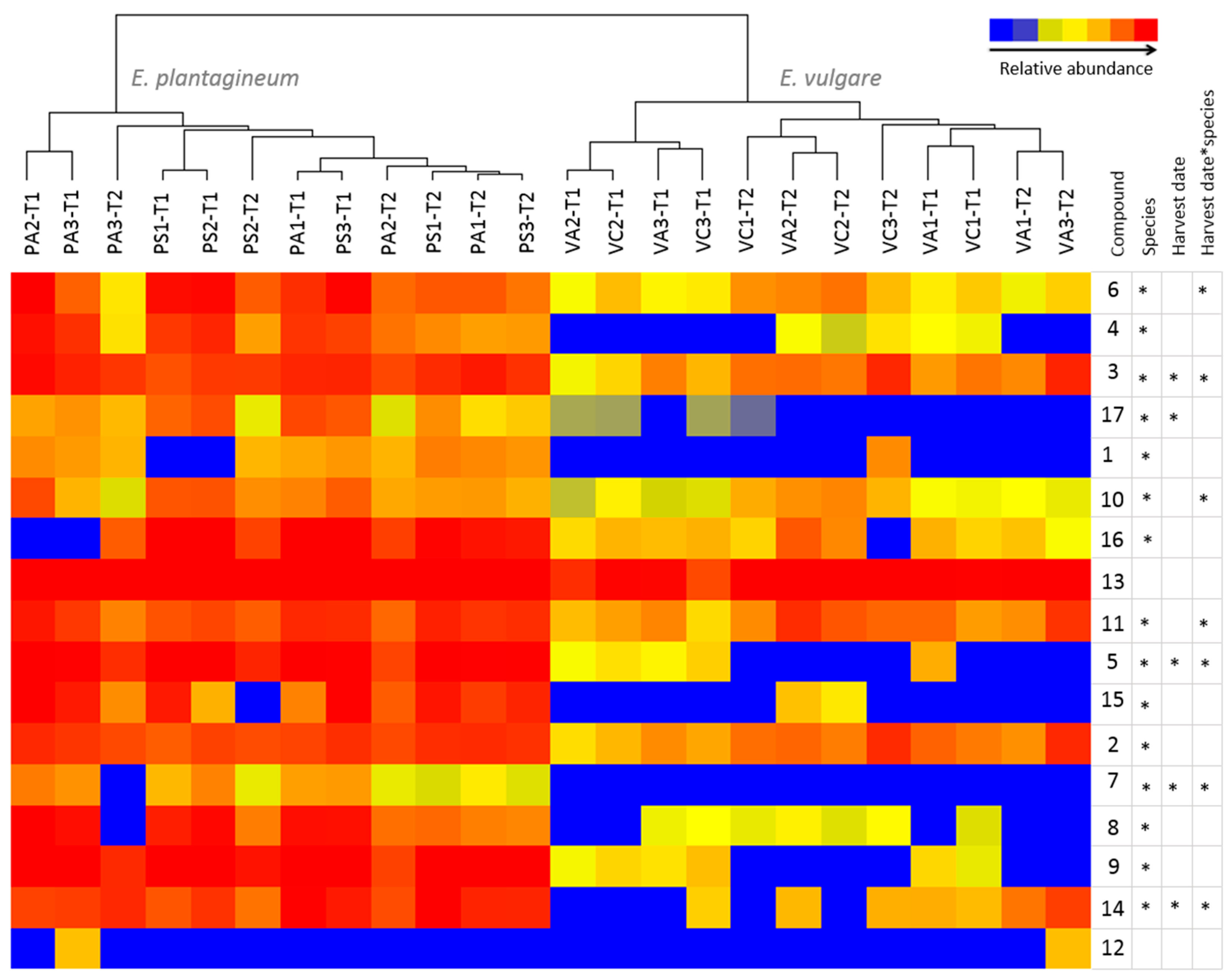
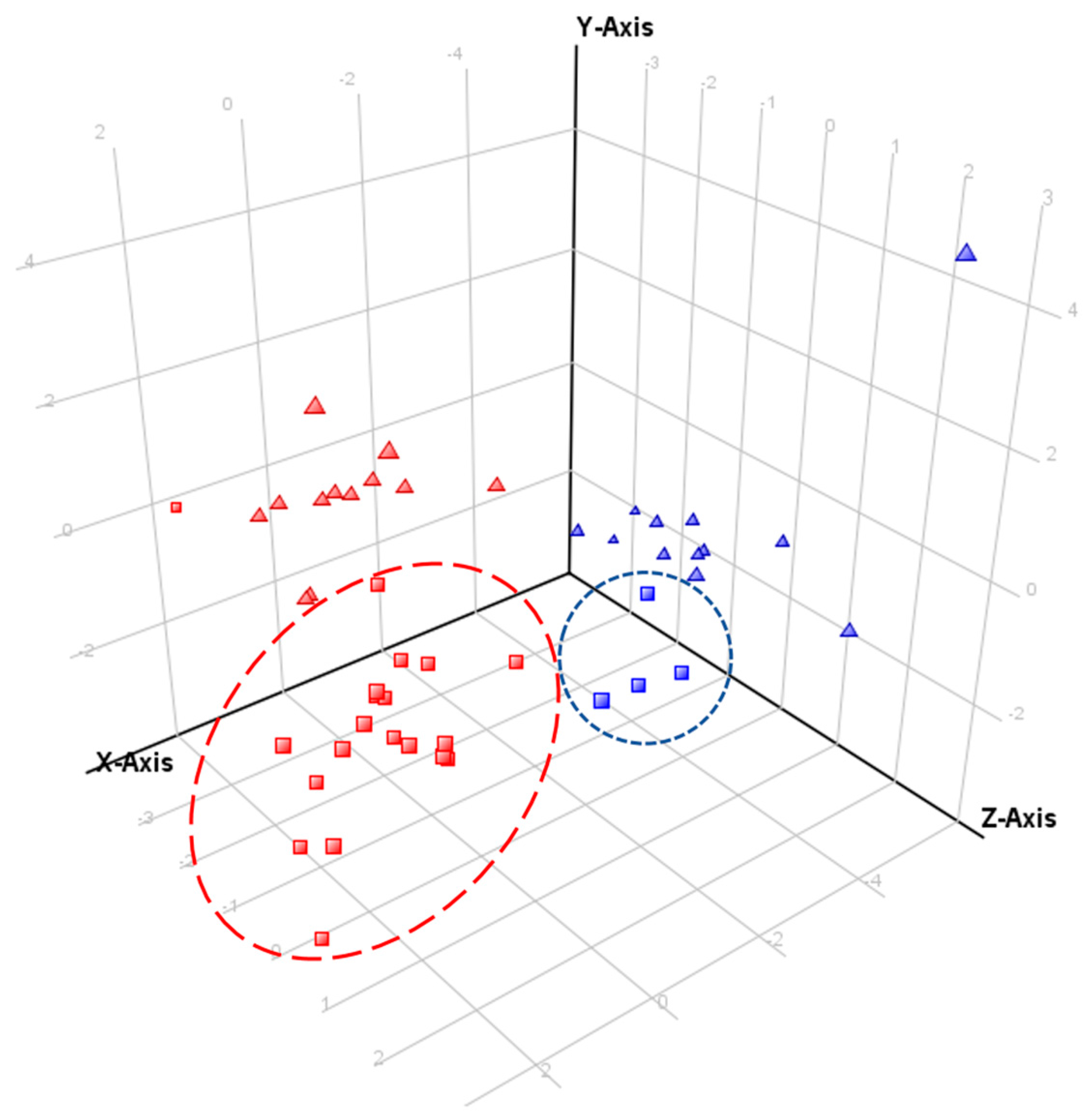
2.4. Comparative Profiling of Echium Plantagineum and E. Vulgare
3. Experimental Section
3.1. Chemicals and Standards
3.2. Instrumentation
3.3. Collection of Field-Grown Plant Material
3.4. Glasshouse Grown Plant Material
3.4.1. Germination and Plant Growth Conditions
3.4.2. Echium Plantagineum Phenological Study—Experiment 1
3.4.3. Comparative Profiling of Echium Species—Experiment 2
3.5. Extraction
3.5.1. Foliar Tissue Extraction
3.5.2. Solid Phase Extraction (SPE)
3.6. UHPLC-MS Analysis
3.7. Data Analysis
3.7.1. UHPLC-MS Data Analysis
3.7.2. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rochfort, S. Metabolomics reviewed: A new “omics” platform technology for systems biology and implications for natural products research. J. Nat. Prod. 2005, 68, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Arbona, V.; Manzi, M.; Ollas, C.D.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [PubMed]
- Weston, L.A.; Skoneczny, D.; Weston, P.A.; Weidenhamer, J.D. Metabolic profiling: An overview—New approaches for the detection and functional analysis of biologically active secondary plant products. J. Allelochem. Interact. 2015, 1, 15–27. [Google Scholar]
- Roessner, U.; Bacic, A. Metabolomics in plant research. Aust. Biochem. 2009, 40, 9–20. [Google Scholar]
- Jaiswal, Y.; Liang, Z.; Ho, A.; Wong, L.; Yong, P.; Chen, H.; Zhao, Z. Distribution of toxic alkaloids in tissues from three herbal medicine aconitum species using laser micro-dissection, UHPLC–QTOF MS and LC–MS/MS techniques. Phytochemistry 2014, 107, 155–174. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.C.; Kessell, A.; Weston, L.A. Secondary plant products causing photosensitization in grazing herbivores: Their structure, activity and regulation. Int. J. Mol. Sci. 2014, 15, 1441–1465. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comp. Funct. Genom. 2001, 2, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Roessner, U.; Luedemann, A.; Brust, D.; Fiehn, O.; Linke, T.; Willmitzer, L.; Fernie, A.R. Metabolic profiling allows comprehensive phenotyping of genetically or environmentally modified plant systems. Plant Cell Online 2001, 13, 11–29. [Google Scholar] [CrossRef]
