
Isolation and Culture of Pig Spermatogonial Stem Cells and Their in Vitro Differentiation into Neuron-Like Cells and Adipocytes




Abstract
:1. Introduction
2. Results
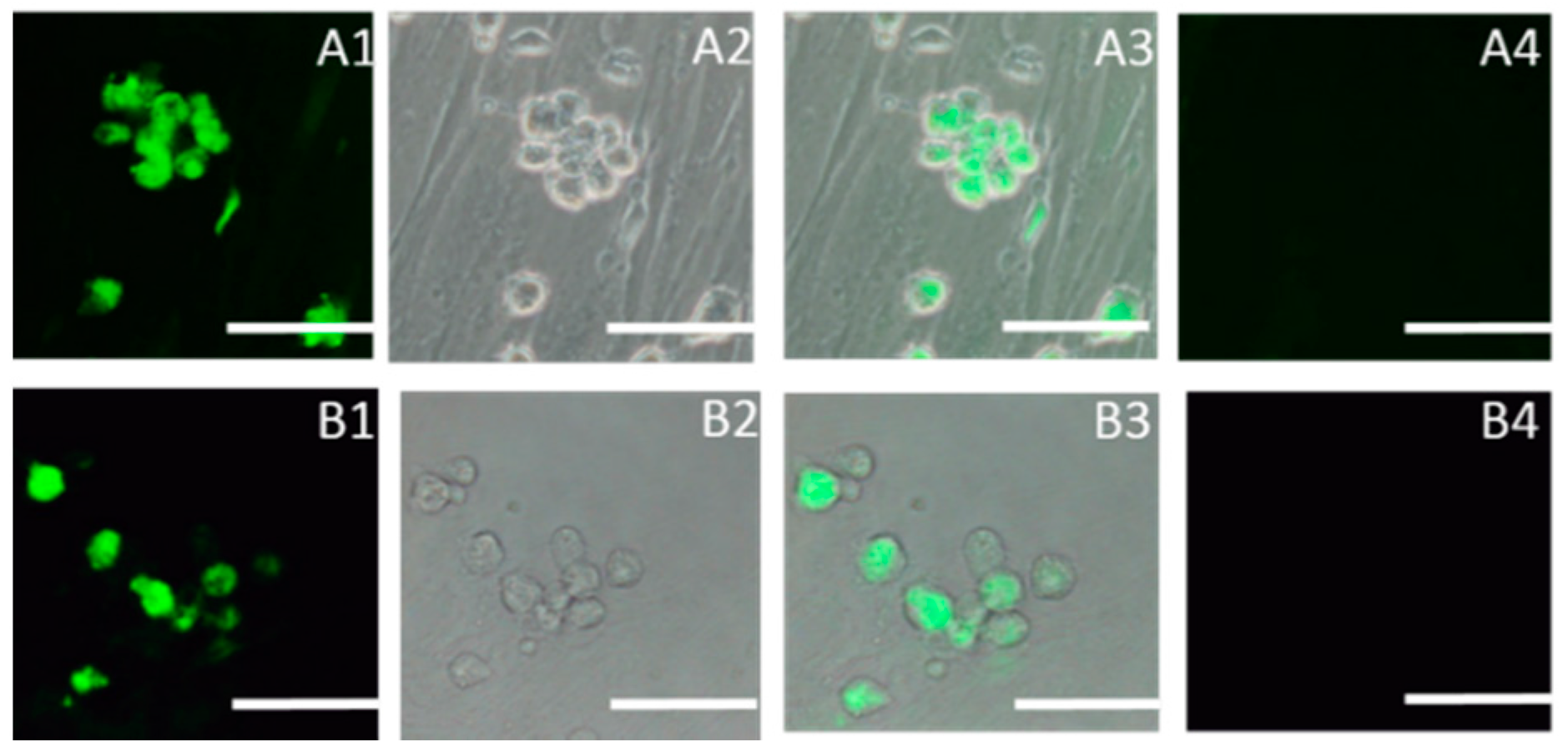
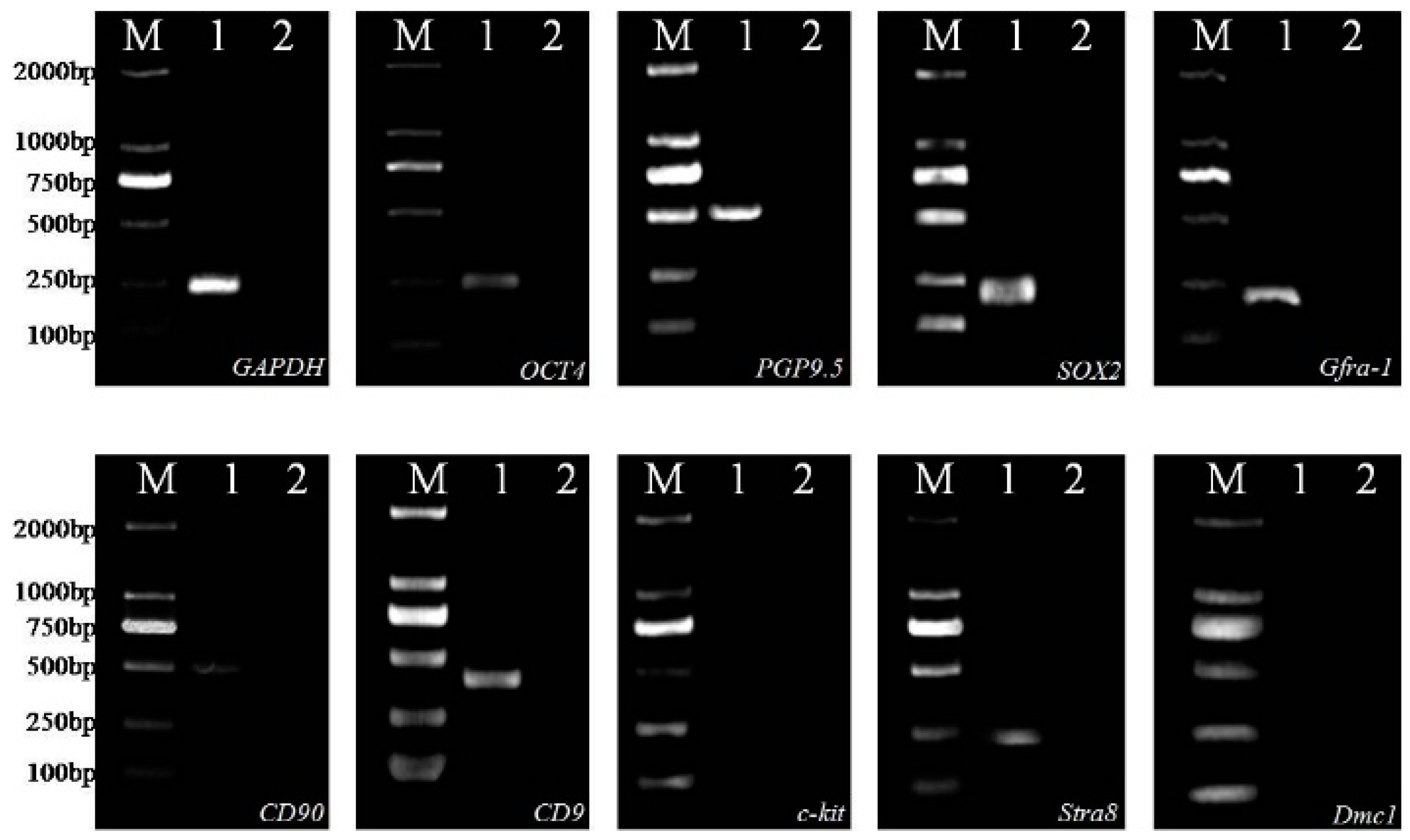
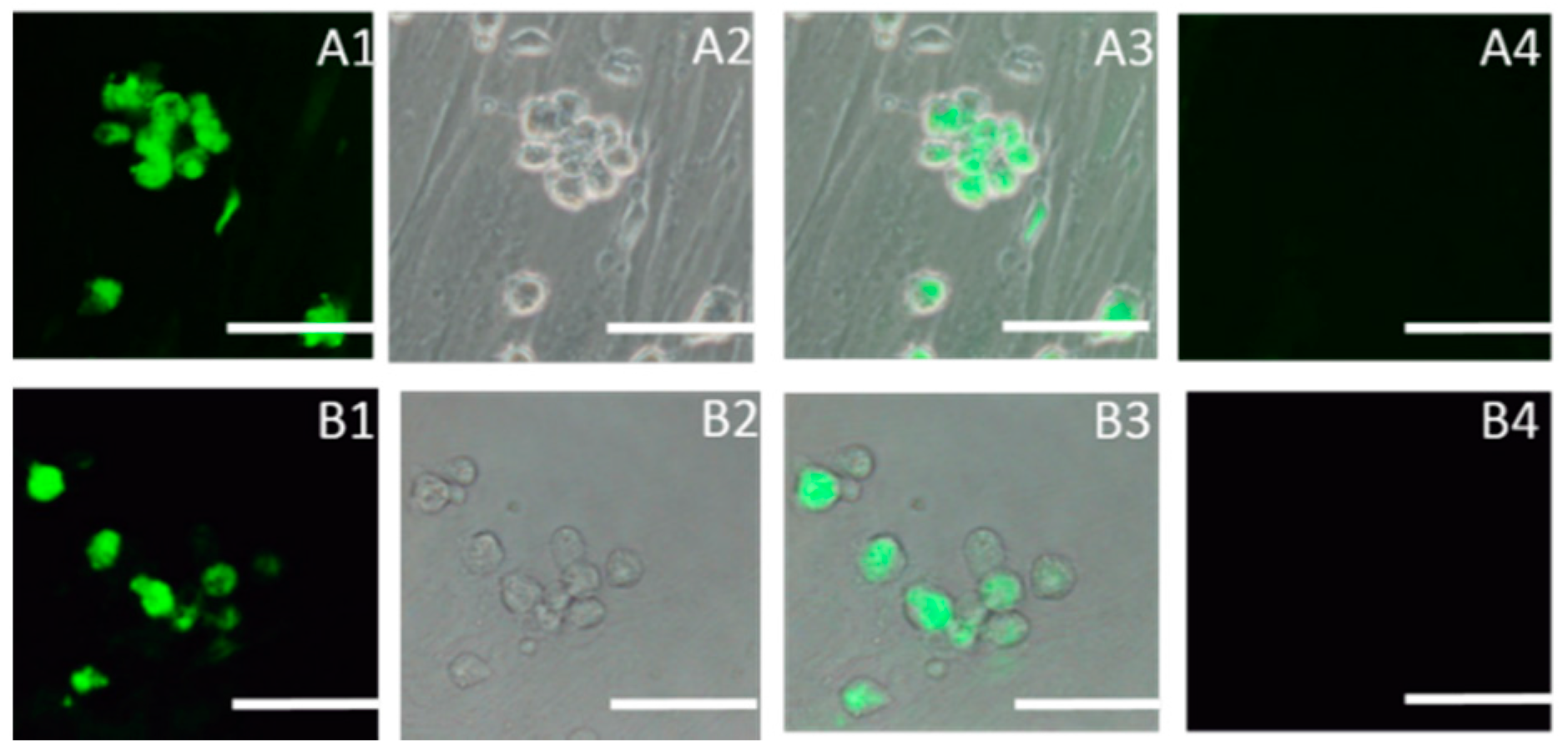
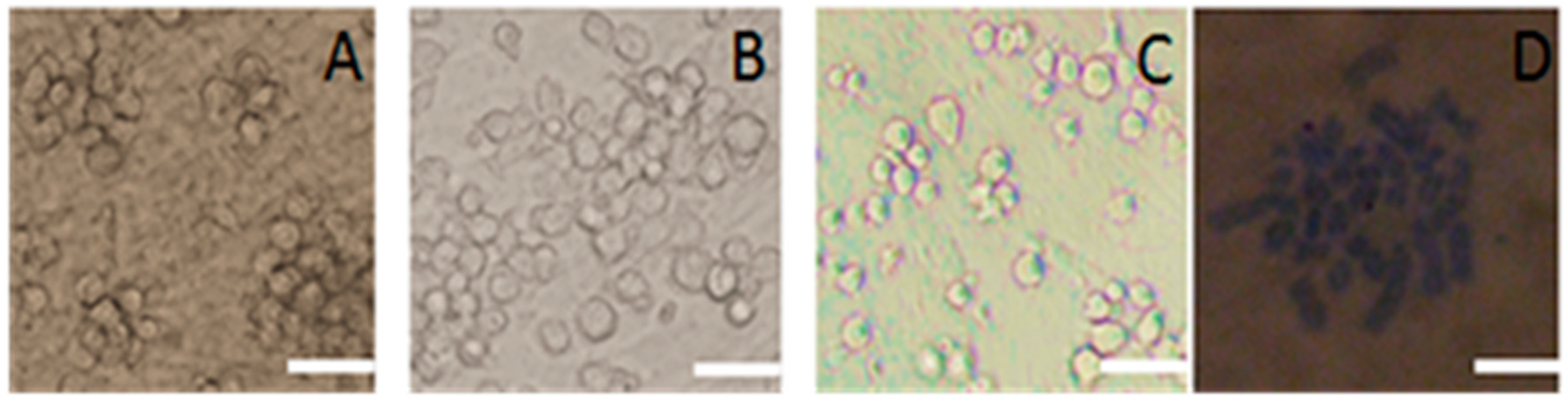
2.1. Identification of SSCs
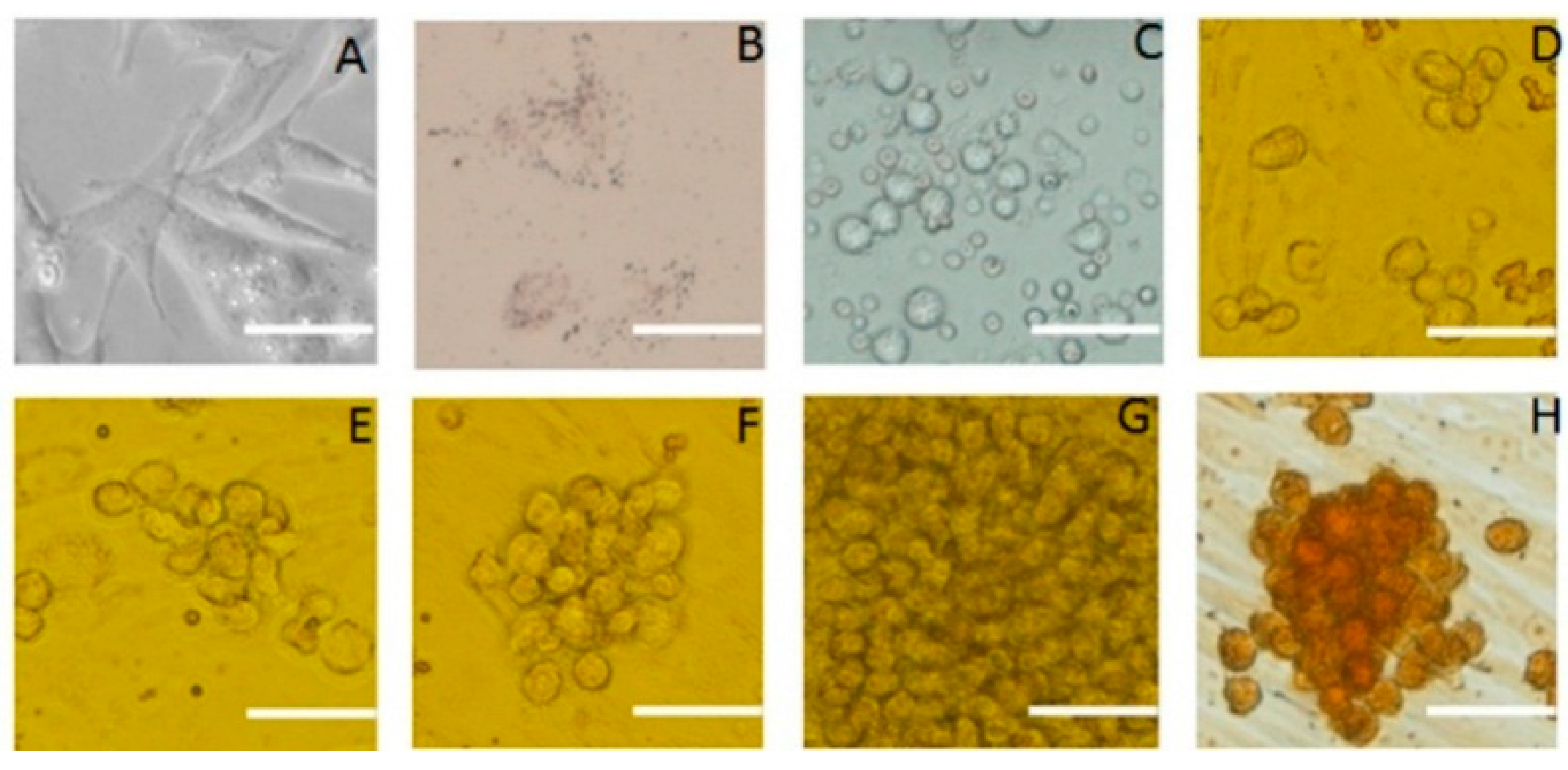
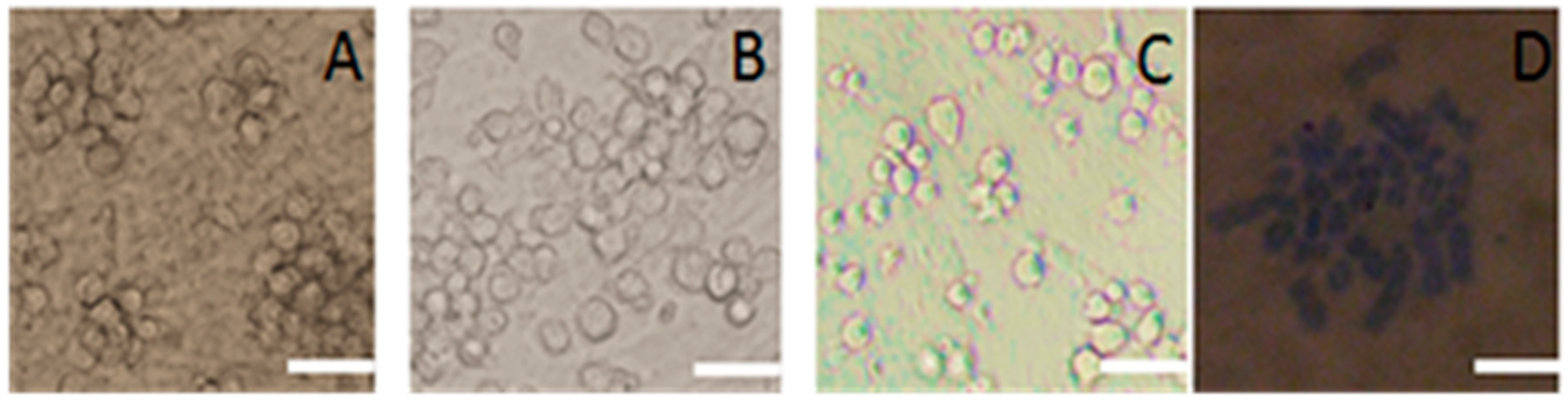
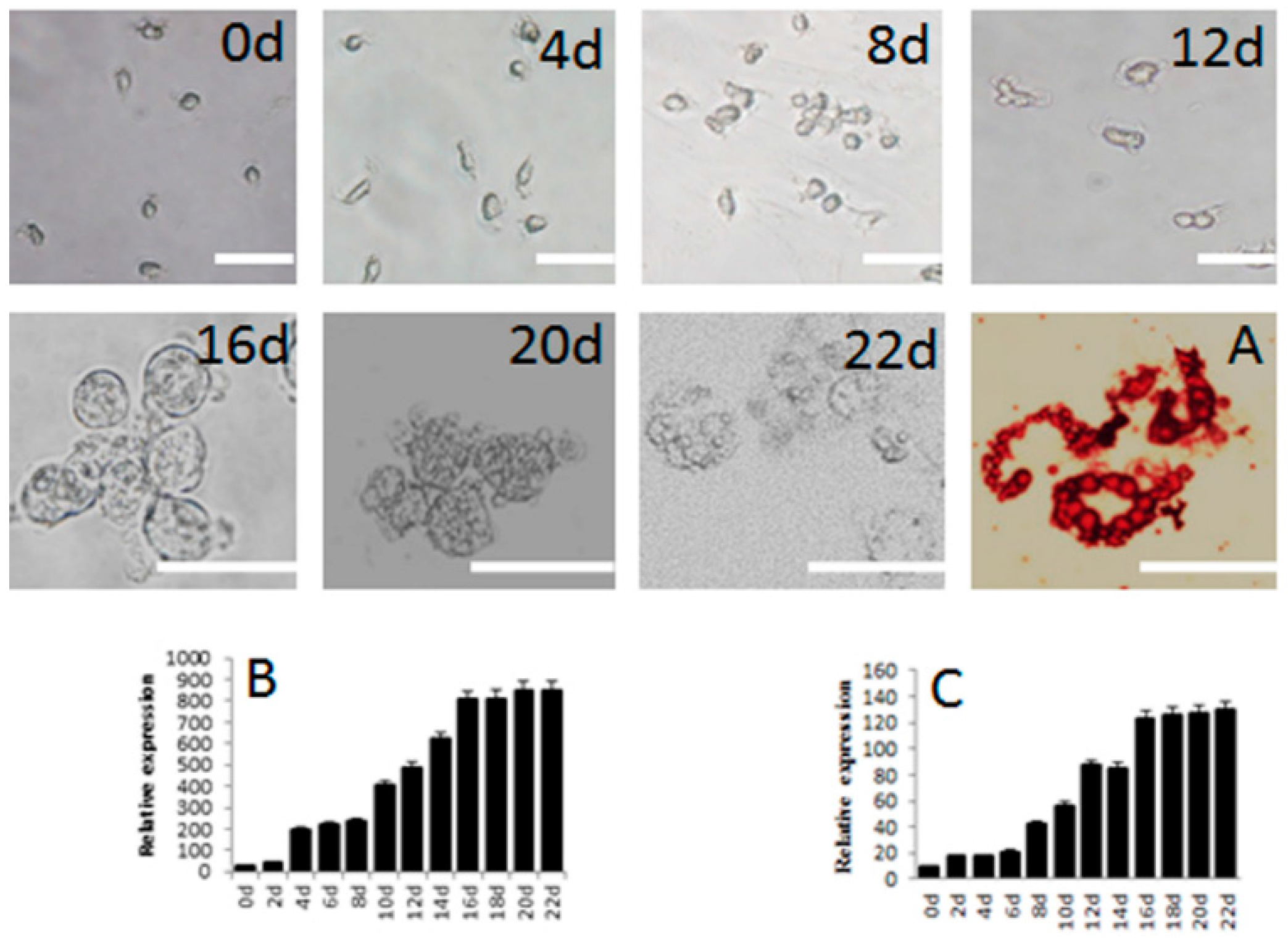
2.2. Subculture of Pig SSCs
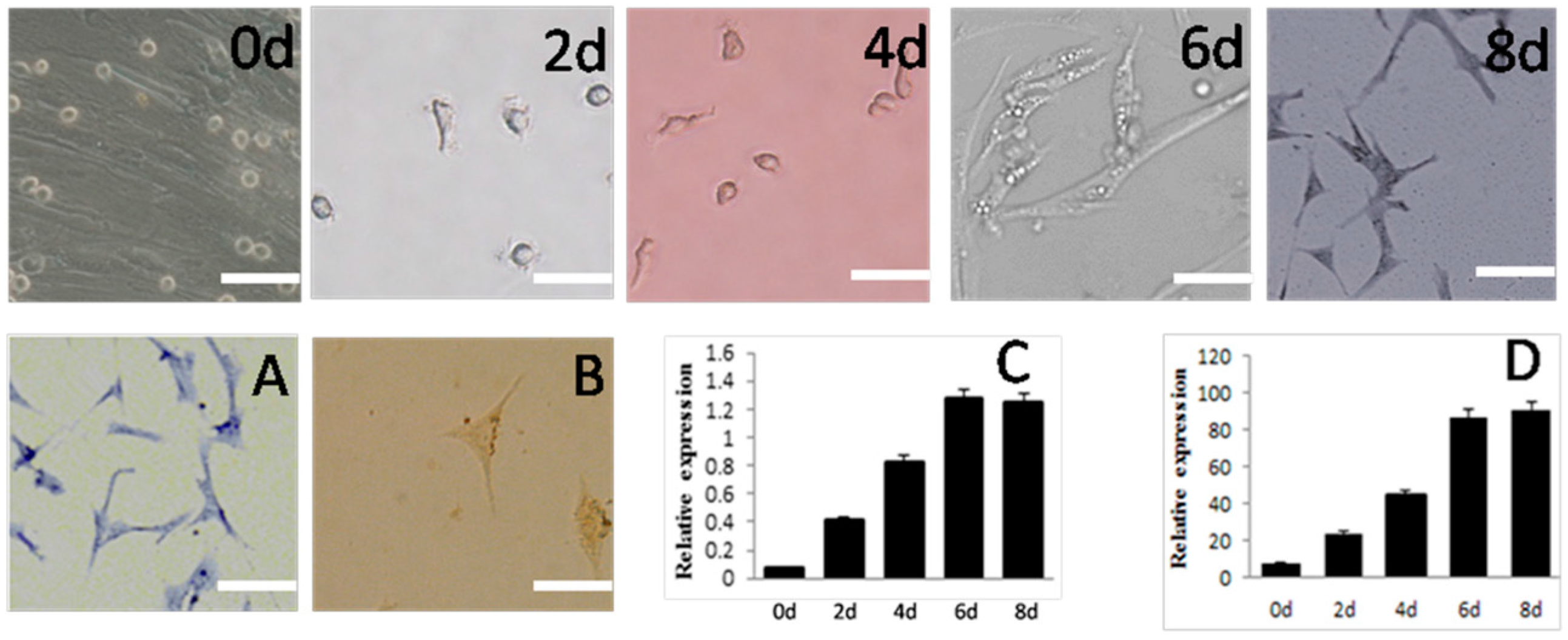
2.3. Differentiation of SSCS into Neuron-Like Cells
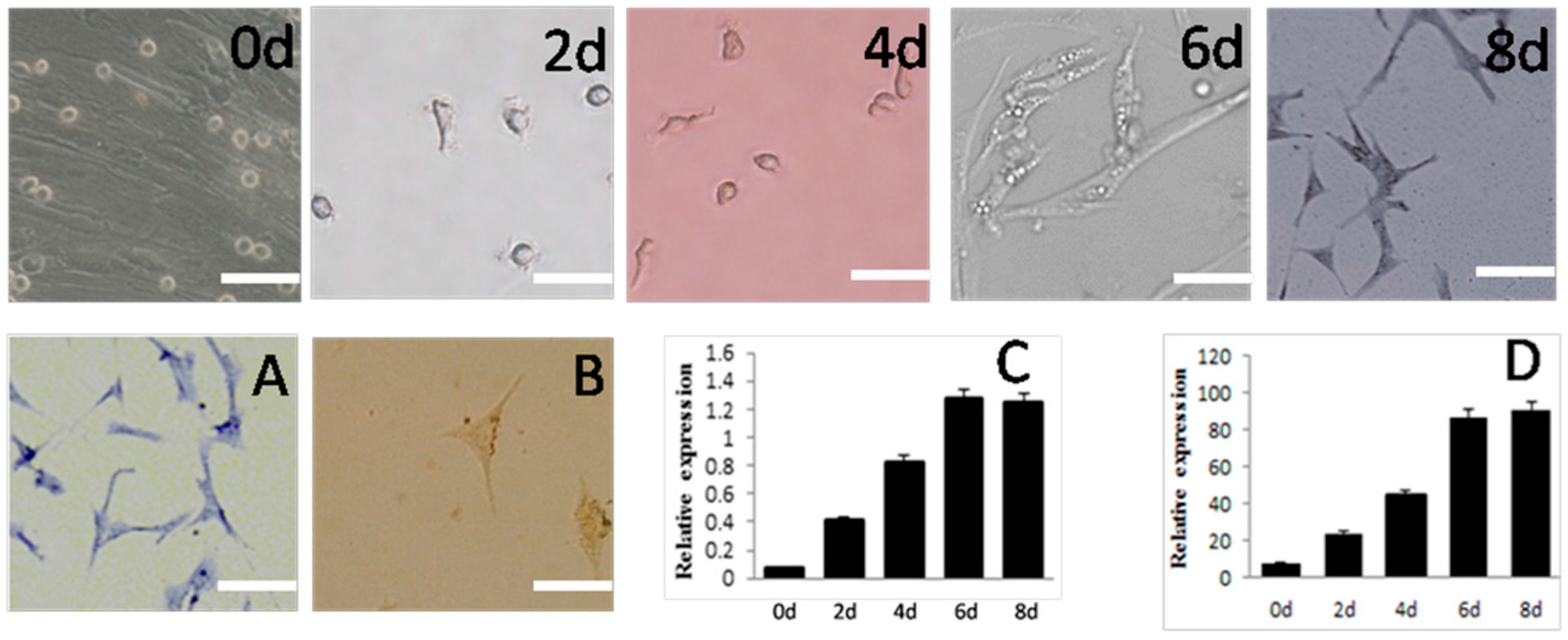
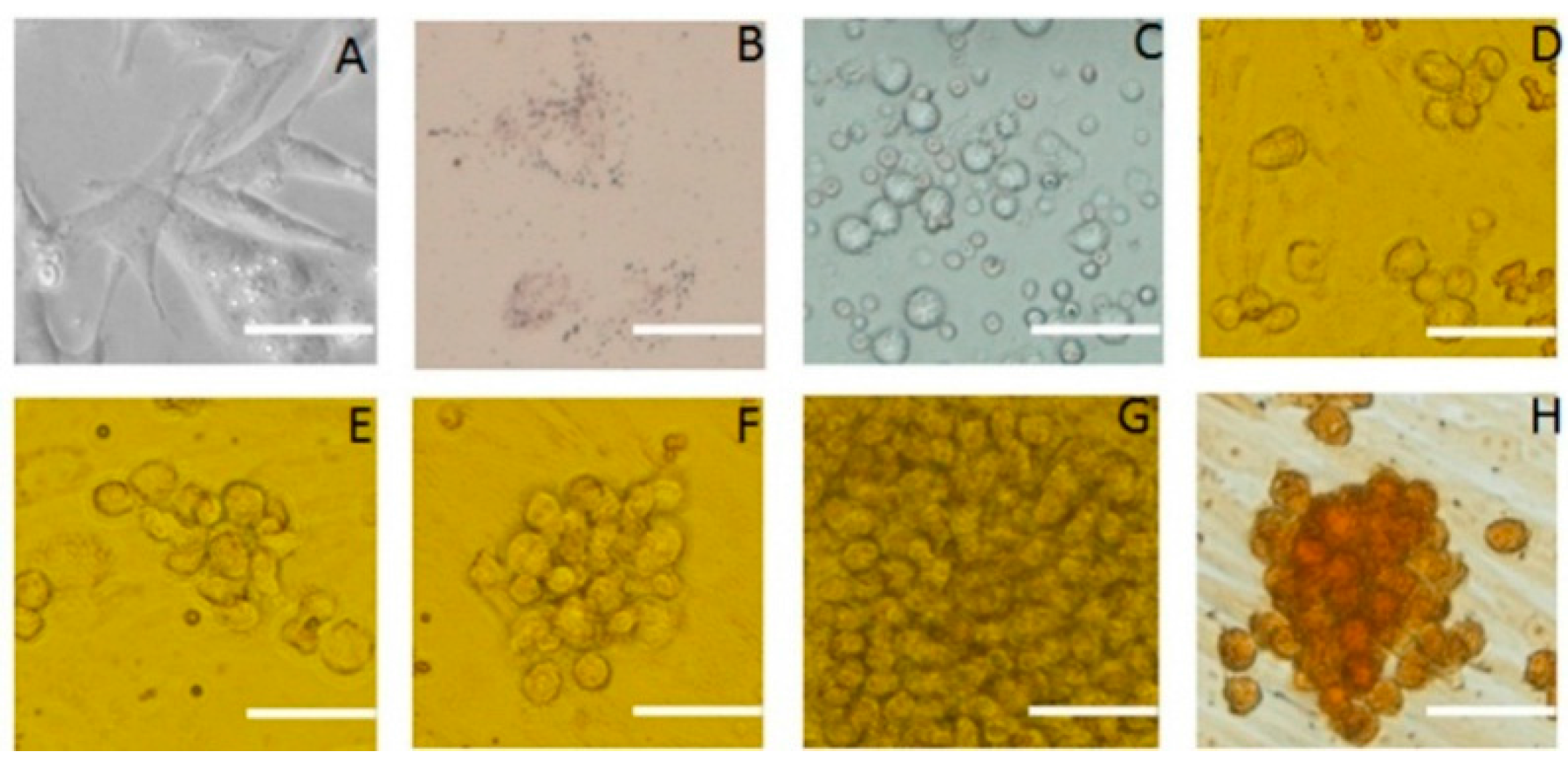
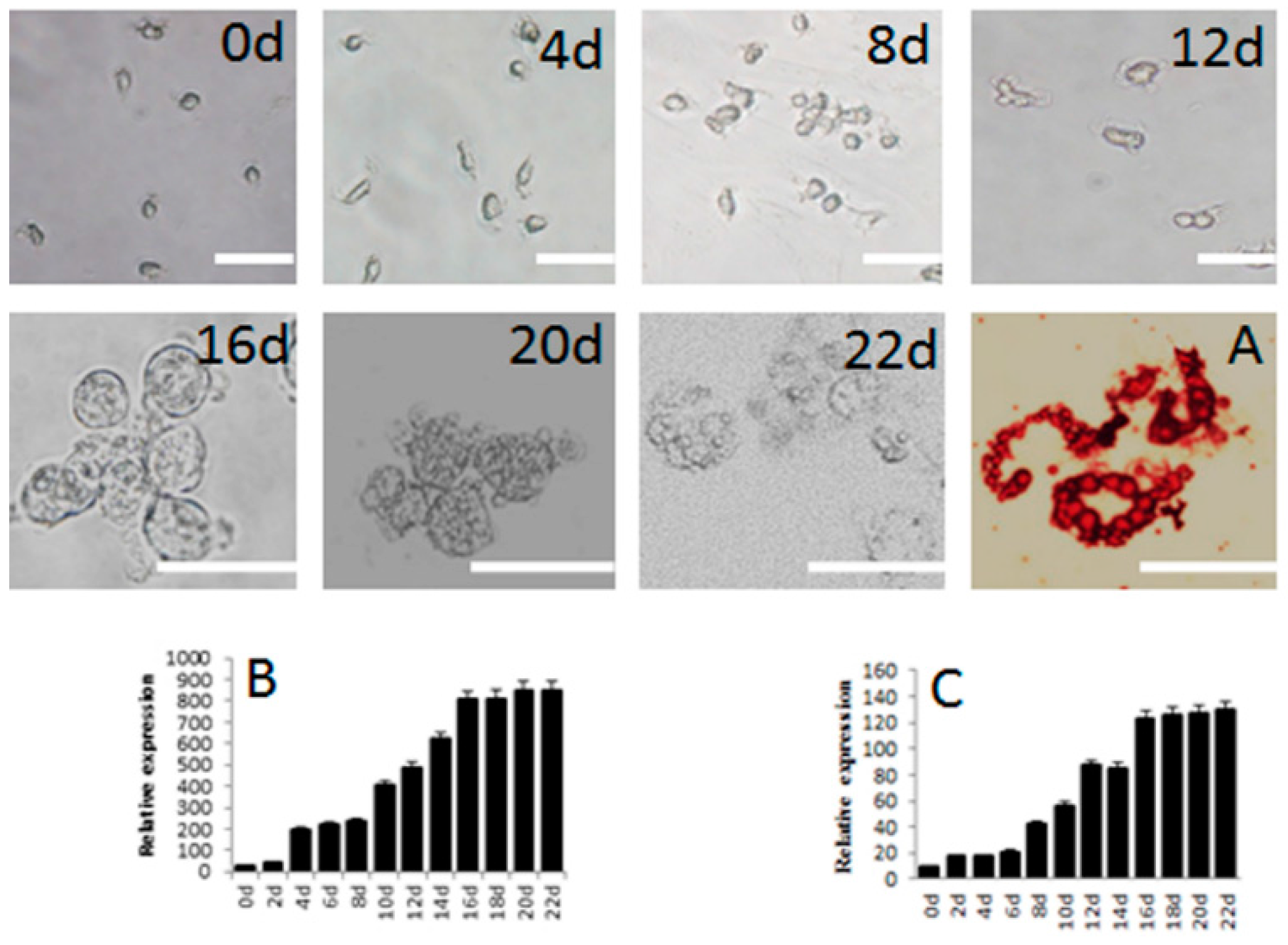
2.4. Differentiation of SSCs into Adipocytes
3. Discussion
4. Experimental Section
4.1. Animal Material
4.2. Isolated and Cultured SSCs
4.2.1. Preparation of Feeder Cells
4.2.2. Isolation and Culture of SSCs
4.3. Identification of SSCs
4.3.1. Staining for Alkaline Phosphatase Activity
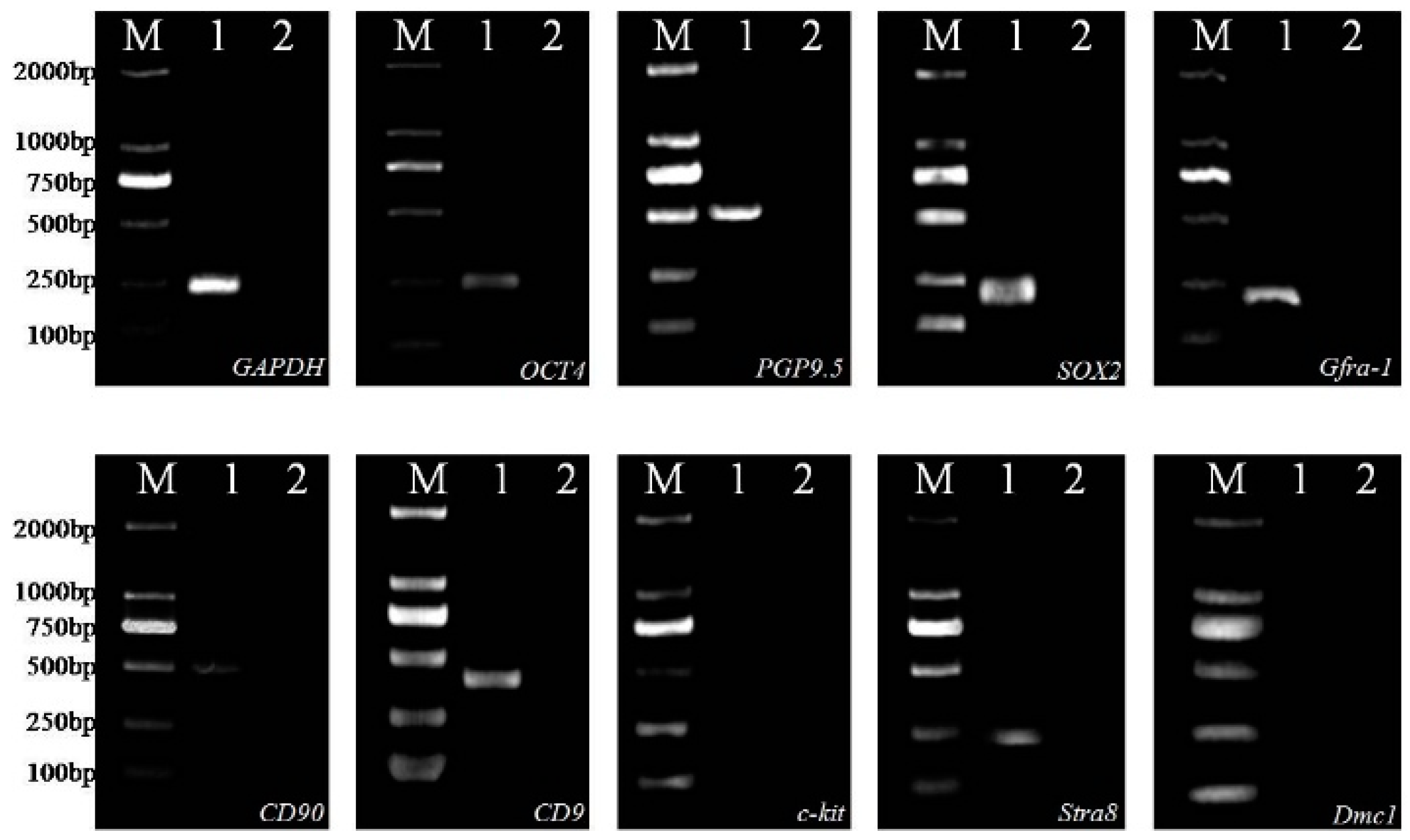
4.3.2. Identification of SSC-Specific Gene Expression by Reverse Transcription-PCR (RT-PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Product Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| OCT4 | GAGGAGTCCCAGGACATCAA | TCGTTGCGAATAGTAACTGC | 248 | 56 |
| SOX2 | CAAGATGCACAACTCGGAGA | TGCTGTAGCTGCAGTTGCTC | 236 | 58 |
| CD9 | TTCTGGCTCGCTGGGATT | CATCGGGAGGCTTGAGAGTA | 468 | 55 |
| PGP9.5 | GAGATGCTGAACAAAGTGCTG | CATGGTTCACCGGAAAAGG | 526 | 56 |
| CD90 | ACCATTGGCATCGCTCTCTT | GCCTTGTGGCTTCGTGTATCT | 512 | 56 |
| Gfra-1 | GAACGGAGGCGGCAGACCAT | AAGCCCAGAGTAGGCGAGGAG | 242 | 58 |
| Stra8 | ACCTCACAGCCTCAAAGTGG | CCTGGGGTTTCTGGAGTACA | 248 | 57 |
| Dmc1 | TGGCTGTGTTTGTGACCAAT | CCCAATTCCTCCAGCAGTTA | 323 | 55 |
| c-kit | gatgccttcaaggatttgga | atggaatctgaggccttcct | 181 | 56 |
| Nestin | GGACAGTGAGGACAAGGCA | AACACGGGCTCTATCACCTC | 281 | 57 |
| β-tubulin | cttcccacgtctccacttct | tcttgttctgcacgttgagc | 233 | 56 |
| PPARγ | TACCAAAGTGCCATCAAA | TGGAGTGGAAATGCTGGA | 110 | 56 |
| C/EBP-α | cacttgcagttccagatcgc | taccgacttcttggctttgc | 222 | 58 |
| GAPDH | TTCCACGGCACAGTCAAGG | TCACGCCCATCACAAACATG | 240 | 55 |
4.3.3. Immunocytochemistry of SSC-Specific Genes
4.3.4. Subculture and Karyotype Analysis
4.4. Induced Differentiation of SSCs
4.4.1. Differentiation of SSCs into Neuron-like Cells
4.4.2. Differentiation of SSCs into Adipocytes
