Enamel Matrix Derivative Promote Primary Human Pulp Cell Differentiation and Mineralization

Abstract
:1. Introduction
2. Results
2.1. Affymetrix Microarray Expression Analysis
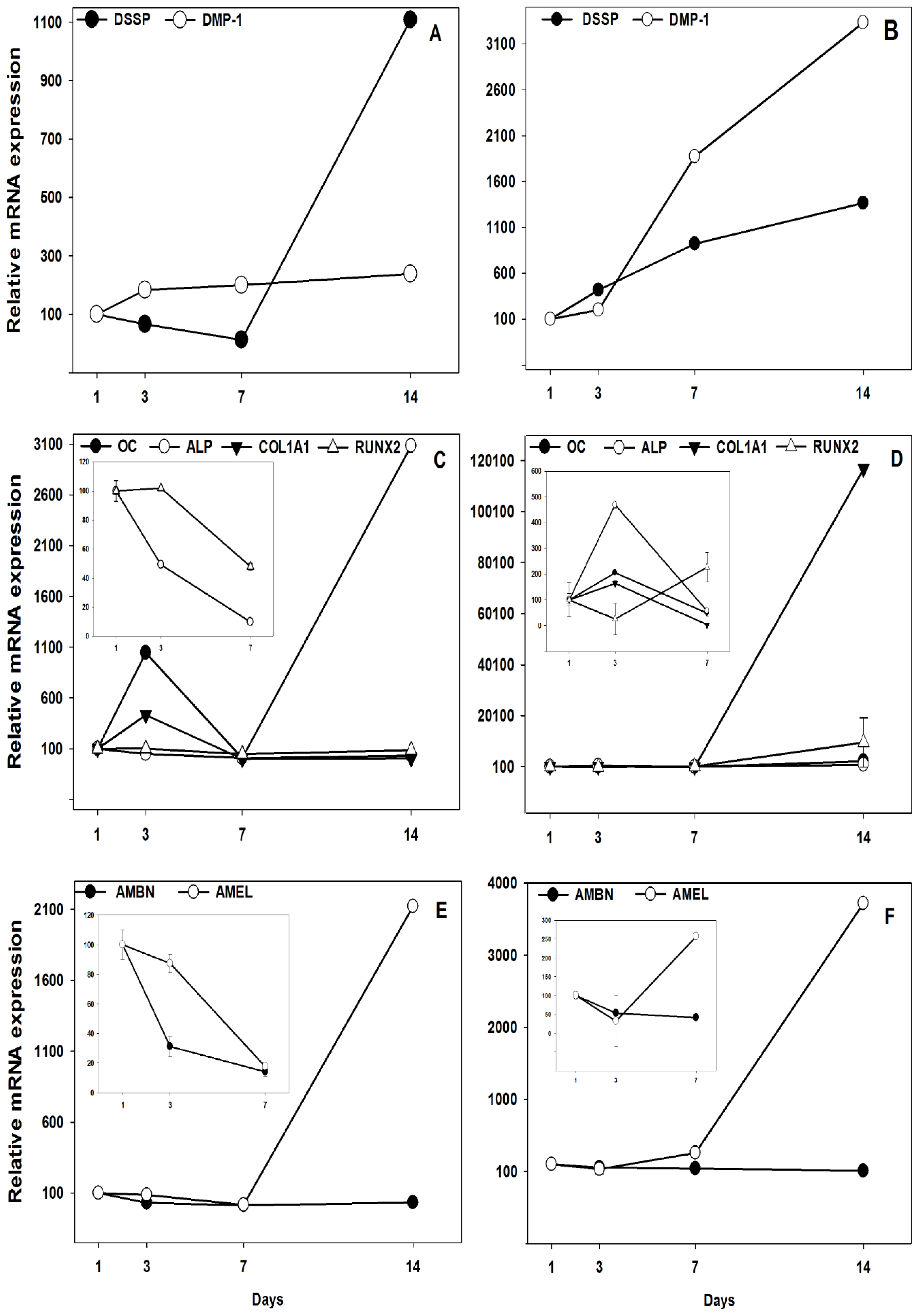
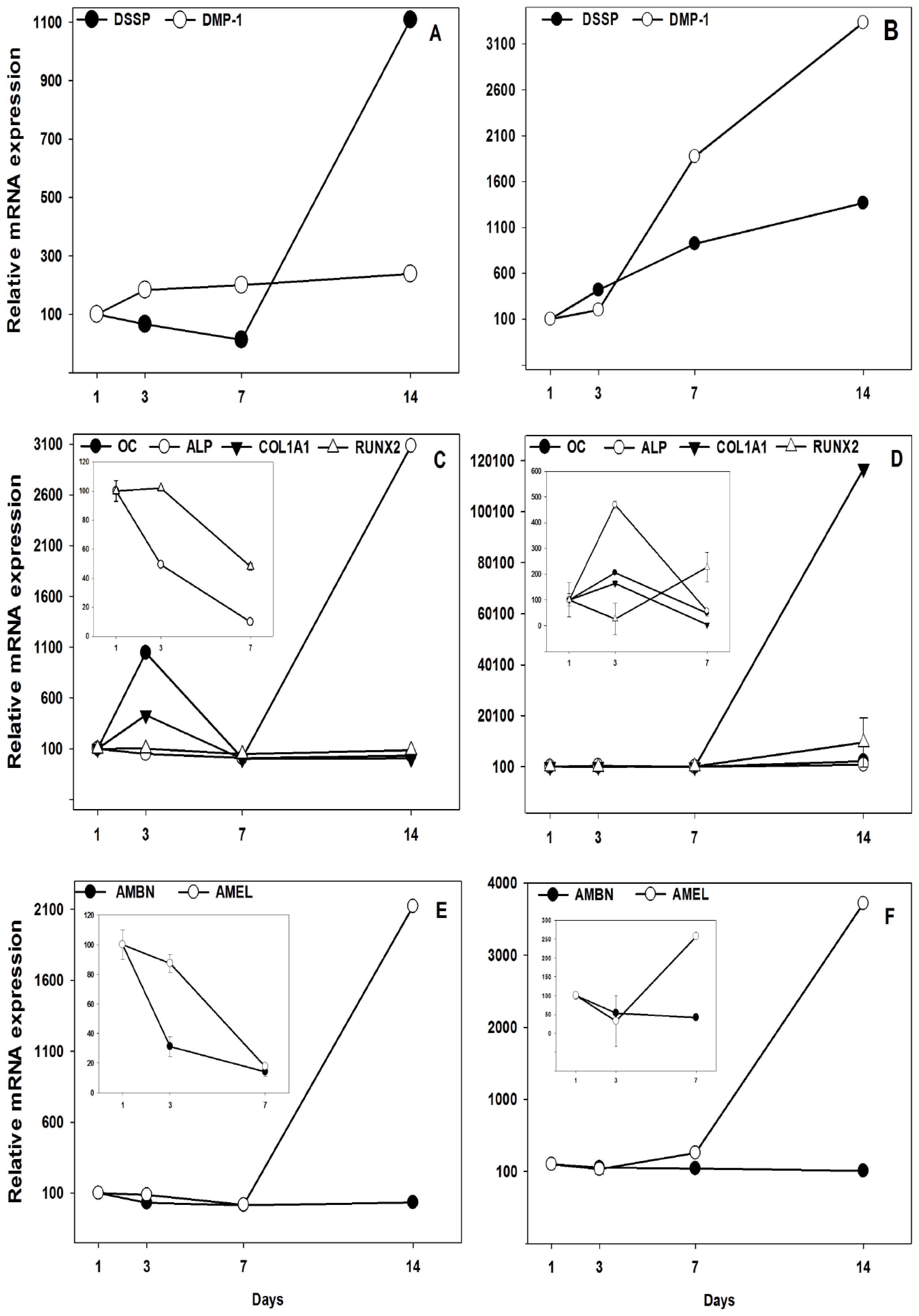
2.2. Gene Expression Analyzed by Real-Time PCR
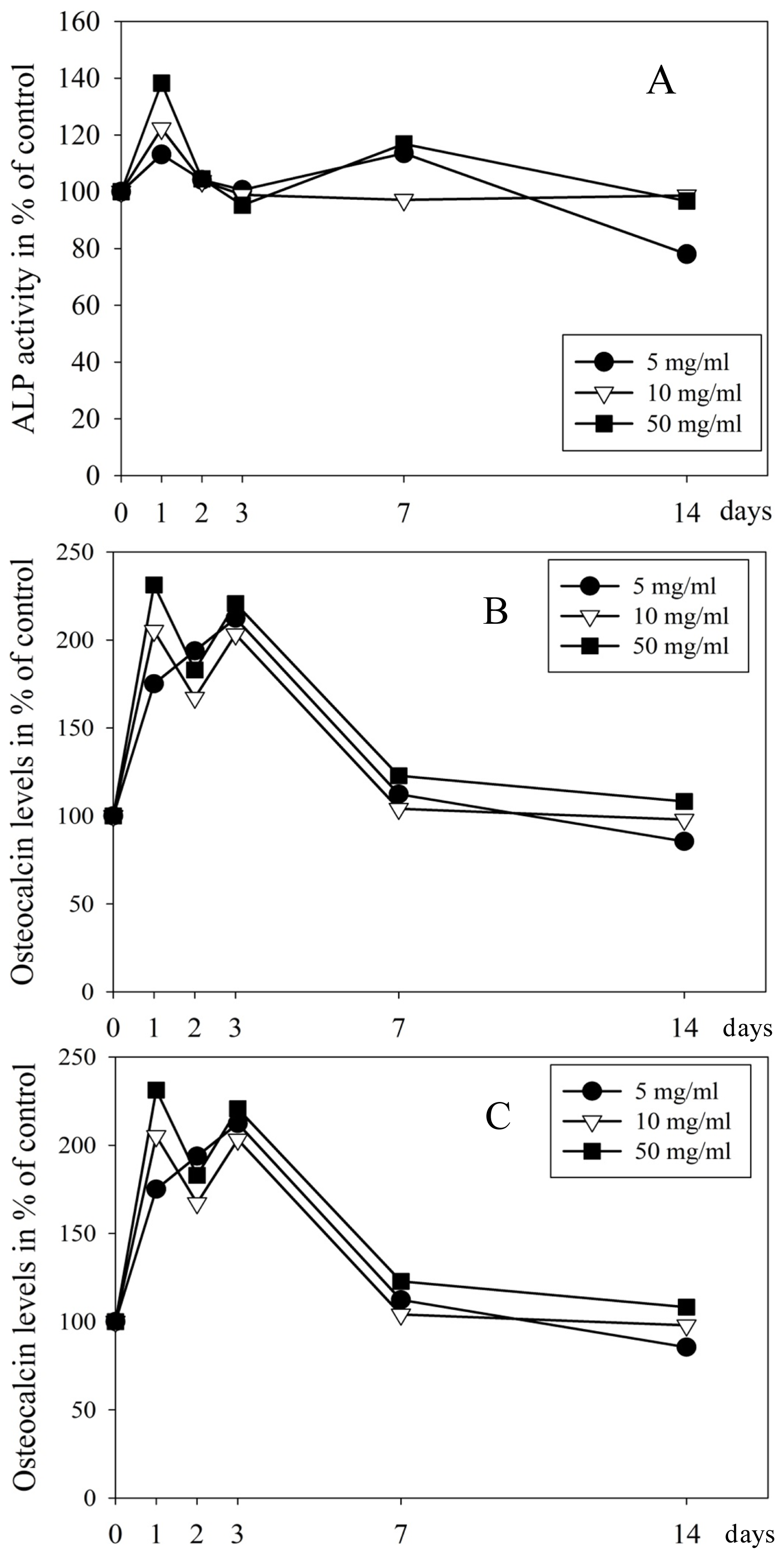
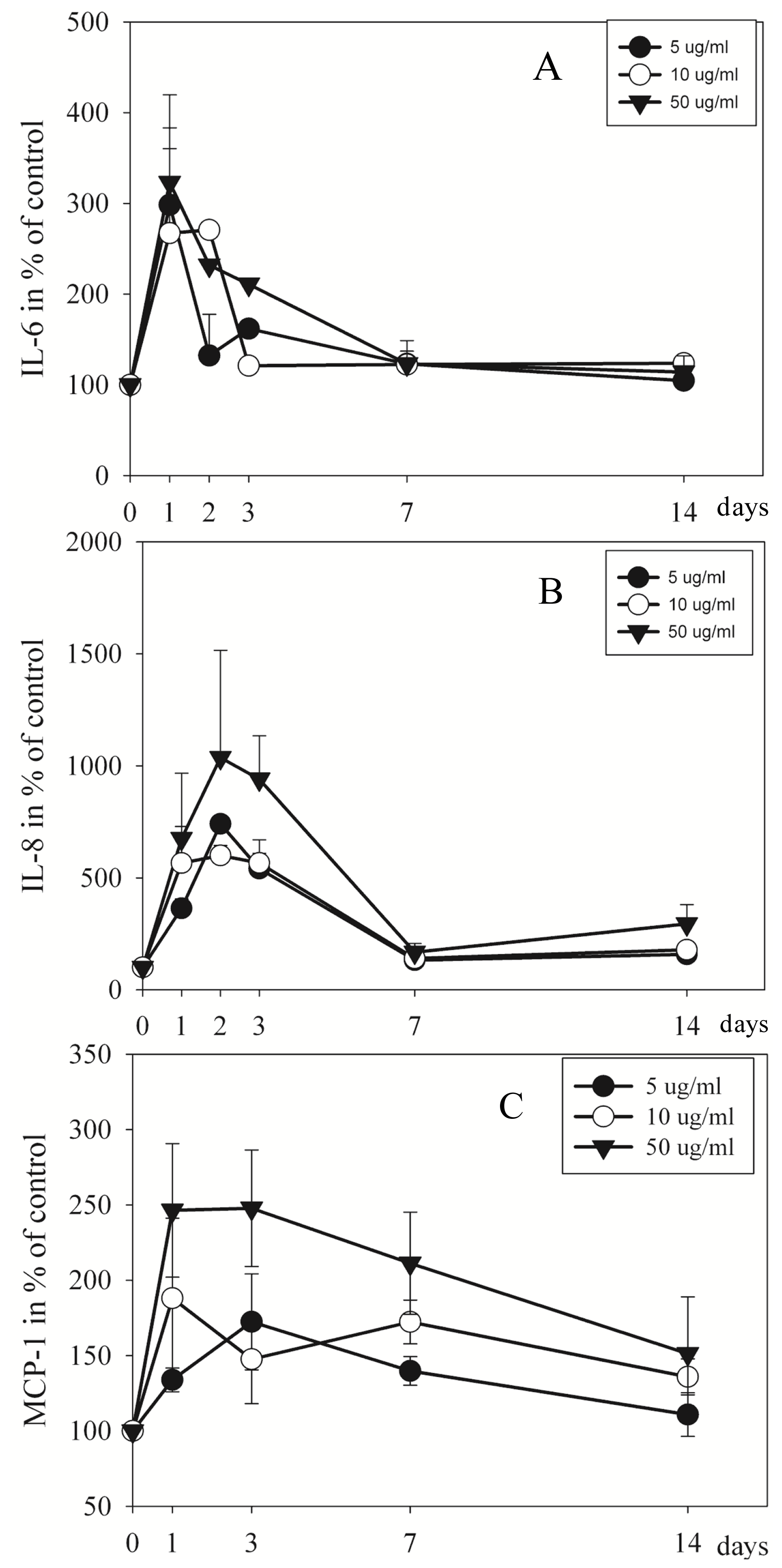
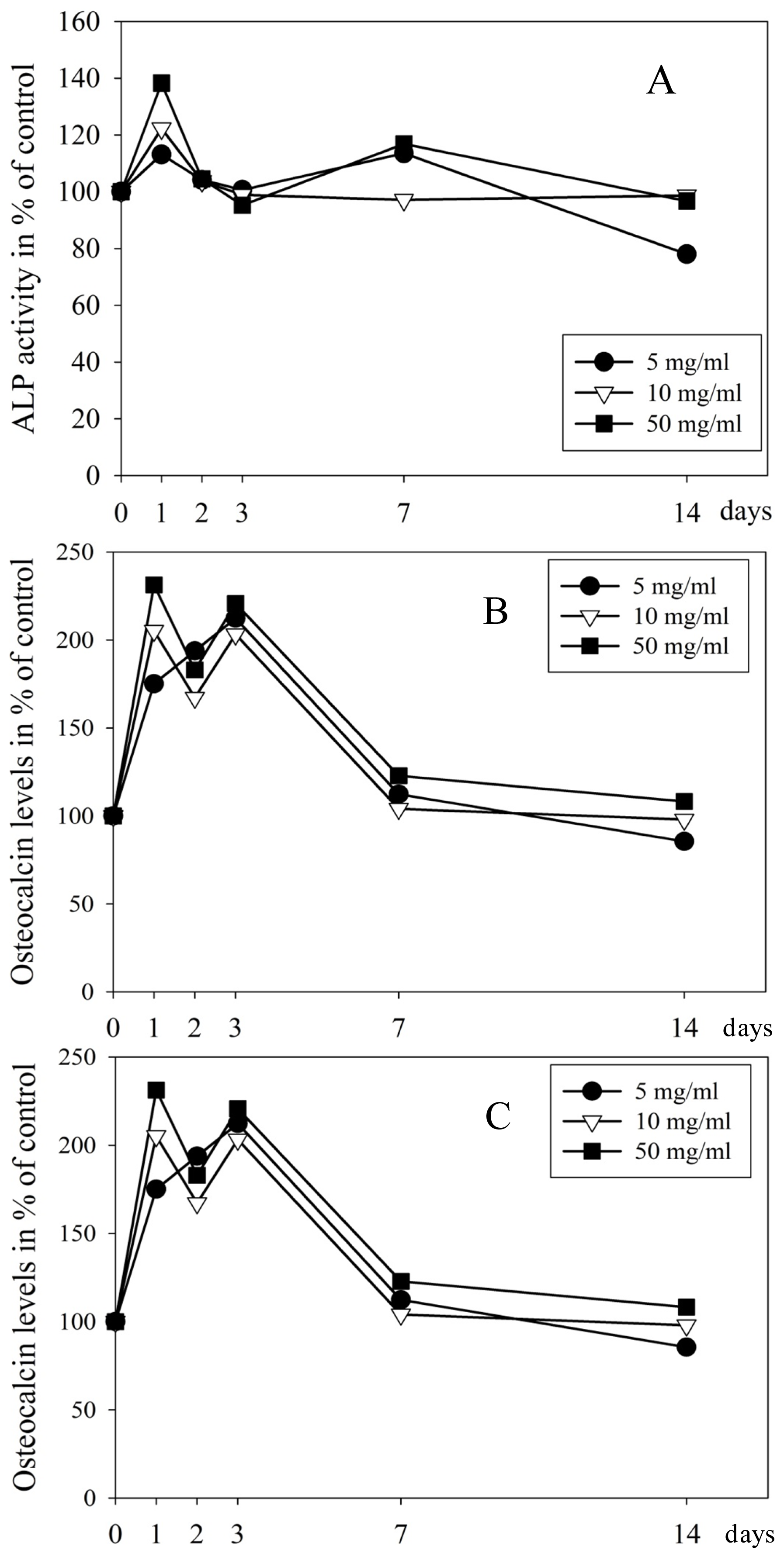
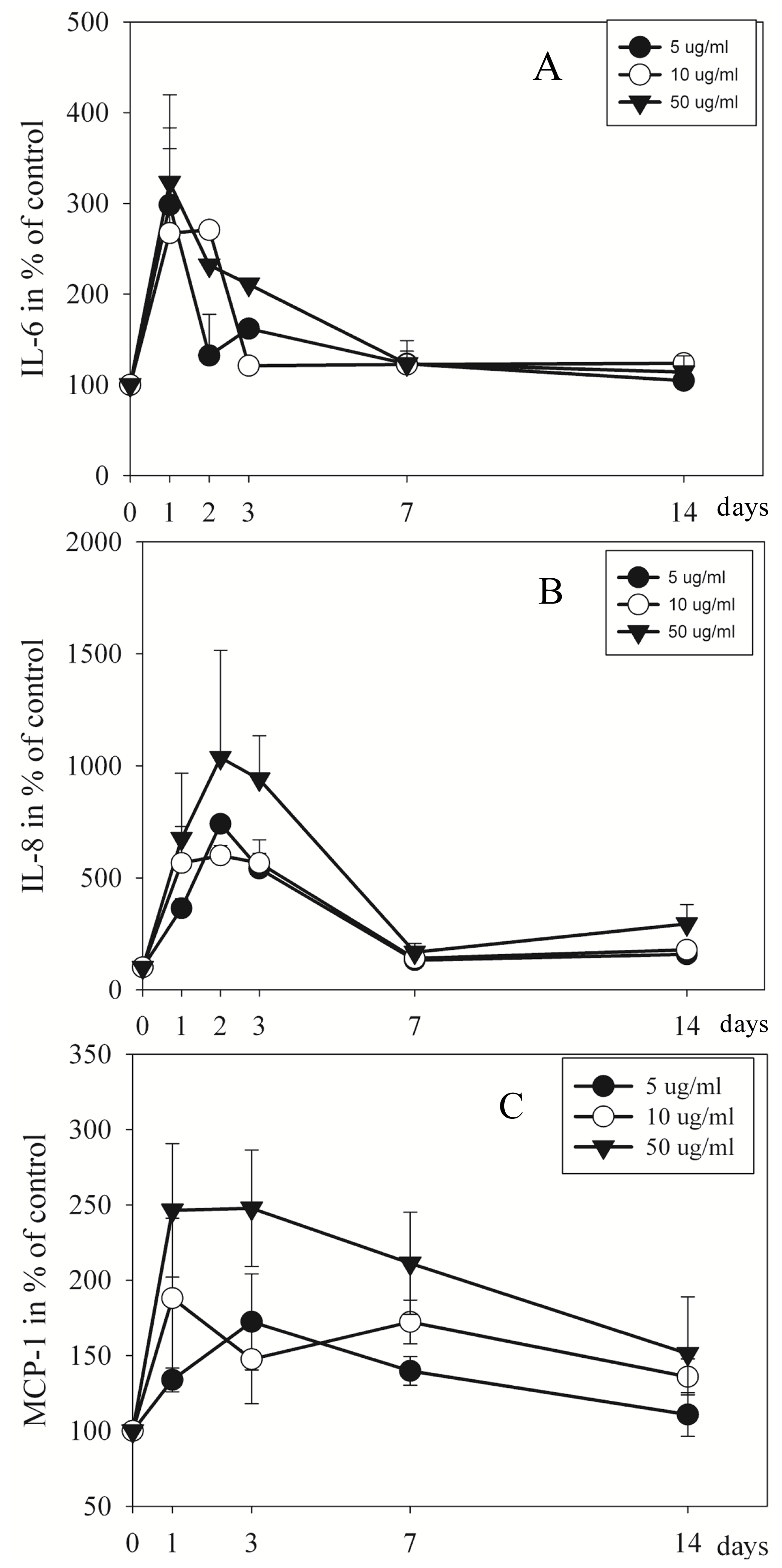
2.3. Proteins in Culture Medium Measured by Elisa and Luminex