- Schauer, N.; Fernie, A.R. Plant metabolomics: Towards biological function and mechanism. Trends Plant Sci. 2006, 11, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Thorpe, A.S.; Thelen, G.C.; Diaconu, A.; Callaway, R.M. Root exudate is allelopathic in invaded community but not in native community: Field evidence for the novel weapons hypothesis. J. Ecol. 2009, 97, 641–645. [Google Scholar] [CrossRef]
- Hartmann, T. Chemical ecology of pyrrolizidine alkaloids. Planta 1999, 207, 483–495. [Google Scholar] [CrossRef]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Inderjit; Wardle, D.A.; Karban, R.; Callaway, R.M. The ecosystem and evolutionary contexts of allelopathy. Trends Eecol. Evol. 2011, 26, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Vivanco, J.M.; Bais, H.P.; Stermitz, F.R.; Thelen, G.C.; Callaway, R.M. Biogeographical variation in community response to root allelochemistry: Novel weapons and exotic invasion. Ecol. Lett. 2004, 7, 285–292. [Google Scholar] [CrossRef]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Biochemistry and Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; Am. Soc. Plant Physiol.: Rockville, MD, USA, 2000; Volume 40, pp. 1250–1319. [Google Scholar]
- Wiedenfeld, H. Plants containing pyrrolizidine alkaloids: Toxicity and problems. Food Addit. Contam. Part A 2011, 28, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Ober, D.; Hartmann, T. Phylogenetic origin of a secondary pathway: The case of pyrrolizidine alkaloids. Plant Mol. Biol. 2000, 44, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Alali, F.Q.; Tahboub, Y.R.; Ibrahim, E.S.; Qandil, A.M.; Tawaha, K.; Burgess, J.P.; Sy, A.; Nakanishi, Y.; Kroll, D.J.; Oberlies, N.H. Pyrrolizidine alkaloids from Echium glomeratum (Boraginaceae). Phytochemistry 2008, 69, 2341–2346. [Google Scholar] [CrossRef] [PubMed]
- El-Shazly, A.; Wink, M. Diversity of pyrrolizidine alkaloids in the boraginaceae structures, distribution, and biological properties. Diversity 2014, 6, 188–282. [Google Scholar] [CrossRef]
- Peterson, J.; Jago, M. Toxicity of Echium plantagineum (Paterson’s curse). 2. Pyrrolizidine alkaloid poisoning in rats. Aust. J. Agric. Res. 1984, 35, 305–315. [Google Scholar] [CrossRef]
