A Novel Poly-Naphthol Compound ST104P Suppresses Angiogenesis by Attenuating Matrix Metalloproteinase-2 Expression in Endothelial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
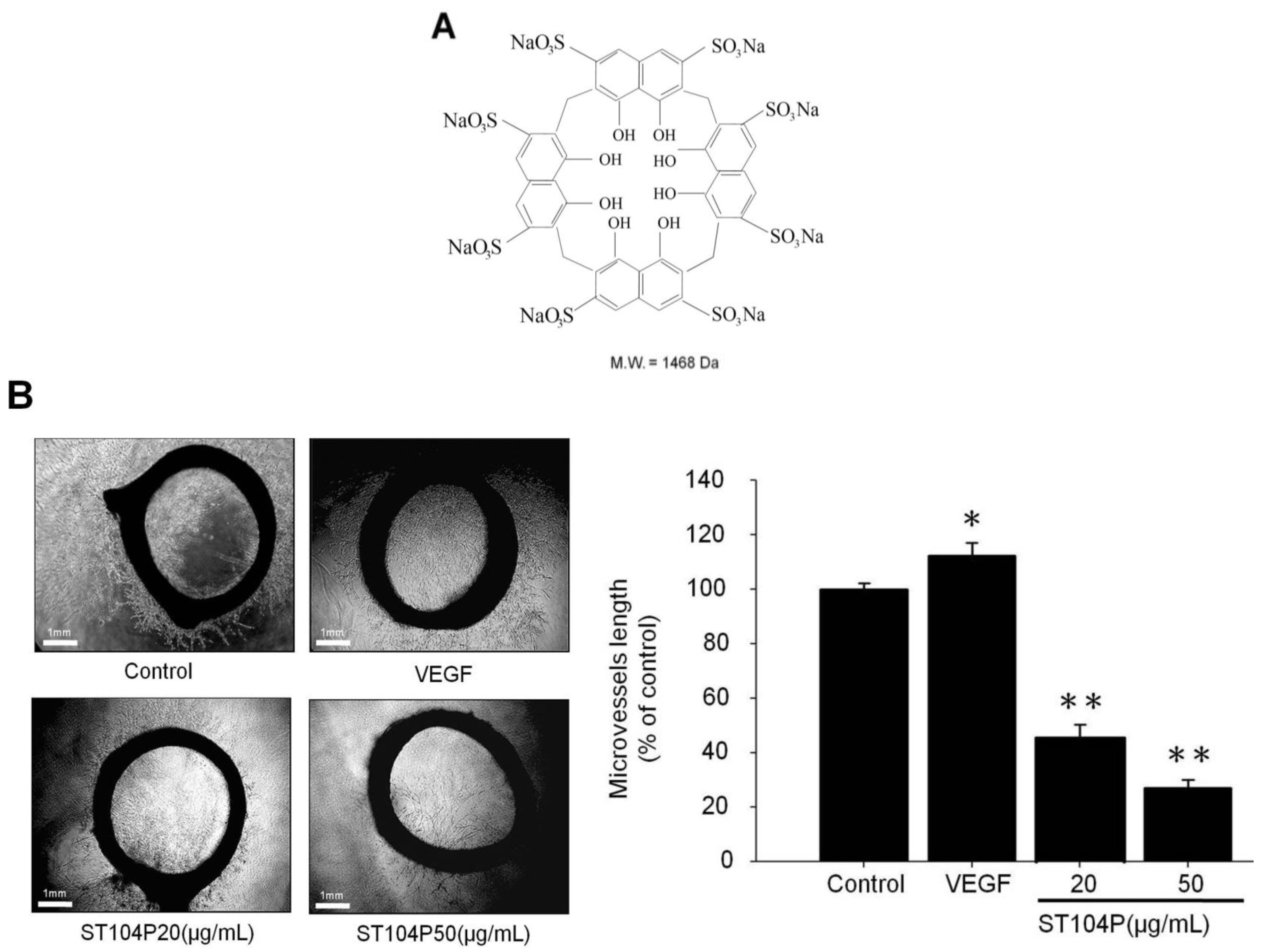
2.1. ST104P Suppressed the Microvessels Sprouting in Organotypic Aortic Rings
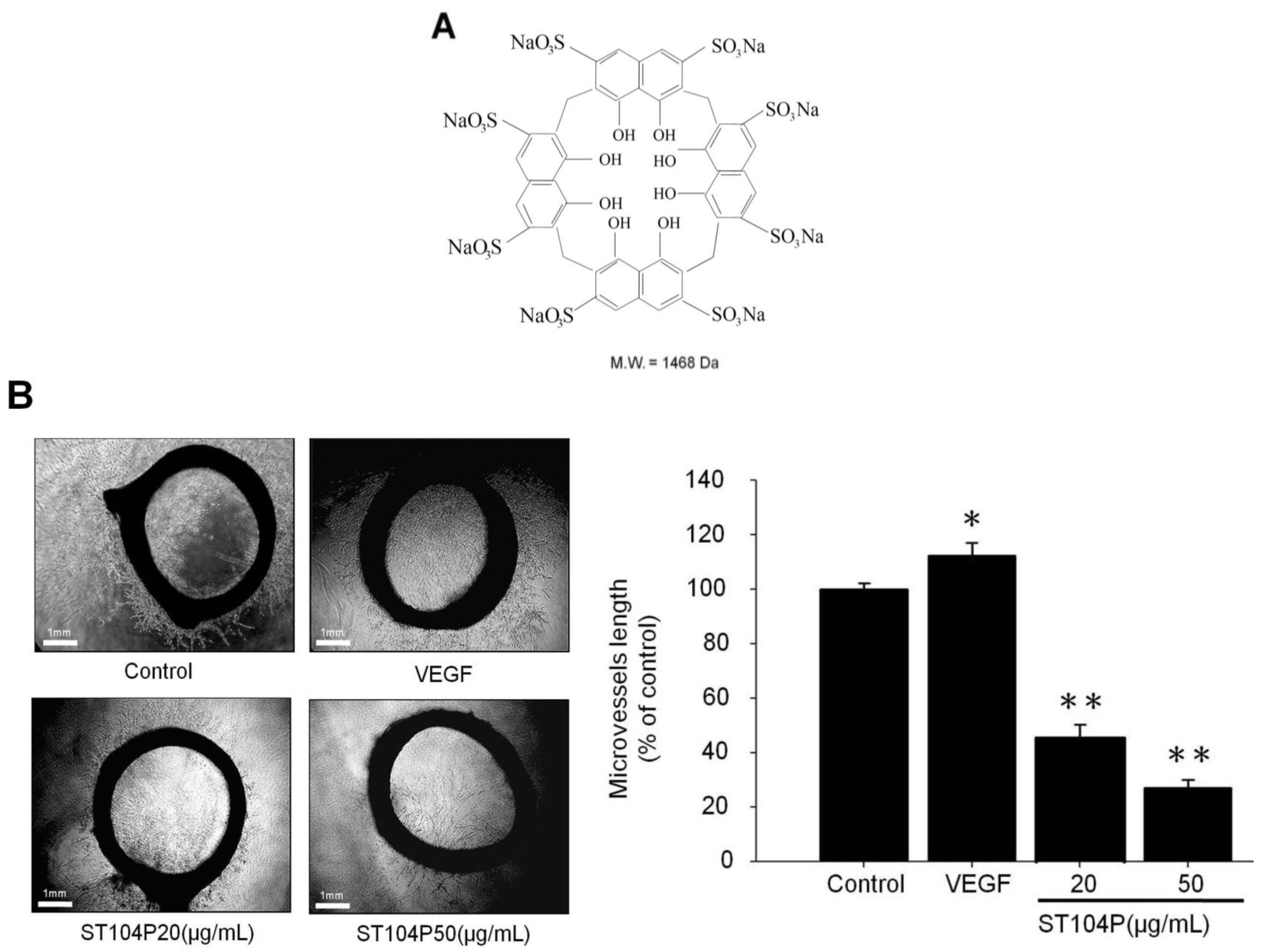
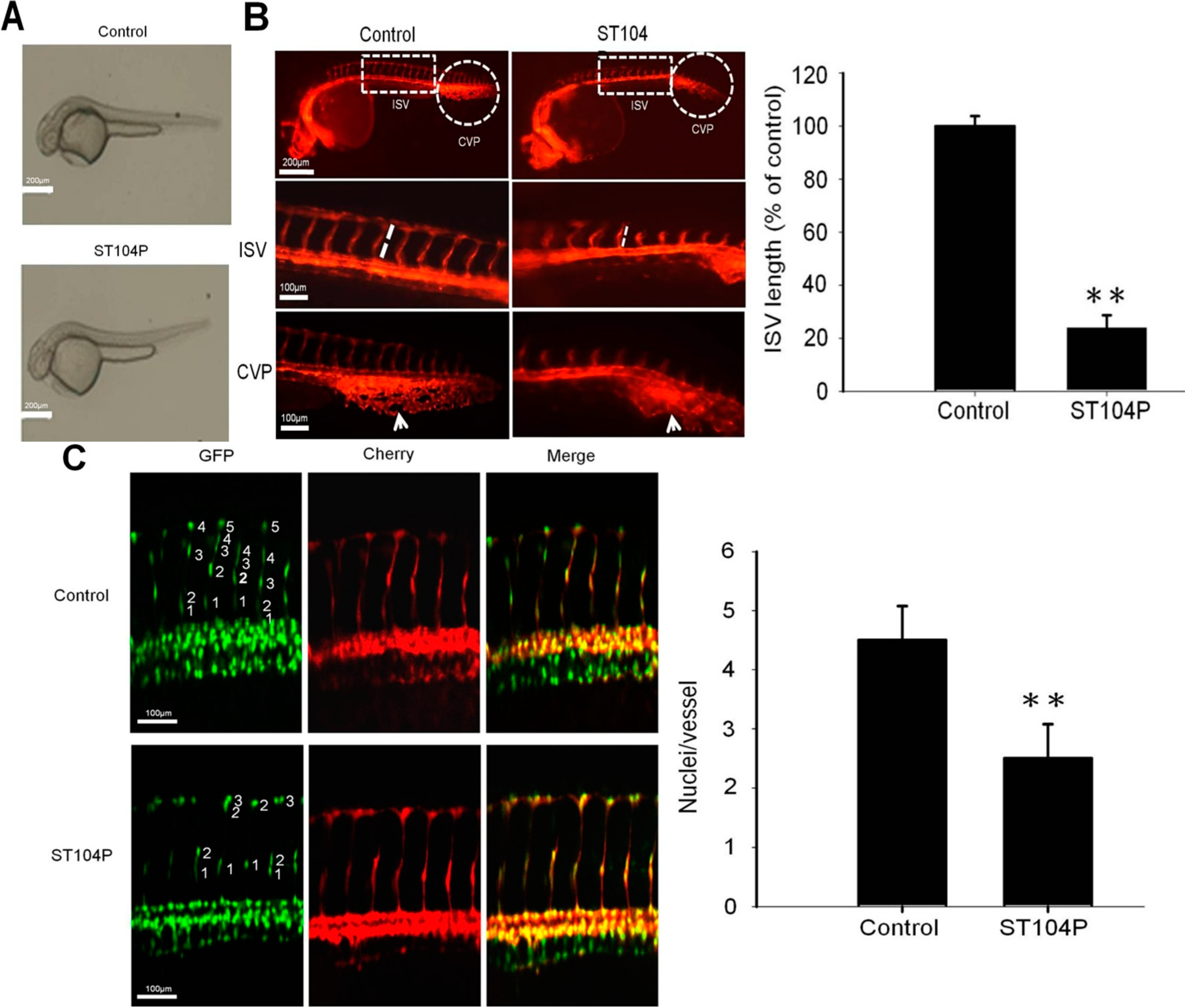
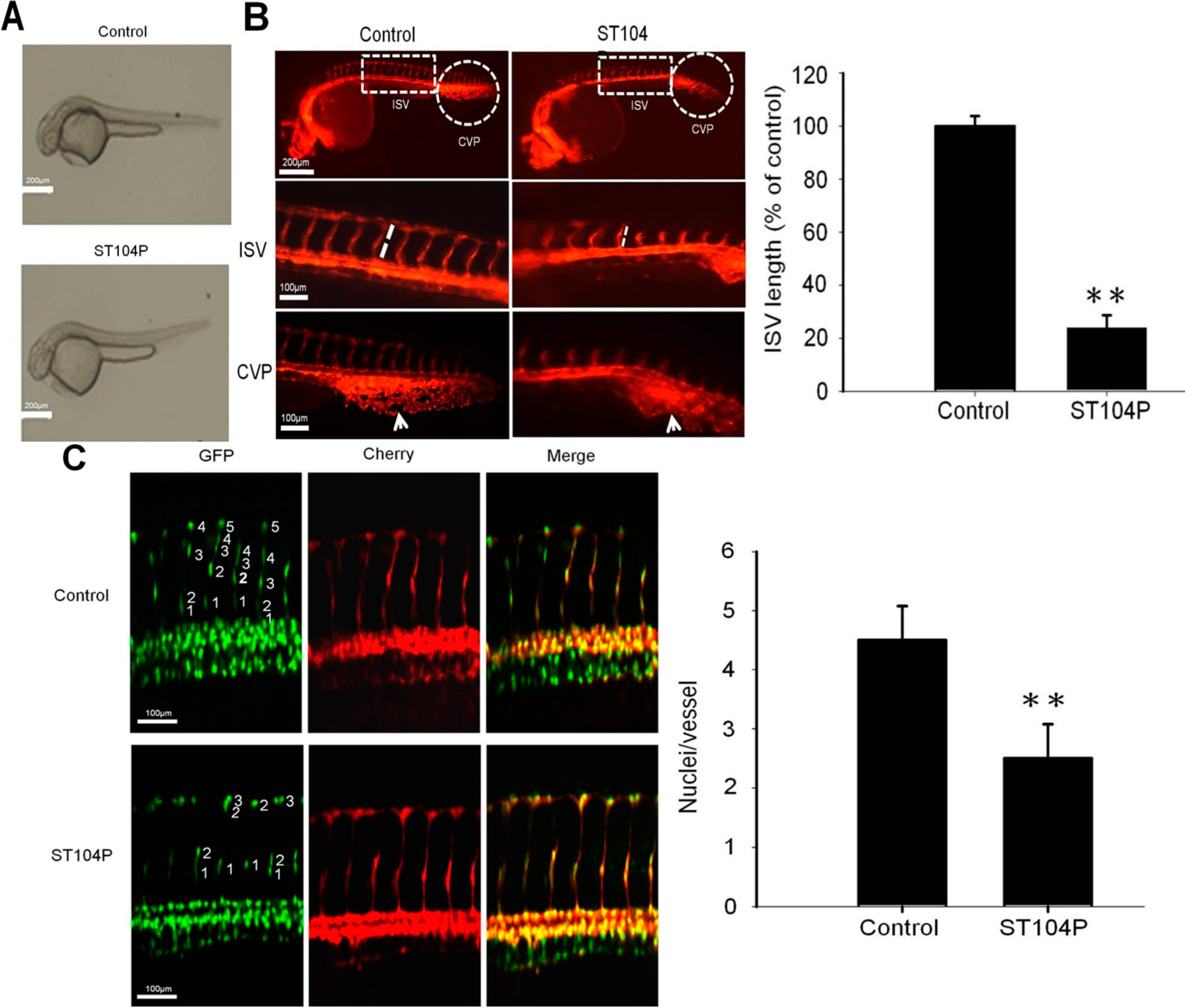
2.2. ST104P Perturbed the Vessels’ Development in Transgenic Zebrafish
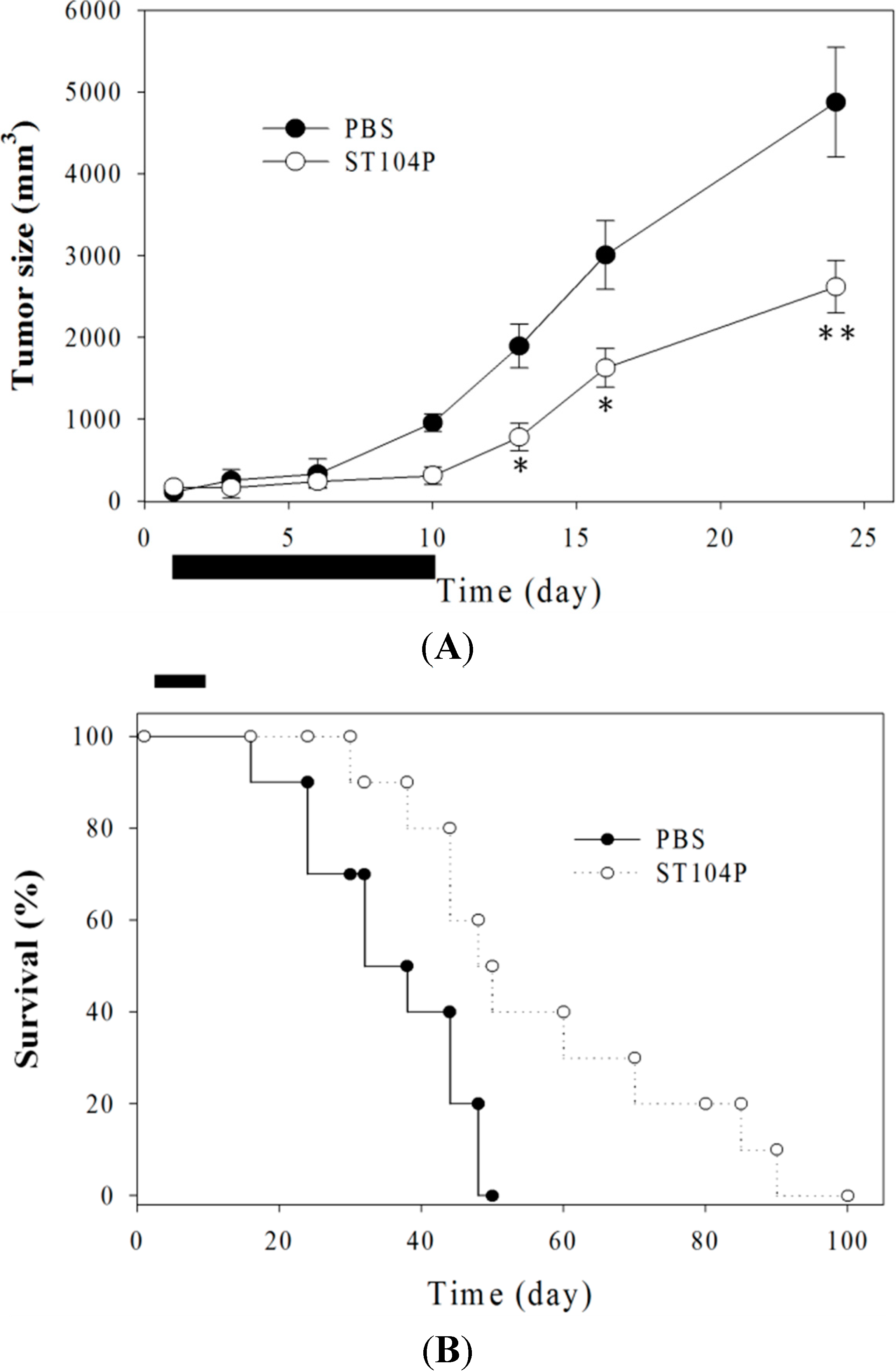
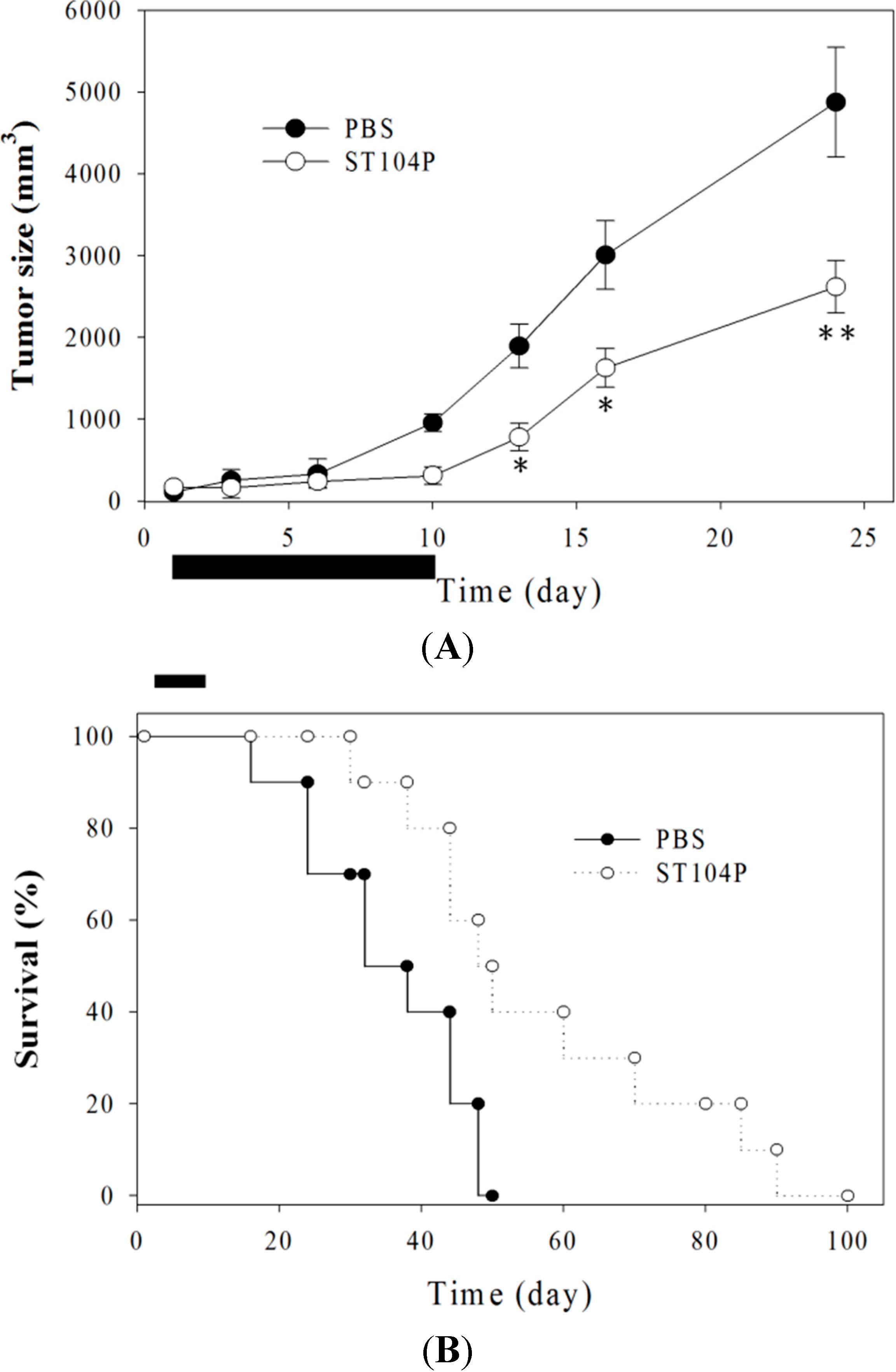
2.3. Injection of ST104P Suppressed Tumor Growth and Prolonged Survival in Mice
2.4. ST104P Attenuated Migration and Tube Formation of Cultured Endothelial Cells

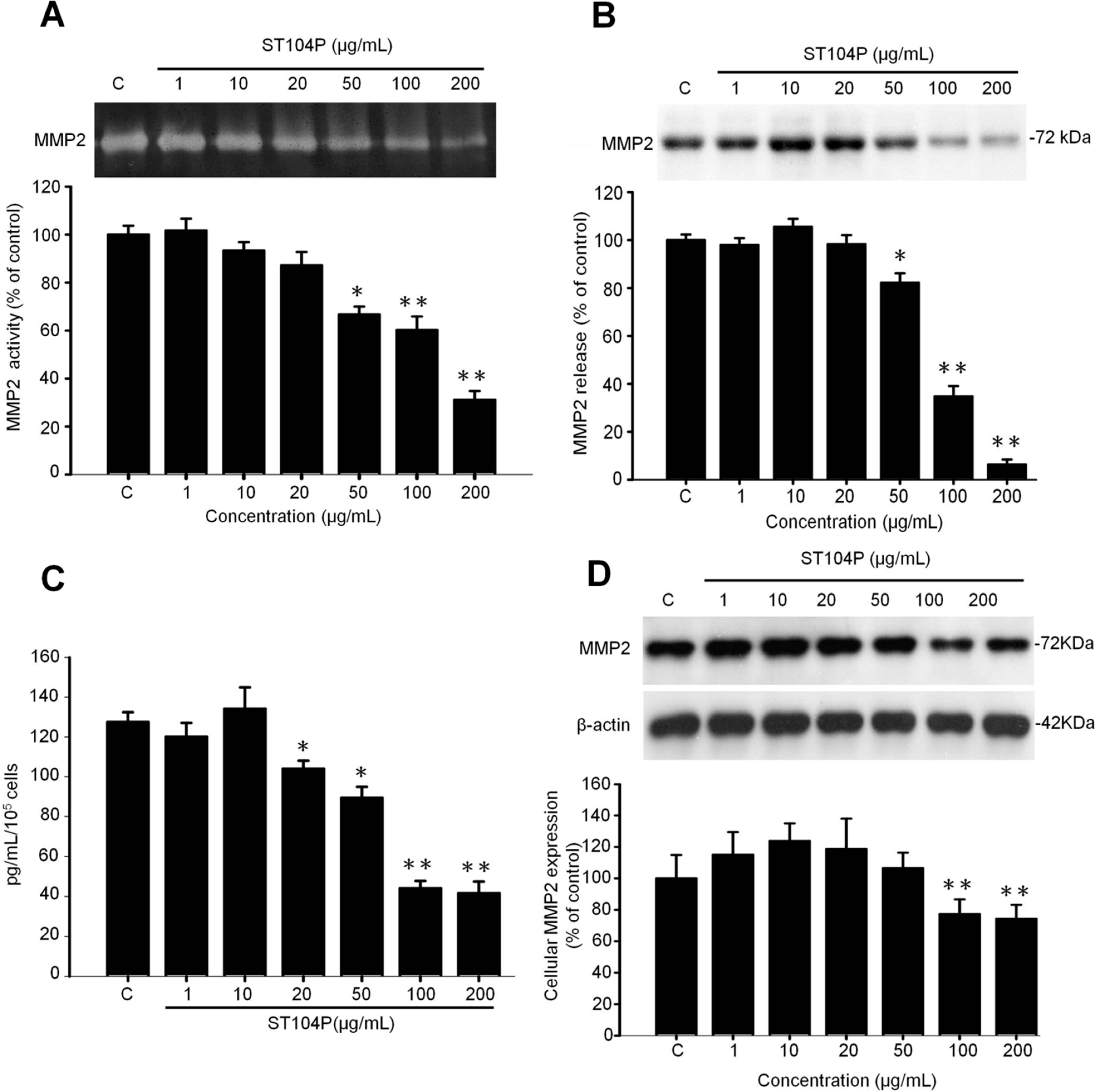
2.5. ST104P Inhibited Secretion and Expression of Matrix Metalloproteinase-2 (MMP-2) in Endothelial Cells
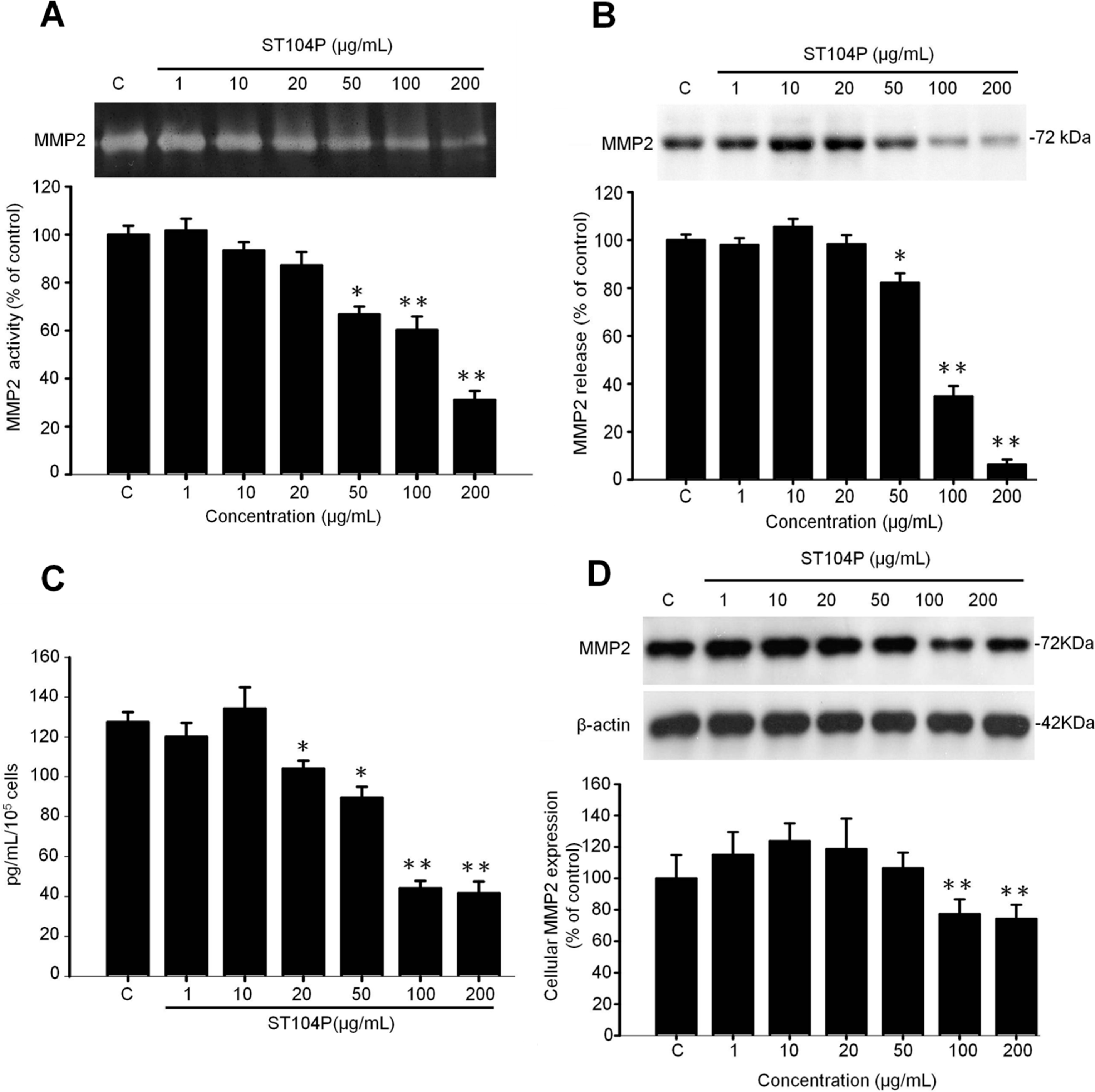
3. Discussion
4. Experimental Section
4.1. Chemical and Antibodies
4.2. Rat Aortic Ring Assay
4.3. Zebrafish Angiogenesis Model
4.4. Cells Cultures
4.5. Western Blot Analysis
4.6. Gelatin Zymography
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Cell Proliferation Assay
4.9. Migration Assay
4.10. Tube Formation Assay
4.11. Scratch Wound Healing Assay