2.4. No Cytotoxic Effects Was Discovered
3. Discussion
3.1. Affymetrix Analysis Showed Effect on Transcription Factors and DNA Repair
3.2. EMD Had Positive Effect on Genes Involved in both Neural and Mesenchymal Healing
3.3. EMD Induces Both Expression and Secretion of Osteogenic Proteins and Increased Mineralization Potential
3.4. EMD Has Immunoregulatory Effect on Pulp Cells
4. Experimental Section
4.1. Cell Culture and Treatment
4.2. Purification of RNA, Microarray Analysis by Affymetrix
4.3. Alkaline Phosphatase (ALP) Activity
4.4. mRNA Isolation Prior to Real Time RT-PCR Quantification
4.5. Real-Time PCR Quantification of Target Genes for Dental Biomineralization
4.6. Analysis of the Cell Culture Medium
4.7. Cytokine Levels in the Culture Medium
4.8. Lactate Dehydrogenase (LDH) Activity
4.9. Statistics
5. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsElisabeth Aurstad Riksen has performed ALP activity, total protein and LDH analysis, analyzed the Affymetrix, ELISA and Luminex results, and performed the statistics. She has written the article as first writer and has contributed to the collection of references. Maria Landin has performed RT-PCR and analyzed the results. Yukio Nakamura has done the cellular experiments. Sjur Reppe has performed affymetrix and analyzed the data. Janne Reseland have designed the study, planned the experiments, and contributed in discussing the results and editing the manuscript. Staale Petter Lyngstadaas has contributed in discussing the results and editing the manuscript.