4.5. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Rooij, D.G. Stem cells in the testis. Int. J. Exp. Pathol. 1998, 79, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Brinster, R.L. Germline stem cell transplantation and transgenesis. Science 2002, 296, 2174–2176. [Google Scholar] [CrossRef] [PubMed]
- Waheeb, R.; Hofmann, M.C. Human spermatogonial stem cells: A possible origin for spermatocytic seminoma. Int. J. Androl. 2011, 34, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Tegelenbosch, R.A.; de Rooij, D.G. A quantitative study of spermatogonial multiplication and stem cell renewal in the C3H/101 F1 hybrid mouse. Mutat Res. 1993, 290, 193–200. [Google Scholar] [CrossRef]
- Kubota, H.; Brinster, R.L. Technology insight: In vitro culture of spermatogonial stem cells and their potential therapeutic uses. Nat. Clin. Pract. Endocrinol. Metab. 2006, 2, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Aponte, P.M.; Soda, T.; Teerds, K.J.; Mizrak, S.C.; van de Kant, H.J.; de Rooij, D.G. Propagation of bovine spermatogonial stem cells in vitro. Reproduction 2008, 136, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Honaramooz, A. Efficient purification of neonatal porcine gonocytes with nycodenz and differential plating. Reprod. Fertil. Dev. 2011, 23, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yarahmadi, M.; Honaramooz, A. Development of novel strategies for the isolation of piglet testis cells with a high proportion of gonocytes. Reprod. Fertil. Dev. 2010, 22, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Park, J.E.; Kim, M.S.; Lee, K.Y.; Park, H.J.; Yun, J.I.; Choi, J.H.; Lee, E.; Lee, S.T. Development of a high-yield technique to isolate spermatogonial stem cells from porcine testes. J. Assist. Reprod. Genet. 2014, 31, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Bahadorani, M.; Hosseini, S.M.; Abedi, P.; Abbasi, H.; Nasr-Esfahani, M.H. Glial cell line-derived neurotrophic factor in combination with insulin-like growth factor 1 and basic fibroblast growth factor promote in vitro culture of goat spermatogonial stem cells. Growth Factors 2015, 33, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Pramod, R.K.; Mitra, A. In vitro culture and characterization of spermatogonial stem cells on sertoli cell feeder layer in goat (capra hircus). J. Assist. Reprod. Genet. 2014, 31, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Heidari, B.; Rahmati-Ahmadabadi, M.; Akhondi, M.M.; Zarnani, A.H.; Jeddi-Tehrani, M.; Shirazi, A.; Naderi, M.M.; Behzadi, B. Isolation, identification, and culture of goat spermatogonial stem cells using c-kit and PGP9.5 markers. J. Assist. Reprod. Genet. 