4.12. Animal Studies
4.13. Treatment of Lewis Lung Carcinoma
4.14. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Folkman, J. Angiogenesis. Annu. Rev. Med. 2006, 57, 1–18. [Google Scholar]
- Griffioen, A.W.; Molema, G. Angiogenesis: Potentials for pharmacologic intervention in the treatment of cancer, cardiovascular diseases, and chronic inflammation. Pharmacol. Rev. 2000, 52, 237–268. [Google Scholar]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar]
- Eichhorn, M.E.; Kleespies, A.; Angele, M.K.; Jauch, K.W.; Bruns, C.J.; Bruns, C.J. Angiogenesis in cancer: Molecular mechanisms, clinical impact. Langenbecks Arch. Surg. 2007, 392, 371–379. [Google Scholar]
- Pandya NM, D.N.; Santani, D.D. Angiogenesis—A new target for future therapy. Vascul. Pharmacol. 2006, 44, 265–274. [Google Scholar]
- Rundhaug, J.E. Matrix metalloproteinases, angiogenesis, and cancer commentary re: A.C. Lockhart et al., Reduction of wound angiogenesis in patients treated with BMS-275291, a broad spectrum matrix metalloproteinase inhibitor. Clin. Cancer Res. 2003, 9, 551–554. [Google Scholar]
- Pratheeshkumar, P.; Kuttan, G. Andrographolide inhibits human umbilical vein endothelial cell invasion and migration by regulating MMP-2 and MMP-9 during angiogenesis. J. Environ. Pathol. Toxicol. Oncol. 2011, 30, 33–41. [Google Scholar]