- Culvenor, C.; Jago, M.; Peterson, J.; Smith, L.; Payne, A.; Campbell, D.; Edgar, J.; Frahn, J. Toxicity of Echium plantagineum (Paterson’s curse). 1. Marginal toxic effects in merino wethers from long-term feeding. Aust. J. Agric. Res. 1984, 35, 293–304. [Google Scholar] [CrossRef]
- Zhu, L.; Ruan, J.-Q.; Li, N.; Fu, P.P.; Ye, Y.; Lin, G. A novel ultra-performance liquid chromatography hyphenated with quadrupole time of flight mass spectrometry method for rapid estimation of total toxic retronecine-type of pyrrolizidine alkaloids in herbs without requiring corresponding standards. Food Chem. 2016, 194, 1320–1328, in press. [Google Scholar] [CrossRef] [PubMed]
- Griffin, C.; O’Mahony, J.; Danaher, M.; Furey, A. Liquid chromatography tandem mass spectrometry detection of targeted pyrrolizidine alkaloids in honeys purchased within ireland. Food Anal. Methods 2015, 8, 18–31. [Google Scholar] [CrossRef]
- Cao, Y.; Colegate, S.M.; Edgar, J.A. Persistence of echimidine, a hepatotoxic pyrrolizidine alkaloid, from honey into mead. J. Food Compos. Anal. 2013, 29, 106–109. [Google Scholar] [CrossRef]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed]
- Macel, M. Attract and deter: A dual role for pyrrolizidine alkaloids in plant-insect interactions. Phytochem. Rev. 2011, 10, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Weston, P.; Weston, L.; Hildebrand, S. Metabolic profiling in Echium plantagineum: Presence of bioactive pyrrolizidine alkaloids and napthoquinones from accessions across southeastern australia. Phytochem. Rev. 2013, 12, 831–837. [Google Scholar] [CrossRef]
- Colegate, S.M.; Edgar, J.A.; Knill, A.M.; Lee, S.T. Solid-phase extraction and HPLC-MS profiling of pyrrolizidine alkaloids and their N-oxides: A case study of Echium plantagineum. Phytochem. Anal. 2005, 16, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, V.P.; Assimopoulou, A.N.; Couladouros, E.A.; Hepworth, D.; Nicolaou, K.C. The chemistry and biology of alkannin, shikonin, and related naphthazarin natural products. Angew. Chem. Int. Ed. 1999, 38, 270–301. [Google Scholar] [CrossRef]
