ESCRT Function in Cytokinesis: Location, Dynamics and Regulation by Mitotic Kinases

Abstract
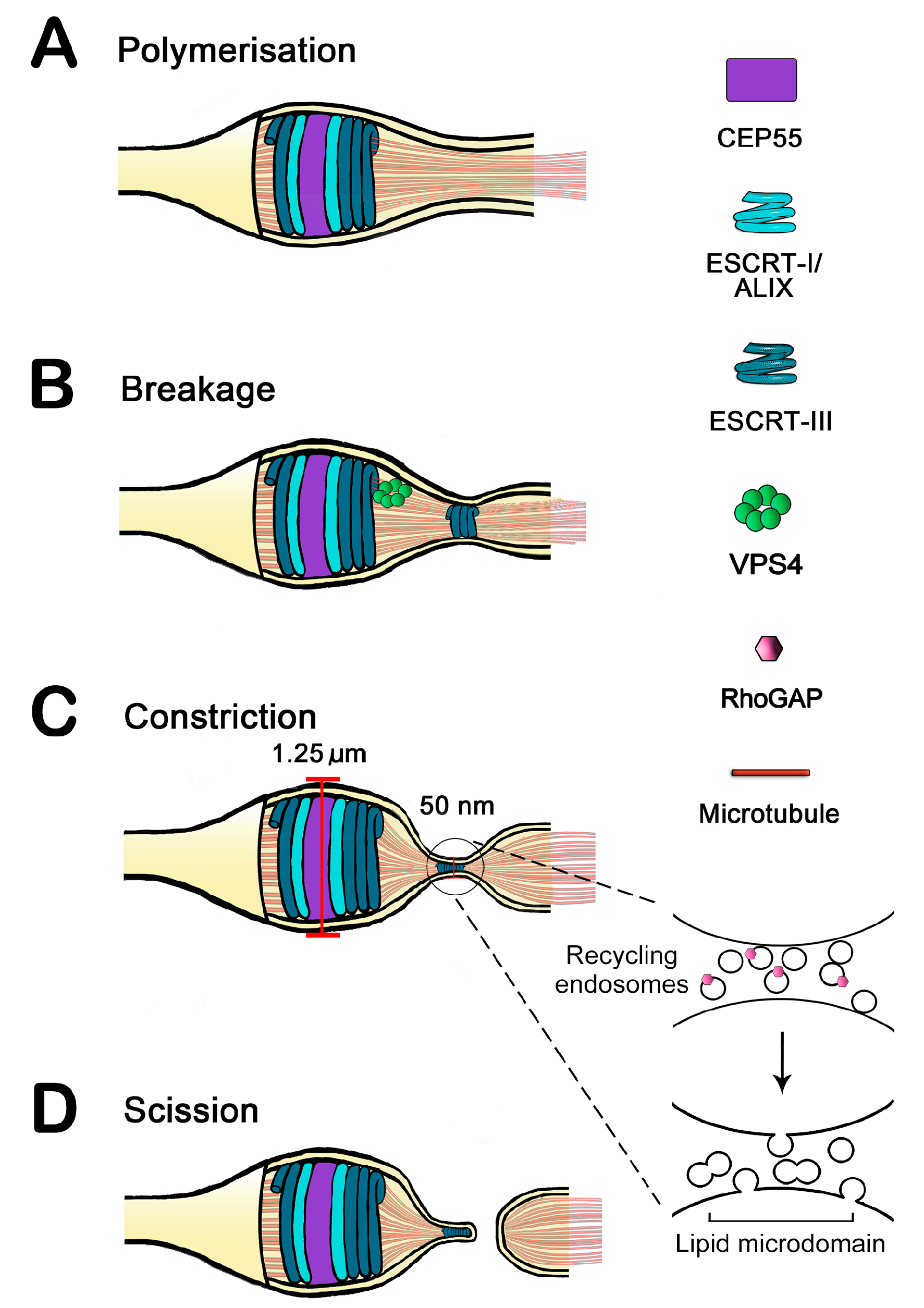
:1. Resolving the Bridge: Cleavage of the Intercellular Bridge Is a Multi-Step, Tightly Regulated Process
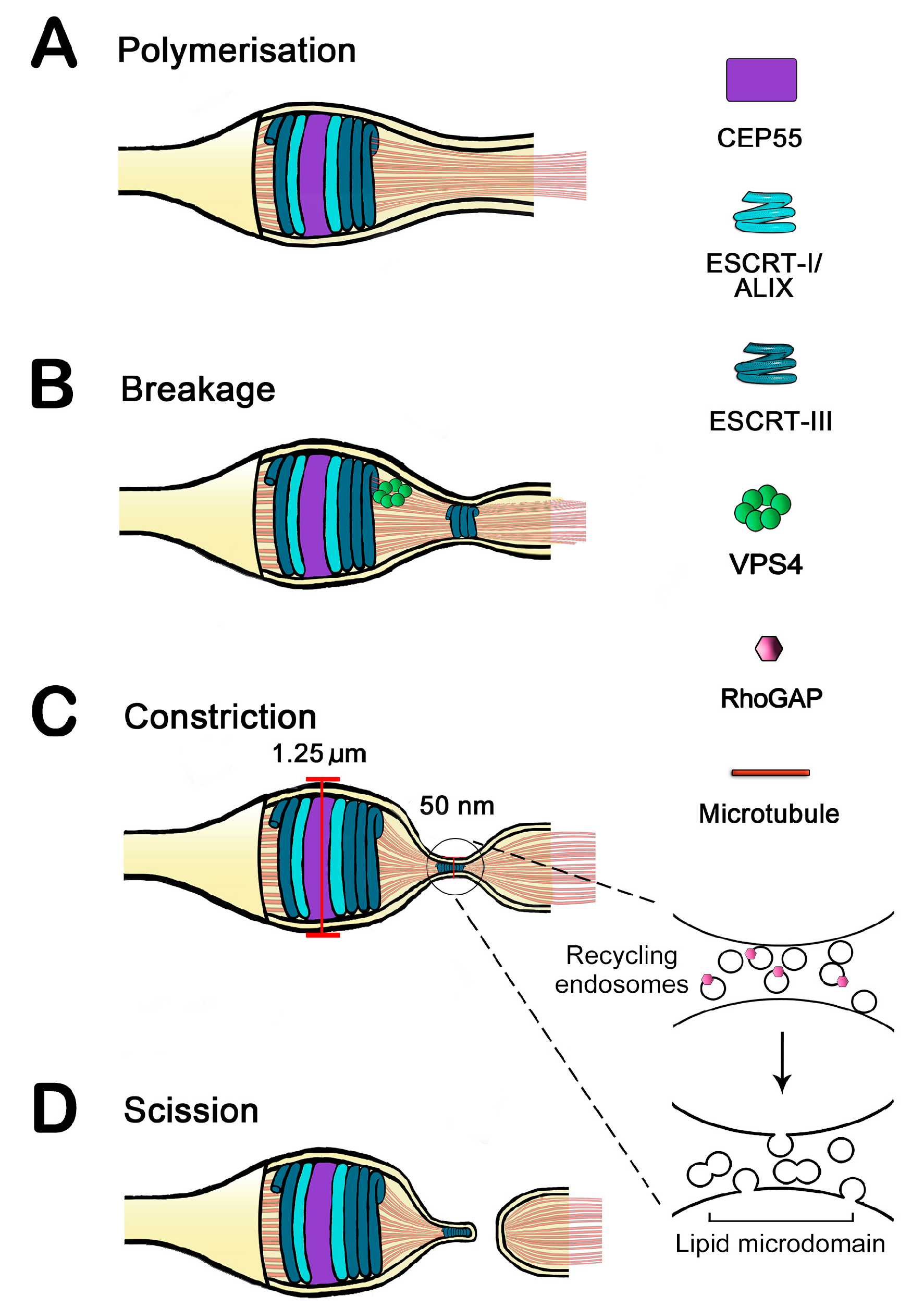
2. Endosomal Sorting Complex Required for Transport (ESCRT) Proteins
3. Function and Dynamics of ESCRTs at Cytokinesis
3.1. Endosomes Drive Secondary Ingression
3.2. ESCRTs Drive Secondary Ingression
3.3. Integrated Models
4. ESCRTs in Other Organisms

{kind=link}
| ESCRT Class | Mammalian Cells | Fission Yeast | Budding Yeast | Worms | Arabidopsis | Archaea | Key Domains | Function |
|---|---|---|---|---|---|---|---|---|
| ESCRT-0 | HRS | Sst4 (Vps27) | Vps27 | Vps27 | Ubiquitin binding domain, phosphatidylinositol 3-phosphate binding domain | Clustering of ubiquitylated cargo | ||
| STAM1/2 | Hse1 | Hse1 | ||||||
| ESCRT-I | TSG101 | Sst6 (Vps23) | Stp22 | TSG-101 | elc | Proline rich linker region that targets ESCRT-I to the midbody during cytokinesis | Membrane budding (with ESCRT-II) and cytokinesis | |
| Vps28 | Vps28 | Vps28 | ||||||
| Vps37A, Vps37B, Vps37C | Vps37 | Vps37 | ||||||
| MVB12A, MVB12B | Mvb12 | MVB-12 | ||||||
| ESCRT-II | EAP30 | Dot2 (Vps22) | Snf8 | Connecting MVBs to microtubules | Membrane budding (with ESCRT-I) | |||
| EAP20 | Vps25 | Vps25 | ||||||
| EAP45 | Vps36 | Vps36 | ||||||
| ESCRT-III | CHMP2A, CHMP2B | Vps2 (Did4) | Did4 | MIM1/2 domain recruits Vps4 to initiate ESCRT-III disassembly. Winged helix domain | Membrane scission/cytokinesis | |||
| CHMP3 | Vps24 | Vps24 | Sacil 373 | |||||
| CHMP4A, CHMP4B, CHMP4C | Vps32 (Snf7) | Snf7 | Vps32 | |||||
| CHMP6 | Vps20 | Vps20 | ||||||
| Vps4 | Vps4A, Vps4B | Vps4 | Vps4 | Skd1 | Sacil 373 | MIT ESCRT-III binding domain, ATPase domain | ESCRT-III disassembly, cytokinesis MVB biogenesis | |
| CHMP5 | SPCC162.06c | Vps60 | ||||||
| Vta1 | Vta1 | Vta1 | ||||||
| ALIX | ALIX | Bro1 | Bro1 | Interacts with apoptosis factors and cytoskeleton. Recruit ESCRT-III to midbody | Targeting functions, cytokinesis/membrane abscission |
4.1. Yeast
4.2. Worms
4.3. Fungi
4.4. Plants
4.5. Archae
5. ESCRT Regulation
5.1. Lipids may Control ESCRT Function
5.2. Polo-Like Kinase and the Control of Early Cytokinesis
5.3. Aurora Kinase and the Abscission Checkpoint
5.4. ANCHRs Away
5.5. Is There More
5.6. Plk and Abscission—Hints from Schizosaccharomyces pombe
6. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Glotzer, M. Animal cell cytokinesis. Ann. Rev. Cell. Dev. Biol. 2001, 17, 351–386. [Google Scholar] [CrossRef]