2012, 29, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Lee, R.; Lee, W.Y.; Kim, D.H.; Chung, H.J.; Kim, J.H.; Kim, N.H.; Choi, S.H.; Song, H. Identification and in vitro derivation of spermatogonia in beagle testis. PLoS ONE 2014, 9, 109963. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, Y.; Qu, R.; He, Y.; Tian, X.; Zeng, W. Spermatogonial stem cells from domestic animals: Progress and prospects. Reproduction 2014, 147, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Sisakhtnezhad, S.; Bahrami, A.R.; Matin, M.M.; Dehghani, H.; Momeni-Moghaddam, M.; Boozarpour, S.; Farshchian, M.; Dastpak, M. The molecular signature and spermatogenesis potential of newborn chicken spermatogonial stem cells in vitro. Vitro Cell Dev. Biol. Anim. 2015, 51, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, S.M.; Costa, G.M.; de Franca, L.R. Biology and identity of fish spermatogonial stem cell. Gen.Comp. Endocrinol. 2014, 207, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Kanatsu-Shinohara, M.; Inoue, K.; Lee, J.; Yoshimoto, M.; Ogonuki, N.; Miki, H.; Baba, S.; Kato, T.; Kazuki, Y.; Toyokuni, S.; et al. Generation of pluripotent stem cells from neonatal mouse testis. Cell 2004, 119, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Guan, K.; Nayernia, K.; Maier, L.S.; Wagner, S.; Dressel, R.; Lee, J.H.; Nolte, J.; Wolf, F.; Li, M.; Engel, W.; et al. Pluripotency of spermatogonial stem cells from adult mouse testis. Nature 2006, 440, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.; Arauzo-Bravo, M.J.; Kim, J.; Stehling, M.; Scholer, H.R. Conversion of adult mouse unipotent germline stem cells into pluripotent stem cells. Nat. Protoc. 2010, 5, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Conrad, S.; Renninger, M.; Hennenlotter, J.; Wiesner, T.; Just, L.; Bonin, M.; Aicher, W.; Buhring, H.J.; Mattheus, U.; Mack, A.; et al. Generation of pluripotent stem cells from adult human testis. Nature 2008, 456, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Mizrak, S.C.; Chikhovskaya, J.V.; Sadri-Ardekani, H.; van Daalen, S.; Korver, C.M.; Hovingh, S.E.; Roepers-Gajadien, H.L.; Raya, A.; Fluiter, K.; de Reijke, T.M.; et al. Embryonic stem cell-like cells derived from adult human testis. Hum. Reprod. 2010, 25, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Qasemi-Panahi, B.; Tajik, P.; Movahedin, M.; Moghaddam, G.; Barzgar, Y.; Heidari-Vala, H. Differentiation of bovine spermatogonial stem cells into osteoblasts. Avicenna J. Med. Biotechnol. 2011, 3, 149–153. [Google Scholar] [PubMed]