- Skoneczny, D.; Weston, P.A.; Zhu, X.; Gurr, G.M.; Callway, R.M.; Weston, L.A. Metabolic profiling and identification of shikonins in root periderm of two invasive Echium spp. Weeds in australia. J. Phytochem. 2015. under review. [Google Scholar]
- Parsons, W.T.; Cuthbertson, E.G. Paterson’s curse, salvation jane. In Noxious Plants of Australia, 2nd ed.; CSIRO Publishing: Melbourne, Australia, 2001; pp. 325–330. [Google Scholar]
- Klemow, K.M.; Clements, D.R.; Threadgill, P.F.; Cavers, P.B. The biology of canadian weeds. 116. Echium vulgare L. Can. J. Plant Sci. 2002, 82, 235–248. [Google Scholar] [CrossRef]
- Grigulis, K.; Sheppard, A.W.; Ash, J.E.; Groves, R.H. The comparative demography of the pasture weed echium plantagineum between its native and invaded ranges. J. Appl. Ecol. 2001, 38, 281–290. [Google Scholar] [CrossRef]
- Piggin, C.M. The biology of australian weeds. 8. Echium plantagineum L. J. Aust. Inst. Agric. 1982, 48, 3–16. [Google Scholar]
- NRM South and the Southern Tasmanian Councils Authority. Availiable online: http://www.nrmsouth.org.au/wp-content/uploads/2014/10/patersons_curse.pdf (accessed on 29 October 2015).
- Mehrabani, M.; Ghannadi, A.; Sajjadi, E.; Ghassemi, N.; Shams-Ardakani, M. Toxic pyrrolizidine alkaloids of Echium amoenum fisch. And mey. DARU J. Pharm. Sci. 2006, 14, 122–127. [Google Scholar]
- Crews, C.; Berthiller, F.; Krska, R. Update on analytical methods for toxic pyrrolizidine alkaloids. Anal. Bioanal. Chem. 2010, 396, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Griffin, C.T.; Danaher, M.; Elliott, C.T.; Glenn Kennedy, D.; Furey, A. Detection of pyrrolizidine alkaloids in commercial honey using liquid chromatography-ion trap mass spectrometry. Food Chem. 2013, 136, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Culvenor, C.C.J.; Edgar, J.A.; Smith, L.W. Pyrrolizidine alkaloids in honey from Echium plantagineum L. J. Agric. Food Chem. 1981, 29, 958–960. [Google Scholar] [CrossRef] [PubMed]
- Culvenor, C. The alkaloids of Echium plantagineum L. Aust. J. Chem. 1956, 9, 512–520. [Google Scholar] [CrossRef]
- Werther, J. Echimidine. Available online: http://commons.wikimedia.org/wiki/File:Echimidine.png (accessed on 24 August 2015).