References
- Stanley, H.R.; Pereira, J.C.; Spiegel, E.; Broom, C.; Schultz, M. The detection and prevalence of reactive and physiologic sclerotic dentin, reparative dentin and dead tracts beneath various types of dental lesions according to tooth surface and age. J. Oral Pathol 1983, 12, 257–289. [Google Scholar]
- Mohammadi, Z.; Dummer, P.M. Properties and applications of calcium hydroxide in endodontics and dental traumatology. Int. Endod. J 2011, 44, 697–730. [Google Scholar]
- Hammarstrom, L. Enamel matrix, cementum development and regeneration. J. Clin. Periodontol 1997, 24, 658–668. [Google Scholar]
- Heijl, L.; Heden, G.; Svardstrom, G.; Ostgren, A. Enamel matrix derivative (emdogain) in the treatment of intrabony periodontal defects. J. Clin. Periodontol 1997, 24, 705–714. [Google Scholar]
- Gestrelius, S.; Lyngstadaas, S.P.; Hammarstrom, L. Emdogain-periodontal regeneration based on biomimicry. Clin. Oral Investig 2000, 4, 120–125. [Google Scholar]
- Nakamura, Y.; Hammarstrom, L.; Matsumoto, K.; Lyngstadaas, S.P. The induction of reparative dentine by enamel proteins. Int. Endod. J 2002, 35, 407–417. [Google Scholar]
- Nakamura, Y.; Slaby, I.; Matsumoto, K.; Ritchie, H.H.; Lyngstadaas, S.P. Immunohistochemical characterization of rapid dentin formation induced by enamel matrix derivative. Calcif. Tissue Int 2004, 75, 243–252. [Google Scholar]
- Butler, W.T. Dentin matrix proteins and dentinogenesis. Connect. Tissue Res 1995, 33, 59–65. [Google Scholar]
- Takeda, T.; Tezuka, Y.; Horiuchi, M.; Hosono, K.; Iida, K.; Hatakeyama, D.; Miyaki, S.; Kunisada, T.; Shibata, T.; Tezuka, K. Characterization of dental pulp stem cells of human tooth germs. J. Dent. Res 2008, 87, 676–681. [Google Scholar]
- Huang, G.T.; Shagramanova, K.; Chan, S.W. Formation of odontoblast-like cells from cultured human dental pulp cells on dentin in vitro. J. Endod. 2006, 32, 1066–1073. [Google Scholar]
- Min, J.H.; Ko, S.Y.; Cho, Y.B.; Ryu, C.J.; Jang, Y.J. Dentinogenic potential of human adult dental pulp cells during the extended primary culture. Hum. Cell 2011, 24, 43–50. [Google Scholar]
- Kiatwateeratana, T.; Kintarak, S.; Piwat, S.; Chankanka, O.; Kamaolmatyakul, S.; Thearmontree, A. Partial pulpotomy on caries-free teeth using enamel matrix derivative or calcium hydroxide: A randomized controlled trial. Int. Endod. J 2009, 42, 584–592. [Google Scholar]
- Olsson, H.; Davies, J.R.; Holst, K.E.; Schroder, U.; Petersson, K. Dental pulp capping: Effect of emdogain gel on experimentally exposed human pulps. Int. Endod. J 2005, 38, 186–194. [Google Scholar]
- Ishizaki, N.T.; Matsumoto, K.; Kimura, Y.; Wang, X.; Yamashita, A. Histopathological study of dental pulp tissue capped with enamel matrix derivative. J. Endod 2003, 29, 176–179. [Google Scholar]
- Kawase, T.; Okuda, K.; Momose, M.; Kato, Y.; Yoshie, H.; Burns, D.M. Enamel matrix derivative (EMDOGAIN) rapidly stimulates phosphorylation of the MAP kinase family and nuclear accumulation of smad2 in both oral epithelial and fibroblastic human cells. J. Periodontal Res 2001, 36, 367–376. [Google Scholar]
- Iwata, T.; Morotome, Y.; Tanabe, T.; Fukae, M.; Ishikawa, I.; Oida, S. Noggin blocks osteoinductive activity of porcine enamel extracts. J. Dent. Res 2002, 81, 387–391. [Google Scholar]
- Kaida, H.; Hamachi, T.; Anan, H.; Maeda, K. Wound healing process of injured pulp tissues with emdogain gel. J. Endod 2008, 34, 26–30. [Google Scholar]
- Guven, E.P.; Yalvac, M.E.; Sahin, F.; Yazici, M.M.; Rizvanov, A.A.; Bayirli, G. Effect of dental materials calcium hydroxide-containing cement, mineral trioxide aggregate, and enamel matrix derivative on proliferation and differentiation of human tooth germ stem cells. J. Endod 2011, 37, 650–656. [Google Scholar]
- Nakamura, Y.; Slaby, I.; Spahr, A.; Pezeshki, G.; Matsumoto, K.; Lyngstadaas, S.P. Ameloblastin fusion protein enhances pulpal healing and dentin formation in porcine teeth. Calcif. Tissue Int 2006, 78, 278–284. [Google Scholar]
- Linde, A. Dentin mineralization and the role of odontoblasts in calcium transport. Connect. Tissue Res 1995, 33, 163–170. [Google Scholar]