- Barr, F.A.; Gruneberg, U. Cytokinesis: Placing and making the final cut. Cell 2007, 131, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Hanson, P.I. Membrane budding and scission by the ESCRT machinery: It’s all in the neck. Nat. Rev. Mol. Cell Biol. 2010, 11, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Guizetti, J.; Gerlich, D.W. Cytokinetic abscission in animal cells. Semin. Cell Dev. Biol. 2010, 21, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Elia, N.; Sougrat, R.; Spurlin, T.A.; Hurley, J.H.; Lippincott-Schwartz, J. Dynamics of endosomal sorting complex required for transport (ESCRT) machinery during cytokinesis and its role in abscission. Proc. Natl. Acad. Sci. USA 2011, 108, 4846–4851. [Google Scholar] [CrossRef] [PubMed]
- Schiel, J.A.; Prekeris, R. Membrane dynamics during cytokinesis. Curr. Opin. Cell Biol. 2013, 25, 92–98. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B.; Martin-Serrano, J. No strings attached: The ESCRT machinery in viral budding and cytokinesis. J. Cell. Sci. 2009, 122, 2167–2177. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.B.; Schonteich, E.; Matheson, J.; Wilson, G.; Yu, X.; Hickson, G.R.; Srivastava, S.; Baldwin, S.A.; Prekeris, R.; Gould, G.W. Rab11-FIP3 and FIP4 interact with Arf6 and the exocyst to control membrane traffic in cytokinesis. EMBO J. 2005, 24, 3389–3399. [Google Scholar] [CrossRef] [PubMed]
- Gromley, A.; Yeaman, C.; Rosa, J.; Redick, S.; Chen, C.-T.; Mirabelle, S.; Guha, M.; Sillibourne, J.; Doxsey, S.J. Centriolin anchoring of Exocyst and SNARE complexes at the midbody is required for secretory-vesicle-mediated abscission. Cell 2005, 123, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Estey, M.P.; di Ciano-Oliveira, C.; Froese, C.D.; Bejide, M.T.; Trimble, W.S. Distinct roles of septins in cytokinesis: SEPT9 mediates midbody abscission. J. Cell Biol. 2010, 191, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Low, S.H.; Li, X.; Miura, M.; Kudo, N.; Quinones, B.; Weimbs, T. Syntaxin 2 and endobrevin are required for the terminal step of cytokinesis in mammalian cells. Dev. Cell 2003, 4, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Neto, H.; Collins, L.L.; Gould, G.W. Vesicle trafficking and membrane remodelling in cytokinesis. Biochem. J. 2011, 437, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Elia, N.; Ghirlando, R.; Lippincott-Schwartz, J.; Hurley, J.H. Midbody targeting of the ESCRT machinery by a non-canonical coiled coil in CEP55. Science 2008, 322, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Teis, D.; Saksena, S.; Emr, S.D. SnapShot: The ESCRT machinery. Cell 2009, 137, 182–182e1. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Boura, E.; Carlson, L.A.; Rozycki, B. Membrane budding. Cell 2010, 143, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Emr, S.D. The ESCRT complexes: Structure and mechanism of a membrane-trafficking network. Annu. Rev. Biophys. Biomol. Struct. 2006, 35, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Wollert, T.; Hurley, J.H. Molecular mechanism of multivesicular body biogenesis by ESCRT complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Wollert, T.; Wunder, C.; Lippincott-Schwartz, J.; Hurley, J.H. Membrane scission by the ESCRT-III complex. Nature 2009, 458, 172–178. [Google Scholar] [PubMed]
- Hanson, P.I.; Shim, S.; Merrill, S.A. Cell biology of the ESCRT machinery. Curr. Opin. Cell Biol. 2009, 21, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Lata, S.; Schoehn, G.; Jain, A.; Pires, R.; Piehler, J.; Gottlinger, H.G.; Weissenhorn, W. Helical structures of ESCRT-III are disassembled by VPS4. Science 2008, 321, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Saksena, S.; Wahlman, J.; Teis, D.; Johnson, A.E.; Emr, S.D. Functional reconstitution of ESCRT-III assembly and disassembly. Cell 2009, 136, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Colf, L.A.; Karren, M.A.; Sandrin, V.; Rodesch, C.K.; Sundquist, W.I. Human ESCRT-III and VPS4 proteins are required for centrosome and spindle maintenance. Proc. Natl. Acad. Sci. USA 2010, 107, 12889–12894. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Kimpler, L.A.; Naismith, T.V.; Lauer, J.M.; Hanson, P.I. Interaction of the mammalian endosomal sorting complex required for transport (ESCRT) III protein hSnf7–1 with itself, membranes, and the AAA+ ATPase SKD1. J. Biol. Chem. 2005, 280, 12799–12809. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.G.; Martin-Serrano, J. Parallels between cytokinesis and retroviral budding: A role for the ESCRT machinery. Science 2007, 316, 1908–1912. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Sandrin, V.; Chung, H.-Y.; Morham, S.G.; Gygi, S.P.; Rodesch, C.K.; Sundquist, W.I. Human ESCRT and ALIX proteins interact with proteins of the midbody and function in cytokinesis. EMBO J. 2007, 26, 4215–4227. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chang, J.; Renvoise, B.; Tipirneni, A.; Yang, S.; Blackstone, C. MITD1 is recruited to midbodies by ESCRT-III and participates in cytokinesis. Mol. Biol. Cell 2012, 23, 4347–4361. [Google Scholar] [CrossRef] [PubMed]
- Renvoise, B.; Parker, R.L.; Yang, D.; Bakowska, J.C.; Hurley, J.H.; Blackstone, C. SPG20 protein spartin is recruited to midbodies by ESCRT-III protein Ist1 and participates in cytokinesis. Mol. Biol. Cell 2010, 21, 3293–3303. [Google Scholar] [CrossRef] [PubMed]