- D’Amato, R.J.; Loughnan, M.S.; Flynn, E.; Folkman, J. Thalidomide is an inhibitor of angiogenesis. Proc. Natl. Acad. Sci. USA 1994, 91, 4082–4085. [Google Scholar]
- O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.R.; Folkman, J. Endostatin: An endogenous inhibitor of angiogenesis and tumor growth. Cell 1997, 88, 277–285. [Google Scholar]
- Folkman, J. Angiogenesis inhibitors: A new class of drugs. Cancer Biol. Ther. 2003, 2, S127–S133. [Google Scholar]
- Yao, V.J.; Ozawa, M.G.; Varner, A.S.; Kasman, I.M.; Chanthery, Y.H.; Pasqualini, R.; Arap, W.; McDonald, D.M. Antiangiogenic therapy decreases integrin expression in normalized tumor blood vessels. Cancer Res. 2006, 66, 2639–2649. [Google Scholar]
- Samant, R.S.; Shevde, L.A. Recent advances in anti-angiogenic therapy of cancer. Oncotarget 2011, 2, 1949–2553. [Google Scholar]
- Cook, K.M.; Figg, W.D. Angiogenesis inhibitors: Current strategies and future prospects. CA Cancer J. Clin. 2010, 60, 222–243. [Google Scholar]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar]
- Zheng, H.; Takahashi, H.; Murai, Y.; Cui, Z.; Nomoto, K.; Niwa, H.; Tsuneyama, K.; Takano, Y. Expressions of MMP-2, MMP-9 and VEGF are closely linked to growth, invasion, metastasis and angiogenesis of gastric carcinoma. Anticancer Res. 2006, 26, 3579–3583. [Google Scholar]