- Prasad, M.; Butler, W.T.; Qin, C. Dentin sialophosphoprotein in biomineralization. Connect. Tissue Res 2010, 51, 404–417. [Google Scholar]
- Gericke, A.; Qin, C.; Sun, Y.; Redfern, R.; Redfern, D.; Fujimoto, Y.; Taleb, H.; Butler, W.T.; Boskey, A.L. Different forms of DMP1 play distinct roles in mineralization. J. Dent. Res 2010, 89, 355–359. [Google Scholar]
- Stockwin, L.H.; Vistica, D.T.; Kenney, S.; Schrump, D.S.; Butcher, D.O.; Raffeld, M.; Shoemaker, R.H. Gene expression profiling of alveolar soft-part sarcoma (ASPS). BMC Cancer 2009, 9. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, L.; Lai, H.; Parada, L.F.; Lei, L. Transcription factor Sox11 is essential for both embryonic and adult neurogenesis. Dev. Dyn 2013. [Google Scholar] [CrossRef]
- Tian, C.; Gong, Y.; Yang, Y.; Shen, W.; Wang, K.; Liu, J.; Xu, B.; Zhao, J.; Zhao, C. Foxg1 has an essential role in postnatal development of the dentate gyrus. J. Neurosci 2012, 32, 2931–2949. [Google Scholar]
- Reseland, J.E.; Reppe, S.; Larsen, A.M.; Berner, H.S.; Reinholt, F.P.; Gautvik, K.M.; Slaby, I.; Lyngstadaas, S.P. The effect of enamel matrix derivative on gene expression in osteoblasts. Eur. J. Oral Sci 2006, 114, 381–202. [Google Scholar]
- Nishikawa, K.; Nakashima, T.; Takeda, S.; Isogai, M.; Hamada, M.; Kimura, A.; Kodama, T.; Yamaguchi, A.; Owen, M.J.; Takahashi, S.; et al. Maf promotes osteoblast differentiation in mice by mediating the age-related switch in mesenchymal cell differentiation. J. Clin. Investig 2010, 120, 3455–3465. [Google Scholar]
- Meier, D.; Schindler, D. Fanconi anemia core complex gene promoters harbor conserved transcription regulatory elements. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Morshedi, A.; Ren, Z.; Li, J.; Droge, P. Probing into the biological processes influenced by ESC factor and oncoprotein HMGA2 using iPSCS. Stem Cell Rev 2013, 9, 514–522. [Google Scholar]
- Kuipers, A.; Zhang, Y.; Cauley, J.A.; Nestlerode, C.S.; Chu, Y.; Bunker, C.H.; Patrick, A.L.; Wheeler, V.W.; Hoffman, A.R.; Orwoll, E.S.; et al. Association of a high mobility group gene (HMGA2) variant with bone mineral density. Bone 2009, 45, 295–300. [Google Scholar]
- Barkana, I.; Alexopoulou, E.; Ziv, S.; Jacob-Hirsch, J.; Amariglio, N.; Pitaru, S.; Vardimon, A.D.; Nemcovsky, C.E. Gene profile in periodontal ligament cells and clones with enamel matrix proteins derivative. J. Clin. Periodontol 2007, 34, 599–609. [Google Scholar]
- Zhang, H.; Somerman, M.J.; Berg, J.; Cunningham, M.L.; Williams, B. Dental anomalies in a child with craniometaphysial dysplasia. Pediatr. Dent 2007, 29, 415–419. [Google Scholar]
- Wu, X.; Li, J.; Chen, C.; Yan, Y.; Jiang, S.; Shao, B.; Xu, J.; Kang, L.; Huang, Y.; Zhu, L.; et al. Involvement of CLEC16A in activation of astrocytes after LPS treated. Neurochem. Res 2012, 37, 5–14. [Google Scholar]
- Djurovic, S.; Gustafsson, O.; Mattingsdal, M.; Athanasiu, L.; Bjella, T.; Tesli, M.; Agartz, I.; Lorentzen, S.; Melle, I.; Morken, G.; et al. A genome-wide association study of bipolar disorder in norwegian individuals, followed by replication in icelandic sample. J. Affect Disord 2010, 126, 312–316. [Google Scholar]
- Kriebel, M.; Wuchter, J.; Trinks, S.; Volkmer, H. Neurofascin: A switch between neuronal plasticity and stability. Int. J. Biochem. Cell. Biol 2012, 44, 694–697. [Google Scholar]
- Miron, R.J.; Bosshardt, D.D.; Zhang, Y.; Buser, D.; Sculean, A. Gene array of primary human osteoblasts exposed to enamel matrix derivative in combination with a natural bone mineral. Clin. Oral Investig 2013, 17, 405–410. [Google Scholar]
- Coppe, C.; Zhang, Y.; Den Besten, P.K. Characterization of primary dental pulp cells in vitro. Pediatr. Dent. 2009, 31, 467–471. [Google Scholar]
- Miron, R.J.; Oates, C.J.; Molenberg, A.; Dard, M.; Hamilton, D.W. The effect of enamel matrix proteins on the spreading, proliferation and differentiation of osteoblasts cultured on titanium surfaces. Biomaterials 2010, 31, 449–460. [Google Scholar]
- Bronckers, A.L.; Farach-Carson, M.C.; van Waveren, E.; Butler, W.T. Immunolocalization of osteopontin, osteocalcin, and dentin sialoprotein during dental root formation and early cementogenesis in the rat. J. Bone Miner. Res 1994, 9, 833–841. [Google Scholar]