- Agromayor, M.; Carlton, J.G.; Phelan, J.P.; Matthews, D.R.; Carlin, L.M.; Ameer-Beg, S.; Bowers, K.; Martin-Serrano, J. Essential role of hIST1 in cytokinesis. Mol. Biol. Cell 2009, 20, 1374–1387. [Google Scholar] [CrossRef] [PubMed]
- Guizetti, J.; Schermelleh, L.; Mantler, J.; Maar, S.; Poser, I.; Leonhardt, H.; Muller-Reichert, T.; Gerlich, D.W. Cortical constriction during abscission involves helices of ESCRT-III-dependent filaments. Science 2011, 331, 1616–1620. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.N.; Barr, F.A. Plk1 negatively regulates Cep55 recruitment to the midbody to ensure orderly abscission. J. Cell Biol. 2010, 191, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Schiel, J.A.; Simon, G.C.; Zaharris, C.; Weisz, J.; Castle, D.; Wu, C.C.; Prekeris, R. FIP3-endosome-dependent formation of the secondary ingression mediates ESCRT-III recruitment during cytokinesis. Nat. Cell Biol. 2012, 14, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Dambournet, D.; Machicoane, M.; Chesneau, L.; Sachse, M.; Rocancourt, M.; el Marjou, A.; Formstecher, E.; Salomon, R.; Goud, B.; Echard, A. Rab35 GTPase and OCRL phosphatase remodel lipids and F-actin for successful cytokinesis. Nat. Cell Biol. 2011, 13, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Im, Y.J.; Wollert, T.; Boura, E.; Hurley, J.H. Structure and function of the ESCRT-II-III interface in multivesicular body biogenesis. Dev. Cell 2009, 17, 234–343. [Google Scholar] [CrossRef] [PubMed]
- Elia, N.; Fabrikant, G.; Kozlov, M.M.; Lippincott-Schwartz, J. Computational model of cytokinetic abscission driven by ESCRT-III polymerization and remodeling. Biophys. J. 2012, 102, 2309–2320. [Google Scholar] [CrossRef] [PubMed]
- Neto, H.; Kaupisch, A.; Collins, L.L.; Gould, G.W. Syntaxin 16 is a master recruitment factor for cytokinesis. Mol. Biol. Cell 2013, 24, 3663–3674. [Google Scholar] [CrossRef] [PubMed]
- Burgess, D.R.; Chang, F. Site selection for the cleavage furrow at cytokinesis. Trends Cell Biol. 2005, 15, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Rismanchi, N.; Renvoise, B.; Lippincott-Schwartz, J.; Blackstone, C.; Hurley, J.H. Structural basis for midbody targeting of spastin by the ESCRT-III protein CHMP1B. Nat. Struct. Mol. Biol. 2008, 15, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.W.; Lindon, C.; Luzio, J.P.; Reid, E. Spastin couples microtubule severing to membrane traffic in completion of cytokinesis and secretion. Traffic 2009, 10, 42–56. [Google Scholar] [CrossRef] [PubMed]