- Thoma, E.C.; Wagner, T.U.; Weber, I.P.; Herpin, A.; Fischer, A.; Schartl, M. Ectopic expression of single transcription factors directs differentiation of a medaka spermatogonial cell line. Stem Cells Dev. 2011, 20, 1425–1438. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, X.Y.; Tian, Z.; Xiao, X.J.; Xu, Q.; Wei, C.X.; Y, F.; Sun, H.C.; Chen, G.H. Directional differentiation of chicken spermatogonial stem cells in vitro. Cytotherapy 2010, 12, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Nazm Bojnordi, M.; Movahedin, M.; Tiraihi, T.; Javan, M. A simple co-culture system for generation of embryonic stem-like cells from testis. Iran. Red Crescent Med. J. 2012, 14, 811–815. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Asadi, M.H.; Javanmardi, S.; Movahedin, M. Derivation of es-like cell from neonatal mouse testis cells in autologous sertoli cells co-culture system. Iran. J. Reprod. Med. 2014, 12, 37–46. [Google Scholar] [PubMed]
- Chen, Z.; Li, Z.; He, Z. Plasticity of male germline stem cells and their applications in reproductive and regenerative medicine. Asian J. Androl. 2015, 17, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Cooke, P.S.; Simon, L.; Nanjappa, M.K.; Medrano, T.I.; Berry, S.E. Plasticity of spermatogonial stem cells. Asian J. Androl. 2015, 17, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Dym, M.; He, Z.; Jiang, J.; Pant, D.; Kokkinaki, M. Spermatogonial stem cells: Unlimited potential. Reprod. Fertil. Dev. 2009, 21, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.X.; Chen, X.F. Updated dedifferentiation and pluripotency of spermatogonial stem cells. Zhonghua Nan Ke Xue 2009, 15, 387–389. [Google Scholar] [PubMed]
- Lee, G.; Kim, H.; Elkabetz, Y.; Al Shamy, G.; Panagiotakos, G.; Barberi, T.; Tabar, V.; Studer, L. Isolation and directed differentiation of neural crest stem cells derived from human embryonic stem cells. Nat. Biotechnol. 2007, 25, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi-Barough, S.; Massumi, M.; Kouchesfahani, H.M.; Ai, J. Derivation of pre-oligodendrocytes from human endometrial stromal cells by using overexpression of microrna 338. J. Mol. Neurosci. 2013, 51, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.M.; Li, F.Z.; Jiang, Z.L.; Sun, D.X.; Li, D.J.; Song, X.X. Isolation, culturationand differentiation of porcince bone marrow mesenchymal stem cells into adipocytes. Chin. J. Vet. Sci. 2015, 35, 91–95. [Google Scholar]