- Narayanan, K.; Gajjeraman, S.; Ramachandran, A.; Hao, J.; George, A. Dentin matrix protein 1 regulates dentin sialophosphoprotein gene transcription during early odontoblast differentiation. J. Biol. Chem 2006, 281, 19064–19071. [Google Scholar]
- Lyngstadaas, S.P.; Wohlfahrt, J.C.; Brookes, S.J.; Paine, M.L.; Snead, M.L.; Reseland, J.E. Enamel matrix proteins; Old molecules for new applications. Orthod. Craniofac. Res 2009, 12, 243–253. [Google Scholar]
- Yan, X.Z.; Rathe, F.; Gilissen, C.; van der Zande, M.; Veltman, J.; Junker, R.; Yang, F.; Jansen, J.A.; Walboomers, X.F. The effect of enamel matrix derivative (emdogain®) on gene expression profiles of human primary alveolar bone cells. J. Tissue Eng. Regen. Med 2012. [Google Scholar] [CrossRef]
- Chai, Z.; Alheim, K.; Lundkvist, J.; Gatti, S.; Bartfai, T. Subchronic glucocorticoid pretreatment reversibly attenuates IL-beta induced fever in rats; IL-6 mRNA is elevated while IL-1 alpha and IL-1 beta mRNAs are suppressed, in the CNS. Cytokine 1996, 8, 227–237. [Google Scholar]
- Sundgren-Andersson, A.K.; Ostlund, P.; Bartfai, T. IL-6 is essential in TNF-alpha-induced fever. Am. J. Physiol 1998, 275, 2028–2034. [Google Scholar]
- Parkar, M.H.; Tonetti, M. Gene expression profiles of periodontal ligament cells treated with enamel matrix proteins in vitro: Analysis using cdna arrays. J. Periodontol 2004, 75, 1539–1546. [Google Scholar]
- Abd-Elmeguid, A.; Yu, D.C.; Kline, L.W.; Moqbel, R.; Vliagoftis, H. Dentin matrix protein 1 activates dental pulp fibroblasts. J. Endod 2012, 38, 75–80. [Google Scholar]
- Dokic, J.; Tomic, S.; Cerovic, S.; Todorovic, V.; Rudolf, R.; Colic, M. Characterization and immunosuppressive properties of mesenchymal stem cells from periapical lesions. J. Clin. Periodontol 2012, 39, 807–816. [Google Scholar]
- Sims, N.A.; Jenkins, B.J.; Nakamura, A.; Quinn, J.M.; Li, R.; Gillespie, M.T.; Ernst, M.; Robb, L.; Martin, T.J. Interleukin-11 receptor signaling is required for normal bone remodeling. J. Bone Miner. Res 2005, 20, 1093–1102. [Google Scholar]
- Schinkel, C.; Wick, M.; Muhr, G.; Koller, M. Analysis of systemic interleukin-11 after major trauma. Shock 2005, 23, 30–34. [Google Scholar]
- Nawa, K.; Ikeno, H.; Matsuhashi, N.; Ogasawara, T.; Otsuka, E. Discovering small molecules that inhibit adipogenesis and promote osteoblastogenesis: Unique screening and Oncostatin M-like activity. Differentiation 2013, 86, 65–74. [Google Scholar]
- Gu, L.; Tseng, S.C.; Rollins, B.J. Monocyte chemoattractant protein-1. Chem. Immunol 1999, 72, 7–29. [Google Scholar]
- Alliot-Licht, B.; Bluteau, G.; Magne, D.; Lopez-Cazaux, S.; Lieubeau, B.; Daculsi, G.; Guicheux, J. Dexamethasone stimulates differentiation of odontoblast-like cells in human dental pulp cultures. Cell Tissue Res 2005, 321, 391–400. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Biological Response | Probe Set Identification (Affymetrix) | Signal Log Ratio | Gene Symbol | Gene Title |
|---|---|---|---|---|
| Apoptosis | 204614_at | 1, 8 | SERPINB2 | serine (or cysteine) proteinase inhibitor, clade B (ovalbumin) |
| 204467_sat | −1 | SNCA | synuclein, alpha (non A4 component of amyloid precursor) | |
| 205681_at | 1, 5 | BCL2A1 | BCL2-related protein A1 | |
| 202095_s_at | 1, 4 | BIRC5 | baculoviral IAP repeat-containing 5 (survivin) | |
| 204237_at | −1 | GULP1 | GULP, engulfment adaptor PTB domain containing 1 | |
| Cell adhesion | 205679_x_at | −1, 2 | AGC1 | aggrecan 1 (chondroitin sulfate proteoglycan 1) |
| 1553418_aat | 3, 8 | CNTNAP5 | contactin associated protein-like 5 | |
| 222853_at | −1, 1 | FLRT3 | fibronectin leucine rich transmembrane protein 3 | |
| 243645_at | 3, 5 | NFASC | neurofascin | |
| 205908_s_at | −1, 5 | OMD | osteomodulin | |
| 204649_at | 1, 2 | TROAP | trophinin associated protein (tastin) | |
| 238871_at | −1, 7 | MLLT4 | Myeloid/lymphoid or mixed-lineage leukemia | |
| 242472_x_at | −3, 7 | FNBP4 | Formin binding protein 4 | |
| 231726_at | −1 | PCDHB14 | protocadherin beta 14 | |
| 214111_at | 1, 1 | OPCML | opioid binding protein/cell adhesion molecule-like | |