- Li, X.; Wu, J.F. Recent developments in patent anti-cancer agents targeting the matrix metalloproteinases (MMPs). Recent Pat. Anticancer Drug Discov. 2010, 5, 109–141. [Google Scholar]
- Poh, B.-L.; Lim, C.S.; Khoo, K.S. A water-soluble cyclic tetramer from reacting chromotropic acid with formaldehyde. Tetrahedron Lett. 1989, 30, 1005–1008. [Google Scholar]
- Poh, B.-L.; Chooi, S.L. Complexations of amines with water-soluble cyclotetrachromotropylene. Tetrahedron 1990, 46, 3651–3658. [Google Scholar]
- Poh, B.-L.; Seah, L.H.; Lim, C.S. Complexations of metal cations with cyclotetracmrohotropylene in water and methanol. Tetrahedron 1990, 46, 4379–4386. [Google Scholar]
- Ben Shoham, A.; Malkinson, G.; Krief, S.; Shwartz, Y.; Ely, Y.; Ferrara, N.; Yaniv, K.; Zelzer, E. S1P1 inhibits sprouting angiogenesis during vascular development. Development 2012, 139, 3859–3869. [Google Scholar]
- Lamalice, L.; Le Boeuf, F.; Huot, J. Endothelial cell migration during angiogenesis. Circ. Res. 2007, 100, 782–794. [Google Scholar]
- Patan, S. Vasculogenesis and angiogenesis as mechanisms of vascular network formation, growth and remodeling. J. Neurooncol. 2000, 50, 1–15. [Google Scholar]