- McMurray, M.A.; Stefan, C.J.; Wemmer, M.; Odorizzi, G.; Emr, S.D.; Thorner, J. Genetic interactions with mutations affecting septin assembly reveal ESCRT functions in budding yeast cytokinesis. Biol. Chem. 2011, 392, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Bhutta, M.S.; Roy, B.; Gould, G.W.; McInerny, C.J. A complex network of interactions between mitotic kinases, phosphatases and ESCRT proteins regulates septation and membrane trafficking in S. pombe. PLoS One 2014, 9, e111789. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, T.; Onishi, M.; Ikeuchi, M.; Kita, A.; Sugiura, R.; Giga-Hama, Y.; Fukui, Y.; Takegawa, K. Essential roles of class E Vps proteins for sorting into multivesicular bodies in Schizosaccharomyces pombe. Microbiology 2007, 153, 2753–2764. [Google Scholar] [CrossRef] [PubMed]
- Green, J.W.; Snoek, L.B.; Kammenga, J.E.; Harvey, S.C. Genetic mapping of variation in dauer larvae development in growing populations of Caenorhabditis elegans. Heredity 2013, 111, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Govindaraghavan, M.; McGuire Anglin, S.L.; Shen, K.F.; Shukla, N.; de Souza, C.P.; Osmani, S.A. Identification of interphase functions for the NIMA kinase involving microtubules and the ESCRT pathway. PLoS Genet. 2014, 10, e1004248. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.G.; Mullen, R.T. Meta-analysis of the expression profiles of the Arabidopsis ESCRT machinery. Plant. Signal. Behav. 2011, 6, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.G.; Howard, A.S.; Khuu, N.; Gidda, S.K.; McCartney, A.; Morphy, B.J.; Mullen, R.T. Protein–protein interaction network and subcellular localization of the Arabidopsis thaliana ESCRT machinery. Front. Plant Sci. 2011, 2, 20. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, C.; Schellmann, S.; Sabovljevic, A.; Shahriari, M.; Keshavaiah, C.; Bechtold, N.; Herzog, M.; Muller, S.; Hanisch, F.G.; Hulskamp, M. The Arabidopsis elch mutant reveals functions of an ESCRT component in cytokinesis. Development 2006, 133, 4679–4689. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, C.; Reyes, F.C.; Buono, R.; Sliwinski, M.K.; Haas, T.J.; Otegui, M.S. The ESCRT-related CHMP1A and B proteins mediate multivesicular body sorting of auxin carriers in Arabidopsis and are required for plant development. Plant Cell 2009, 21, 749–766. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhuang, X.; Gao, C.; Wang, X.; Jiang, L. The Arabidopsis endosomal sorting complex required for transport III regulates internal vesicle formation of the prevacuolar compartment and is required for plant development. Plant Physiol. 2014, 165, 1328–1343. [Google Scholar] [CrossRef] [PubMed]