- Borjigin, U.; Davey, R.; Hutton, K.; Herrid, M. Expression of promyelocytic leukaemia zinc-finger in ovine testis and its application in evaluating the enrichment efficiency of differential plating. Reprod. Fertil. Dev. 2010, 22, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Dores, C.; Rancourt, D.; Dobrinski, I. Stirred suspension bioreactors as a novel method to enrich germ cells from pre-pubertal pig testis. Andrology 2015, 3, 590–597. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, D.G. The spermatogonial stem cell niche. Microsc. Res. Tech. 2009, 72, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Spermatogonial stem cells share some, but not all, phenotypic and functional characteristics with other stem cells. Proc. Natl. Acad. Sci. USA 2003, 100, 6487–6492. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, T.; Orwig, K.E.; Avarbock, M.R.; Brinster, R.L. Spermatogonial stem cell enrichment by multiparameter selection of mouse testis cells. Proc. Natl. Acad. Sci. USA 2000, 97, 8346–8351. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, T.; Avarbock, M.R.; Brinster, R.L. β1- and α6-integrin are surface markers on mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 1999, 96, 5504–5509. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Growth factors essential for self-renewal and expansion of mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16489–16494. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Bai, Y.; Li, S.; Wei, H.; Zhang, X.; Li, L.; Tian, X.C.; Jiang, Q.; Wang, C.; Qin, L.; et al. Characteristics of spermatogonial stem cells derived from neonatal porcine testis. Andrologia 2015, 47, 765–778. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, X.; Zhu, H.; Wang, D. Developments in techniques for the isolation, enrichment, main culture conditions and identification of spermatogonial stem cells. Cytotechnology 2015. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Megee, S.; Rathi, R.; Dobrinski, I. Protein gene product 9.5 is a spermatogonia-specific marker in the pig testis: Application to enrichment and culture of porcine spermatogonia. Mol. Reprod. Dev. 2006, 73, 1531–1540. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.Y.; Lee, K.H.; Heo, Y.T.; Kim, N.H.; Kim, J.H.; Moon, S.H.; Chung, H.J.; Yoon, M.J.; Song, H. Transcriptional coactivator undifferentiated embryonic cell transcription factor 1 expressed in spermatogonial stem cells: A putative marker of boar spermatogonia. Anim. Reprod. Sci. 2014, 150, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; He, Y.; An, J.; Qin, J.; Wang, Y.; Zhang, Y.; Tian, X.; Zeng, W. Thy1 is a surface marker of porcine gonocytes. Reprod. Fertil. Dev. 2014, 26, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Lee, W.Y.; Kim, J.H.; Yoon, M.J.; Kim, N.H.; Uhm, S.J.; Kim, D.H.; Chung, H.J.; Song, H. Characterization of GFRα-1-positive and GFRα-1-negative spermatogonia in neonatal pig testis. Reprod. Domest. Anim. 2013, 48, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.; Fujihara, M.; Minami, N.; Yamada, M.; Imai, H. Expression of nanog, but not POU5F1, points to the stem cell potential of primitive germ cells in neonatal pig testis. Reproduction 2008, 135, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, B.J.; Kim, B.G.; Lee, Y.A.; Kim, K.J.; Chung, H.J.; Hwang, S.; Woo, J.S.; Park, J.K.; Schmidt, J.A.; et al. Stage-specific embryonic antigen-1 expression by undifferentiated spermatogonia in the prepubertal boar testis. J. Anim. Sci. 2013, 91, 3143–3154. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Honaramooz, A. Characterization and quenching of autofluorescence in piglet testis tissue and cells. Anat. Res. Int. 2012, 2012, 820120. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Yongda, L.; Jun, M.; Yingyu, S.; Shaoju, Z.; Xinwen, Z.; Mingxue, Z. Culture and neural differentiation of rat bone marrow mesenchymal stem cells in vitro. Cell. Biol. Int. 2007, 31, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xu, Y.; Qin, T.; Wang, F.P.; Ma, L.L.; Luo, X.G.; Zhang, T.C. Myocardin-related transcription factor-a is a key regulator in retinoic acid-induced neural-like differentiation of adult bone marrow-derived mesenchymal stem cells. Gene 2013, 523, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, M.L.; Rao, A.J.; Demerdash, O.N.; Kalil, R.E. Expression of nestin by neural cells in the adult rat and human brain. PLoS ONE 2011, 6, 18535. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Johansson, C.B.; Eliasson, C.; Stakeberg, J.; Wallen, A.; Perlmann, T.; Lendahl, U.; Betsholtz, C.; Berthold, C.H.; Frisen, J. Abnormal reaction to central nervous system injury in mice lacking glial fibrillary acidic protein and vimentin. J. Cell Biol. 1999, 145, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Lendahl, U.; Zimmerman, L.B.; McKay, R.D. Cns stem cells express a new class of intermediate filament protein. Cell 1990, 60, 585–595. [Google Scholar] [CrossRef]