| Cell cycle | 227552_at | 1, 8 | sep.01 | septin 1 |
| 204456_s_at | −1 | GAS1 | growth arrest-specific 1 | |
| 204991_s_at | 1 | NF2 | neurofibromin 2 (bilateral acoustic neuroma) | |
| 211094_s_at | −1, 8 | NF1 | neurofibromin 1 (neurofibromatosis) | |
| 225123_at | −1, 1 | SESN3 | Sestrin 3 | |
| 202859_x_at | 1, 9 | IL8 | interleukin 8 | |
| Growth factor | 222722_at | −1, 4 | OGN | osteoglycin (osteoinductive factor, mimecan) |
| 219304_s_at | −1, 5 | PDGFD | platelet derived growth factor D | |
| 205782_at | −1, 1 | FGF7 | fibroblast growth factor 7 (keratinocyte growth factor) | |
| 211518_s_at | −1, 3 | BMP4 | bone morphogenetic protein 4 | |
| Transcription | 1569098_sat | −1, 2 | TP53BP1 | tumor protein p53 binding protein, 1 |
| 203359_s_at | 1, 1 | MYCBP | c-myc binding protein | |
| 201417_at | −1 | SOX4 | SRY (sex determining region Y)-box 4 | |
| 227162_at | −1, 1 | ZBTB26 | zinc finger and BTB domain containing 26 | |
| 1568873_at | 1, 2 | ZNF519 | zinc finger protein 519 | |
| 221530_s_at | −1, 2 | BHLHB3 | basic helix-loop-helix domain containing, class B, 3 | |
| 204915_s_at | 1 | SOX11 | SRY (sex determining region Y)-box 11 | |
| 206018_at | 1, 4 | FOXG1B | forkhead box G1B | |
| microtubule cytoskeleton | 202890_at | −1 | MAP7 | microtubule-associated protein 7 |
| 206364_at | 1 | KIF14 | kinesin family member 14 | |
| 221258_sat | 1, 4 | KIF18A | kinesin family member 18A | |
| 219570_at | −1 | C20orf23 | chromosome 20 open reading frame 23 | |
| 219306_at | 1, 1 | KIF15 | kinesin family member 15 | |
| 209408_at | 1, 2 | KIF2C | kinesin family member 2C | |
| 218755_at | 1, 1 | KIF20A | kinesin family member 20A | |
| 210052_sat | 1, 2 | TPX2 | TPX2, microtubule-associated protein homolog (Xenopus laevis) | |
| 219588_sat | 1, 1 | MTB | more than blood homolog | |
| 204162_at | 1, 2 | KNTC2 | kinetochore associated 2 | |
| 209680_sat | 3, 1 | KIFC1 | kinesin family member C1 | |
| 38158_at | 1, 4 | ESPL1 | extra spindle poles like 1 (S. cerevisiae) | |
| 218009_sat | 1, 3 | PRC1 | protein regulator of cytokinesis 1 | |
| 2047at | 1, 4 | KIF23 | kinesin family member 23 | |
| 225540_at | −1, 8 | MAP2 | Microtubule-associated protein 2 | |
| Gene | Sequence of Left Primer | Sequence of Right Primer |
|---|---|---|
| AMELX | 5′-CTCATCACCACATCCCAGTG-3′ | 5′-TGTTGGATTGGAGTCATGGA-3′ |
| AMBN | 5′-AGAGCACAGTGCATGTCAGC-3′ | 5′-AAGAACGGCACTGCAAAACT-3′ |
| DSPP | 5′-GGGAAAGTGGTGGTGGTGCT-3′ | 5′-CACCAGGGCATGGCTGTAAG-3′ |
| DMP1 | 5′-AAGCAGACAGCGAATCCAGT-3′ | 5′-CTGCTGAGCTGCTGTGAGAC-3′ |
| OC | 5′-GCAAGTAGCGCCAATCTAGG-3′ | 5′-GCTTCACCCTCGAAATGGTA-3′ |
| ALP | 5′-GACAAGAAGCCCTTCACTGC-3′ | 5′-AGACTGCGCCTGGTAGTTGT-3′ |
| COL1A1 | 5′-CATCTCCCCTTCGTTTTTGA-3′ | 5′-CCAAATCCGATGTTTCTGCT-3′ |
| RUNX2 | 5′-TTACTTACACCCCGCCAGTC-3′ | 5′-CACTCTGGCTTTGGGAAGAG-3′ |
| GAPDH | 5′-CTCTGCTCCTCCTGTTCGAC-3′ | 5′-ACGACCAAATCCGTTGACTC-3′ |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Riksen, E.A.; Landin, M.A.; Reppe, S.; Nakamura, Y.; Lyngstadaas, S.P.; Reseland, J.E. Enamel Matrix Derivative Promote Primary Human Pulp Cell Differentiation and Mineralization. Int. J. Mol. Sci. 2014, 15, 7731-7749. https://doi.org/10.3390/ijms15057731
Riksen EA, Landin MA, Reppe S, Nakamura Y, Lyngstadaas SP, Reseland JE. Enamel Matrix Derivative Promote Primary Human Pulp Cell Differentiation and Mineralization. International Journal of Molecular Sciences. 2014; 15(5):7731-7749. https://doi.org/10.3390/ijms15057731
Chicago/Turabian StyleRiksen, Elisabeth Aurstad, Maria A. Landin, Sjur Reppe, Yukio Nakamura, Ståle Petter Lyngstadaas, and Janne E. Reseland. 2014. "Enamel Matrix Derivative Promote Primary Human Pulp Cell Differentiation and Mineralization" International Journal of Molecular Sciences 15, no. 5: 7731-7749. https://doi.org/10.3390/ijms15057731
APA StyleRiksen, E. A., Landin, M. A., Reppe, S., Nakamura, Y., Lyngstadaas, S. P., & Reseland, J. E. (2014). Enamel Matrix Derivative Promote Primary Human Pulp Cell Differentiation and Mineralization. International Journal of Molecular Sciences, 15(5), 7731-7749. https://doi.org/10.3390/ijms15057731