- Burbridge, M.F.; Cog, F.; Galizzi, J.P.; Boutin, J.A.; West, D.C.; Tucker, G.C. The role of the matrix metalloproteinases during in vitro vessel formation. Angiogenesis 2002, 5, 215–226. [Google Scholar]
- Kim, T.S.; Kim, Y.B. Correlation between expression of matrix metalloproteinase-2 (MMP-2), and matrix metalloproteinase-9 (MMP-9) and angiogenesis in colorectal adenocarcinoma. J. Korean Med. Sci. 1999, 14, 263–270. [Google Scholar]
- Folkman, J. Angiogenesis: an organizing principle for drug discovery? Nat. Rev. Drug Dis. 2007, 6, 273–286. [Google Scholar]
- Moulton, K.S.; Heller, E.; Konerding, M.A.; Flynn, E.; Palinski, W.; Folkman, J. Angiogenesis inhibitors endostatin or TNP-470 reduce intimal neovascularization and plaque growth in apolipoprotein E-deficient mice. Circulation 1999, 99, 1726–1732. [Google Scholar]
- O’Reilly, M.S.; Holmgren, L.; Shing, Y.; Chen, C.; Rosenthal, R.A.; Moses, M.; Lane, W.S.; Cao, Y.; Sage, E.H.; Folkman, J. Angiostatin: A novel angiogenesis inhibitor that mediates the suppression of metastases by a Lewis lung carcinoma. Cell 1994, 79, 315–328. [Google Scholar]
- Shi, Z.G.; Li, J.P.; Shi, L.L.; Li, X. An updated patent therapeutic agents targeting MMPs. Recent Pat. Anticancer Drug Discov. 2012, 7, 74–101. [Google Scholar]