- Reyes, F.C.; Buono, R.A.; Roschzttardtz, H.; di Rubbo, S.; Yeun, L.H.; Russinova, E.; Otegui, M.S. A novel endosomal sorting complex required for transport (ESCRT) component in Arabidopsis thaliana controls cell expansion and development. J. Biol. Chem. 2014, 289, 4980–4988. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.Y.; Obita, T.; Freund, S.M.; Williams, R.L.; Bell, S.D. A role for the ESCRT system in cell division in Archaea. Science 2008, 322, 1710–1713. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.C.; Samson, R.Y.; Brumfield, S.K.; Bell, S.D.; Young, M.J. Functional interplay between a virus and the ESCRT machinery in archaea. Proc. Natl. Acad. Sci. USA 2013, 110, 10783–70787. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.Y.; Obita, T.; Hodgson, B.; Shaw, M.K.; Chong, P.L.; Williams, R.L.; Bell, S.D. Molecular and structural basis of ESCRT-III recruitment to membranes during archaeal cell division. Mol. Cell 2011, 41, 186–196. [Google Scholar]
- Dobro, M.J.; Samson, R.Y.; Yu, Z.; McCullough, J.; Ding, H.J.; Chong, P.L.; Bell, S.D.; Jensen, G.J. Electron cryotomography of ESCRT assemblies and dividing Sulfolobus cells suggests that spiraling filaments are involved in membrane scission. Mol. Biol. Cell 2013, 24, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Whitley, P.; Reaves, B.J.; Hashimoto, M.; Riley, A.M.; Potter, B.V.L.; Holman, G.D. Identification of mammalian Vps24p as an effector of phosphatidylinositol 3,5-bisphosphate-dependent endosome compartmentalization. J. Biol. Chem. 2003, 278, 38786–38795. [Google Scholar] [CrossRef] [PubMed]
- Yorikawa, C.; Shibata, H.; Waguri, S.; Hatta, K.; Horii, M.; Katoh, K.; Kobayashi, T.; Uchiyama, Y.; Maki, M. Human CHMP6, a myristoylated ESCRT-III protein, interacts directly with an ESCRT-II component EAP20 and regulates endosomal cargo sorting. Biochem. J. 2005, 387, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.M.; Chang, F.; Burgess, D.R. Movement of membrane domains and requirement of membrane signaling molecules for cytokinesis. Curr. Biol. 2005, 9, 781. [Google Scholar]
- Chen, C.T.; Hehnly, H.; Doxsey, S.J. Orchestrating vesicle transport, ESCRTs and kinase surveillance during abscission. Nat. Rev. Mol. Cell Biol. 2012, 13, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.A.; Sillje, H.H.W.; Nigg, E.A. Polo-like kinases and the orchestration of cell division. Nat. Rev. Mol. Cell Biol. 2004, 5, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, J.R.; Plowman, G.D. The Aurora/Ipl1p kinase family: Regulators of chromosome segregation and cytokinesis. Trends Cell Biol. 1999, 9, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.K.; Coughlin, M.; Mitchison, T.J. Midbody assembly and its regulation during cytokinesis. Mol. Biol. Cell 2012, 23, 1024–1034. [Google Scholar] [CrossRef] [PubMed]