- Spira, M.E.; Oren, R.; Dormann, A.; Gitler, D. Critical calpain-dependent ultrastructural alterations underlie the transformation of an axonal segment into a growth cone after axotomy of cultured aplysia neurons. J. Comp. Neurol. 2003, 457, 293–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xiong, W.; Chen, Y.; Ma, Q.; Ma, J.; Ge, Y.; Han, D. Evaluation on the phagocytosis of apoptotic spermatogenic cells by sertoli cells in vitro through detecting lipid droplet formation by oil red o staining. Reproduction 2006, 132, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Boschi, F.; Rizzatti, V.; Zamboni, M.; Sbarbati, A. Models of lipid droplets growth and fission in adipocyte cells. Exp. Cell Res. 2015, 336, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [PubMed]
- Seo, J.B.; Noh, M.J.; Yoo, E.J.; Park, S.Y.; Park, J.; Lee, I.K.; Park, S.D.; Kim, J.B. Functional characterization of the human resistin promoter with adipocyte determination- and differentiation-dependent factor 1/sterol regulatory element binding protein 1c and ccaat enhancer binding protein-α. Mol. Endocrinol. 2003, 17, 1522–1533. [Google Scholar] [CrossRef] [PubMed]
- Pascual, G.; Fong, A.L.; Ogawa, S.; Gamliel, A.; Li, A.C.; Perissi, V.; Rose, D.W.; Willson, T.M.; Rosenfeld, M.G.; Glass, C.K. A sumoylation-dependent pathway mediates transrepression of inflammatory response genes by PPAR-γ. Nature 2005, 437, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Hamm, J.K.; Park, B.H.; Farmer, S.R. A role for c/ebpbeta in regulating peroxisome proliferator-activated receptor gamma activity during adipogenesis in 3t3-l1 preadipocytes. J. Biol. Chem. 2001, 276, 18464–18471. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPalpha induces adipogenesis through PPARγ: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, T.; Song, C.; Gao, B.; Zhang, Y. Cloning and expression characteristics of the pig stra8 gene. Int. J. Mol. Sci. 2014, 15, 12480–12494. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.Y. The Study of Isolation and Culture of Spermatogonial Stem Cells of Wzsp; Liaoning Normal University: Dalian, China, 2009. [Google Scholar]
- Wang, X.; Xue, C.; Liu, H.; Xu, Y.; Zhao, R.; Jiang, Z.; Dodson, M.V.; Chen, J. Differential display of expressed genes reveals a novel function of SFRS18 in regulation of intramuscular fat deposition. Int. J. Biol. Sci. 2009, 5, 28–33. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, W.H.; Sun, X.F.; Kong, S.; Jiang, Y.H.; Zheng, Y.H.; Yin, Y.F. Cultivation and karyotype analysis of the human embryonic stem cells HUES4. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2007, 24, 275–278. [Google Scholar] [PubMed]