- Srivastava, K.; Kundumani-Sridharan, V.; Zhang, B.; Bajpai, A.K.; Rao, G.N. 15(S)-hydroxyeicosatetraenoic acid-induced angiogenesis requires STAT3-dependent expression of VEGF. Cancer Res. 2007, 67, 4328–4336. [Google Scholar]
- Bee, Y.S.; Sheu, S.J.; Ma, Y.-L.; Lin, H.-C.; Weng, W.-T.; Kuo, H.-M.; Hsu, H.-C.; Tang, C.-H.; Liou, J.-C.; Tai, M.-H.; et al. Topical application of recombinant calreticulin peptide, vasostatin 48, alleviates laser-induced choroidal neovascularization in rats. Mol. Vis. 2010, 16, 756–767. [Google Scholar]
- Roman, B.L.; Pham, V.N.; Lawson, N.D.; Kulik, M.; Childs, S.; Lekven, A.C.; Garrity, D.M.; Moon, R.T.; Fishman, M.C.; Lechleider, R.J.; et al. Disruption of acvrl1 increases endothelial cell number in zebrafish cranial vessels. Development 2002, 129, 3009–3019. [Google Scholar]
- Proulx, K.; Lu, A.; Sumanas, S. Cranial vasculature in zebrafish forms by angioblast cluster-derived angiogenesis. Dev. Biol. 2010, 348, 34–46. [Google Scholar]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar]
- Kuo, H.M.; Lin, C.-Y.; Lam, H.-C.; Lin, P.-R.; Chan, H.-H.; Tseng, J.-C.; Sun, C.-K.; Hsu, T.-F.; Wu, C.-C.; Yang, C.-Y.; et al. PTEN overexpression attenuates angiogenic processes of endothelial cells by blockade of endothelin-1/endothelin B receptor signaling. Atherosclerosis 2012, 221, 341–349. [Google Scholar]