- Norden, C.; Mendoza, M.; Dobbelaere, J.; Kotwaliwale, C.V.; Biggins, S.; Barral, Y. The NoCut pathway links completion of cytokinesis to spindle midzone function to prevent chromosome breakage. Cell 2006, 125, 85. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.G.; Caballe, A.; Agromayor, M.; Kloc, M.; Martin-Serrano, J. ESCRT-III governs the Aurora B-mediated abscission checkpoint through CHMP4C. Science 2012, 336, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, L.; Montembault, E.; Takeda, T.; Bassi, Z.I.; Glover, D.M.; D’Avino, P.P. The chromosomal passenger complex controls the function of endosomal sorting complex required for transport-III Snf7 proteins during cytokinesis. Open Biol. 2012, 2, 120070. [Google Scholar] [CrossRef] [PubMed]
- Thoresen, S.B.; Campsteijn, C.; Vietri, M.; Schink, K.O.; Liestol, K.; Andersen, J.S.; Raiborg, C.; Stenmark, H. ANCHR mediates Aurora-B-dependent abscission checkpoint control through retention of VPS4. Nat. Cell Biol. 2014, 16, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.; Schiebel, E. Separase regulates INCENP-Aurora B anaphase spindle function through Cdc14. Science 2003, 302, 2120–2124. [Google Scholar] [CrossRef] [PubMed]
- Lanzetti, L.; Margaria, V.; Melander, F.; Virgili, L.; Lee, M.H.; Bartel, J.; Jensen, S. Regulation of the Rab5 GTPase-activating protein RN-tre by the dual-specificity phosphatase Cdc14A in human cells. J. Biol. Chem. 2007, 282, 15258–15270. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhutta, M.S.; McInerny, C.J.; Gould, G.W. ESCRT Function in Cytokinesis: Location, Dynamics and Regulation by Mitotic Kinases. Int. J. Mol. Sci. 2014, 15, 21723-21739. https://doi.org/10.3390/ijms151221723
Bhutta MS, McInerny CJ, Gould GW. ESCRT Function in Cytokinesis: Location, Dynamics and Regulation by Mitotic Kinases. International Journal of Molecular Sciences. 2014; 15(12):21723-21739. https://doi.org/10.3390/ijms151221723
Chicago/Turabian StyleBhutta, Musab S., Christopher J. McInerny, and Gwyn W. Gould. 2014. "ESCRT Function in Cytokinesis: Location, Dynamics and Regulation by Mitotic Kinases" International Journal of Molecular Sciences 15, no. 12: 21723-21739. https://doi.org/10.3390/ijms151221723
APA StyleBhutta, M. S., McInerny, C. J., & Gould, G. W. (2014). ESCRT Function in Cytokinesis: Location, Dynamics and Regulation by Mitotic Kinases. International Journal of Molecular Sciences, 15(12), 21723-21739. https://doi.org/10.3390/ijms151221723