- Li, B.C.; Tian, Z.Q.; Sun, M.; Xu, Q.; Wang, X.Y.; Qin, Y.R.; Xu, F.; Gao, B.; Wang, K.H.; Sun, H.C.; et al. Directional differentiation of chicken primordial germ cells into adipocytes, neuron-like cells, and osteoblasts. Mol. Reprod. Dev. 2010, 77, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Woodbury, D.; Schwarz, E.J.; Prockop, D.J.; Black, I.B. Adult rat and human bone marrow stromal cells differentiate into neurons. J. Neurosci. Res. 2000, 61, 364–370. [Google Scholar] [CrossRef]
- Liu, Q.; Cheng, G.; Wang, Z.; Zhan, S.; Xiong, B.; Zhao, X. Bone marrow-derived mesenchymal stem cells differentiate into nerve-like cells in vitro after transfection with brain-derived neurotrophic factor gene. Vitro Cell Dev. Biol. Anim. 2015, 51, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Bagher, Z.; Ebrahimi-Barough, S.; Azami, M.; Mirzadeh, H.; Soleimani, M.; Ai, J.; Nourani, M.R.; Joghataei, M.T. Induction of human umbilical wharton’s jelly-derived mesenchymal stem cells toward motor neuron-like cells. Vitro Cell Dev. Biol. Anim. 2015, 51, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Tara, S.; Krishnan, L.K. Bioengineered fibrin-based niche to direct outgrowth of circulating progenitors into neuron-like cells for potential use in cellular therapy. J. Neural Eng. 2015, 12, 36011. [Google Scholar] [CrossRef] [PubMed]
- MacDougald, O.A.; Lane, M.D. Transcriptional regulation of gene expression during adipocyte differentiation. Annu. Rev. Biochem. 1995, 64, 345–373. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chen, T.; Zhang, Y.; Li, B.; Xu, Q.; Song, C. Isolation and Culture of Pig Spermatogonial Stem Cells and Their in Vitro Differentiation into Neuron-Like Cells and Adipocytes. Int. J. Mol. Sci. 2015, 16, 26333-26346. https://doi.org/10.3390/ijms161125958
Wang X, Chen T, Zhang Y, Li B, Xu Q, Song C. Isolation and Culture of Pig Spermatogonial Stem Cells and Their in Vitro Differentiation into Neuron-Like Cells and Adipocytes. International Journal of Molecular Sciences. 2015; 16(11):26333-26346. https://doi.org/10.3390/ijms161125958
Chicago/Turabian StyleWang, Xiaoyan, Tingfeng Chen, Yani Zhang, Bichun Li, Qi Xu, and Chengyi Song. 2015. "Isolation and Culture of Pig Spermatogonial Stem Cells and Their in Vitro Differentiation into Neuron-Like Cells and Adipocytes" International Journal of Molecular Sciences 16, no. 11: 26333-26346. https://doi.org/10.3390/ijms161125958
APA StyleWang, X., Chen, T., Zhang, Y., Li, B., Xu, Q., & Song, C. (2015). Isolation and Culture of Pig Spermatogonial Stem Cells and Their in Vitro Differentiation into Neuron-Like Cells and Adipocytes. International Journal of Molecular Sciences, 16(11), 26333-26346. https://doi.org/10.3390/ijms161125958