- Tai, M.H.; Kuo, S.M.; Liang, H.T.; Chiou, K.R.; Lam, H.C.; Hsu, C.M.; Pownall, H.J.; Chen, H.-H.; Huang, M.T.; Yang, C.-Y. Modulation of angiogenic processes in cultured endothelial cells by low density lipoproteins subfractions from patients with familial hypercholesterolemia. Atherosclerosis 2006, 186, 448–457. [Google Scholar]
- Zhu, N.; Lalla, R.; Eves, P.; Brown, T.L.; King, A.; Kemp, E.H.; Haycock, J.W.; MacNeil, S. Melanoma cell migration is upregulated by tumour necrosis factor-α and suppressed by α-melanocyte-stimulating hormone. Br. J. Cancer 2004, 90, 1457–1463. [Google Scholar]
- Tsai, H.E.; Liu, L.F.; Dusting, G.J.; Weng, W.T.; Chen, S.C.; Kung, M.L.; Tee, R.; Liu, G.-S.; Tai, M.-H. Pro-opiomelanocortin gene delivery suppresses the growth of established Lewis lung carcinoma through a melanocortin-1 receptor-independent pathway. J. Gene Med. 2012, 14, 44–53. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ma, Y.-L.; Lin, S.-W.; Fang, H.-C.; Chou, K.-J.; Bee, Y.-S.; Chu, T.-H.; Chang, M.-C.; Weng, W.-T.; Wu, C.-Y.; Cho, C.-L.; et al. A Novel Poly-Naphthol Compound ST104P Suppresses Angiogenesis by Attenuating Matrix Metalloproteinase-2 Expression in Endothelial Cells. Int. J. Mol. Sci. 2014, 15, 16611-16627. https://doi.org/10.3390/ijms150916611