- Ryan, B. The role of leaf surface chemistry and morphology in plant defense of two related invasive weeds. In Echium Plantagineum and Echium Vulgare; Research, Charles Sturt University: Wagga Wagga, Australia, 2014. [Google Scholar]
- Ben Farhat, M.; Jordán, M.J.; Chaouch-Hamada, R.; Landoulsi, A.; Sotomayor, J.A. Changes in phenolic profiling and antioxidant capacity of Salvia aegyptiaca L. By-products during three phenological stages. LWT—Food Sci. Technol. 2015, 63, 791–797. [Google Scholar] [CrossRef]
- Molyneux, R.J.; Johnson, A.E. Extraordinary levels of production of pyrrolizidine alkaloids in senecio riddellii. J. Nat. Prod. 1984, 47, 1030–1032. [Google Scholar] [CrossRef] [PubMed]
- Frölich, C.; Ober, D.; Hartmann, T. Tissue distribution, core biosynthesis and diversification of pyrrolizidine alkaloids of the lycopsamine type in three Boraginaceae species. Phytochemistry 2007, 68, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- El-Shazly, A.; Sarg, T.; Ateya, A.; Abdel Aziz, A.; El-Dahmy, S.; Witte, L.; Wink, M. Pyrrolizidine alkaloids from Echium setosum and Echium vulgare. J. Nat. Prod. 1996, 59, 310–313. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Prentis, P.J.; Wilson, J.R.U.; Dormontt, E.E.; Richardson, D.M.; Lowe, A.J. Adaptive evolution in invasive species. Trends Plant Sci. 2008, 13, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Jamil, S.Z.; Weston, P.A.; Skoneczny, D.; Weston, L.A.; Gurr, G.M. Induction of chemical defences in the invasive weed Paterson’s curse (Echium plantaginuem) by a generalist herbivore insect and a specialist biological control agent. In Proceedings of the 7th Word Congress on Allelopathy, Vigo, Spain, 28 July–1 August 2014; Reigosa Roger, M., Sánchez-Moreiras, A., Eds.; Publisher: Vigo, Spain, 2014. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skoneczny, D.; Weston, P.A.; Zhu, X.; Gurr, G.M.; Callaway, R.M.; Weston, L.A. Metabolic Profiling of Pyrrolizidine Alkaloids in Foliage of Two Echium spp. Invaders in Australia—A Case of Novel Weapons? Int. J. Mol. Sci. 2015, 16, 26721-26737. https://doi.org/10.3390/ijms161125979
Skoneczny D, Weston PA, Zhu X, Gurr GM, Callaway RM, Weston LA. Metabolic Profiling of Pyrrolizidine Alkaloids in Foliage of Two Echium spp. Invaders in Australia—A Case of Novel Weapons? International Journal of Molecular Sciences. 2015; 16(11):26721-26737. https://doi.org/10.3390/ijms161125979
Chicago/Turabian StyleSkoneczny, Dominik, Paul A. Weston, Xiaocheng Zhu, Geoff M. Gurr, Ragan M. Callaway, and Leslie A. Weston. 2015. "Metabolic Profiling of Pyrrolizidine Alkaloids in Foliage of Two Echium spp. Invaders in Australia—A Case of Novel Weapons?" International Journal of Molecular Sciences 16, no. 11: 26721-26737. https://doi.org/10.3390/ijms161125979
APA StyleSkoneczny, D., Weston, P. A., Zhu, X., Gurr, G. M., Callaway, R. M., & Weston, L. A. (2015). Metabolic Profiling of Pyrrolizidine Alkaloids in Foliage of Two Echium spp. Invaders in Australia—A Case of Novel Weapons? International Journal of Molecular Sciences, 16(11), 26721-26737. https://doi.org/10.3390/ijms161125979