Ma Y-L, Lin S-W, Fang H-C, Chou K-J, Bee Y-S, Chu T-H, Chang M-C, Weng W-T, Wu C-Y, Cho C-L, et al. A Novel Poly-Naphthol Compound ST104P Suppresses Angiogenesis by Attenuating Matrix Metalloproteinase-2 Expression in Endothelial Cells. International Journal of Molecular Sciences. 2014; 15(9):16611-16627. https://doi.org/10.3390/ijms150916611
Chicago/Turabian StyleMa, Yi-Ling, Shih-Wei Lin, Hua-Chang Fang, Kang-Ju Chou, Youn-Shen Bee, Tian-Huei Chu, Ming-Chi Chang, Wen-Tsan Weng, Chang-Yi Wu, Chung-Lung Cho, and et al. 2014. "A Novel Poly-Naphthol Compound ST104P Suppresses Angiogenesis by Attenuating Matrix Metalloproteinase-2 Expression in Endothelial Cells" International Journal of Molecular Sciences 15, no. 9: 16611-16627. https://doi.org/10.3390/ijms150916611
APA StyleMa, Y.-L., Lin, S.-W., Fang, H.-C., Chou, K.-J., Bee, Y.-S., Chu, T.-H., Chang, M.-C., Weng, W.-T., Wu, C.-Y., Cho, C.-L., & Tai, M.-H. (2014). A Novel Poly-Naphthol Compound ST104P Suppresses Angiogenesis by Attenuating Matrix Metalloproteinase-2 Expression in Endothelial Cells. International Journal of Molecular Sciences, 15(9), 16611-16627. https://doi.org/10.3390/ijms150916611